

Abstract
The potential of the developing zebrafish model for toxicology and drug discovery is limited by inefficient approaches to manipulating and chemically exposing zebrafish embryos—namely, manual placement of embryos into 96- or 384-well plates and exposure of embryos while still in the chorion, a barrier of poorly characterized permeability enclosing the developing embryo. We report the automated dechorionation of 1600 embryos at once at 4 h postfertilization (hpf) and placement of the dechorionated embryos into 96-well plates for exposure by 6 hpf. The process removed ≥95% of the embryos from their chorions with 2% embryo mortality by 24 hpf, and 2% of the embryos malformed at 120 hpf. The robotic embryo placement allocated 6-hpf embryos to 94.7% ± 4.2% of the wells in multiple 96-well trials. The rate of embryo mortality was 2.8% (43 of 1536) from robotic handling, the rate of missed wells was 1.2% (18 of 1536), and the frequency of multipicks was <0.1%. Embryo malformations observed at 24 hpf occurred nearly twice as frequently from robotic handling (16 of 864; 1.9%) as from manual pipetting (9 of 864; 1%). There was no statistical difference between the success of performing the embryo placement robotically or manually.
Keywords: zebrafish, automation, chorion, robotic
Introduction
Toxicology is undergoing a paradigm shift recognized by the U.S. Environmental Protection Agency, the National Toxicology Program, and the National Research Council. This is nowhere more evident than the Tox21 agenda1,2 and its European counterpart, the REACH initiative,3 which will fundamentally rely on high-throughput screening methods made possible by enormous advances in robotics, digital imaging, and computational and informational tools to assess a staggering backlog of more than 60 000 chemicals now in production, for most of which little if any toxicology data exist. Although the Tox21 vision emphasizes complete transition to in vitro screening for toxicity prediction, for the foreseeable future, rapid in vitro methods will not eliminate animal toxicity testing but serve to prioritize chemicals for further screening in animals. However, a bottleneck of time and money stands in the way of evaluating even a small fraction of the in vitro prioritized chemicals with conventional rodent models of human risk. A solution to the bottleneck is much faster, predictive animal models that are amenable to automation platforms. Zebrafish has emerged as the choice for such a model. No other vertebrate is better suited to high-throughput chemical screening.4
Zebrafish development is the most sensitive life stage to chemical exposure. The bulk of the model’s utility, and hence the bulk of zebrafish toxicology work, is centered on development. Because zebrafish embryos remain transparent throughout much of organogenesis, adverse effects of chemical exposure on development of the brain, notochord, heart, jaw, body segmentation, and body size can be continuously assessed in the living animal under low magnification. An important developmental feature is that zebrafish embryos that are malformed, missing organs, or displaying organ dysfunction usually survive well beyond the point at which those organs normally start to function. This feature is in stark contrast to rodents, in which heart and other organ malformations, as well as missing or dysfunctional organs, typically cause a generalized in utero lethality. In a large-scale (>1000 animal) rodent screen, such end points would be missed with anything less than Herculean efforts at detection.4
As the only vertebrate model meeting the rapid, predictive toxicology needs of the 21st century, zebrafish are increasingly used by public- and private-sector interests to conduct discovery screens of chemical libraries containing ≥1000 compounds (reviewed in Sukardi et al.5 and Zon and Peterson6; see also Kokel et al.,7 Paik et al.,8 Rihel et al.,9 Wang et al.,10 and Yen et al.11). These same studies also highlight near-exclusive reliance on inefficient approaches to manipulating and chemically exposing zebrafish embryos— namely, manual placement of embryos into 96- or 384-well plates and exposure of embryos while still in the chorion, an acellular barrier of poorly characterized permeability enclosing the developing embryo.
Manual placement of embryos to microtiter wells is not a cost-effective use of laboratory personnel and, although barely feasible for screens of a few thousand compounds, is completely impractical for the scale needed to address the rapidly growing backlog of conventional and nanomaterial chemistries already in use. When working with dechorionated embryos, manual placement by humans requires a considerably more refined handling technique than embryos in the chorion. High-precision, repetitive motions necessitate many breaks for lab technicians, and the mundane nature of the task equates to frequent personnel turnover and inefficiencies associated with continual retraining.
