

Abstract
The brain changes in response to experience and altered environment. To do that, the nervous system often remodels the structures of neuronal circuits. This structural plasticity of the neuronal circuits appears to be controlled not only by intrinsic factors, but also by extrinsic mechanisms including modification of the extracellular matrix. Recent studies employing a range of animal models implicate that matrix metalloproteinases regulate multiple aspects of the neuronal development and remodeling in the brain. This paper aims to summarize recent advances of our knowledge on the neuronal functions of matrix metalloproteinases and discuss how they might relate in neuronal disease.
1. Introduction
In higher vertebrates, the space between neural cells in the brain is filled with material of the extracellular matrix (ECM). Both neurons and glial cells contribute to the production of the ECM components, and the ECM in turn mediates various structural and functional interactions between these cells [1, 2]. During early development, the ECM plays crucial roles in proliferation, migration and differentiation of neural cells. In the mature brain, the ECM undergoes a slow turnover and supports multiple physiological processes. In general, the mature ECM environment seems inhibitory for structural plasticity of neuronal circuits. For example, chondroitin sulfate proteoglycans appear to be one of inhibitory components in the ECM because their degradation by chondroitinase can reactivate ocular dominance plasticity [3]. It is thus likely that regulated proteolytic alteration of the ECM microenvironment should be required for the structural plasticity of neuronal circuits.
The ECM modifications in the nervous system are likely achieved by the concerted actions of several different proteases that are secreted by neurons and glial cells [4–6]. Among these proteinases, the matrix metalloproteinases (MMPs) family stands out as likely regulators of the neural plasticity. The mammalian central nervous system (CNS) contains over 10 different MMPs with detectable levels of transcripts or proteins [7, 8]. Studies of the temporal and spatial expression patterns of MMPs in the developing nervous system suggest that MMPs play important roles in neuronal development. In addition, expression of many MMPs has shown to change in response to injury or neurological disease [9]. Knockouts of particular MMPs significantly affect the injury and pathology, indicating that MMPs function as the crucial mediators of neuronal disease [10–12]. Interestingly, several MMP knockouts show deficits in learning and memory [10, 11]. Consistent with these notions, MMPs likely mediate the structural changes of dendritic spines as well as axon/dendrite structures in response to neuronal activity and in mental diseases [10, 11]. This paper considers potential roles for MMPs in neuronal development and plasticity, and discusses its alteration in injury and disease states.
2. MMPs in the Nervous System
Currently, 24 mammalian MMPs have been identified with distinct yet overlapping substrate specificities (Figure 1). Several MMPs are membrane anchored by transmembrane domains (MMP-14, -15, -16, -24) or by GPI links (MMP-17, -25). The expression of many MMPs has been detected in the nervous system and shown to change in response to injury and neurological disease [6–8]. Analysis of mRNA expression in the brain and spinal cord showed that MMP-2, -9, -11, -12, -13, -14, -15, and -24 are developmentally regulated, whereas mRNA levels of MMP-3, -7, and -10 remain unchanged throughout the neural development [6–8]. So far, two secreted types of MMPs, MMP-2, and MMP-9, are most frequently investigated MMPs in the brain because they are relatively easily detectable. MMP-2 is detected in various brain structures including astroglia and some pyramidal neurons in the cortex and Purkinje cells in the cerebellum, whereas MMP-9 is expressed in the hippocampus, cerebellum, and cortex, predominantly in neurons [6–8]. Levels of MMP-2 and MMP-9 are significantly elevated following ischemia, brain injury, and kainate treatment [6, 11], implying a role for MMP-2 and MMP-9 in remodeling of neural circuits in response to neural activity and brain damages.
Figure 1.
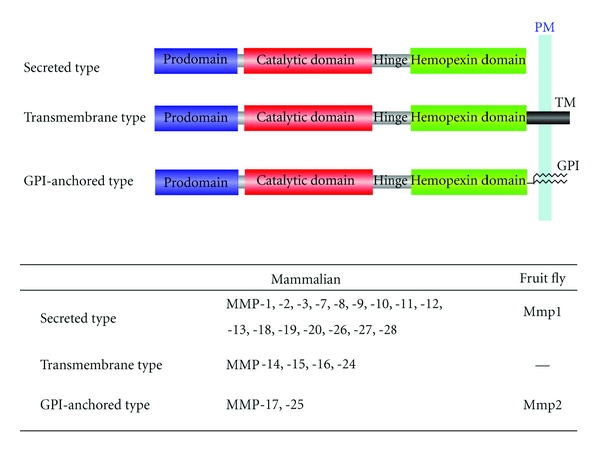
Domain structures of typical MMP family members in mammal and fruit fly. Hinge, hinge region; PM, plasma membrane; TM, transmembrane domain; GPI, glycosyl-phosphatidylinositol linker. The signal sequence located at the amino terminus of all prodomain is not shown.
In the fruit fly Drosophila melanogaster, there are only two MMP family members: Mmp1 and Mmp2 [13]. Mmp1 is a secreted proteinase, whereas Mmp2 is a GPI-anchored protein (Figure 1). Expression levels of Mmp1 and Mmp2 are dramatically elevated during metamorphosis in many tissues including the nervous system [13]. In agreement with the expression pattern, recent reports indicate that Mmp1 and Mmp2 play critical roles in remodeling of neural circuits in the peripheral nervous system [14, 15].
