

Application of nonmaterial's in medicine is an emerging field with potentials to positively impact human health care [1]. Nanoconjugates of monoclonal antibodies including EGFR (epidermal growth factor receptor) antibody cetuximab (C225), herceptin, are potential candidates for various biomedical applications including targeted delivery, detection/diagnosis and imaging [2][3]. However, mechanisms by which C225 and its nanoconjugates transport to the cell and molecular machineries involved for intracellular delivery are not well understood. A better understanding of the regulatory components involved in endocytosis of these nanoconjugates will aide us for successful nanodesign to achieve specific intracellular targeting. Exploiting strong affinity of gold nanoparticles (AuNPs) to bind to –SH and –NH2 moieties present in the cysteine and lysine residues of C225, we design two Au-C225 conjugates having variable number of antibody to demonstrate that the mechanism of endocytosis of C225 in PANC-1 cells can be tailored by precise design of the nanoconjugates [4]. Using pharmacological inhibitors (Supplementary Figure 1), genetic approaches and Cdc42 null cells we demonstrate that both C225 and gold-conjugated C225 with complete surface coverage (Au-C225-C; ∼3 C225/particle) requires clustering in the GSL (Glycosphingolipid) domains at the plasma membrane followed by dynamin-2 (dyn-2) dependent caveolar endocytosis. However, partially covered nanoconjugate (Au-C225-P; ∼1 C225/particle) is internalized via dyn-2 independent Cdc42 dependent pinocytosis/phagocytosis that requires actin polymerization. Regulating the endocytosis of cetuximab between caveolar and pinocytic internalization by appropriate nanodesign may be useful to target specific intracellular pathways for better therapeutic intervention with reduced side effects.
To determine the mechanism and molecular machinery involved in the uptake of C225 and its nanoconjugates we designed two nanoconjugates as we have described recently; a) AuNP surface partially covered by C225, termed as Au-C225-P and b) AuNP surface fully covered by cetuximab, termed as Au-C225-C (Figure 1a). The C225 to AuNP ratio in Au-C225-P and Au-C225-C is ∼ 1 and 3, respectively [5]. To study the interaction between C225 and its nanoconjugates with EGFR by confocal microscopy, we transfected PANC-1 cells with EGFR-GFP (see methods) and treated with C225, Au-C225-P or Au-C225-C for 1h where C225 was Cy3 labeled. Both C225 and Au-C225-C demonstrated similar co-localization with EGFR at the cell membrane indicating that the receptor-binding motif of C225 remained unaffected upon assembly on AuNPs. However, relatively lower extent of co-localization of Au-C225-P with EGFR at the plasma membrane suggests other modes of interaction with the cell membrane (Supplementary Figure 2).
Figure 1. Effect on dynamin-2 requirement upon nano surface coverage and mechanism of C225-nanoconjugate uptake; role of CME vs. CI.

(a) Cartoon presentation of the reaction scheme and structure of the antibody-nanoconjugates (Au-C225-P and Au-C225-C). (b) To study the dependency on dynamin-2, PANC-1 cells infected with dyn-2 WT and dominant negative mutant (K44A) adenovirus, show switching of endocytosis pathway upon increasing AuNP surface coverage (Au-C225-C) with C225 compared to Au-C225-P (showing dyn-2 independent internalization, Scale bar 20 μm). (c) Quantification of antibody uptake by Au-C225-C treatment with Dyn-2 WT and DynK44A infected cell. (d) PANC-1 cells were preincubated with 10 mM βMCD (methyl-β-cyclodextrin) for 30 min in a chamber slides and processed for confocal microscopy. Fluorescence images show the inhibition of internalization of Cy3-labeled C225 and its different nanoconjugates (Au-C225-P and Au-C225-C) for 1 h at 37 °C (Scale bar 20 μm). (e) To study the clathrin independent (CI) endocytosis, PANC-1 cell transfected with GFP-EGFR constructs were preincubated with clostridium difficile toxin B (660 ng/mL) for 1 h and then treated with Cy3 labeled C225, Au-C225-P and Au-C225-C for 1h at 37 °C. Fluorescence images show that uptake was only inhibited for Au-C225-P by toxin B treatment (Scale bar 20 μm). (f) Quantification of antibody uptake as affected by the pharmacological inhibitors treatments.