No chemically comprehensive assessment of chorion permeability has been reported, but the chorion is widely suspected to influence chemical uptake, and several reports confirm that it is an uptake barrier for metal nanomaterials.12,13 Reported attempts at zebrafish chorion removal prior to 24 h postfertilization (hpf) on a large (>100 embryo) scale have been plagued by generally low survival,14 and exposure in the chorion continues to be a common practice in large screens. Such screens have yielded much information and potentially invaluable therapeutic discovery (reviewed in Zon and Peterson6), but it is tempting to speculate how much information has been missed because of permeability limitations of the chorion. A second, important limitation of not removing the chorion prior to exposure is that compounds that specifically inhibit the hatching process lead to secondary phenotypic responses. For example, the widely used insecticide cartap inhibits hatching, resulting in secondary effects on the embryo such as wavy notochord, axis malformation, and somite defects.15 These effects are due to the lack of hatching, rather than a primary response to exposure to the chemical. The chorion can significantly confound the early life stage zebrafish toxicity assay by leading to false positives. When we consider the obvious potential for false negatives due to the aforementioned barrier effect, chorion removal is critical to improve the predictivity of the assay.
Herein we report rapid and cost-effective automated removal of the chorion from 2000 embryos at once at the 4-hpf stage and placement of the dechorionated embryos into 96-well plates for exposure at 6 hpf. Two approaches were key to developing these platforms: (1) the use of pronase degradation of the chorion combined with automated agitation and washing of the embryos and (2) the application of machine vision-guided robotics to rapidly select and place the dechorionated embryos into plate wells. A single station is used in our laboratory to plate >1000 dechorionated embryos per day, requiring approximately 4 h and with a survival rate better than 95% by 120 hpf.
Materials and Methods
Zebrafish
Embryonic zebrafish were obtained from a Tropical 5D strain of zebrafish (Danio rerio) reared in the Sinnhuber Aquatic Research Laboratory (SARL) at Oregon State University, Corvallis. Adults were kept at standard laboratory conditions of 28 °C on a 14-h light/10-h dark photoperiod in fish water (FW) consisting of reverse-osmosis water supplemented with a commercially available salt (Instant Ocean, www.instantocean.com) to create a salinity of 600 microsiemens. Sodium bicarbonate was added as needed to adjust the pH to 7.4. Zebrafish were group spawned, and embryos were collected and staged as described by Kimmel et al.16
Automated Chorion Removal
To establish an inexpensive and highly reproducible method of removing chorions from about 1600 embryos at a time at 4 hpf, a Belly Dancer shaker (ATR, Inc., Laurel, MD) was modified to accommodate a custom-machined, anodized aluminum shaker plate that holds 4 glass Petri dish bottoms (100 × 15 mm; VWR, Radnor, PA) and attached water delivery tubing, stainless steel nozzles, and a drain port (Fig. 1). The internal workings of the Belly Dancer were modified with a small pump and a parametric motion controller (Revolution Robotics, Inc., Corvallis, OR). The front panel was modified with an LED display and push button control. The on-board pump supplied rinse water from an external heated (28 °C) carboy via the tubing and nozzles to each glass dish at the appropriate time. The movement of the shaker was controlled by the same system to deliver pulsed agitation or gentle swirling, precisely when needed, to dislodge partially hydrolyzed chorions. The only manually performed steps were the addition of a pronase aliquot to commence digestion and pressing of the start button. No other steps were necessary to operate the device. The pronase digestion of the chorion was performed at 4 hpf. Approximately 2000 zebrafish Tropical 5D strain embryos were received from the Sinnhuber Aquatic Research Laboratory’s mass spawning facility in a 135-mm plastic dish and quickly cleaned by removing all dead, unfertilized, or obviously abnormal embryos with an aspirator, a 5- to 10-min process for a trained technician. Approximately 400 to 500 embryos were placed in each of the 4 glass dishes in 25 mL of FW with 50 μL of 50 mg/mL pronase (#81748; Fluka, St. Louis, MO) for 6.5 min while the dechorionator platform constantly agitated. The pronase was then flushed away by gently overflowing the dish with the pumped-in FW for 10 min with 45-s agitation cycles separated by 15 s while still. The total volume of fish water consumed was about 1 L. After the pronase and rinse phases, the embryos were incubated for 20 min at 28 °C, agitated once more to dislodge any remaining chorions, and rinsed again to remove the dislodged chorions. The dechorionation was evaluated (Table 1) by gently removing approximately 100 embryos with a flame-polished Pasteur pipette after the final rinse and examination under a dissecting microscope for pronase or mechanical damage. No further cleanup of the dechorionated batches was performed prior to allocation to 96-well plates.