3. Secreted MMPs in Structural and Functional Plasticity of Synapses
Dendritic spine morphology and synaptic potentiation can both be dynamically modulated by proteins of the ECM and the cell-surface proteins with which they interact, which has long fueled the idea that regulated ECM remodeling has an important role in synaptic plasticity [16]. In the brain, MMPs are secreted by neurons and glial cells in an inactive (pro)form, and they become proteolytically active when several regulatory steps that result in removal of the propeptide are triggered in response to specific stimuli [5]. For example, studies have shown that in response to long-term potentiation (LTP) induction, MMP-9 rapidly becomes proteolytically active at perisynaptic sites and is essential for maintenance of LTP [17, 18]. Thus, perisynaptic MMP-9 proteolysis in response to LTP induction is likely critical for local remodeling of dendritic spine structures and functions necessary to support long-term synaptic plasticity [19–21].
LTP is a widely used cellular model for long-lasting synaptic plasticity that is thought to underlie learning and memory. MMP-9 knockout mice show behavioral impairments in hippocampus-dependent associative learning [11]. Furthermore, hippocampal slice cultures from MMP-9 knockout mice show impaired LTP, which can be restored by the application of recombinant MMP-9 [11]. In other studies, transient induction of hippocampal MMP-9 and MMP-3 levels was observed when animals were run in the Morris water maze, further supporting a role for MMPs in hippocampus-dependent learning [18]. Although the precise molecular mechanisms involved remain unclear, MMP-mediated synaptic potentiation can be inhibited in hippocampal slices by blocking integrin signaling, which suggests that MMP-9 regulates synaptic plasticity and LTP through integrins [11]. MMP-9 may exert its effects on integrins through the cleavage of laminin or other ECM components, exposing otherwise inaccessible RGD sites that can induce integrin signaling. It is also possible that MMP-9 may yield signaling functions through CD44 and integrins as shown in B cell lymphomas [22].
Alternatively, MMP-9 could regulate LTP by promoting lateral movement of glutamate receptors in excitatory synapses. Many synapses in the mature CNS are wrapped by a dense ECM, which likely acts as a spatial obstacle for glutamate receptors including N-methyl-D-aspartic acid- (NMDA-) type and alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic-acid- (AMPA) type receptors. It is thus assumed that modification of the ECM components might facilitate structural and functional plasticity in synapses. Indeed, a recent study using a high-resolution fluorescent in situ zymography (ISZ) shows colocalization of MMP-9 with synaptic glutamate receptors [23]. Michaluk et al. [24] showed that addition of recombinant MMP-9 in hippocampal cultured neurons increased lateral diffusion of NMDA receptors but not AMPA receptors. They further showed that the motility change was not mediated by alteration in the net ECM structure nor by direct cleavage of NMDA receptors, but rather through modification of an integrin-dependent pathway [24]. On the other hand, Frischknecht et al. [25] indicated that enzymatic removal of the ECM increased extrasynaptic receptor diffusion and the exchange of synaptic AMPA receptors but not NMDA receptors. It is thus possible that membrane diffusion of AMPA and NMDA receptors is differently regulated by the ECM and MMPs in synapses, which may contribute to functional difference of AMPA and NMDA receptors in synaptic plasticity.
4. MMPs in Axon Regeneration
Several lines of evidence point toward a potential role of MMPs in axonal regeneration following an injury. First, expression of MMPs has been shown to correspond with periods of recovery [26]. In regenerating sciatic nerve fibers, MMP-9 expression is colocalized with phosphorylated neurofilament M, a marker for regenerative elongation [27]. Furthermore, the phosphorylated neurofilament M is also induced by MMP-9 treatment and inhibited by an anti-MMP-9 antibody treatment [27]. Regenerating axons also show immunoreactivity for MMP-2 and MMP-3 [28].
A second line of evidence comes from studies of their application to nonpermissive substrates. When primary dorsal root ganglion (DRG) neurons are grown on cryostat sections of normal adult nerves, neurite outgrowth is poor as the adult nerves constitute an unfavorable substrate. However, when the nerve sections are treated with MMP-2, which degrades inhibitory chondroitin sulfate proteoglycans (CSPGs) to expose permissive laminin, neurite outgrowth from DRG neurons is promoted [29]. In a chronically denervated distal tibial nerve segments, successful regeneration of axons is facilitated by supernatant from a neural stem cell line containing large quantities of secreted MMP-2, which likely degrades the CSPGs in the chronically denerved nerve sections [30].
Wallerian degeneration occurs after injury and involves the breakdown of myelin and axons, and removal of degenerating nerve components. This clearance is necessary for eventual repair. MMP levels increase during Wallerian degeneration, and this correlation is functionally significant as Wallerian degeneration of a transected nerve is delayed by MMP inhibitors [31].
In summary, the data for MMPs in axonal regeneration is scant but still suggestive. More definitive evidence, perhaps in the form of application of MMPs to an area of injury in vivo to improve axonal regrowth, is required.