We earlier demonstrated that C225 internalization in PANC-1 cell is dyn-2 dependent [6]. Here, we wanted to investigate whether the dyn-2 dependent internalization of C225 could be altered by nanodesign. We infected the PANC-1 cells with wild type dyn-2 (Dyn-2-WT) and mutant dyn-2 (Dyn-2-K44A) adenovirus and monitored the internalization of Cy3 labeled C225 and its different nanoconjugates (Au-C225-P and Au-C225-C) by confocal microscopy. Substantial inhibition of Au-C225-C internalization (Figure 1b, c) was observed in PANC-1 cells expressing mutant dyn-2-K44A as compared to dyn2-WT expressing cells, whereas the same cells did not inhibit the uptake of Au-C225-P (Figure 1b). Thus, dyn-2 dependent endocytosis of C225 can be altered by appropriate nanodesign.
Because dyn-2 is involved in both CME (Clathrin Mediated Endocytosis) and CI (Clathrin Independent) pathways, we wanted to delineate further specific endocytic pathways involved during internalization of C225-nanoconjugates[7]. Chlorpromazine, a CME inhibitor, has no apparent effect on the internalization of either C225 or its different nanoconjugates (Au-C225-P and Au-C225-C uptake) (Supplementary Figure 3) [8]. However, as a positive control, it inhibited the internalization of transferrin (Supplementary Figure 4). These data clearly suggest that C225 and its different nanoconjugates do not require the CME pathway as their primary route of internalization.
Since C225 and its nanoconjugates do not internalize by CME, we then investigated CI pathways. To determine the involvement of lipid raft we used β-methyl cyclodextrin (BmCD), a reagent that extracts cholesterol from the plasma membrane (PM), thereby inhibiting lipid raft dependent endocytosis [8-9]. Preincubation of PANC-1 cells with 5 mM BmCD for 30 minutes selectively blocked (∼70 %) internalization of C225 (Supplementary Figure 5) while higher concentration of BmCD (10 mM) was required to inhibit Au-C225-P and Au-C225-C internalization to a similar extent (Figure 1d). Taken together, these data suggest the possible involvement of lipid raft mediated caveolar endocytosis for both Au-C225-P and Au-C225-C. Preincubation of PANC-1 cell with another Cav-ME inhibitor nystatin (50 μg/ml) also inhibited the internalization of both the nanoconjugates (Supplementary Figure 5), thus reconfirming the involvement of lipid-rafts in caveolar endocytosis of C225 and its nanoconjugates [8].
To investigate a possible involvement of other CI pathways apart from Cav-ME, we used cytochalasin D, a fungal metabolite, primarily known to block macropinocytic/phagocytic pathways by inhibiting actin polymerization[10]. Pretreatment of PANC-1 cells with cytochalasin D selectively inhibited internalization of Au-C225-P, whereas it had no effect on C225 and Au-C225-C uptake. (Supplementary Figure 5). Similarly, treatment of PANC-1 cells with Clostridium difficile Toxin B specifically inhibited the internalization of Au-C225-P, whereas it too had no effect for C225 and Au-C225-C internalization (Figure 1e, f). Toxin B inhibits the RhoA and Cdc42 dependent CI pathways[8, 11]. These results suggest that, possibly RhoA/Cdc42 dependent pinocytosis and actin polymerization are required for Au-C225-P uptake.