Figure 1.

A modified shaker platform-based instrument for the automation of chorion removal from zebrafish embryos at 4 h postfertilization. (A) A frontal view of the modified Belly Dancer shaker. The custom machined and anodized aluminum plate is at top and holds 4 × 100-mm glass dishes. The control panel consists solely of a start-and-stop button and a small LCD status display. (B) A closer view of the shaker platform during a rinse phase of chorion removal. Embryos are visible in the plates. The rinse water is pumped via the onboard pump from an offboard heated source and delivered via the hose and nozzle assembly to gently overflow the plates and not suspend the embryos. The rinse water is channeled to a drain port at the rear of the platform. Agitation and pump control are via the custom onboard microcomputer.
Table 1.
Results of Automated Chorion Removal at 4 h Postfertilization (hpf), observed at 24 and 120 hpf
| Results |
||||||
|---|---|---|---|---|---|---|
| Trial Date | Embryos Sampled from ≈1600a | Normal | Dead at 24 hpf | Malformed at 24 hpf | Dead at 120 hpfb | Malformed at 120 hpfc |
| 7/18/2011 | 86 | 83 | 3 | 0 | 0 | 1 |
| 7/25/2011 | 100 | 97 | 3 | 0 | 0 | 1 |
| 7/26/2011 | 100 | 98 | 1 | 1 | 1 | 2 |
| 7/27/2011 | 100 | 92 | 1 | 7 | 6 | 2 |
| 7/28/2011 | 97 | 95 | 2 | 0 | 1 | 2 |
| 7/29/2011 | 99 | 95 | 0 | 4 | 3 | 1 |
| Total | 582 | 560 | 10 | 12 | 11 | 9 |
| % | 100.0 | 96.2 | 1.7 | 2.1 | 1.9 | 1.5 |
The dechorionator holds 4 × 100-mm dishes with approximately 400 embryos each, from which roughly 100 were removed at once by Pasteur pipette after the process for evaluation.
Additional larval mortality at 120 hpf. Excludes mortality observed at 24 hpf.
Additional larval malformation at 120 hpf. Excludes malformation observed at 24 hpf.
Automated Allocation of Dechorionated Embryos to 96-Well Plates
After the rest period, dechorionated embryos at approximately 5 to 6 hpf were transferred to individual wells of a 96-well BD Falcon (BD Biosciences, Franklin Lakes, NJ) tissue culture polystyrene plate by a custom robotic pick and place system (Fig. 2; video of the robotic system in operation can be viewed at http://tanguaylab.com/Automation.html). We noted that the use of nontissue culture-treated polystyrene plates caused rapid disintegration of 100% of embryos once removed from the chorion. The system consisted of a four-axis Selective Compliant Assembly Robot Arm (SCARA; Denso, Inc., Long Beach, CA) with a custom end-effector designed to replicate a handheld, wide-bore, flame-polished Pasteur pipette (Fig. 2). A 100-mm glass Petri dish with approximately 400 embryos was loaded into a well-lit area below a rigidly mounted machine vision camera (Allied Vision, Inc., Stadtroda, Germany). Custom software was used to determine the precise coordinates of a suitable embryo, which were then passed to the robot control unit. Under the lighting conditions used, normal embryos appeared semitransparent, whereas dead embryos appeared bright white, a parameter easily distinguished by the machine vision. The robot was programmed to first draw 100 μL of embryo medium into the flame-polished pipette from a filling station at the beginning of each cycle, drive the pipette to coordinates several centimeters above the machine vision-selected embryo, and place the pipette tip 20 μm above the embryo coordinates. The embryo was gently aspirated along with an additional ~20 μL of embryo medium into the pipette via the onboard syringe pump, and the robot returned the pipette tip to the coordinates of the liquid surface of the next empty well of the plate. The wells had been prefilled with 50 μL of embryo medium. A quick, gentle touch of the liquid surface was all that was required to cause the embryo to be released to the well by capillary action. Positive dispensing pressure was not needed as the embryo had settled to the bottom of the liquid column while the robot pivoted between the source plate and 96-well plate. We noted that an additional 2 to 3 μL of embryo medium was transferred to the well by capillary release of the embryo. The cycle was completed with an aspirate and total dispense step at a wash station and a subsequent 100-μL recharge at the filling station. The cycle was then repeated 95 times. Because of the affinity of dechorionated 6-hpf embryos for each other, the source Petri dish had to be given a brief, 1-s swirl once during the loading of a plate to redisperse the embryos, improving machine vision selection.
