

Abstract
Many genes have been implicated in schizophrenia as have viral prenatal or adult infections and toxoplasmosis or Lyme disease. Several autoantigens also target key pathology-related proteins. These factors are interrelated. Susceptibility genes encode for proteins homologous to those of the pathogens while the autoantigens are homologous to pathogens' proteins, suggesting that the risk-promoting effects of genes and risk factors are conditional upon each other, and dependent upon protein matching between pathogen and susceptibility gene products. Pathogens' proteins may act as dummy ligands, decoy receptors, or via interactome interference. Many such proteins are immunogenic suggesting that antibody mediated knockdown of multiple schizophrenia gene products could contribute to the disease, explaining the immune activation in the brain and lymphocytes in schizophrenia, and the preponderance of immune-related gene variants in the schizophrenia genome. Schizophrenia may thus be a “pathogenetic” autoimmune disorder, caused by pathogens, genes, and the immune system acting together, and perhaps preventable by pathogen elimination, or curable by the removal of culpable antibodies and antigens.
1. Introduction
Over 600 genes have been implicated in schizophrenia in association studies, supporting the contention that multiple genes of small effect contribute to this condition [1, 2] (see http://www.polygenicpathways.co.uk/schizgenesandfunc.htm for association references). These genes cluster together in clearly defined signalling networks related to the diverse subpathologies of schizophrenia [3–7]. Epistasis between genes within these same signalling networks markedly affects the degree of risk-promotion [8–10], in part, explaining the inconsistency in genetic association studies.
Schizophrenia has also been associated with prenatal complications including maternal rubella (German measles) [11], influenza [12, 13], Varicella zoster (chicken pox) [14], Herpes (HSV-2) [15], common cold infection with fever [16], or poliovirus infection [17] while in childhood or adulthood, coxsackie virus infection (in neonates [18]) or Lyme disease (vectored by the Ixodes tick and Borrelia Burgdorferri) or Toxoplasmosis have been reported as risk factors [19, 20] (see Table 1). The human endogenous retrovirus, HERV-W, has also been implicated in schizophrenia [21]. A number of schizophrenia-related genes are implicated in the life cycles of these pathogens, suggesting an interplay between genes and risk factors [22].
Table 1.
Some of the pathogens implicated in Schizophrenia, either in relation to maternal infection, or to infection in later life.
| Pre- and perinatal maternal infection | Juvenile (in offspring) | Adult |
|---|---|---|
| Rubella (first trimester) [76]: Influenza (first trimester) [13] Influenza or common cold with fever (second trimester) [16] |
Mumps or cytomegalovirus infection (0–12 years old) [77] |
HSV-1 seropositivity related to grey matter volume [78] |
|
| ||
| Poliovirus (second trimester) [17] | Coxsackie B5 infection perinatally [18] | HSV-1 (in Afro-Americans) or HHV-6 seropositivity: Inverse correlation with HSV-2 and cytomegalovirus [79] |
|
| ||
| Measles, Varicella zoster or polio (seropositivity at birth) [14] | Childhood meningitis (0–4 years old) [80] | Borna disease virus seropositivty [81] |
|
| ||
| HSV-2 (antibodies assayed at the end of pregnancy) [82] | Coronavirus seropositivity [83] | |
|
| ||
| Influenza B (seropositivity at birth) [84] | Elevated retrovirus HERV-W transcripts [85] | |
|
| ||
| Toxoplasmosis (antibodies during pregnancy) [86] | Measles virus seropositivity [87] | |
|
| ||
| Hepatitis C [38] | ||
|
| ||
| Toxoplasmosis [88] | ||
|
| ||
| Correlation with the incidence of Lyme disease (Borrelia) [20] | ||
Many schizophrenia genes relate to the immune network [5, 6, 22, 23]. Immune activation is also observed in the schizophrenic brain [24, 25] or in lymphocytes [26–29]. A number of autoantigens/autoantibodies to key schizophrenia-related proteins have also been reported. These include dopamine, serotonin, acetylcholine, and NMDA receptors; inter alia (Table 2). Maternal immune activation in animal models has also been shown to generate phenotypes relevant to schizophrenia in the offspring [30].
Table 2.
Pathogens expressing proteins with homology to the autoantigens reported in schizophrenia. The size of the tags is proportional the number of pathogen's proteins that are homologous to the autoantigen. Note that the profile is different for each pathogen. The original BLAST files can be found at http://www.polygenicpathways.co.uk/blasts.htm.

|
As shown below, genes, risk factors, and immunity can be linked together forming a unifying pathway whose elements are interdependent. Dysfunction of this network which is conditional upon interactions between its three branches may be responsible for schizophrenia.
2. Methods
The human herpesvirus 2 genome (NC_001798) as well as those of the rhinovirus (NC_001490), rubella (NC_001545.1) and Varicella zoster (NC_001348.1) and HERV-W (NP_055405.3: env polyprotein) viruses, Borrelia Burgdorferri (NC_011728) and T. Gondii (NC_001799: Partial genome) were screened against the human proteome using the NCBI BLAST server and the Entrez query filter “schizophrenia”. The HERV-W, influenza, HSV-2 and rubella viruses were also screened unfiltered (Translated pathogen genome versus human proteins: BlastX) [31]. The BLAST algorithm detects overall homology between entire gene or protein sequences, and it is necessary to set parameters to low significance levels in order to detect short intraprotein consensus homology. The parameters used were: Expect 20,000, E value = 100,000; matrix PAM30. The original BLAST results are stocked at http://www.polygenicpathways.co.uk/blasts.htm. Information for all abbreviations is available at this site, provided by the NextBio highlighting service.
BLAST files were scanned by an online tag cloud generator producing tags sized according to gene word occurrences http://www.tagcloud-generator.com/generator.php#anker. Word occurrences were counted using a “Highlightall add-in” for Firefox https://addons.mozilla.org/en-US/firefox/addon/4240/.
Antigenicity (B-cell epitope prediction) was estimated using the BepiPred server http://www.cbs.dtu.dk/services/BepiPred/ [32] (Table 4).
Table 4.
Viruses reported to bind to DISC1 interactome partners.
| DISC1 partner gene symbol | Protein name | Viral binder |
|---|---|---|
| ACTG1 | Actin, cytoplasmic 2 | HIV-1 [97] HSV1 [98] |
| ACTN2 | Actinin, alpha 2 | Hepatitis C [99] |
| AKAP9 | A-kinase anchor protein 9 | Epstein-Barr [97] |
| ATF4 | Cyclic AMP-dependent transcription factor ATF-4 | HSV1 [98] |
| ATF5 | Cyclic AMP-dependent transcription factor ATF-5 | HTLV1 [100] |
| BICD1 | Protein bicaudal D homolog 1 | Cytomegalovirus [101] |
| C14orf135 | Uncharacterized protein C14orf135 precursor | Hepatitis C [102] |
| DCTN1 | Dynactin-1 | HSV1 [98] |
| DCTN2 | Dynactin subunit 2 | Dynactins are involved in the transport of the adenoviruses, HSV-1, the hantaan virus, HTLV-1 and the poliovirus [103–108] |
| DNAJC7 | DnaJ homolog subfamily C member 7 | Part of a complex forming the coxsackie virus receptor [109] |
| DYNC1H1 | Dynein heavy chain, cytosolic | Adenovirus (in a complex with dynactin and NDEL1) [110] |
| EEF2 | Elongation factor 2 | Epstein Barr [111] |
| EIF3S3 | Eukaryotic translation initiation factor 3 subunit 3 | Hepatitis C [112] |
| FEZ1 | Fasciculation and elongation protein zeta 1 (zygin I) | JC Polyomavirus [113] |
| HERC2 | HECT domain and RCC1-like domain-containing protein 2 | Papillomavirus 16 [114] |
| KIF3C | Kinesin-like protein KIF3C | HIV-1 [115] |
| MATR3 | Matrin-3 | HSV1 [98] |
| NDEL1 | Nuclear distribution protein nudE-like 1 | Part of a complex involved in Adenovirus transport (with dynactin and cytoplasmic dynein) [110] |
| PAFAH1B1 | Platelet-activating factor acetylhydrolase IB subunit alpha | Binds to Poliovirus P3 protein and HIV-1 Tat [116, 117] |
| PCNT | Pericentrin | Involved in the microtubular transport of the adenovirus [118] |
| PGK1 | Phosphoglycerate kinase 1 | Epstein-Barr [119] |
| SMARCE1 | SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily E member 1 | HSV-1 [97] |
| STX18 | Syntaxin-18 | Papillomavirus [119] |
| TNKS | Tankyrase-1 | Epstein-Barr [120] |
| TUBB | Tubulin beta chain | Epstein-Barr [119] |
| YWHAE | 14-3-3 protein epsilon | Hepatitis C [97] : L : Epstein-Barr [119] |
| YWHAQ | 14-3-3 protein theta | HIV [97] HSV1 [98] |
| YWHAZ | 14-3-3 protein zeta/delta | HSV1 [98] : Epstein-Barr [119] |
Kegg pathway analysis [33] of 632 schizophrenia susceptibility gene candidates was performed using Kegg mapper http://www.genome.jp/kegg/tool/color_pathway.html. The results of this analysis are available at http://www.polygenicpathways.co.uk/keggszgenes.htm. Venn diagrams were constructed online at http://www.bioinformatics.org/gvenn/index.htm [34].
Genes and risk factors with at least one positive association are included in this study. Although certain genes and risk factors are clearly more important than others, and problems of replication in both gene and risk factor studies abound, gene, gene, and gene/environment interactions may explain some of the heterogeneity. For example many schizophrenia-related genes are involved in the life cycle of T. Gondii, but may be irrelevant if this pathogen is not encountered. Similarly T. Gondii infection may have little effect is such gene variants are not present. Pathway analyses of genome wide association data, and previous studies, are showing that the risk-promoting effects of many genes in similar pathways are better predictors of risk, than when treating each gene in isolation (see Section 1). Although some of these factors may be false positives, many genes and risk factors may have a role to play in certain conditions, but the greater import of genes such as DISC1 or neuregulin is recognised.