To further confirm the involvement of RhoA/Cdc42 during Au-C225-P internalization, we overexpressed dominant negative RhoA (RhoAT19N) and Cdc42 (Cdc42T17N) mutants in PANC-1 cells and studied internalization of C225, Au-C225-C and Au-C225-P. We used Alexa fluor 594 labeled dextran particles as a positive control, which requires Cdc42 for internalization (Supplementary Figure 6) [12]. The uptake of Au-C225-P was completely inhibited by overexpression of Cdc42T17N whereas it remained unaffected for C225 and Au-C225-C (Figure 2a, b). Internalization of C225-nanocinjugates were, however found to be RhoA independent in RhoA dominant negative mutant expressing PANC-1 cells (Supplementary Figure 7). The results with cytochalasin D, toxin B and Cdc42 dominant negative construct independently demonstrate that endocytosis of Au-C225-P involves actin polymerization and entry into the cell is primarily by Cdc42 dependent macropinocytosis pathways. To visualize the endocytosis of C225, Au-C225-P and Au-C225-C at the molecular level, we performed transmission electron microscopy (TEM) analysis of PANC-1 cells after treatment with C225-nanoconjugates. Figure 2c shows an induction of a caveolar structure at the plasma membrane after incubation with C225. Similarly, AuNPs were also found inside a flask and grape shaped caveolar architecture in Au-C225-C treated PANC-1 cells (Figure 2c), further confirming the involvement of Cav-ME for C225 and Au-C225-C. On the other hand, AuNPs were found in close vicinity of pinocytosis invaginations at the PM in Au-C225-P treated PANC-1 cells (Supplementary Figure 8). TEM image of Au-C225-P uptake at the pinocytosis morphology corroborates the results obtained from Cdc42 dominant mutant and Toxin B experiments.
Figure 2. Role of Cdc42 GTPase on the internalization of C225, Au-C225-P and Au-C225-C and structural Elucidation of CI Pathways.

(a) PANC-1 cell transfected with GFP-Cdc42F17 dominant negative mutants were treated with Cy3 labeled C225, Au-C225-P and Au-C225-C for 1 h at 37 °C. Fluorescence images shows the inhibition of uptake of AuNPs partially covered with C225 (Au-C225-P) while the uptake of C225, Au-C225-C remains unaffected. Lower panel represent the images for Dex-Red particles (positive control) treated transfected cell (Scale bar 10 μm). (b) Quantification of nanoconjugates internalization in the presence of dominant negative mutant, GFP-Cdc42F17 transfected PANC-1 cell. (c) TEM images represent the arrest of the caveolar invagination, triggered by the treatment with C225 and Au-C225-C to PANC-1 cell at 4 °C, incubated for 2 h (Scale bar 100 nm).
Since C225 and its nanoconjugates required lipid raft for endocytosis, next we investigated the involvement of specific lipid microdomains at the plasma membrane during Cav-ME and other CI pathways. We treated PANC-1 cells with ceramide synthase inhibitor fumonosin B1 (FB1) to inhibit SL (sphingolipid) biosynthesis [12-13]. Treatment with FB1 led to the inhibition of internalization of C225 and all of its nanoconjugates (Figure 3a). While uptake of C225 and Au-C225-C could be restored by exogenous addition of ganglioside GM3, internalization of Au-C225-P could not (Figure 3a, b) [13-14]. These results suggest that internalization of C225 and Au-C225-C involve Cav-ME that requires glycosphingolipid domain to build the endocytotic vesicles whereas Au-C225-P primarily requires fluid phase mediated uptake (Figure 3c).
Figure 3. Lipid micro domain involved during cetuximab-nanoconjugates uptake.