Figure 2.

A custom robotic station for the automated transfer of dechorionated zebrafish embryos from the dish in which chorion removal occurred to a 96-well plate. (A, B) A front and side view of the workstation. An extruded aluminum strut assembly rigidly supports the overhead camera. The station is supported from below by a steel tooling plate welded to a rigid steel table. The four-axis Denso SCARA robot is bolted to the tooling plate as it generates strong inertial forces during its movement. The syringe pump supplying the pipetting force is visible next to the base of the robot. Also visible are the lighted arena beneath the camera and the holder for the destination plate and a second plate for pipette rinsing and preloading with water. (C) The custom end effecter gently cradles a single, wide-bore, flame-polished Pasteur pipette connected to the syringe pump line. (D) The lighted arena consists of an aluminum square with precisely located pins in the corners that serve as visible references for the machine vision software. The circular array of LED units provides the optimal amount of contrast needed for the machine vision to clearly see the embryos. The units are commercially available, high-intensity LED assemblies. When operating, the station is protected by a light curtain that, if interrupted, stops the robot’s motion in less than 50 ms.
Statistical Analysis
Performance of the robotic embryo placement was evaluated over 16 trials in parallel with manual embryo loading of 96-well plates. The frequencies of successful well allocations between each method were compared by one-way analysis of variance (ANOVA) where p < 0.05 was the threshold for no significant difference between the two methods.
Results
Automated Chorion Removal at 4 hpf
The performance of six trials of enzymatic chorion removal from approximately 1600 zebrafish embryos at once was assessed from random samples of about 100 embryos from each trial and summarized in Table 1. In each trial, at least 95% of the 4-hpf embryos were successfully removed from the chorion where success was defined as alive at 24 hpf with no malformations evident by 120 hpf. The automated chorion removal resulted in only about 2% embryo mortality by 24 hpf, and only 2% of the embryos were malformed at 120 hpf. Figure 3 depicts the type of mortal damage that 1%-2% of the embryos were observed to have immediately following the automated dechorionation process. Early damage that may have led to 2% of the embryos being malformed at 120 hpf was not visibly detected.
Figure 3.
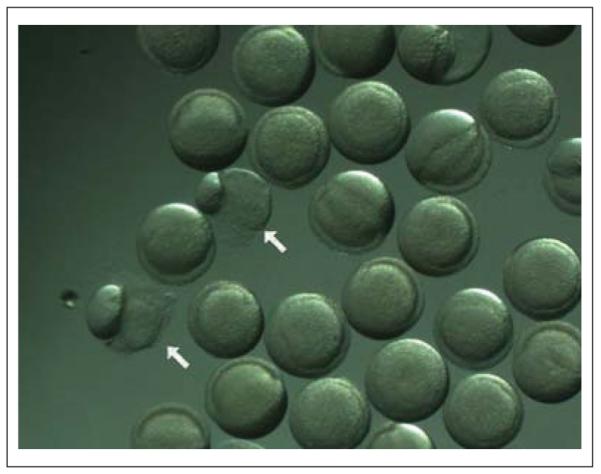
Mortal damage typically observed in 1%-2% of 5-h postfertilization (hpf) zebrafish embryos immediately after the automated dechorionation process. The damaged embryos are indicated by arrows; all other embryos in the field are normal. Whether the low-frequency damage is due to the effects of pronase digestion or motion is unknown.