3. Results
Pictograms of selected BLAST results are shown in Figure 1. The initial sweep of unfiltered BLASTs returned 125–304 hits, but this number was markedly increased when using the filter “schizophrenia” (14,088 Hits for HSV-2). For unfiltered sweeps, the viral homologues are longer, while the filtered sweeps return shorter contiguous sequences nevertheless including multiple matches of pentapeptides or more.
Figure 1.

Screenshots of the pictorial representation of the viral BLAST results against the human proteome. The streaks dotted throughout the human genome/proteome represent the areas of homology, some with contiguous sequences of 5 or more amino acids. The number of hits is shown for each virus or pathogen. The figure also shows the total coverage of human chromosome 10 by viral gene homologues. The top set of figures were from unfiltered blasts while the bottom set of 6 figures represent filtered blasts using the query “schizophrenia”.
Viral-human matches are characterised by short contiguous amino acid matches of 5 or more amino acids, that are identical in viral and human proteins, defined as vatches (viral matches). These are exemplified, for DISC1 in Figure 2. Hexapeptide matches have also been described for the influenza H5N1 virus and this study also highlighted homologies with DISC1, reelin and neurexin, inter alia [35]. The entire length of a human protein can be composed of many overlapping, intercalated vatches, related to multiple viral species. However, the viral spectrum is distinct for each protein as shown in Figure 3 for DISC1, neuregulin, the D2 dopamine receptor and transcription factor 4. Each is homologous to proteins from a large spectrum of viruses, but this spectrum is distinct for each protein. Interestingly, all are homologous to proteins from the hepatitis C virus. Several studies have noted that Hepatitis C infection is associated with schizophrenia, but this has generally been interpreted in terms of a schizophrenia life style that favours infection, rather than viewing Hepatitis C as a risk-promoting factor [36–39]. These data may challenge this assumption.
Figure 2.

(a) Varicella protein alignments within DISC1: the boxed regions show the region of alignment, and the blue letters denote 100% identity. This is not an alignment of the whole Varicella proteome but represents fragments of the same or different Varicella proteins that align with DISC1 fragments (vatches). The larger font delineates highly antigenic regions of DISC1 with an antigenicity index of >0.8 (Figure 4). (b) Other viral vatches within the DISC1 protein. The vatches are colour or format coded in relation to the different viruses. (c) Viral vatches for the risk factors implicated in schizophrenia in relation to the highly immunogenic regions of DISC1.
Figure 3.
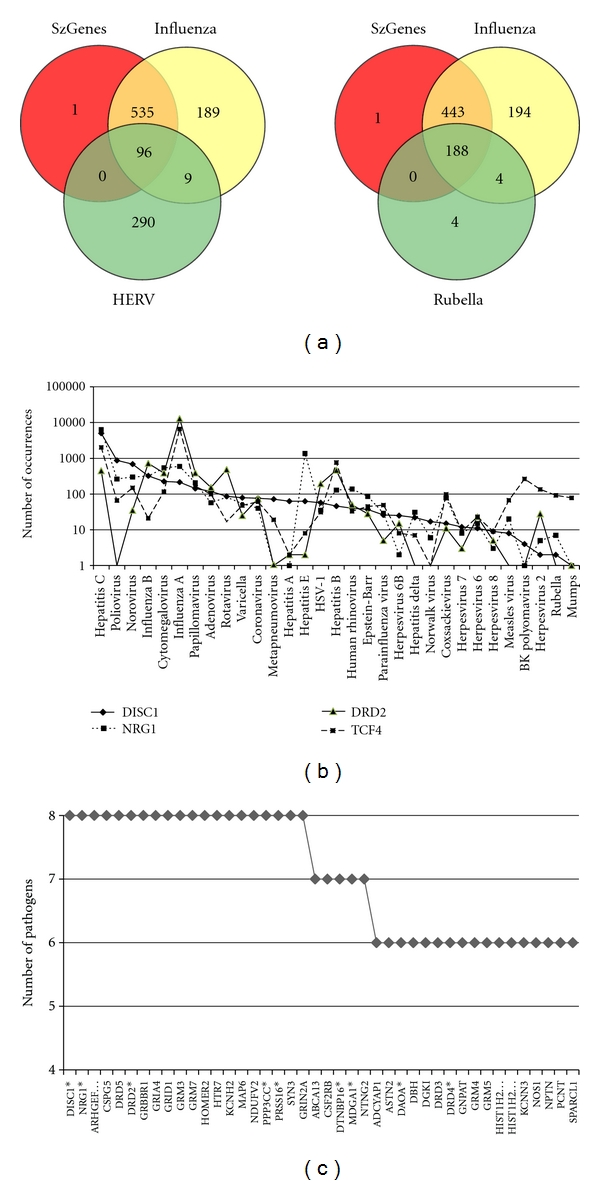
(a) Venn diagrams of the number of Schizophrenia gene products (N = 632) with homology to the rubella, HERV-W and influenza viruses. The singleton in SZ-genes was different on each occasion: Thus, all genes are covered. (b) The viral matching spectra of DISC1, neuregulin, the dopamine D2 receptor and transcription factor 4. The Y-axis depicts the number of word occurrences on the original BLAST results page. Note the logarithmic axis. (c) The number of pathogens expressing proteins with homology to the protein products of schizophrenia susceptibility genes. Those marked by an asterisk are within the 30 top-ranked genes in SZ-gene http://www.szgene.org/.
All of the pathogens implicated in schizophrenia express proteins with homology to multiple schizophrenia susceptibility gene products (Table 3). The profile of each individual pathogen is again specific for different types of gene product, but all target key members of the schizophrenia network including dopamine, serotonin and glutamate receptors as well as neuregulin and growth-related or DISC1 related pathways. This is the case even when no filter is used. Interestingly, both the rubella and the influenza viruses target members of the translation initiation complex, which has been implicated in myelination and oligodendrocyte survival [4, 40]. Oligodendrocyte cell loss and myelination defects are prominent in the schizophrenic brain [41–44].
Table 3.
Human proteins with homology to proteins expressed by pathogens. The size of the tags reflects the number of pathogen's proteins that are homologous to the human protein: the filters used are described. The number of schizophrenia susceptibility genes within each of these datasets is shown in the left-hand column. Certain genes are classified according to family and are highlighted in red. Gene definitions and the original BLAST files can be found at http://www.polygenicpathways.co.uk/blasts.htm. Note that the homologues are often clustered in families (e.g., HTR1A, HTR2A, HTR3A, HTR3B, HTR3E, HTR5A, and HTR7).
















|
The degree of overlap between the rubella, HERV and influenza viruses and schizophrenia gene products is shown by the Venn diagrams in Figure 3. All but one schizophrenia gene product was covered by various permutations and similar data were recovered for other pathogens. All schizophrenia gene products (N = 632) were homologous to proteins expressed by one or more of these pathogens. However, only 16 proteins were common to all 8 pathogens (Figure 3). These included neuregulin (NRG1) and DISC1, dopamine (DRD5), glutamate (GRIA4, GRID1, GRM3, GRM7) GABA (GABBR1) and serotonin (HTR7) receptors, a presynaptic protein regulating glutamate release (synapsin SYN3) and HOMER2, a member of the postsynaptic scaffold, all of which are key elements relating to the pathology of schizophrenia.
Other proteins within this class included neurocan (CSPG5), a chondroitin sulphate proteoglycan expressed in oligodendrocytes that inhibits neurite outgrowth and regulates axonal growth [45–47]. It is also involved thalamocortical projection development [48]. ARHGEF10 is a rho Guanine-nucleotide exchange factor that controls myelination [49]. NDUFV2 is a subunit of the mitochondrial respiratory chain and its protein expression levels are reduced in the frontal cortex and striatum in schizophrenia [50]. PPP3CC Calcineurin gamma (PPP3CC) plays a role in dopamine receptor signalling [51, 52]. Calcineurin knockout mice show defects in prepulse inhibition and other phenotypes related to schizophrenia [53]. Calcineurin is highly expressed in the immune system and regulates the expression of numerous cytokines [54]. MAP6 is a microtubule protein that controls synaptic organisation, in particular of glutamatergic synapses where it controls the expression of the glutamate transporter and presynaptic genes, synaptophysin and GAP-43, spinophilin and MAP2. [55, 56] KCNH2 is a potassium channel that plays a role in the development of neural crest cells [57] and in lymphocyte proliferation [58]. PRSS16 is a serine protease involved in autoimmunity and the presentation of self-antigens within the thymus [59].
So, by a random bioinformatics process, trawling the entire human proteome, asking simply which proteins are homologous to those of the pathogens implicated in schizophrenia, we arrive at a small set of proteins related to synaptic and dendritic function, myelination, neuregulin and DISC1 pathways, glutamate, dopamine, GABA and serotonin transmission, and immune regulation that are the cornerstones of schizophrenia pathology [3, 60–62].
3.1. Autoantigens in Schizophrenia
Many autoantibodies have been reported in schizophrenia. The pathogens implicated in schizophrenia also express proteins that are homologous to these autoantigens. Again the profile of each autoantigen or pathogen is distinct as shown in Table 2.
3.2. DISC1
DISC1 is a key “hub gene” in schizophrenia linked, via its interactome, to many other schizophrenia susceptibility gene products [3, 63–66]. Its viral homology is illustrated in Figure 2. The Varicella virus is homologous to DISC1 in several regions, over its entire length, many matches in regions of high immunogenicity. These figures illustrate the types of matches seen in other proteins and shows that the vatches are often part of larger gapped consensus sequences. Interestingly, Varicella infection also results in the production of antibodies to pericentrin, a DISC1 binding partner [67].
DISC1 is a highly immunogenic protein, as predicted by B-cell epitope prediction (Figure 4). Autoantibodies to DISC1 have not been reported in schizophrenia. However, the viral risk factors implicated in schizophrenia express proteins that are homologous to the highly antigenic regions of the DISC1 protein, as shown in Figure 2. These viral proteins are equally antigenic and antiviral antibodies might also thus be expected to target multiple regions of the DISC1 protein.
Figure 4.