5. Membrane-Anchored MMPs Promote Dendrite Remodeling
The ECM exerts a strong influence on dendrite morphogenesis in cultured neurons, as the ECM can affect dendrite patterning in part through ECM-neurite adhesive contacts mediated by cell adhesion molecules such as integrins [32]. ECM-neurite interactions have also been implicated in regulating structural plasticity of dendrites in vivo. For instance, blockage of the integrin-ECM interaction in RGCs [33] or genetic ablation of the integrin-mediated signaling in adult cortical neurons [34] causes progressive retraction of dendritic branches.
The ECM modifications in the nervous system are likely achieved by the concerted actions of several different proteases that are secreted by neurons and glial cells [4–6]. Among these many proteases, MMPs stand out as likely regulators of the dendrite development and pathology since MMPs are dramatically upregulated in particular neurons of the developing brain and are often colocalized with dendrites [35, 36]. However, in large part due to issues of redundancy and compensation among over 20 vertebrate MMP family members, the in vivo role of MMPs in the nervous system remains to be established (Table 1).
Table 1.
MMP family proteins in neural circuit remodeling.
A recent study on Drosophila PNS neurons has provided in vivo evidence that ECM degradation facilitates dendrite remodeling [15]. During metamorphosis, the larval dendritic arbors of class IV sensory neurons are completely replaced with adult-specific processes as a result of extensive pruning and subsequent regeneration of dendritic arbors [14, 46]. Similar to their larval counterparts, dendrites of adult class IV neurons initially elaborate dendritic trees in a radial fashion and covered the whole body wall prior to eclosion. However, in contrast to what is observed during larval development [47–49], this radial arrangement of the dendritic arbor is rapidly rearranged to a lattice-like shape within 24 h after eclosion [15]. Time-lapse imaging revealed that this radial-to-lattice reshaping is largely due to rearrangement of the existing radial processes into a lattice-like pattern, rather than extensive pruning of the radially-arranged dendrites followed by regrowth of new arbors into a lattice pattern (Figure 2). Mutations in Mmp2, which encodes a GPI-anchored matrix metalloproteinase, block this radial-to-lattice reshaping of class IV dendrites without affecting other aspects of dendrite growth or development, and Mmp2 expression in epithelial cells adjacent to class IV dendrites is transiently increased at exactly the time when class IV dendrites undergo the radial-to-lattice reshaping. Therefore, epithelial Mmp2 promotes the dendrite reshaping through local modification of the basement membrane (BM) upon which class IV dendrites grow. These observations indicate that alteration of the ECM microenvironment might be a general mechanism for driving the structural plasticity of dendritic arbors in vivo (Figure 2).
Figure 2.
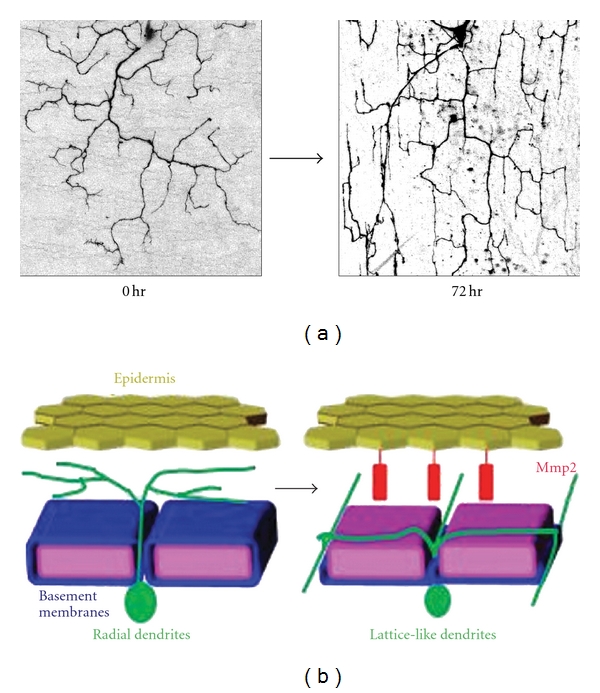
Dendrite reshaping in Drosophila sensory neurons is mediated by matrix metalloproteinases. (a) Dendritic pattern of a single class IV neurons in 0 hr and 72 hr posteclosion adults. (b) Dendrites of Drosophila sensory neurons are rapidly reshaped within 24 hours after eclosion. This dendrite reshaping is promoted by the matrix metalloproteinase Mmp2-mediated degradation of the basement membrane, suggesting that proteolytic alteration of the extracellular matrix plays a fundamental role in remodeling of dendritic structures during reorganization of neuronal circuits. This model is predominantly based on data from [15].
In the mouse cerebellum, membrane-type 5 MMP (MT5-MMP; also named as MMP-24) is expressed in developing dendrites of Purkinje cells, implicating a role for MT5-MMP in dendrogenesis [35]. The precise roles of MT5-MMP in the cerebellum are not known. Since structural remodeling in dendrites as well as synapses is reported in developing Purkinje cells [50], it is of interest to examine if membrane-anchored MMPs including MT5-MMP may play a role in dendrite remodeling the mammalian nervous system.