Automated Placement of Dechorionated Embryos into 96-Well Plates
We evaluated the success of robotic 96-well loading of embryos dechorionated at 4 hpf that were 5-6 hpf at the time of plate loading. The standard for comparison was our routine method of manual placement of dechorionated embryos using a handheld Pasteur pipette. Sixteen 96-well plate comparisons were performed in parallel, and the results are summarized in Table 2. The manual loading data were derived from several of the personnel in our laboratory who are equally adept at the technique. The robotic system successfully allocated embryos to 94.7% ± 4.2% of the wells, and manual loading successfully allocated embryos to 94.9% ± 3.6% of the wells. There was no statistical difference between the success of the two methods (ANOVA F = 0.53; p < 0.01 that a significant difference existed). The criterion for success was that each well received only 1 embryo and that the embryo was alive and not visibly damaged or malformed at 24 hpf. We note that 2.8% (43 of 1536) of the unsuccessful wells were from mortality directly as a result of robotic handling. Mortality from manual loading accounted for 4% (62 of 1536) of the unsuccessful wells. The robotic system missed 1.2% (18 of 1536) of the wells, but only one miss occurred from manual loading. The frequency of multipicks (2 embryos allocated to a single well) was similar for the robotic and manual loading (3 and 2 embryos, respectively, out of 1536). Embryo malformations observed 18 h after plate loading (24 hpf) occurred nearly twice as frequently from robotic handling (16 of 864; 1.9%) as they did from manual pipetting (9 of 864; 1%). No bias toward dead or malformed embryos occurring in certain wells was ever observed.
Table 2.
Robotic Pick-and-Place (P&P) Performance on 6–h Postfertilization (hpf) Dechorionated Zebrafish Embryos Compared to Manual Plate Loading
| Overall P&P Performance |
Specific P&P Failures |
||||||
|---|---|---|---|---|---|---|---|
| Trial Date | Wells | Wells Allocated Successfully, No. (%)a |
Wells Unsuccessful, No. (%)b |
P&P Mortalityc |
Missed Wellsd |
Multipickse | Malformedf |
| 8/1/2011 | 96 | 86 (89.6) | 10 (10.4) | 2 | 5 | 0 | 3 |
| 8/2/2011 | 96 | 90 (93.8) | 6 (6.3) | 4 | 0 | 0 | 2 |
| 8/2/2011 | 96 | 93 (96.9) | 3 (3.1) | 2 | 0 | 0 | 1 |
| 8/2/2011 | 96 | 95 (99.0) | 1 (1.0) | 0 | 0 | 0 | 1 |
| 8/4/2011 | 96 | 85 (88.5) | 11 (11.5) | 9 | 0 | 0 | 2 |
| 8/4/2011 | 96 | 91 (94.8) | 5 (5.2) | 2 | 1 | 0 | 2 |
| 8/5/2011 | 96 | 88 (91.7) | 8 (8.3) | 6 | 0 | 0 | 2 |
| 8/8/2011 | 96 | 87 (90.6) | 9 (9.4) | 6 | 0 | 1 | 2 |
| 8/9/2011 | 96 | 89 (92.7) | 7 (7.3) | 4 | 2 | 0 | 1 |
| 9/6/2011 | 96 | 95 (99.0) | 1 (1.0) | 0 | 1 | 0 | — |
| 9/6/2011 | 96 | 91 (94.8) | 5 (5.2) | 0 | 5 | 1 | — |
| 9/6/2011 | 96 | 96 (100.0) | 0 | 0 | 0 | 0 | — |
| 9/7/2011 | 96 | 84 (87.5) | 12 (12.5) | 8 | 1 | 3 | — |
| 9/7/2011 | 96 | 95 (99.0) | 1 (1.0) | 0 | 1 | 0 | — |
| 9/7/2011 | 96 | 95 (99.0) | 1 (1.0) | 0 | 1 | 0 | — |
| 9/7/2011 | 96 | 95 (99.0) | 1 (1.0) | 0 | 1 | 0 | — |
| Ave = 94.7% ± 4.2%g | 2.8% | 1.2% | 0.3% | 1.9% | |||
| n = 16 trials | |||||||
|
| |||||||
| Overall Manual Performance |
Specific Manual Failures |
||||||
| Trial Date | Wells | Wells Allocated Successfully, No. (%) |
Wells Unsuccessful, No. (%) |
Manual Load Mortality |
Missed Wells |
Multipicks | Malformed |
|
| |||||||
| 8/1/2011h | 96 | 87 (90.6) | 9 (9.4) | 6 | 1 | 1 | 1 |
| 8/2/2011h | 96 | 89 (92.7) | 7 (7.3) | 7 | 0 | 0 | 0 |
| 8/2/2011h | 96 | 87 (90.6) | 9 (9.4) | 8 | 0 | 0 | 1 |
| 8/2/2011h | 96 | 91 (94.8) | 5 (5.2) | 1 | 0 | 1 | 3 |
| 8/4/2011h | 96 | 86 (89.6) | 10 (10.4) | 10 | 0 | 0 | 0 |
| 8/4/2011 | 96 | 91 (94.8) | 5 (5.2) | 5 | 0 | 0 | 0 |
| 8/5/2011 | 96 | 91 (94.8) | 5 (5.2) | 3 | 0 | 0 | 2 |
| 8/8/2011 | 96 | 88 (91.7) | 8 (8.3) | 6 | 0 | 0 | 2 |
| 8/9/2011 | 96 | 96 (100.0) | 0 | 0 | 0 | 0 | 0 |
| 9/6/2011 | 96 | 96 (100.0) | 0 | 0 | 0 | 0 | — |
| 9/6/2011 | 96 | 96 (100.0) | 0 | 0 | 0 | 0 | — |
| 9/6/2011 | 96 | 89 (92.7) | 7 (7.3) | 7 | 0 | 0 | — |