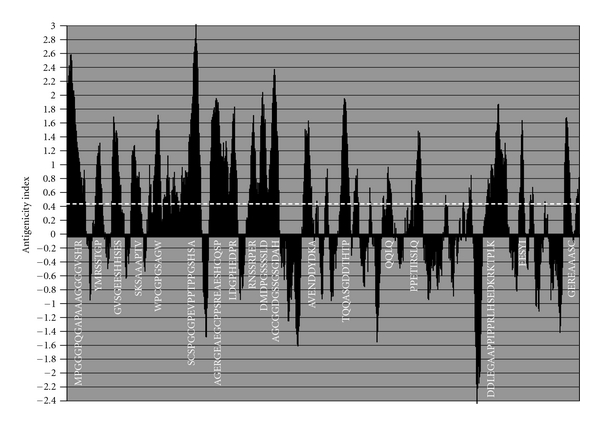
The antigenicity (B-cell epitope prediction) of DISC1: the amino acid sequences with an index of >0.35 are considered as epitopes. A value of 0.8 was chosen to define highly antigenic regions as seen in Figure 2. The amino acid sequences of these highly antigenic regions are shown.
3.3. Viral Proteins Are Part of the DISC1 Interactome
DISC1 and many of its binding partners, or other members of its interactome, contain vatches that are homologous to proteins expressed by the Rubella virus (Figure 5). (Other viruses also display this property, although the interactome members targeted are distinct, and specific for each virus (see http://www.polygenicpathways.co.uk/vatches.htm). Upon infection, viruses might therefore be considered as extraneous spurs to these types of protein/protein networks, and are likely to markedly affect their integrity. Indeed, several viruses, including herpes simplex, hepatitis C, Epstein-Barr, the cytomegalovirus, adenovirus and Coxsackie virus are known to bind to DISC1 interaction partners (Table 4).
Figure 5.
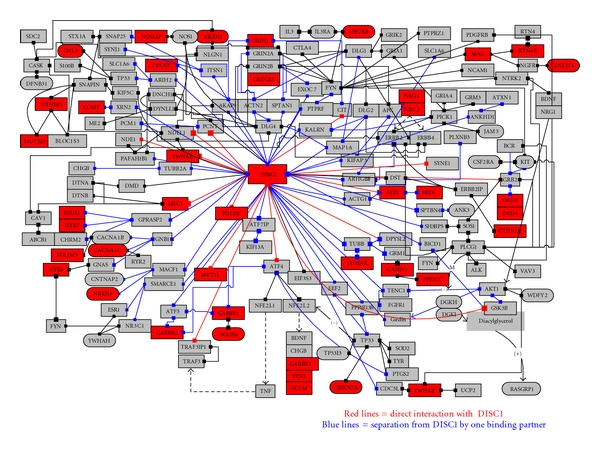
The DISC1 interactome see http://www.polygenicpathways.co.uk/discforum.htm. Proteins in red are homologous to Rubella proteins.
3.4. Viral DNA within the Human Genome
The insertion of viral DNA into the human genome had until recently been thought to be the preserve of retroviruses. However the incorporation of DNA into mammalian genomes has recently been demonstrated on a large scale for both RNA and DNA viruses. Viral integration may be mediated by nonhomologous recombination with chromosomal DNA or, in the case of RNA viruses, by interactions with host chromosomal retrotransposons [68, 69]. It has also been shown the herpes virus HHV-6 can be transmitted from parent to child via chromosomal integration [70]. The BLAST analyses of the viruses detailed in this paper, and of others at http://www.polygenicpathways.co.uk/blasts.htm clearly show that viral DNA from many species is present within the human genome. This viral homology may well cover the entire human genome. For example, a Blast of human chromosome 10 against all viral genomes (almost 3,000 viral forms) yielded 119,857 hits with entire coverage of 135.5 million bases. Viral DNA is thus both inter and intragenic (Figure 1). It has been proposed that retroviral integration, into paternal and maternal gene lines, inserting several genes at once and effectively creating a new being, is responsible for evolutionary saccades [71]. The fact that RNA and DNA nonretroviruses can also be so incorporated has important implications in this area.
The HSV-2 virus is homologous to several dopamine receptors and the BLAST pictogram shows how the same virus provokes repeating patterns in the human proteome (Figure 6). The same is true of the Herpes simplex virus (HSV-1) which is homologous to multiple lipoprotein receptors as well as to multiple kinases or of the cytomegalovirus which expresses proteins homologous to many chemokine receptors (see http://www.polygenicpathways.co.uk/blasts.htm). One interpretation of this, given the ability of chromosomal integration, is that repeated viral visits to the human genome over millions of years are responsible for the creation of gene families.
Figure 6.
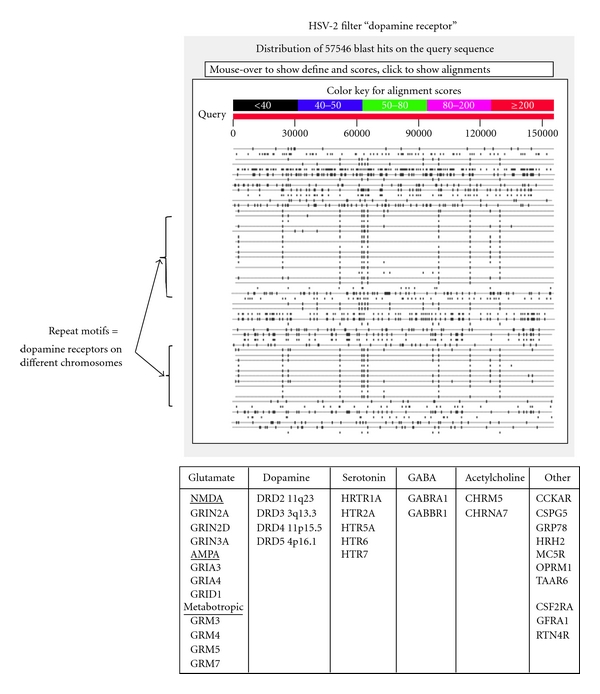
A screen shot of the HSV-2 BLAST results using the filter “dopamine receptor”. The repeated patterns correspond to dopamine receptors on different chromosomes as shown in Table 1. Homology with glutamate, serotonin, GABA, acetylcholine and other receptors is also noted.
It is also possible that viral/human homology reflects convergent viral evolution, although this is difficult to reconcile with the presence of viral DNA in intergenic regions, for which there would be little evolutionary drive or selective pressure. It is also plausible that a bidirectional transfer of human and viral DNA could be at work.
For whatever reason, the result is that human proteins resemble those expressed by a multitude of today's viruses and other pathogens. Upon infection, these pathogens are thus able to interfere with the function of their human counterparts in a number of ways (see below).
3.5. Copy Number Variations and the Effects of Parental Age on Risk
Repeated viral insertion could well explain copy number variations, which are associated with a number of diseases, including schizophrenia [72, 73]. As their number increases, so will the number of matches to the same viral proteins, thus increasing the risk of viral interference and autoimmunity. As viral infection can be passed from parent to child via chromosomal integration, perhaps this is also why both paternal and maternal older age have been reported as risk factors in schizophrenia and other disorders [74, 75].
3.6. KEGG Pathway Analysis of Schizophrenia Susceptibility Genes
The color-coded pathways for this analysis are posted at http://www.polygenicpathways.co.uk/keggszgenes.htm. It confirmed the involvement of a number of polygenic pathways, including long-term potentiation and oxidative stress [3] growth factor/neuregulin pathways [121], neuroactive ligand pathways (dopamine/serotonin/glutamate and others) as well as dopamine metabolism pathways [9]. In the context of this review, a large number of immune-related pathways are traced out by these genes, together with many pathogen-related pathways, including toxoplasmosis, which heads the list (Table 5). The involvement of schizophrenia related genes in the life cycles of pathogens has been the subject of a previous review [22] and this relationship is supported by this analysis. Other pathogen related pathways relating to amoebiasis, Staphylococcus aureus and Helicobacter pylori infection, might indicate the involvement of other pathogens in schizophrenia, although such pathways could also be considered as generic pathways related to many pathogens.
Table 5.
The number of schizophrenia gene products in KEGG pathways related to immunity, and viral or pathogen life cycles.
| Pathogen pathways | Viral pathways | Immune | |||
|---|---|---|---|---|---|
| Toxoplasmosis | 16 | Focal adhesion | 20 | Cytokine-cytokine receptor interaction | 26 |
| Chagas disease | 15 | Cell adhesion molecules (CAMs) | 19 | Jak-STAT signaling pathway | 16 |
| Amoebiasis | 13 | Regulation of actin cytoskeleton | 17 | Systemic lupus erythematosus | 13 |
| Leishmaniasis | 12 | Protein processing in endoplasmic reticulum | 13 | T cell receptor signaling pathway | 13 |
| Viral myocarditis | 8 | Endocytosis | 12 | Phagosome | 12 |
| Staphylococcus aureus infection | 7 | Phagosome | 12 | Allograft rejection | 11 |
| Epithelial cell signaling in Helicobacter pylori infection | 6 | Gap junction | 11 | Hematopoietic cell lineage | 11 |
| Malaria | 6 | Tight junction | 11 | Antigen processing and presentation | 10 |
| Tryptophan metabolism | 6 | Adherens junction | 6 | Fc epsilon RI signaling pathway | 10 |
| NOD-like receptor signaling pathway | 4 | ECM-receptor interaction | 6 | Apoptosis | 10 |
| Vibrio cholerae infection | 4 | Oocyte meiosis | 5 | Graft-versus-host disease | 9 |
| Bacterial invasion of epithelial cells | 3 | SNARE interactions in vesicular transport | 4 | Autoimmune thyroid disease | 8 |
| E.coli infection | 3 | Chemokine signaling pathway | 8 | ||
| RIG-I-like receptor signaling pathway | 3 | Basal transcription factors | 3 | Leukocyte transendothelial migration | 8 |
| Cytosolic DNA-sensing pathway | 2 | Spliceosome | 2 | Natural killer cell mediated cytotoxicity | 8 |
| Shigellosis | 2 | Aminoacyl-tRNA biosynthesis | 1 | Adipocytokine signaling pathway | 7 |
| Base excision repair | 1 | Asthma | 7 | ||
| RNA degradation | 1 | Intestinal IgA production | 5 | ||
| Toll-like receptor signaling pathway | 5 | ||||
| Complement and coagulation cascades | 4 | ||||
| B cell receptor signaling pathway | 3 | ||||
| TGF-beta signaling pathway | 3 | ||||
| Lysosome | 2 | ||||
| Regulation of autophagy | 2 | ||||
| Fc gamma R-mediated phagocytosis | 1 | ||||
| Primary immunodeficiency | 1 | ||||
There is no specific viral life cycle pathway within the KEGG dataset. However, viruses use adhesion molecules as receptors, endocytosis for cellular entry and the intracellular actin and tubulin networks for migration to and from the nucleus, mediated via dynein and kinesin motors. They also subjugate intracellular vesicular trafficking pathways, and are able to subvert both lysosomal and phagosomal pathways. Their exit may depend upon exocytosis, or by apoptotic or other means of killing their host cell [122]. These pathways are heavily represented within the schizophrenia gene analysis.