6. Potential Roles of MMPs in Neurogenesis
MMPs have recently been considered to be involved in the neurogenic response of adult neural stem/progenitor cells. MMPs are expressed abundantly in neural stem cells isolated from the human CNS [42]. Brain injuries including ischemia enhance neurogenesis in neuroproliferative regions of the adult rodent brain such as the subventricular zone (SVZ) of the lateral ventricles and the subgranular zone (SGZ) of the dentate gyrus (DG) of the hippocampus [43]. Although most MMPs are expressed at very low levels in the adult CNS, mRNA expression of both MMP-9 and MMP-2 increased severalfold in neural progenitor cells of SVZ after ischemic insult in adult rats [44]. Similarly, upregulation of MMP-9 and MMP-2 in the SGZ of the dentate gyrus was compatible with the peak of postischemic neurogenesis in adult primate brains [45]. These observations suggest that MMPs could be an important component in neurogenesis-associated processes in postischemic brain hippocampus. Physiological significance of the MMP expressions in the neurogenic regions remains to be determined. One likely possibility is that, as shown in the developing brain, MMPs might promote proliferation, neurite extension, and migration of newly born neurons. Alternatively, MMPs might play a role in providing an optimal niche for neural stem/progenitor cells by modulating the ECM environment.
7. MMPs in Neuronal Injury and Disease
MMP expression levels are elevated after nervous system injury and in a number of neuronal pathologies. For example, MMP-9 expression is elevated shortly after ischemia and stroke [9]. Likewise, multiple MMPs including MMP-3, -7, -10, -11, -19, and -20 are immediately induced within 24 h of an acute insult such as spinal cord compression injury [51]. During the period of peak signs in experimental autoimmune encephalomyelitis (EAE), an animal model of multiple sclerosis and the transcripts encoding the majority of MMPs are elevated [52]. In epileptogenesis, serum MMP-9 levels and the ratio of MMP-9 to tissue inhibitor of metalloproteinase-1 are elevated in children with various febrile seizures and convulsive status epilepticus [53]. After seizure, MMP-9 mRNA is transported to dendrites and synapses in the hippocampal DG of kainic acid-treated rats [54]. Wilczynski et al. showed that the sensitivity to PTZ epileptogenesis was decreased in MMP-9 knockout mice but is increased in transgenic rats overexpressing MMP-9 [41].
The increase of many MMPs in CNS pathology raises the question of which of these enzymes are important for the pathophysiological process. The alleviation of disease pathology in response to MMP inhibitors shows that the net effect of the expression of MMPs in CNS injury is detrimental. In this regard, MMP-9 null mice have been found to be less afflicted than wild-type mice by EAE and stroke, whereas MMP-9 and MMP-12 null mice recover better from spinal cord injury [55, 56]. MMP-2 null mice have been found to be more suspective to EAE, which have been attributed to compensatory increase in MMP-9 in these animals [57]. In the mouse model of EAE, MMP-9 promotes development and progression of the disease whereas MMP-12 has a role in its resolution [58], suggesting that different MMPs might have distinct roles in EAE development. Indeed, double MMP-2 and -9 knockout mice are resistant to EAE development [59], indicating that combined MMP-2 and -9 activities are crucial EAE.
In addition to the brain injury and diseases, MMPs are implicated in drug addiction. The MMP inhibitors often suppressed acquisition of cocain-induced conditioned place preference (CPP) [60]. In consistent with the inhibitor studies, MMP protein levels in the brain were changed in cocain abusers as well as cocain-treated animals [61]. The role of MMPs in drug addiction was further confirmed by the findings that MMP-2 and MMP-9 deficient mice displayed attenuated sensitization and cocain CPP [62, 63].
MMPs are recently shown to be involved in development of neuropathic pain, which is characterized by mechanical allodynia that is, painful responses to previously nonpainful mechanical stimuli [64]. In L5 spinal nerve ligation, MMP-9 shows a rapid and transient upregulation in the injured DRG sensory neurons, whereas MMP-2 shows a delayed response in DRG satellite cells and spinal astrocytes. Local inhibition of MMP-9 by an intrathecal route inhibits the early phase of neuropathic pain, whereas inhibition of MMP-2 suppresses the late phase of neuropathic pain. These results suggest that distinct MMPs play different roles in early and late-phase development of neuropathic pain and that MMP inhibitors may provide a therapeutic approach for the treatment of neuropathic pain.
A recent report suggests a possible role of MMPs in Huntington's disease (HD) pathogenesis [65]. HD, the most frequent of a group of nine inherited neurodegenerative polyglutamine disorders, is caused by an expanded CAG triplet repeat in exon 1 of the huntingtin gene that encodes a stretch of polyglutamine (polyQ) residues close to the N-terminus of the huntingtin (Htt) protein. Htt is known to be cleaved by various proteases including Caspases and calpains, and inhibition of the mutant Htt proteolysis reduces neurotoxicity, indicating an important role for Htt proteolysis in HD pathogenesis. Miller et al. examined Htt proteolytic processing by screening for enzymes that cleave mutant Htt and found that MMP-10 is responsible for the Htt proteolysis to produce small N-terminal toxic fragments. A nonpeptidic inhibitor of MMPs NNHG, and two known endogenous MMPs inhibitors, TIMP1 and TIMP3, blocked Htt-mediated toxicity and produced beneficial therapeutic effects, providing new insights into Htt proteolysis and its potential as a therapeutic target.