| 9/7/2011 | 96 | 89 (92.7) | 7 (7.3) | 7 | 0 | 0 | — |
| 9/7/2011 | 96 | 96 (100.0) | 0 | 0 | 0 | 0 | — |
| 9/7/2011 | 96 | 94 (97.9) | 2 (2.1) | 1 | 0 | 1 | — |
| 9/7/2011 | 96 | 95 (99.0) | 1 (1.0) | 1 | 0 | 0 | — |
| Ave = 94.9% ± 3.6% | 4.0% | — | 0.2% | 1.0% | |||
| n = 16 trials | |||||||
Well allocation is placement of one dechorionated 6-hpf embryo.
An unsuccessful well allocation (errorsc-e) was determined immediately after the trial and at 24 hpf for.
Embryo dead, usually partly or completely disintegrated, immediately after trial.
Embryo or disintegrated residue absent from well immediately after trial.
More than one embryo allocated to the same well.
Malformations observed at 24 hpf; Dash indicates not recorded 9/6–9/7.
Robotic performance was not different from the overall success of manual loading (F = 0.53; p < 0.01).
Lower apparent performance success of earlier trials reflects the performance of earlier software versions.
Discussion
We have introduced automated platforms for high-throughput chorion removal at 4 hpf and 96-well plate allocation at 6 hpf that consistently yielded 95% healthy embryos. Together, these automation platforms provide (1) a rapid and inexpensive circumventing of the potential for false-negative and false-positive results imposed by the chorion on high-throughput applications of the developmental zebrafish model and (2) a much less labor-intensive and more reliable means of carefully allocating dechorionated embryos to 96-well plates at a rate amenable to high-throughput screening.
This is the first report of an en masse chorion removal method for zebrafish embryos prior to 24 hpf, with a reproducibly high survival rate. Although chorion removal at 24 hpf from 50 to 100 zebrafish embryos at once has been reported, initiating embryo exposure so late in development is likely to be of limited utility for large-scale screens. Such screens lack a priori knowledge of compound activity and must therefore maximize opportunities for chemical “hits” by chemically exposing during the widest practical window of development. A recent report sought to quantify the success of pronase-supported dechorionation at 6 hpf, as described by Westerfield,17 for replicates of 50 embryos.14 That study concluded that the use of pronase was generally damaging to 6-hpf embryos and demonstrated a normal development rate of only 75% and a mortality rate of nearly 40%.14 Our demonstration of pronase-supported, automated chorion removal, at 4 hpf, from 1600 embryos at once, consistently yielded ≥95% survival and normal development to 120 hpf, indicating that a pronase-supported approach can be both practical and scalable to meet the embryo demands of high-throughput screening.
To our knowledge, this is also the first report of a reproducible method for robot-automated allocation of embryos to microtiter plates. A recent report described an image-based fluidic approach to rapid allocation of embryos to 96-well plates.18 Evidence of that system’s performance was largely limited to the handling of embryos still in the chorion with only cursory performance data from dechorionated embryos. We have demonstrated the highly reproducible use of a small, industrial robotic arm approach to retrieve single dechorionated embryos at 6 to 7 hpf from an unsorted dish and allocate them to a 96-well plate with a better than 95% survival rate. It would also be straightforward and, in some instances, desirable to allocate more than one embryo per well, such as for monitoring subtle locomotor activity where a higher signal-to-noise ratio is achieved with multiple embryos.7 For gross malformation end points, one embryo per well minimizes the potential effects of dose titration from uptake by multiple animals in the same 100-μL volume. Another practical extension of the automation would be for sorting transient transgenic reporter animals fluorescing at the embryonic stage and fluorescent-tagged morpholino-injected embryos in high-throughput gene knockdown assays.