3.7. Mechanisms of Action
Individual proteins are homologous to multiple viral proteins, which nevertheless are specific for a spectrum of viruses, while individual viruses are homologous to a large but specific subset of human proteins.
Our proteomes therefore contain proteins with sequences exactly matching those in the current virome, and in the proteomes of bacteria and other pathogens, which are also subject to phage or viral infection. Pathogens' proteins are therefore homologous to receptors, transporters, peptide messengers, growth factors, and other protein products of diverse gene families. Upon infection, surrogate dopamine, NMDA serotonin and other receptors, as well as transporters and enzymes are made available, which in effect may steal the ligands of their human counterparts. It is already known that the dopaminergic ligand, amantadine, binds to the influenza virus [123], which expresses proteins homologous to dopamine receptors (Table 3). When homologous to peptide ligands, viral proteins may occupy and block or perhaps stimulate their cognate receptors, or use them for entry, as is the case with the AIDS virus and the CCR5 and CXCR4 chemokine receptors [124].
This is illustrated by the Norovirus (Norwalk) which causes vomiting sickness. The virus expresses proteins homologous to monoamine and other amine oxidases as well as to a number of dopamine and monoamine transporters (Table 6). Dopamine subversion by the viral homologues would be expected to increase dopamine levels resulting in emesis, thus explaining the recurrent vomiting produced by infection.
Table 6.
Human homologues of Norwalk virus proteins.
| Dopamine metabolisers | Amine transporters | Others |
|---|---|---|
|
AOC2 amine oxidases AOC3“” KDM1A amine oxidase demethylase KDM1B“” MAOA monoamine oxidase MAOB“” RNLS renalase amine oxidase SMOX spermine oxidase SPR sepiapterin reductase Monoamine synthesis cofactor SULT1A1 sulphotransferases SULT1A3 monoamine metabolite sulphation SULT1A4 |
SLC6A2 (Noradrenaline)
SLC6A3 (Dopamine) SLC18A1vesicular monoamine SLC18A2“” SLC22A2 organic cation SLC22A3 extraneuronal monoamine SLC29A4 (Na+/H+) |
CADPS2 amine release activator
CDCA7 cell division cycle associated 7 CDCA7L IL4I1 cytokine PICK1 postsynaptic scaffold |
The potential interference by viruses within protein/protein networks is well illustrated by the homology of rubella proteins to DISC1 and other members of its interactome, and by the fact that many viruses have indeed been found to bind to these components (Table 4).
The homologous human proteins of the viral risk factors implicated in schizophrenia correspond to the genomic locations of 632 schizophrenia susceptibility genes (see Venn diagrams). Both negative and positive genetic association results have been reported for these many genes and it now seems plausible that, in some cases, this may be due to the presence or absence of active infection with these and other pathogens, and that DNA assays have been detecting pathogen as well as human DNA in the blood samples used for assay. There is evidently no way of discriminating viral or bacterial double-stranded DNA from human DNA.
This is not specific to schizophrenia, as the viruses implicated in Alzheimer's disease (HSV-1, HIV-1, HHV-6 and the cytomegalovirus) [125–127] are also homologous to proteins encoded by Alzheimer's disease susceptibility genes see http://www.polygenicpathways.co.uk/blasts.htm [128].
It seems that a viable interpretation, given the same phenomenon in these diseases, is that these genes are susceptibility genes precisely because they encode for proteins with homology to the viral risk factors. Infection and genetics therefore appear to be interdependent. The pathogens may promote disease if the human genes encode for homologous products, and the genes promote disease if the homologous pathogen is encountered. Such interdependence likely explains the heterogeneous data in both gene and risk factor association studies.
Other pathogens, including Borrelia Burgdorferri and T. Gondii have also been implicated in schizophrenia. These too express many homologous proteins to both viral and human proteomes. These parasites tend to be associated with schizophrenia in adulthood, while viral infections are predominantly prenatal risk factors. These may have primed the antibody network to respond to homologous antigens expressed by Borrelia or T. Gondii, suggesting that detection and elimination of these pathogens may be of therapeutic benefit in adult life.
Schizophrenia is a neurodevelopmental disorder [129, 130] and, as the risk-promoting effects of viruses are related to maternal infection, it is possible that knockdown or interference of foetal proteins by viral-induced antibodies targeting their human counterparts may contribute to the neurodevelopmental disturbances observed in schizophrenia. Indeed DISC1, neuregulin, ERBB4, FEZ1 or COMT knockout mice display many of the pathological and behavioural symptoms associated with schizophrenia [131–135]. Viral interference with these same proteins might be expected to promote the same effects, but on a massive scale, targeting many relevant proteins at once. It is also possible that such autoantibodies play a role in the comorbid conditions associated with schizophrenia, for example autoimmune disease such as Thyrotoxicosis, celiac disease, acquired haemolytic anaemia, interstitial cystitis, or Sjogren's syndrome [136].
Autoantibodies to several proteins have been reported in schizophrenia (muscarinic, nicotinic, dopaminergic and NMDA receptors, inter alia, (Table 2) and all are homologous to proteins expressed by the risk factors in schizophrenia. The effects of antibody knockdown have not been analysed for any schizophrenia related proteins, but have been reported for the microtubule-related protein tau, in relation to Alzheimer's disease. In mice, tau immunisation produces tau hyperphosphorylation, neurofibrillary tangles and axonal damage as seen in the human condition [137]. Tau (MAPT) is homologous to Herpes simplex (HSV-1) and a number of other pathogens. Such effects are relevant to the autoantigens observed in schizophrenia.
Schizophrenia is also a degenerative disease in adolescence or adulthood, characterised by oligodendrocyte cell loss, impaired synaptic connectivity and pyramidal cell dendrite shrinkage [41, 138–140], In the light of the above homologies it seems likely that such degenerative changes may relate to autoimmune-related attack of these diverse compartments. Indeed there is evidence for microglial activation in the schizophrenic brain [141] and several studies have reported changes in the cytokine profile in the brain, CSF or peripheral immune compartments [24, 142–146].
3.8. Clinical Implications in Schizophrenia and Other Conditions
These data suggest that susceptibility gene products are the vehicles enabling the risk-promoting effects of pathogenic risk factors, via the interactions described above, and that the two are indispensable for the genesis of schizophrenia. Pathogen detection and elimination or vaccination, particularly prior to pregnancy might be expected to reduce the incidence of schizophrenia and also to be of clinical benefit in adulthood. Interestingly, vitamin D is able to stunt the growth of T. Gondii [147] and low levels of this vitamin, both prenatally and in adulthood, have been associated with schizophrenia risk, although abnormally high levels are also a risk factor [148]. Pharmaceutical effort in this direction may also vastly improve the armoury and safety of drugs against parasites such as T. Gondii and Borrelia.
Autoimmunity, involving several key schizophrenia-related proteins may well be a consequence of pathogen infection, and related to viral/human protein homology. Antigen and antibody removal by immunoadsorption techniques might therefore also be if clinical benefit.
This scenario suggests a novel and probably common class of “pathogenetic” autoimmune disease caused by pathogens but dependent on our genes. Indeed, the same phenomenon has been observed in Alzheimer's disease where the risk factor herpes simplex expresses proteins containing peptide matches to the products of multiple susceptibility genes [128]. Work from Kanduc's laboratory has also shown that 30 viral proteomes, including many nonretroviruses, contain multiple pentapeptide matches to many human proteins [149]. This is corroborated by data posted at http://www.polygenicpathways.co.uk/blasts.htm which shows, inter alia, that Bornavirus proteins, a virus implicated in Bipolar disorder [150], display this type of homology in relation to Bipolar disorder susceptibility gene products, that the coronavirus implicated in Parkinson's disease [151] expresses proteins homologous to the PARK7 gene product and to dopaminergic and oxidative stress-related proteins, and that multiple sclerosis autoantigens are homologous to the products of the Epstein-Barr virus which has been implicated in this disorder [152]. Our genomes and polymorphisms determine which vatches we possess, which pathogens match these sequences and which pathogen-related disorder we might develop. Environmental variables, and vaccination, determine which pathogens we encounter and our immune system (HLA-antigens and immune background determined soon after birth) may determine how we deal with these pathogens. With the power of current day bioinformatics, it should be possible to rapidly identify all vatches in the human proteome and to pair them with the various pathogenic species and human diseases. This would greatly aid our understanding of the implication of pathogens in disease and may lead to radically new therapies and prevention strategies in many disorders.