8. Future Perspectives
This paper highlights the diversity and importance of MMPs in neuronal development, plasticity, and maintenance of neuronal health. Many questions await further studies, including which member or members of the MMPs are important in a particular condition or pathological state, how they achieve their effects, and what roles each MMP has in the overall scheme. MMPs are also implicated in diseases of the nervous system, and it is important to target their activity for therapy. The study of MMP functions should open up new vistas in the nervous system physiology and pathology.
Authers' Contribution
These authors contribute equally to this paper.
References
- 1.Sorokin L. The impact of the extracellular matrix on inflammation. Nature Reviews Immunology. 2010;10(10):712–723. doi: 10.1038/nri2852. [DOI] [PubMed] [Google Scholar]
- 2.Faissner A, Pyka M, Geissler M, et al. Contributions of astrocytes to synapse formation and maturation—potential functions of the perisynaptic extracellular matrix. Brain Research Reviews. 2010;63(1-2):26–38. doi: 10.1016/j.brainresrev.2010.01.001. [DOI] [PubMed] [Google Scholar]
- 3.Pizzorusso T, Medini P, Berardi N, Chierzi S, Fawcett JW, Maffei L. Reactivation of ocular dominance plasticity in the adult visual cortex. Science. 2002;298(5596):1248–1251. doi: 10.1126/science.1072699. [DOI] [PubMed] [Google Scholar]
- 4.Dzwonek J, Rylski M, Kaczmarek L. Matrix metalloproteinases and their endogenous inhibitors in neuronal physiology of the adult brain. FEBS Letters. 2004;567(1):129–135. doi: 10.1016/j.febslet.2004.03.070. [DOI] [PubMed] [Google Scholar]
- 5.Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nature Reviews Molecular Cell Biology. 2007;8(3):221–233. doi: 10.1038/nrm2125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yong VW. Metalloproteinases: mediators of pathology and regeneration in the CNS. Nature Reviews Neuroscience. 2005;6(12):931–944. doi: 10.1038/nrn1807. [DOI] [PubMed] [Google Scholar]
- 7.Pagenstecher A, Stalder AK, Kincaid CL, Shapiro SD, Campbell IL. Differential expression of matrix metalloproteinase and tissue inhibitor of matrix metalloproteinase genes in the mouse central nervous system in normal and inflammatory states. American Journal of Pathology. 1998;152(3):729–741. [PMC free article] [PubMed] [Google Scholar]
- 8.Ulrich R, Gerhauser I, Seeliger F, Baumgärtner W, Alldinger S. Matrix metalloproteinases and their inhibitors in the developing mouse brain and spinal cord: a reverse transcription quantitative polymerase chain reaction study. Developmental Neuroscience. 2005;27(6):408–418. doi: 10.1159/000088455. [DOI] [PubMed] [Google Scholar]
- 9.Yong VW, Power C, Forsyth P, Edwards DR. Metalloproteinases in biology and pathology of the nervous system. Nature Reviews Neuroscience. 2001;2(7):502–511. doi: 10.1038/35081571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hu J, Van den Steen PE, Sang QXA, Opdenakker G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nature Reviews Drug Discovery. 2007;6(6):480–498. doi: 10.1038/nrd2308. [DOI] [PubMed] [Google Scholar]
- 11.Ethell IM, Ethell DW. Matrix metalloproteinases in brain development and remodeling: synaptic functions and targets. Journal of Neuroscience Research. 2007;85(13):2813–2823. doi: 10.1002/jnr.21273. [DOI] [PubMed] [Google Scholar]
- 12.Van Den Steen PE, Dubois B, Nelissen I, Rudd PM, Dwek RA, Opdenakker G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9) Critical Reviews in Biochemistry and Molecular Biology. 2002;37(6):375–536. doi: 10.1080/10409230290771546. [DOI] [PubMed] [Google Scholar]
- 13.Page-McCaw A, Serano J, Santë JM, Rubin GM. Drosophila matrix metalloproteinases are required for tissue remodeling, but not embryonic development. Developmental Cell. 2003;4(1):95–106. doi: 10.1016/s1534-5807(02)00400-8. [DOI] [PubMed] [Google Scholar]
- 14.Kuo CT, Jan LY, Jan YN. Dendrite-specific remodeling of Drosophila sensory neurons requires matrix metalloproteases, ubiquitin-proteasome, and ecdysone signaling. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(42):15230–15235. doi: 10.1073/pnas.0507393102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yasunaga KI, Kanamori T, Morikawa R, Suzuki E, Emoto K. Dendrite Reshaping of Adult Drosophila Sensory Neurons Requires Matrix Metalloproteinase-Mediated Modification of the Basement Membranes. Developmental Cell. 2010;18(4):621–632. doi: 10.1016/j.devcel.2010.02.010. [DOI] [PubMed] [Google Scholar]