We did not include the time to complete plate loading for any of the robotic trials shown in Table 2, focusing instead on the ability of the modified SCARA robot to handle embryos gently with high survival and low malformation rates. Figure 4 summarizes the entire process with approximate times for completion. We note that although several variables affected the time required for the robotic system to complete a 96-well plate, by far the most important variable was density and dispersal of embryos in the source dish. Having more than 300 embryos in the source dish, or failing to keep the embryos well dispersed with periodic swirling, noticeably slowed the rate at which the machine vision camera and software could select a sufficiently isolated embryo to map and direct the robot to retrieve. Once the source dish was depleted to less than 300 embryos and the dispersal was kept at a maximum, the system consistently loaded one 96-well plate every 15 min. For comparison, laboratory personnel that perform the task on a daily basis consistently loaded one 96-well plate every 6 to 10 min. However, three experienced loaders in our laboratory could only load a total of 18 to 20 plates before fatigue resulted in a successful loading rate of <95% when the plates were observed at 24 hpf. The robotic system, facing no such limitation, offers an obvious advantage when throughput demands require ≥200 plates per week. Moreover, having developed the software for a single system, the cost of scaling the system to multiple SCARA robotic loading stations will be limited to hardware only.
Figure 4.

Process summary and timeline for automated chorion removal and embryo allocation to plate wells. Step 1, beginning at 4 h postfertilization (hpf), consists of removal of dead or obviously abnormal embryos from the mass of 2000 embryos in a 135-mm Petri dish. It requires 5 to 10 min to complete the cleanup using a Pasteur pipette connected to a vacuum aspirator. Step 2, beginning immediately after 1, requires the approximate division of the mass of embryos among 4, 100-mm glass Petri dishes, the addition of a pronase aliquot to each, and pressing the start button. The automated gentle shaking, rinsing, and rest period requires 40 to 45 min. Step 3, beginning immediately after the post-dechorionation rest period and rinse to remove any remaining traces of chorion, simply requires the movement of the plate of 400 embryos to the Selective Compliant Assembly Robot Arm (SCARA) robot station and pressing the start button. A single embryo is delivered to each of the 96 wells in 15 min.
The automation platforms herein obviate persistent concerns about the chorion and its potential to limit the effects of chemical exposures, and they relieve a serious bottleneck to high-throughput use of the developing zebrafish by auto-mating embryo allocations to assay plates. We believe that these advances, coupled with advances in automated imaging and phenotype analysis, will quickly enable researchers to expand the scale and scope of toxicology and discovery research.
Acknowledgments
The authors wish to express their gratitude to Greg Gonnerman, Lindsey Chalker, Carrie Barton, Cari Buchner and Chapell Miller of the Tanguay Laboratory for providing a steady supply of high quality zebrafish eggs and for their insights on how to improve the automation. We thank Dr. Siba Das for Photoshop help with the figures.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The presented work was funded in part by NIH grants RC4 ES019764, P42 ES016465, and P30 ES000210.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
- 1.Hartung T. Lessons Learned from Alternative Methods and Their Validation for a New Toxicology in the 21st Century. J. Toxicol. Environ. Health B Crit. Rev. 2010;13(2-4):277–290. doi: 10.1080/10937404.2010.483945. [DOI] [PubMed] [Google Scholar]