References
- 1.Crow TJ. How and why genetic linkage has not solved the problem of psychosis: review and hypothesis. American Journal of Psychiatry. 2007;164(1):13–21. doi: 10.1176/ajp.2007.164.1.13. [DOI] [PubMed] [Google Scholar]
- 2.Owen MJ, Craddock N, O’Donovan MC. Schizophrenia: genes at last? Trends in Genetics. 2005;21(9):518–525. doi: 10.1016/j.tig.2005.06.011. [DOI] [PubMed] [Google Scholar]
- 3.Carter CJ. Schizophrenia susceptibility genes converge on interlinked pathways related to glutamatergic transmission and long-term potentiation, oxidative stress and oligodendrocyte viability. Schizophrenia Research. 2006;86(1–3):1–14. doi: 10.1016/j.schres.2006.05.023. [DOI] [PubMed] [Google Scholar]
- 4.Carter CJ. eIF2B and oligodendrocyte survival: where nature and nurture meet in bipolar disorder and schizophrenia? Schizophrenia Bulletin. 2007;33(6):1343–1353. doi: 10.1093/schbul/sbm007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Khler AK, Djurovic S, Rimol LM, et al. Candidate gene analysis of the human natural killer-1 carbohydrate pathway and perineuronal nets in schizophrenia: B3GAT2 Is associated with disease risk and cortical surface area. Biological Psychiatry. 2011;69(1):90–96. doi: 10.1016/j.biopsych.2010.07.035. [DOI] [PubMed] [Google Scholar]
- 6.Jia P, Wang L, Meltzer HY, Zhao Z. Common variants conferring risk of schizophrenia: a pathway analysis of GWAS data. Schizophrenia Research. 2010;122(1–3):38–42. doi: 10.1016/j.schres.2010.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.O’Dushlaine C, Kenny E, Heron E, et al. Molecular pathways involved in neuronal cell adhesion and membrane scaffolding contribute to schizophrenia and bipolar disorder susceptibility. Molecular Psychiatry. 2010;16(3):286–292. doi: 10.1038/mp.2010.7. [DOI] [PubMed] [Google Scholar]
- 8.Nicodemus KK, Law AJ, Radulescu E, et al. Biological validation of increased schizophrenia risk with NRG1, ERBB4, and AKT1 epistasis via functional neuroimaging in healthy controls. Archives of General Psychiatry. 2010;67(10):991–1001. doi: 10.1001/archgenpsychiatry.2010.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Talkowski ME, Kirov G, Bamne M, et al. A network of dopaminergic gene variations implicated as risk factors for schizophrenia. Human Molecular Genetics. 2008;17(5):747–758. doi: 10.1093/hmg/ddm347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nicodemus KK, Callicott JH, Higier RG, et al. Evidence of statistical epistasis between DISC1, CIT and NDEL1 impacting risk for schizophrenia: biological validation with functional neuroimaging. Human Genetics. 2010;127(4):441–452. doi: 10.1007/s00439-009-0782-y. [DOI] [PubMed] [Google Scholar]
- 11.Brown AS, Cohen P, Greenwald S, Susser E. Nonaffective psychosis after prenatal exposure to rubella. American Journal of Psychiatry. 2000;157(3):438–443. doi: 10.1176/appi.ajp.157.3.438. [DOI] [PubMed] [Google Scholar]
- 12.Adams W, Kendell RE, Hare EH, Munk-Jorgensen P. Epidemiological evidence that maternal influenza contributes to the aetiology of schizophrenia. An analysis of Scottish, English, and Danish data. British Journal of Psychiatry. 1993;163:522–534. doi: 10.1192/bjp.163.4.522. [DOI] [PubMed] [Google Scholar]
- 13.Brown AS, Begg MD, Gravenstein S, et al. Serologic evidence of prenatal influenza in the etiology of schizophrenia. Archives of General Psychiatry. 2004;61(8):774–780. doi: 10.1001/archpsyc.61.8.774. [DOI] [PubMed] [Google Scholar]
- 14.Torrey EF, Rawlings R, Waldman IN. Schizophrenic births and viral diseases in two states. Schizophrenia Research. 1988;1(1):73–77. doi: 10.1016/0920-9964(88)90043-6. [DOI] [PubMed] [Google Scholar]
- 15.Mortensen PB, Pedersen CB, Hougaard DM, et al. A Danish National Birth Cohort study of maternal HSV-2 antibodies as a risk factor for schizophrenia in their offspring. Schizophrenia Research. 2010;(1–3):257–263. doi: 10.1016/j.schres.2010.06.010. [DOI] [PubMed] [Google Scholar]
- 16.Stober G, Franzek E, Beckmann H. Pregnancy infections in mothers of chronic schizophrenic patients. The significance of differential nosology [Schwangerschaftsinfektionen bei Muttern von chronisch Schizophrenen. Die Bedeutung einer differenzierten Nosologie] Nervenarzt. 1994;65(3):175–182. [PubMed] [Google Scholar]
- 17.Suvisaari J, Haukka J, Tanskanen A, Hovi T, Lönnqvist J. Association between prenatal exposure to poliovirus infection and adult schizophrenia. American Journal of Psychiatry. 1999;156(7):1100–1102. doi: 10.1176/ajp.156.7.1100. [DOI] [PubMed] [Google Scholar]
- 18.Rantakallio P, Jones P, Moring J, von Wendt L. Association between central nervous system infections during childhood and adult onset schizophrenia and other psychoses: a 28-year follow-up. International Journal of Epidemiology. 1997;26(4):837–843. doi: 10.1093/ije/26.4.837. [DOI] [PubMed] [Google Scholar]
- 19.Yolken RH, Torrey EF. Are some cases of psychosis caused by microbial agents? A review of the evidence. Molecular Psychiatry. 2008;13(5):470–479. doi: 10.1038/mp.2008.5. [DOI] [PubMed] [Google Scholar]
- 20.Fritzsche M. Seasonal correlation of sporadic schizophrenia to Ixodes ticks and Lyme borreliosis. International Journal of Health Geographics. 2002;1, article 2 doi: 10.1186/1476-072X-1-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Karlsson H, Bachmann S, Schröder J, McArthur J, Torrey EF, Yolken RH. Retroviral RNA identified in the cerebrospinal fluids and brains of individuals with schizophrenia. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(8):4634–4639. doi: 10.1073/pnas.061021998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Carter CJ. Schizophrenia susceptibility genes directly implicated in the life cycles of pathogens: cytomegalovirus, influenza, herpes simplex, rubella, and Toxoplasma gondii. Schizophrenia Bulletin. 2009;35(6):1163–1182. doi: 10.1093/schbul/sbn054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sun J, Jia P, Fanous AH, et al. Schizophrenia gene networks and pathways and their applications for novel candidate gene selection. PLoS One. 2010;5(6) doi: 10.1371/journal.pone.0011351. Article ID e11351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Müller N. The role of the cytokine network in the CNS and psychic disorders. Nervenarzt. 1997;68(1):11–20. doi: 10.1007/s001150050092. [DOI] [PubMed] [Google Scholar]
- 25.Rothermundt M, Arolt V, Bayer TA. Review of immunological and immunopathological findings in schizophrenia. Brain, Behavior, and Immunity. 2001;15(4):319–339. doi: 10.1006/brbi.2001.0648. [DOI] [PubMed] [Google Scholar]
- 26.Theodoropoulou S, Spanakos G, Baxevanis CN, et al. Cytokine serum levels, autologous mixed lymphocyte reaction and surface marker analysis in never medicated and chronically medicated schizophrenic patients. Schizophrenia Research. 2001;47(1):13–25. doi: 10.1016/s0920-9964(00)00007-4. [DOI] [PubMed] [Google Scholar]
- 27.Tanaka KF, Shintani F, Fujii Y, Yagi G, Asai M. Serum interleukin-18 levels are elevated in schizophrenia. Psychiatry Research. 2000;96(1):75–80. doi: 10.1016/s0165-1781(00)00196-7. [DOI] [PubMed] [Google Scholar]
- 28.O’Donnell MC, Catts SV, Ward PB, et al. Increased production of interleukin-2 (IL-2) but not soluble interleukin-2 receptors (sIL-2R) in unmedicated patients with schizophrenia and schizophreniform disorder. Psychiatry Research. 1996;65(3):171–178. doi: 10.1016/s0165-1781(96)02824-7. [DOI] [PubMed] [Google Scholar]
- 29.Ganguli R, Rabin BS, Kelly RH, Lyte M, Ragu U. Clinical and laboratory evidence of autoimmunity in acute schizophrenia. Annals of the New York Academy of Sciences. 1987;496:676–685. doi: 10.1111/j.1749-6632.1987.tb35829.x. [DOI] [PubMed] [Google Scholar]
- 30.Makinodan M, Tatsumi K, Manabe T, et al. Maternal immune activation in mice delays myelination and axonal development in the hippocampus of the offspring. Journal of Neuroscience Research. 2008;86(10):2190–2200. doi: 10.1002/jnr.21673. [DOI] [PubMed] [Google Scholar]
- 31.Altschul SF, Madden TL, Schäffer AA, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research. 1997;25(17):3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Larsen JE, Lund O, Nielsen M. Improved method for predicting linear B-cell epitopes. Immunome Research. 2006;2:p. 2. doi: 10.1186/1745-7580-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Goto S, Bono H, Ogata H, et al. Organizing and computing metabolic pathway data in terms of binary relations. Pacific Symposium on Biocomputing. 1997:175–186. [PubMed] [Google Scholar]