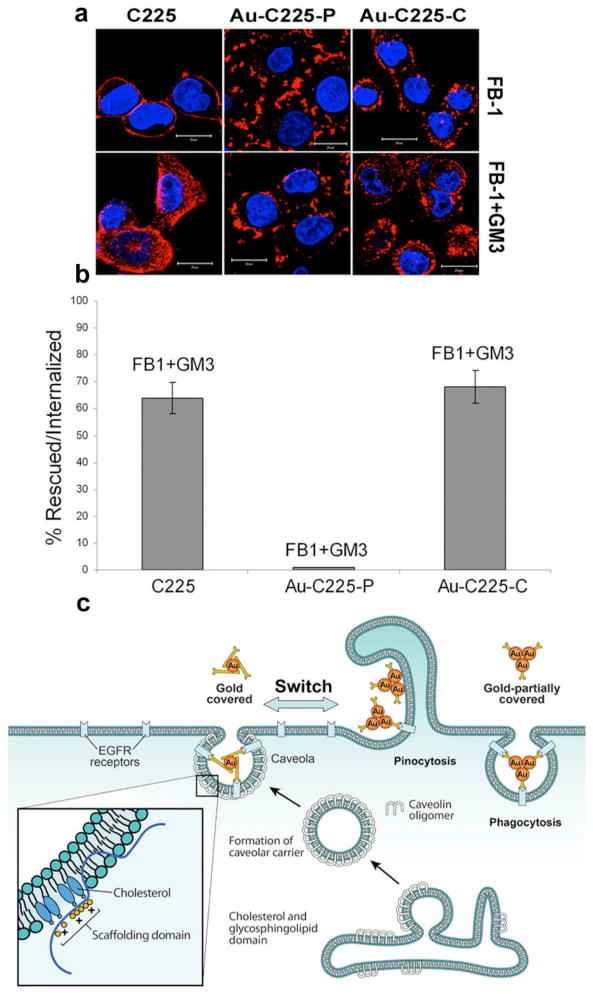
(a) PANC-1 cell was preincubated Fumonisin Synthase B1 (FB1) for 48 h followed by treatment with Cy3 labeled C225, Au-C225-P and Au-C225-C for 1h at 37 °C. Fluorescence images show the inhibition of internalization (> 90 %, upper panel) of C225and its gold conjugate (Au-C225-P and Au-C225-C). Uptake of the C225 and Au-C225-C was restored upon addition of exogenous monosialoganglioside (GM3) (200 μg/ml) for 1 h at 37 °C. Lower panel images show the restoration of uptake of C225 and Au-C225-C when incubated with GM3, whereas inhibition of Au-C225-P cannot be restored by GM3 (Scale bar 20 μm). (b) Quantification of uptake of C225 and its nanoconjugates rescued by the addition of GM3 to FB-1 treated PANC-1 cell. (c) Proposed mechanism of pathway switch as triggered by the nanoconjugated antibody (Au-C225-P and Au-C225-C).
Understanding the nanoconjugate interaction with the cell is critical for a successful clinical translation of a nanomaterial-based targeted drug delivery system with minimum side effects[3b]. Cetuximab utilizes mainly Cav-ME pathway for its endocytosis that involves cholesterol and GSLs lipid raft microdomain at PM.[15] When AuNP was partially covered by C225; it allows the nanoconjugate to interact with the proteins at the plasma membrane through available gold surface. Such interactions help internalization of C225 through an alternative pathway that requires Cdc42 and actin polymerization. This hypothesis was further validated by quantifying the uptake of naked AuNPs in Cdc42 null cells.[16] Uptake of AuNPs was substantially inhibited in Cdc42 null cells (Supplementary Figure 6) confirming the importance of availability of gold surface during uptake via Cdc42 dependent pinocytic/phagocytosis pathways.
In conclusion, we have demonstrated that the mechanism of caveolar uptake of cetuximab could be shifted to dynmin-2 independent Cdc42 dependent pinocytotic/phagocytosis pathway with proper nanodesign and antibody loading. Such pinocytotic uptake of Au-C225-P could be reversed to original dynamin-2 dependent caveolar pathway by fully coating the nanosurface with C225 (Figure 3c). Tailoring the endocytosis mechanism of antibody might be useful to modulate intracellular signaling pathway and be effective even if the cells have a defect in endocytosis and transport mechanisms [17].
Supplementary Material
Acknowledgments
Work is supported by National Institutes of Health (NIH) Grant CA135011, CA136494 to P.M. We are thankful to John Charlesworth and Cindy B. Uhl for helping with TEM image analysis. We gratefully acknowledge James E. Tarara for helping with confocal image analysis. We also acknowledge Prof. Cord Brakebusch and M. McNiven for providing us Cdc42 null cell and EGFR-GFP construct, respectively.