- 16.Dityatev A, Schachner M, Sonderegger P. The dual role of the extracellular matrix in synaptic plasticity and homeostasis. Nature Reviews Neuroscience. 2010;11(11):735–746. doi: 10.1038/nrn2898. [DOI] [PubMed] [Google Scholar]
- 17.Nagy V, Bozdagi O, Matynia A, et al. Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory. Journal of Neuroscience. 2006;26(7):1923–1934. doi: 10.1523/JNEUROSCI.4359-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Meighan SE, Meighan PC, Choudhury P, et al. Effects of extracellular matrix-degrading proteases matrix metalloproteinases 3 and 9 on spatial learning and synaptic plasticity. Journal of Neurochemistry. 2006;96(5):1227–1241. doi: 10.1111/j.1471-4159.2005.03565.x. [DOI] [PubMed] [Google Scholar]
- 19.Wang XB, Bozdagi O, Nikitczuk JS, Zu WZ, Zhou Q, Huntley GW. Extracellular proteolysis by matrix metalloproteinase-9 drives dendritic spine enlargement and long-term potentiation coordinately. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(49):19520–19525. doi: 10.1073/pnas.0807248105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Conant K, Wang Y, Szklarczyk A, Dudak A, Mattson MP, Lim ST. Matrix metalloproteinase-dependent shedding of intercellular adhesion molecule-5 occurs with long-term potentiation. Neuroscience. 2010;166(2):508–521. doi: 10.1016/j.neuroscience.2009.12.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Michaluk P, Wawrzyniak M, Alot P, et al. Influence of matrix metalloproteinase MMP-9 on dendritic spine morphology. Journal of Cell Science. 2011;124(19):3369–3380. doi: 10.1242/jcs.090852. [DOI] [PubMed] [Google Scholar]
- 22.Redondo-Muñoz J, Ugarte-Berzal E, Terol MJ, et al. Matrix metalloproteinase-9 promotes chronic lymphocytic leukemia B cell survival through its hemopexin domain. Cancer Cell. 2010;17(2):160–172. doi: 10.1016/j.ccr.2009.12.044. [DOI] [PubMed] [Google Scholar]
- 23.Gawlak M, Górkiewicz T, Gorlewicz A, Konopacki FA, Kaczmarek L, Wilczynski GM. High resolution in situ zymography reveals matrix metalloproteinase activity at glutamatergic synapses. Neuroscience. 2009;158(1):167–176. doi: 10.1016/j.neuroscience.2008.05.045. [DOI] [PubMed] [Google Scholar]
- 24.Michaluk P, Mikasova L, Groc L, Frischknecht R, Choquet D, Kaczmarek L. Matrix metalloproteinase-9 controls NMDA receptor surface diffusion through integrin β1 signaling. Journal of Neuroscience. 2009;29(18):6007–6012. doi: 10.1523/JNEUROSCI.5346-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Frischknecht R, Heine M, Perrais D, Seidenbecher CI, Choquet D, Gundelfinger ED. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nature Neuroscience. 2009;12(7):897–904. doi: 10.1038/nn.2338. [DOI] [PubMed] [Google Scholar]
- 26.Duchossoy Y, Horvat JC, Stettler O. MMP-related gelatinase activity is strongly induced in scar tissue of injured adult spinal cord and forms pathways for ingrowing neurites. Molecular and Cellular Neuroscience. 2001;17(6):945–956. doi: 10.1006/mcne.2001.0986. [DOI] [PubMed] [Google Scholar]
- 27.Shubayev VI, Myers RR. Matrix metalloproteinase-9 promotes nerve growth factor-induced neurite elongation but not new sprout formation in vitro. Journal of Neuroscience Research. 2004;77(2):229–239. doi: 10.1002/jnr.20160. [DOI] [PubMed] [Google Scholar]
- 28.Ahmed Z, Dent RG, Leadbeater WE, Smith C, Berry M, Logan A. Matrix metalloproteases: degradation of the inhibitory environment of the transected optic nerve and the scar by regenerating axons. Molecular and Cellular Neuroscience. 2005;28(1):64–78. doi: 10.1016/j.mcn.2004.08.013. [DOI] [PubMed] [Google Scholar]
- 29.Zuo J, Neubauer D, Dyess K, Ferguson TA, Muir D. Degradation of chondroitin sulfate proteoglycan enhances the neurite- promoting potential of spinal cord tissue. Experimental Neurology. 1998;154(2):654–662. doi: 10.1006/exnr.1998.6951. [DOI] [PubMed] [Google Scholar]
- 30.Heine W, Conant K, Griffin JW, Höke A. Transplanted neural stem cells promote axonal regeneration through chronically denervated peripheral nerves. Experimental Neurology. 2004;189(2):231–240. doi: 10.1016/j.expneurol.2004.06.014. [DOI] [PubMed] [Google Scholar]
- 31.Siebert H, Dippel N, Mäder M, Weber F, Brück W. Matrix metalloproteinase expression and inhibition after sciatic nerve axotomy. Journal of Neuropathology and Experimental Neurology. 2001;60(1):85–93. doi: 10.1093/jnen/60.1.85. [DOI] [PubMed] [Google Scholar]