- 34.Pirooznia M, Nagarajan V, Deng Y. GeneVenn—a web application for comparing gene lists using Venn diagrams. Bioinformation. 2007;1:420–422. doi: 10.6026/97320630001420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kanduc D. Describing the hexapeptide identity platform between the influenza A H5N1 and Homo sapiens proteomes. Biologics. 2010;4:245–261. doi: 10.2147/btt.s12097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Himelhoch S, McCarthy JF, Ganoczy D, et al. Understanding associations between serious mental illness and hepatitis C virus among veterans: a national multivariate analysis. Psychosomatics. 2009;50(1):30–37. doi: 10.1176/appi.psy.50.1.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Baillargeon JG, Paar DP, Wu H, et al. Psychiatric disorders, HIV infection and HIV/hepatitis co-infection in the correctional setting. AIDS Care. 2008;20(1):124–129. doi: 10.1080/09540120701426532. [DOI] [PubMed] [Google Scholar]
- 38.Carney CP, Jones L, Woolson RF. Medical comorbidity in women and men with schizophrenia: a population-based controlled study. Journal of General Internal Medicine. 2006;21(11):1133–1137. doi: 10.1111/j.1525-1497.2006.00563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cournos F, McKinnon K, Sullivan G. Schizophrenia and comorbid human immunodeficiency virus or hepatitis C virus. Journal of Clinical Psychiatry. 2005;66(6):27–33. [PubMed] [Google Scholar]
- 40.van der Knaap MS, Pronk JC, Scheper GC. Vanishing white matter disease. The Lancet Neurology. 2006;5(5):413–423. doi: 10.1016/S1474-4422(06)70440-9. [DOI] [PubMed] [Google Scholar]
- 41.Uranova N, Orlovskaya D, Vikhreva O, et al. Electron microscopy of oligodendroglia in severe mental illness. Brain Research Bulletin. 2001;55(5):597–610. doi: 10.1016/s0361-9230(01)00528-7. [DOI] [PubMed] [Google Scholar]
- 42.Vostrikov V, Orlovskaya D, Uranova N. Deficit of pericapillary oligodendrocytes in the prefrontal cortex in schizophrenia. World Journal of Biological Psychiatry. 2008;9(1):34–42. doi: 10.1080/15622970701210247. [DOI] [PubMed] [Google Scholar]
- 43.Vostrikov VM. Decreased numerical density of pericapillary oligodendrocytes in the cortex in schizophrenia. Zhurnal Nevrologii i Psihiatrii imeni S.S. Korsakova. 2007;107(12):58–65. [PubMed] [Google Scholar]
- 44.Hakak Y, Walker JR, Li C, et al. Genome-wide expression analysis reveals dysregulation of myelination-related genes in chronic schizophrenia. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(8):4746–4751. doi: 10.1073/pnas.081071198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Friedlander DR, Milev P, Karthikeyan L, Margolis RK, Margolis RU, Grumet M. The neuronal chondroitin sulfate proteoglycan neurocan binds to the neural cell adhesion molecules Ng-CAM/L1/NILE and N-CAM, and inhibits neuronal adhesion and neurite outgrowth. Journal of Cell Biology. 1994;125(3):669–680. doi: 10.1083/jcb.125.3.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sandvig A, Berry M, Barrett LB, Butt A, Logan A. Myelin-, reactive glia-, and scar-derived CNS axon growth inhibitors: expression, receptor signaling, and correlation with axon regeneration. Glia. 2004;46(3):225–251. doi: 10.1002/glia.10315. [DOI] [PubMed] [Google Scholar]
- 47.Chen ZJ, Ughrin Y, Levine JM. Inhibition of axon growth by oligodendrocyte precursor cells. Molecular and Cellular Neuroscience. 2002;20(1):125–139. doi: 10.1006/mcne.2002.1102. [DOI] [PubMed] [Google Scholar]
- 48.Li HP, Oohira A, Ogawa M, Kawamura K, Kawano H. Aberrant trajectory of thalamocortical axons associated with abnormal localization of neurocan immunoreactivity in the cerebral neocortex of reeler mutant mice. European Journal of Neuroscience. 2005;22(11):2689–2696. doi: 10.1111/j.1460-9568.2005.04491.x. [DOI] [PubMed] [Google Scholar]
- 49.Verhoeven K, de Jonghe P, van de Putte T, et al. Slowed conduction and thin myelination of peripheral nerves associated with mutant rho Guanine-nucleotide exchange factor 10. American Journal of Human Genetics. 2003;73(4):926–932. doi: 10.1086/378159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ben-Shachar D, Karry R. Neuroanatomical pattern of mithochondrial complex I pathology varies between schizoprenia, bipolar disorder and major depression. PLoS One. 2008;3(11) doi: 10.1371/journal.pone.0003676. Article ID e3676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yuen EY, Yan Z. Dopamine D4 receptors regulate AMPA receptor trafficking and glutamatergic transmission in GABAergic interneurons of prefrontal cortex. Journal of Neuroscience. 2009;29(2):550–562. doi: 10.1523/JNEUROSCI.5050-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fukunaga K, Takeuchi Y. Novel intracellular signal transduction of dopamine D2 receptor in schizophrenia. Tanpakushitsu Kakusan Koso. 2006;51(11):1602–1608. [PubMed] [Google Scholar]
- 53.Miyakawa T, Leiter LM, Gerber DJ, et al. Conditional calcineurin knockout mice exhibit multiple abnormal behaviors related to schizophrenia. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(15):8987–8992. doi: 10.1073/pnas.1432926100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Viola JPB, Rao A. Role of the cyclosporin-sensitive transcription factor NFAT1 in the allergic response. Memorias do Instituto Oswaldo Cruz. 1997;92(supplement 2):147–155. doi: 10.1590/s0074-02761997000800020. [DOI] [PubMed] [Google Scholar]
- 55.Andrieux A, Salin PA, Vernet M, et al. The suppression of brain cold-stable microtubules in mice induces synaptic defects associated with neuroleptic-sensitive behavioral disorders. Genes and Development. 2002;16(18):2350–2364. doi: 10.1101/gad.223302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Andrieux A, Salin PA, Job D. A role for the cytoskeleton in mental diseases? [Un rôle pour les microtubules dans les pathologies psychiatriques?] Pathologie Biologie. 2004;52(2):89–92. doi: 10.1016/j.patbio.2003.04.007. [DOI] [PubMed] [Google Scholar]
- 57.Arcangeli A, Rosati B, Cherubini A, et al. HERG- and IRK-like inward rectifier currents are sequentially expressed during neuronal development of neural crest cells and their derivatives. European Journal of Neuroscience. 1997;9(12):2596–2604. doi: 10.1111/j.1460-9568.1997.tb01689.x. [DOI] [PubMed] [Google Scholar]
- 58.Smith GAM, Tsui HW, Newell EW, et al. Functional up-regulation of HERG K channels in neoplastic hematopoietic cells. Journal of Biological Chemistry. 2002;277(21):18528–18534. doi: 10.1074/jbc.M200592200. [DOI] [PubMed] [Google Scholar]
- 59.Gommeaux J, Grégoire C, Nguessan P, et al. Thymus-specific serine protease regulates positive selection of a subset of CD4+ thymocytes. European Journal of Immunology. 2009;39(4):956–964. doi: 10.1002/eji.200839175. [DOI] [PubMed] [Google Scholar]
- 60.Buonanno A, Kwon OB, Yan L, et al. Neuregulins and neuronal plasticity: possible relevance in schizophrenia. Novartis Foundation Symposium. 2008;289:165–177. doi: 10.1002/9780470751251.ch13. [DOI] [PubMed] [Google Scholar]
- 61.Seeman P. Glutamate and dopamine components in schizophrenia. Journal of Psychiatry and Neuroscience. 2009;34(2):143–149. [PMC free article] [PubMed] [Google Scholar]
- 62.Uranova NA, Vostrikov VM, Orlovskaya DD, Rachmanova VI. Oligodendroglial density in the prefrontal cortex in schizophrenia and mood disorders: a study from the Stanley Neuropathology Consortium. Schizophrenia Research. 2004;67(2-3):269–275. doi: 10.1016/S0920-9964(03)00181-6. [DOI] [PubMed] [Google Scholar]
- 63.Winter C, Reutiman TJ, Folsom TD, et al. Dopamine and serotonin levels following prenatal viral infection in mouse-Implications for psychiatric disorders such as schizophrenia and autism. European Neuropsychopharmacology. 2008;18(10):712–716. doi: 10.1016/j.euroneuro.2008.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Millar JK, Wilson-Annan JC, Anderson S, et al. Disruption of two novel genes by a translocation co-segregating with schizophrenia. Human Molecular Genetics. 2000;9(9):1415–1423. doi: 10.1093/hmg/9.9.1415. [DOI] [PubMed] [Google Scholar]
- 65.Camargo LM, Wang Q, Brandon NJ. What can we learn from the disrupted in schizophrenia 1 interactome: lessons for target identification and disease biology? Novartis Foundation Symposium. 2008;289:208–216. doi: 10.1002/9780470751251.ch17. [DOI] [PubMed] [Google Scholar]