References
- 1.Giljohann DA, Mirkin CA. Nature. 2009;462:461–464. doi: 10.1038/nature08605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.a) Patra CR, Bhattacharya R, Wang E, Katarya A, Lau JS, Dutta S, Muders M, Wang S, Buhrow SA, Safgren SL, Yaszemski MJ, Reid JM, Ames MM, Mukherjee P, Mukhopadhyay D. Cancer Res. 2008;68:1970–1978. doi: 10.1158/0008-5472.CAN-07-6102. [DOI] [PubMed] [Google Scholar]; b) Jiang W, KimBetty YS, Rutka JT, Chan WCW. Na,t Nano. 2008;3:145–150. doi: 10.1038/nnano.2008.30. [DOI] [PubMed] [Google Scholar]
- 3.a) Dobrovolskaia MA, McNeil SE. Nat Nano. 2007;2:469–478. doi: 10.1038/nnano.2007.223. [DOI] [PubMed] [Google Scholar]; b) Dobrovolskaia MA, Germolec DR, Weaver JL. Nat Nano. 2009;4:411–414. doi: 10.1038/nnano.2009.175. [DOI] [PubMed] [Google Scholar]
- 4.Masui H, Kawamoto T, Sato JD, Wolf B, Sato G, Mendelsohn J. Cancer Res. 1984;44:1002–1007. [PubMed] [Google Scholar]
- 5.Khan JA, Kudgus RA, Szabolcs A, Dutta S, Wang E, Cao S, Curran GL, Shah V, Curley S, Mukhopadhyay D, Robertson JD, Bhattacharya R, Mukherjee P. PLoS ONE. 2011;6(6):e20347. doi: 10.1371/journal.pone.0020347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bhattacharyya S, Bhattacharya R, Curley S, McNiven MA, Mukherjee P. Proc Natl Acad Sci USA. 2010;107:14541–14546. doi: 10.1073/pnas.1006507107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mayor S, Pagano RE. Nat Rev Mol Cell Biol. 2007;8:603–612. doi: 10.1038/nrm2216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Singh RD, Marks DL, Pagano RE. Current Protoc Cell Biol / editorial board, Juan S Bonifacino … [et al] 2007;Chapter 24(Unit 24):21. doi: 10.1002/0471143030.cb2401s35. [DOI] [PubMed] [Google Scholar]
- 9.Parton RG, Simons K. Nat Rev Mol Cell Biol. 2007;8:185–194. doi: 10.1038/nrm2122. [DOI] [PubMed] [Google Scholar]
- 10.a Casella JF, Flanagan MD, Lin S. Nature. 1981;293:302–305. doi: 10.1038/293302a0. [DOI] [PubMed] [Google Scholar]; b Chimini G, Chavrier P. Nat Cell Biol. 2000;2:E191–196. doi: 10.1038/35036454. [DOI] [PubMed] [Google Scholar]
- 11.Aktories K. J Clin Invest. 1997;99:827–829. doi: 10.1172/JCI119245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cheng ZJ, Singh RD, Sharma DK, Holicky EL, Hanada K, Marks DL, Pagano RE. Mol Biol Cell. 2006;17:3197–3210. doi: 10.1091/mbc.E05-12-1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Merrill AH, Jr, Liotta DC, Riley RT. Trends Cell Biol. 1996;6:218–223. doi: 10.1016/0962-8924(96)10021-0. [DOI] [PubMed] [Google Scholar]
- 14.Singh RD, Marks DL, Holicky EL, Wheatley CL, Kaptzan T, Sato SB, Kobayashi T, Ling K, Pagano RE. Traffic. 2010;11:348–360. doi: 10.1111/j.1600-0854.2009.01022.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lin J, Zhang H, Chen Z, Zheng Y. ACS Nano. 2010;4:5421–5429. doi: 10.1021/nn1010792. [DOI] [PubMed] [Google Scholar]
- 16.Czuchra A, Wu X, Meyer H, van Hengel J, Schroeder T, Geffers R, Rottner K, Brakebusch C. Mol Biol Cell. 2005;16:4473–4484. doi: 10.1091/mbc.E05-01-0061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wang Y, Pennock S, Chen X, Wang Z. Mol Cell Biol. 2002;22:7279–7290. doi: 10.1128/MCB.22.20.7279-7290.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.