- 32.Reichardt LF, Tomaselli KJ. Extracellular matrix molecules and their receptors: functions in neural development. Annual Review of Neuroscience. 1991;14:531–570. doi: 10.1146/annurev.ne.14.030191.002531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Marrs GS, Honda T, Fuller L, et al. Dendritic arbors of developing retinal ganglion cells are stabilized by β1-integrins. Molecular and Cellular Neuroscience. 2006;32(3):230–241. doi: 10.1016/j.mcn.2006.04.005. [DOI] [PubMed] [Google Scholar]
- 34.Moresco EMY, Donaldson S, Williamson A, Koleske AJ. Integrin-mediated dendrite branch maintenance requires Abelson (Abl) family kinases. Journal of Neuroscience. 2005;25(26):6105–6118. doi: 10.1523/JNEUROSCI.1432-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sekine-Aizawa Y, Hama E, Watanabe K, et al. Matrix metalloproteinase (MMP) system in brain: identification and characterization of brain-specific MMP highly expressed in cerebellum. European Journal of Neuroscience. 2001;13(5):935–948. doi: 10.1046/j.0953-816x.2001.01462.x. [DOI] [PubMed] [Google Scholar]
- 36.Szklarczyk A, Lapinska J, Rylski M, McKay RDG, Kaczmarek L. Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. Journal of Neuroscience. 2002;22(3):920–930. doi: 10.1523/JNEUROSCI.22-03-00920.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Saygili E, Schauerte P, Pekassa M, et al. Sympathetic neurons express and secrete MMP-2 and MT1-MMP to control nerve sprouting via pro-NGF conversion. Cellular and Molecular Neurobiology. 2011;31:17–25. doi: 10.1007/s10571-010-9548-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Miller CM, Liu N, Page-Mccaw A, Broihier HT. Drosophila Mmp2 regulates the matrix molecule faulty attraction (Frac) to promote motor axon targeting in Drosophila . Journal of Neuroscience. 2011;31(14):5335–5347. doi: 10.1523/JNEUROSCI.4811-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Miller CM, Page-McCaw A, Broihier HT. Matrix metalloproteinases promote motor axon fasciculation in the Drosophila embryo. Development. 2008;135(1):95–109. doi: 10.1242/dev.011072. [DOI] [PubMed] [Google Scholar]
- 40.Gonthier B, Koncina E, Satkauskas S, et al. A PKC-dependent recruitment of MMP-2 controls semaphorin-3A growth-promoting effect in cortical dendrites. PLoS One. 2009;4(4) doi: 10.1371/journal.pone.0005099. Article ID e5099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wilczynski GM, Konopacki FA, Wilczek E, et al. Important role of matrix metalloproteinase 9 in epileptogenesis. Journal of Cell Biology. 2008;180(5):1021–1035. doi: 10.1083/jcb.200708213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Frölichsthal-Schoeller P, Vescovi AL, Krekoski CA, Murphy G, Edwards DR, Forsyth P. Expression and modulation of matrix metalloproteinase-2 and tissue inhibitors of metalloproteinases in human embryonic CNS stem cells. NeuroReport. 1999;10(2):345–351. doi: 10.1097/00001756-199902050-00025. [DOI] [PubMed] [Google Scholar]
- 43.Wójcik-Stanaszek L, Sypecka J, Szymczak P, et al. The potential role of metalloproteinases in neurogenesis in the gerbil hippocampus following global forebrain ischemia. PLoS ONE. 2011;6(7) doi: 10.1371/journal.pone.0022465. Article ID e22465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Morris DC, Zhang ZG, Zhang R, LeTourrneau Y, Gregg SR, Chopp M. Stroke increases expression of matrix metalloproteinases and P21-activeted protein kinase in neural progenitor cells. Academic Emergency Medicine. 2006;13:p. S194. [Google Scholar]
- 45.Lu L, Tonchev AB, Kaplamadzhiev DB, et al. Expression of matrix metalloproteinases in the neurogenic niche of the adult monkey hippocampus after ischemia. Hippocampus. 2008;18(10):1074–1084. doi: 10.1002/hipo.20466. [DOI] [PubMed] [Google Scholar]
- 46.Williams DW, Truman JW. Cellular mechanisms of dendrite pruning in Drosophila: insights from in vivo time-lapse of remodeling dendritic arborizing sensory neurons. Development. 2005;132(16):3631–3642. doi: 10.1242/dev.01928. [DOI] [PubMed] [Google Scholar]
- 47.Emoto K, He Y, Ye B, et al. Control of dendritic branching and tiling by the tricornered-kinase/furry signaling pathway in Drosophila sensory neurons. Cell. 2004;119(2):245–256. doi: 10.1016/j.cell.2004.09.036. [DOI] [PubMed] [Google Scholar]
- 48.Emoto K, Parrish JZ, Jan LY, Jan YN. The tumour suppressor Hippo acts with the NDR kinases in dendritic tiling and maintenance. Nature. 2006;443(7108):210–213. doi: 10.1038/nature05090. [DOI] [PubMed] [Google Scholar]