- 66.Hennah W, Porteous D. The DISC1 pathway modulates expression of neurodevelopmental, synaptogenic and sensory perception genes. PLoS One. 2009;4(3) doi: 10.1371/journal.pone.0004906. Article ID e4906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rossner S, Ueberham U, Schlichs R, Perez-Polo JR, Bigl V. Neurotrophin binding to the p75 neurotrophin receptor is necessary but not sufficient to mediate NGF-effects on APP secretion in PC-12 cells. Journal of Neural Transmission, Supplement. 1998;(54):279–285. doi: 10.1007/978-3-7091-7508-8_27. [DOI] [PubMed] [Google Scholar]
- 68.Katzourakis A, Gifford RJ. Endogenous viral elements in animal genomes. PLoS Genetics. 2010;6(11) doi: 10.1371/journal.pgen.1001191. Article ID e1001191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Geuking MB, Weber J, Dewannieux M, et al. Recombination of retrotransposon and exogenous RNA virus results in nonretroviral cDNA integration. Science. 2009;323(5912):393–396. doi: 10.1126/science.1167375. [DOI] [PubMed] [Google Scholar]
- 70.Tanaka-Taya K, Sashihara J, Kurahashi H, et al. Human herpesvirus 6 (HHV-6) is transmitted from parent to child in an integrated form and characterization of cases with chromosomally integrated HHV-6 DNA. Journal of Medical Virology. 2004;73(3):465–473. doi: 10.1002/jmv.20113. [DOI] [PubMed] [Google Scholar]
- 71.Khodosevich K, Lebedev Y, Sverdlov E. Endogenous retroviruses and human evolution. Comparative and Functional Genomics. 2002;3(6):494–498. doi: 10.1002/cfg.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Brown AS. Prenatal infection as a risk factor for schizophrenia. Schizophrenia Bulletin. 2006;32(2):200–202. doi: 10.1093/schbul/sbj052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ozawa K, Hashimoto K, Kishimoto T, Shimizu E, Ishikura H, Iyo M. Immune activation during pregnancy in mice leads to dopaminergic hyperfunction and cognitive impairment in the offspring: a neurodevelopmental animal model of schizophrenia. Biological Psychiatry. 2006;59(6):546–554. doi: 10.1016/j.biopsych.2005.07.031. [DOI] [PubMed] [Google Scholar]
- 74.Kinnell HG. Parental age in schizophrenia. British Journal of Psychiatry. 1983;142:p. 204. doi: 10.1192/bjp.142.2.204a. [DOI] [PubMed] [Google Scholar]
- 75.Lopez-Castroman J, Gómez DD, Belloso JJC, et al. Differences in maternal and paternal age between Schizophrenia and other psychiatric disorders. Schizophrenia Research. 2010;116(2-3):184–190. doi: 10.1016/j.schres.2009.11.006. [DOI] [PubMed] [Google Scholar]
- 76.Brown AS, Cohen P, Greenwald S, Susser E. Nonaffective psychosis after prenatal exposure to rubella. American Journal of Psychiatry. 2000;157(3):438–443. doi: 10.1176/appi.ajp.157.3.438. [DOI] [PubMed] [Google Scholar]
- 77.Dalman C, Allebeck P, Gunnell D, et al. Infections in the CNS during childhood and the risk of subsequent psychotic illness: a cohort study of more than one million Swedish subjects. American Journal of Psychiatry. 2008;165(1):59–65. doi: 10.1176/appi.ajp.2007.07050740. [DOI] [PubMed] [Google Scholar]
- 78.Prasad KM, Bamne MN, Shirts BH, et al. Grey matter changes associated with host genetic variation and exposure to Herpes Simplex Virus 1 (HSV1) in first episode schizophrenia. Schizophrenia Research. 2010;118(1–3):232–239. doi: 10.1016/j.schres.2010.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Niebuhr DW, Millikan AM, Yolken R, Li Y, Weber NS. Results from a hypothesis generating case-control study: herpes family viruses and schizophrenia among military personnel. Schizophrenia Bulletin. 2008;34(6):1182–1188. doi: 10.1093/schbul/sbm139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Abrahao AL, Focaccia R, Gattaz WF. Childhood meningitis increases the risk for adult schizophrenia. World Journal of Biological Psychiatry. 2005;6(2):44–48. doi: 10.1080/15622970510030063. [DOI] [PubMed] [Google Scholar]
- 81.Terayama H, Nishino Y, Kishi M, Ikuta K, Itoh M, Iwahashi K. Detection of anti-Borna Disease Virus (BDV) antibodies from patients with schizophrenia and mood disorders in Japan. Psychiatry Research. 2003;120(2):201–206. doi: 10.1016/s0165-1781(03)00190-2. [DOI] [PubMed] [Google Scholar]
- 82.Buka SL, Cannon TD, Torrey EF, Yolken RH. Maternal exposure to herpes simplex virus and risk of psychosis among adult offspring. Biological Psychiatry. 2008;63(8):809–815. doi: 10.1016/j.biopsych.2007.09.022. [DOI] [PubMed] [Google Scholar]
- 83.Severance EG, Dickerson FB, Viscidi RP, et al. Coronavirus immunoreactivity in individuals with a recent onset of psychotic symptoms. Schizophrenia Bulletin. 2011;37(1):101–107. doi: 10.1093/schbul/sbp052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ellman LM, Yolken RH, Buka SL, Torrey EF, Cannon TD. Cognitive functioning prior to the onset of psychosis: the role of fetal exposure to serologically determined influenza infection. Biological Psychiatry. 2009;65(12):1040–1047. doi: 10.1016/j.biopsych.2008.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yao Y, Schröder J, Nellåker C, et al. Elevated levels of human endogenous retrovirus-W transcripts in blood cells from patients with first episode schizophrenia. Genes, Brain and Behavior. 2008;7(1):103–112. doi: 10.1111/j.1601-183X.2007.00334.x. [DOI] [PubMed] [Google Scholar]
- 86.Brown AS, Schaefer CA, Quesenberry CP, Liu L, Babulas VP, Susser ES. Maternal exposure to toxoplasmosis and risk of schizophrenia in adult offspring. American Journal of Psychiatry. 2005;162(4):767–773. doi: 10.1176/appi.ajp.162.4.767. [DOI] [PubMed] [Google Scholar]
- 87.Dickerson F, Stallings C, Origoni A, Copp C, Khushalani S, Yolken R. Antibodies to measles in individuals with recent onset psychosis. Schizophrenia Research. 2010;119(1–3):89–94. doi: 10.1016/j.schres.2009.12.010. [DOI] [PubMed] [Google Scholar]
- 88.Amminger GP, McGorry PD, Berger GE, et al. Antibodies to infectious agents in individuals at ultra-high risk for psychosis. Biological Psychiatry. 2007;61(10):1215–1217. doi: 10.1016/j.biopsych.2006.09.034. [DOI] [PubMed] [Google Scholar]
- 89.Chandley MJ, Miller MN, Kwasigroch CN, Wilson TD, Miller BE. Increased antibodies for the α7 subunit of the nicotinic receptor in schizophrenia. Schizophrenia Research. 2009;109(1–3):98–101. doi: 10.1016/j.schres.2009.01.023. [DOI] [PubMed] [Google Scholar]
- 90.Borda T, Gomez R, Berría MI, Sterin-Borda L. Antibodies against astrocyte M1 and M2 muscarinic cholinoceptor from schizophrenic patients' sera. Glia. 2004;45(2):144–154. doi: 10.1002/glia.10312. [DOI] [PubMed] [Google Scholar]
- 91.Suzuki Y, Kurita T, Sakurai K, Takeda Y, Koyama T. Case report of anti-NMDA receptor encephalitis suspected of schizophrenia. Seishin Shinkeigaku Zasshi. 2009;111(12):1479–1484. [PubMed] [Google Scholar]
- 92.Shcherbakova IV, Siryachenko TM, Mazaeva NA, Kaleda VG, Krasnolobova SA, Klyushnik TP. Leukocyte elastase and autoantibodies to nerve growth factor in the acute phase of schizophrenia and their relationship to symptomatology. World Journal of Biological Psychiatry. 2004;5(3):143–148. doi: 10.1080/15622970410029926. [DOI] [PubMed] [Google Scholar]
- 93.Tanaka S, Matsunaga H, Kimura M, et al. Autoantibodies against four kinds of neurotransmitter receptors in psychiatric disorders. Journal of Neuroimmunology. 2003;141(1-2):155–164. doi: 10.1016/s0165-5728(03)00252-2. [DOI] [PubMed] [Google Scholar]
- 94.Wang XF, Wang D, Zhu W, Delrahim KK, Dolnak D, Rapaport MH. Studies characterizing 60 kDa autoantibodies in subjects with schizophrenia. Biological Psychiatry. 2003;53(5):361–375. doi: 10.1016/s0006-3223(02)01972-8. [DOI] [PubMed] [Google Scholar]
- 95.Kim JJ, Lee SJ, Toh KY, Lee CU, Lee C, Paik IH. Identification of antibodies to heat shock proteins 90 kDa and 70 kDa in patients with schizophrenia. Schizophrenia Research. 2001;52(1-2):127–135. doi: 10.1016/s0920-9964(00)00091-8. [DOI] [PubMed] [Google Scholar]
- 96.Poletaev AB, Morozov SG, Gnedenko BB, Zlunikin VM, Korzhenevskey DA. Serum anti-S100b, anti-GFAP and anti-NGF autoantibodies of IgG class in healthy persons and patients with mental and neurological disorders. Autoimmunity. 2000;32(1):33–38. doi: 10.3109/08916930008995985. [DOI] [PubMed] [Google Scholar]
- 97.Chatr-Aryamontri A, Ceol A, Peluso D, et al. VirusMINT: a viral protein interaction database. Nucleic Acids Research. 2009;37(1):D669–D673. doi: 10.1093/nar/gkn739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Carter CJ. Herpes simplex: host viral protein interactions. WikiGenes, 2010.