- 2.Schmidt CW. TOX 21: New Dimensions of Toxicity Testing. Environ. Health Perspect. 2009;117(8):A348–A353. doi: 10.1289/ehp.117-a348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lilienblum W, Dekant W, Foth H, Gebel T, Hengstler JG, Kahl R, Kramer PJ, Schweinfurth H, Wollin KM. Alternative Methods to Safety Studies in Experimental Animals: Role in the Risk Assessment of Chemicals under the New European Chemicals Legislation (REACH) Arch. Toxicol. 2008;82(4):211–236. doi: 10.1007/s00204-008-0279-9. [DOI] [PubMed] [Google Scholar]
- 4.Zon LI, Peterson RT. In Vivo Drug Discovery in the Zebrafish. Nat. Rev. Drug Discov. 2005;4(1):35–44. doi: 10.1038/nrd1606. [DOI] [PubMed] [Google Scholar]
- 5.Sukardi H, Chng HT, Chan EC, Gong Z, Lam SH. Zebrafish for Drug Toxicity Screening: Bridging the In Vitro Cell-Based Models and In Vivo Mammalian Models. Expert Opin. Drug Metab. Toxicol. 2011;7(5):579–589. doi: 10.1517/17425255.2011.562197. [DOI] [PubMed] [Google Scholar]
- 6.Zon LI, Peterson R. The New Age of Chemical Screening in Zebrafish. Zebrafish. 2010;7(1):1. doi: 10.1089/zeb.2010.9996. [DOI] [PubMed] [Google Scholar]
- 7.Kokel D, Bryan J, Laggner C, White R, Cheung CY, Mateus R, Healey D, Kim S, Werdich AA, Haggarty SJ, et al. Rapid Behavior-Based Identification of Neuroactive Small Molecules in the Zebrafish. Nat. Chem. Biol. 2010;6(3):231–237. doi: 10.1038/nchembio.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Paik EJ, de Jong JL, Pugach E, Opara P, Zon LI. A Chemical Genetic Screen in Zebrafish for Pathways Interacting with cdx4 in Primitive Hematopoiesis. Zebrafish. 2010;7(1):61–68. doi: 10.1089/zeb.2009.0643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, Jang S, Haggarty SJ, Kokel D, Rubin LL, Peterson RT, et al. Zebrafish Behavioral Profiling Links Drugs to Biological Targets and Rest/Wake Regulation. Science. 2010;327(5963):348–351. doi: 10.1126/science.1183090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang C, Tao W, Wang Y, Bikow J, Lu B, Keating A, Verma S, Parker TG, Han R, Wen XY. Rosuvastatin, Identified from a Zebrafish Chemical Genetic Screen for Anti-angiogenic Compounds, Suppresses the Growth of Prostate Cancer. Eur. Urol. 2010;58(3):418–426. doi: 10.1016/j.eururo.2010.05.024. [DOI] [PubMed] [Google Scholar]
- 11.Yen J, Donerly S, Levin ED, Linney EA. Differential Acetyl cholinesterase Inhibition of Chlorpyrifos, Diazinon and Parathion in Larval Zebrafish. Neurotoxicol. Teratol. 2011;33(6):735–741. doi: 10.1016/j.ntt.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Johnson A, Carew E, Sloman KA. The Effects of Copper on the Morphological and Functional Development of Zebrafish Embryos. Aquat. Toxicol. 2007;84(4):431–438. doi: 10.1016/j.aquatox.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 13.Bar-Ilan O, Louis KM, Yang SP, Pedersen JA, Hamers RJ, Peterson RE, Heideman W. Titanium Dioxide Nanoparticles Produce Phototoxicity in the Developing Zebrafish. Nanotoxicology. 2011 Aug 10; doi: 10.3109/17435390.2011.604438. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 14.Henn K, Braunbeck T. Dechorionation as a Tool to Improve the Fish Embryo Toxicity Test (FET) with the Zebrafish (Danio rerio) Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011;153(1):91–98. doi: 10.1016/j.cbpc.2010.09.003. [DOI] [PubMed] [Google Scholar]
- 15.Zhou S, Dong Q, Li S, Guo J, Wang X, Zhu G. Developmental Toxicity of Cartap on Zebrafish Embryos. Aquat. Toxicol. 2009;95(4):339–346. doi: 10.1016/j.aquatox.2009.10.006. [DOI] [PubMed] [Google Scholar]
- 16.Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995;203(3):253–310. doi: 10.1002/aja.1002030302. [DOI] [PubMed] [Google Scholar]
- 17.Westerfield M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio) 5th ed University of Oregon Press; Eugene: 2007. [Google Scholar]
- 18.Graf SF, Hotzel S, Liebel U, Stemmer A, Knapp HF. Image-Based Fluidic Sorting System for Automated Zebrafish Egg Sorting into Multiwell Plates. J. Lab. Autom. 2011;16(2):105–111. doi: 10.1016/j.jala.2010.11.002. [DOI] [PubMed] [Google Scholar]