- 49.Koike-Kumagai M, Yasunaga KI, Morikawa R, Kanamori T, Emoto K. The target of rapamycin complex 2 controls dendritic tiling of Drosophila sensory neurons through the Tricornered kinase signalling pathway. EMBO Journal. 2009;28(24):3879–3892. doi: 10.1038/emboj.2009.312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kaneko M, Yamaguchi K, Eiraku M, et al. Remodeling of monoplanar purkinje cell dendrites during cerebellar circuit formation. PLoS One. 2011;6(5) doi: 10.1371/journal.pone.0020108. Article ID e20108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wells JEA, Hurlbert RJ, Fehlings MG, Yong VW. Neuroprotection by minocycline facilitates significant recovery from spinal cord injury in mice. Brain. 2003;126(7):1628–1637. doi: 10.1093/brain/awg178. [DOI] [PubMed] [Google Scholar]
- 52.Weaver A, Goncalves Da Silva A, Nuttall RK, et al. An elevated matrix metalloproteinase (MMP) in an animal model of multiple sclerosis is protective by affecting Th1/Th2 polarization. FASEB Journal. 2005;19(12):1668–1670. doi: 10.1096/fj.04-2030fje. [DOI] [PubMed] [Google Scholar]
- 53.Suenaga N, Ichiyama T, Kubota M, Isumi H, Tohyama J, Furukawa S. Roles of matrix metalloproteinase-9 and tissue inhibitors of metalloproteinases 1 in acute encephalopathy following prolonged febrile seizures. Journal of the Neurological Sciences. 2008;266(1-2):126–130. doi: 10.1016/j.jns.2007.09.011. [DOI] [PubMed] [Google Scholar]
- 54.Konopacki FA, Rylski M, Wilczek E, et al. Synaptic localization of seizure-induced matrix metalloproteinase-9 mRNA. Neuroscience. 2007;150(1):31–39. doi: 10.1016/j.neuroscience.2007.08.026. [DOI] [PubMed] [Google Scholar]
- 55.Dubois B, Masure S, Hurtenbach U, et al. Resistance of young gelatinase B-deficient mice to experimental autoimmune encephalomyelitis and necrotizing tail lesions. Journal of Clinical Investigation. 1999;104(11):1507–1515. doi: 10.1172/JCI6886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Noble LJ, Donovan F, Igarashi T, Goussev S, Werb Z. Matrix metalloproteinases limit functional recovery after spinal cord injury by modulation of early vascular events. Journal of Neuroscience. 2002;22(17):7526–7535. doi: 10.1523/JNEUROSCI.22-17-07526.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Esparza J, Kruse M, Lee J, Michaud M, Madri JA. MMP-2 null mice exhibit an early onset and severe experimental autoimmune encephalomyelitis due to an increase in MMP-9 expression and activity. FASEB Journal. 2004;18(14):1682–1691. doi: 10.1096/fj.04-2445com. [DOI] [PubMed] [Google Scholar]
- 58.DaSilva AG, Yong VW. Matrix metalloproteinase-12 deficiency worsens relapsing-remitting experimental autoimmune encephalomyelitis in association with cytokine and chemokine dysregulation. American Journal of Pathology. 2009;174(3):898–909. doi: 10.2353/ajpath.2009.080952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Agrawal S, Anderson P, Durbeej M, et al. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. Journal of Experimental Medicine. 2006;203(4):1007–1019. doi: 10.1084/jem.20051342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mash DC, ffrench-Mullen J, Adi N, Qin Y, Buck A, Pablo J. Gene expression in human hippocampus from cocaine abusers identifies genes which regulate extracellular matrix remodeling. PLoS ONE. 2007;2(11):p. e1187. doi: 10.1371/journal.pone.0001187. Article ID e1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Brown TE, Forquer MR, Harding JW, Wright JW, Sorg BA. Increase in matrix metalloproteinase-9 levels in the rat medial prefrontal cortex after cocaine reinstatement of conditioned place preference. Synapse. 2008;62(12):886–889. doi: 10.1002/syn.20562. [DOI] [PubMed] [Google Scholar]
- 62.Mizoguchi H, Yamada K, Niwa M, et al. Reduction of methamphetamine-induced sensitization and reward in matrix metalloproteinase-2 and -9-deficient mice. Journal of Neurochemistry. 2007;100(6):1579–1588. doi: 10.1111/j.1471-4159.2006.04288.x. [DOI] [PubMed] [Google Scholar]
- 63.Liu Y, Brown S, Shaikh J, Fishback JA, Matsumoto RR. Relationship between methamphetamine exposure and matrix metalloproteinase 9 expression. NeuroReport. 2008;19(14):1407–1409. doi: 10.1097/WNR.0b013e32830dd606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kawasaki Y, Xu ZZ, Wang X, et al. Distinct roles of matrix metalloproteases in the early- and late-phase development of neuropathic pain. Nature Medicine. 2008;14(3):331–336. doi: 10.1038/nm1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Miller JP, Holcomb J, Al-Ramahi I, et al. Matrix metalloproteinases are modifiers of huntingtin proteolysis and toxicity in Huntington’s disease. Neuron. 2010;67(2):199–212. doi: 10.1016/j.neuron.2010.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]