- 99.Lan S, Wang H, Jiang H, et al. Direct interaction between α-actinin and hepatitis C virus NS5B. FEBS Letters. 2003;554(3):289–294. doi: 10.1016/s0014-5793(03)01163-3. [DOI] [PubMed] [Google Scholar]
- 100.Forgacs E, Gupta SK, Kerry JA, Semmes OJ. The bZIP transcription factor ATFx binds human T-cell leukemia virus type 1 (HTLV-1) tax and represses HTLV-1 long terminal repeat-mediated transcription. Journal of Virology. 2005;79(11):6932–6939. doi: 10.1128/JVI.79.11.6932-6939.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Indran SV, Ballestas ME, Britt WJ. Bicaudal D1-dependent trafficking of human cytomegalovirus tegument protein pp150 in virus-infected cells. Journal of Virology. 2010;84(7):3162–3177. doi: 10.1128/JVI.01776-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Huang YP, Cheng J, Zhang SL, et al. Screening of hepatocyte proteins binding to F protein of hepatitis C virus by yeast two-hybrid system. World Journal of Gastroenterology. 2005;11(36):5659–5665. doi: 10.3748/wjg.v11.i36.5659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Radtke K, Kieneke D, Wolfstein A, et al. Plus- and minus-end directed microtubule motors bind simultaneously to herpes simplex virus capsids using different inner tegument structures. PLoS Pathogens. 2010;6(7) doi: 10.1371/journal.ppat.1000991. Article ID e1000991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Bremner KH, Scherer J, Yi J, Vershinin M, Gross SP, Vallee RB. Adenovirus transport via direct interaction of cytoplasmic dynein with the viral capsid hexon subunit. Cell Host and Microbe. 2010;6(6):523–535. doi: 10.1016/j.chom.2009.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Si H, Verma SC, Lampson MA, Cai Q, Robertson ES. Kaposi’s sarcoma-associated herpesvirus-encoded LANA can interact with the nuclear mitotic apparatus protein to regulate genome maintenance and segregation. Journal of Virology. 2008;82(13):6734–6746. doi: 10.1128/JVI.00342-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ramanathan HN, Chung DH, Plane SJ, et al. Dynein-dependent transport of the Hantaan virus nucleocapsid protein to the endoplasmic reticulum-Golgi intermediate compartment. Journal of Virology. 2007;81(16):8634–8647. doi: 10.1128/JVI.00418-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Schramm B, de Haan CAM, Young J, et al. Vaccinia-virus-induced cellular contractility facilitates the subcellular localization of the viral replication sites. Traffic. 2006;7(10):1352–1367. doi: 10.1111/j.1600-0854.2006.00470.x. [DOI] [PubMed] [Google Scholar]
- 108.Dorweiler IJ, Ruone SJ, Wang H, Burry RW, Mansky LM. Role of the human T-cell leukemia virus type 1 PTAP motif in Gag targeting and particle release. Journal of Virology. 2006;80(7):3634–3643. doi: 10.1128/JVI.80.7.3634-3643.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kobayashi K, Sueyoshi T, Inoue K, Moore R, Negishi M. Cytoplasmic accumulation of the nuclear receptor CAR by a tetratricopeptide repeat protein in HepG2 cells. Molecular Pharmacology. 2003;64(5):1069–1075. doi: 10.1124/mol.64.5.1069. [DOI] [PubMed] [Google Scholar]
- 110.Bremner KH, Scherer J, Yi J, Vershinin M, Gross SP, Vallee RB. Adenovirus transport via direct interaction of cytoplasmic dynein with the viral capsid hexon subunit. Cell Host and Microbe. 2010;6(6):523–535. doi: 10.1016/j.chom.2009.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Johannsen E, Luftig M, Chase MR, et al. Proteins of purified Epstein-Barr virus. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(46):16286–16291. doi: 10.1073/pnas.0407320101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Collier AJ, Gallego J, Klinck R, et al. A conserved RNA structure within the HCV IRES eIF3-binding site. Nature Structural Biology. 2002;9(5):375–380. doi: 10.1038/nsb785. [DOI] [PubMed] [Google Scholar]
- 113.Sawa H, Suzuki T, Orba Y, Sunden Y, Nagashima K. Recent research on the JC virus. Brain and Nerve. 2007;59(2):101–108. [PubMed] [Google Scholar]
- 114.Vos RM, Altreuter J, White EA, Howley PM. The ubiquitin-specific peptidase USP15 regulates human papillomavirus type 16 E6 protein stability. Journal of Virology. 2009;83(17):8885–8892. doi: 10.1128/JVI.00605-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.König R, Zhou Y, Elleder D, et al. Global analysis of host-pathogen interactions that regulate early-stage HIV-1 replication. Cell. 2008;135(1):49–60. doi: 10.1016/j.cell.2008.07.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kondratova AA, Neznanov N, Kondratov RV, Gudkov AV. Poliovirus protein 3A binds and inactivates LIS1, causing block of membrane protein trafficking and deregulation of cell division. Cell Cycle. 2005;4(10):1403–1410. doi: 10.4161/cc.4.10.2041. [DOI] [PubMed] [Google Scholar]
- 117.Epie N, Ammosova T, Sapir T, et al. HIV-1 Tat interacts with LIS1 protein. Retrovirology. 2005;2 doi: 10.1186/1742-4690-2-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Verdier Y, Penke B. Binding sites of amyloid β-peptide in cell plasma membrane and implications for Alzheimer's disease. Current Protein and Peptide Science. 2004;5(1):19–31. doi: 10.2174/1389203043486937. [DOI] [PubMed] [Google Scholar]
- 119.Bossis I, Roden RBS, Gambhira R, et al. Interaction of tSNARE syntaxin 18 with the papillomavirus minor capsid protein mediates infection. Journal of Virology. 2005;79(11):6723–6731. doi: 10.1128/JVI.79.11.6723-6731.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Brunetti CR, Dingwell KS, Wale C, Graham FL, Johnson DC. Herpes simplex virus gD and virions accumulate in endosomes by mannose 6-phosphate-dependent and -independent mechanisms. Journal of Virology. 1998;72(4):3330–3339. doi: 10.1128/jvi.72.4.3330-3339.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Balu DT, Coyle JT. Neuroplasticity signaling pathways linked to the pathophysiology of schizophrenia. Neuroscience and Biobehavioral Reviews. 2011;35(3):848–870. doi: 10.1016/j.neubiorev.2010.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Flint SJ, Enquist LW, Racaniello VR, Skalka AM. Principles of Virology. Herndon, Va, USA: ASM Press; 2008. [Google Scholar]
- 123.Rosenberg MR, Casarotto MG. Coexistence of two adamantane binding sites in the influenza A M2 ion channel. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(31):13866–13871. doi: 10.1073/pnas.1002051107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Toma J, Whitcomb JM, Petropoulos CJ, Huang W. Dual-tropic HIV type 1 isolates vary dramatically in their utilization of CCR5 and CXCR4 coreceptors. AIDS. 2010;24(14):2181–2186. doi: 10.1097/QAD.0b013e32833c543f. [DOI] [PubMed] [Google Scholar]
- 125.Carter CJ. Interactions between the products of the Herpes simplex genome and Alzheimer’s disease susceptibility genes: relevance to pathological-signalling cascades. Neurochemistry International. 2008;52(6):920–934. doi: 10.1016/j.neuint.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 126.Honjo K, van Reekum R, Verhoeff NPLG. Alzheimer’s disease and infection: do infectious agents contribute to progression of Alzheimer’s disease? Alzheimer’s and Dementia. 2009;5(4):348–360. doi: 10.1016/j.jalz.2008.12.001. [DOI] [PubMed] [Google Scholar]
- 127.Itzhaki RF, Wozniak MA, Appelt DM, Balin BJ. Infiltration of the brain by pathogens causes Alzheimer’s disease. Neurobiology of Aging. 2004;25(5):619–627. doi: 10.1016/j.neurobiolaging.2003.12.021. [DOI] [PubMed] [Google Scholar]
- 128.Carter CJ. Alzheimer's disease: a pathogenetic autoimmune disorder caused by herpes simplex in a gene-dependent manner. International Journal of Alzheimer's Disease. 2010;2010:17 pages. doi: 10.4061/2010/140539. Article ID 140539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Chance SA, Esiri MM, Crow TJ. Ventricular enlargement in schizophrenia: a primary change in the temporal lobe? Schizophrenia Research. 2003;62(1-2):123–131. doi: 10.1016/s0920-9964(02)00344-4. [DOI] [PubMed] [Google Scholar]
- 130.Hennah W, Porteous D. The DISC1 pathway modulates expression of neurodevelopmental, synaptogenic and sensory perception genes. PLoS One. 2009;4(3) doi: 10.1371/journal.pone.0004906. Article ID e4906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Desbonnet L, Waddington JL, O’Tuathaigh CMP. Mutant models for genes associated with schizophrenia. Biochemical Society Transactions. 2009;37(1):308–312. doi: 10.1042/BST0370308. [DOI] [PubMed] [Google Scholar]
- 132.Jaaro-Peled H. Gene models of schizophrenia: DISC1 mouse models. Progress in Brain Research. 2009;179:75–86. doi: 10.1016/S0079-6123(09)17909-8. [DOI] [PubMed] [Google Scholar]
- 133.Sakae N, Yamasaki N, Kitaichi K, et al. Mice lacking the schizophrenia-associated protein FEZ1 manifest hyperactivity and enhanced responsiveness to psychostimulants. Human Molecular Genetics. 2008;17(20):3191–3203. doi: 10.1093/hmg/ddn215. [DOI] [PubMed] [Google Scholar]
- 134.Barros CS, Calabrese B, Chamero P, et al. Impaired maturation of dendritic spines without disorganization of cortical cell layers in mice lacking NRG1/ErbB signaling in the central nervous system. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(11):4507–4512. doi: 10.1073/pnas.0900355106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Babovic D, O’Tuathaigh CM, O’Connor AM, et al. Phenotypic characterization of cognition and social behavior in mice with heterozygous versus homozygous deletion of catechol-O-methyltransferase. Neuroscience. 2008;155(4):1021–1029. doi: 10.1016/j.neuroscience.2008.07.006. [DOI] [PubMed] [Google Scholar]
- 136.Eaton WW, Byrne M, Ewald H, et al. Association of schizophrenia and autoimmune diseases: linkage of Danish national registers. American Journal of Psychiatry. 2006;163(3):521–528. doi: 10.1176/appi.ajp.163.3.521. [DOI] [PubMed] [Google Scholar]
- 137.Rosenmann H, Grigoriadis N, Karussis D, et al. Tauopathy-like abnormalities and neurologic deficits in mice immunized with neuronal tau protein. Archives of Neurology. 2006;63(10):1459–1467. doi: 10.1001/archneur.63.10.1459. [DOI] [PubMed] [Google Scholar]
- 138.Black JE, Kodish IM, Grossman AW, et al. Pathology of layer V pyramidal neurons in the prefrontal cortex of patients with schizophrenia. American Journal of Psychiatry. 2004;161(4):742–744. doi: 10.1176/appi.ajp.161.4.742. [DOI] [PubMed] [Google Scholar]
- 139.Kolomeets NS, Uranova NA. Synaptic contacts in schizophrenia: the study with immunocytochemical identification of dopaminergic neurons [Sinapticheskie kontakty pri shizofrenii: issledovaniie s immunotsitokhimicheskoi identifikatsiei dofaminergicheskikh neironov] Zhurnal Nevropatolgii i Psikhiatrii im. S S Korsakova. 1997;97(12):39–43. [PubMed] [Google Scholar]
- 140.Thompson PM, Vidal C, Giedd JN, et al. Mapping adolescent brain change reveals dynamic wave of accelerated gray matter loss in very early-onset schizophrenia. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(20):11650–11655. doi: 10.1073/pnas.201243998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bayer TA, Buslei R, Havas L, Falkai P. Evidence for activation of microglia in patients with psychiatric illnesses. Neuroscience Letters. 1999;271(2):126–128. doi: 10.1016/s0304-3940(99)00545-5. [DOI] [PubMed] [Google Scholar]
- 142.Holden RJ, Pakula IS. Immunological influences in attention-deficit disorder and schizophrenia: is there a link between these two conditions? Medical Hypotheses. 1995;45(6):575–587. doi: 10.1016/0306-9877(95)90242-2. [DOI] [PubMed] [Google Scholar]
- 143.Söderlund J, Schröder J, Nordin C, et al. Activation of brain interleukin-1β in schizophrenia. Molecular Psychiatry. 2009;14(12):1069–1071. doi: 10.1038/mp.2009.52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Drzyzga Ł, Obuchowicz E, Marcinowska A, Herman ZS. Cytokines in schizophrenia and the effects of antipsychotic drugs. Brain, Behavior, and Immunity. 2006;20(6):532–545. doi: 10.1016/j.bbi.2006.02.002. [DOI] [PubMed] [Google Scholar]
- 145.Smith RS, Maes M. The macrophage-T-lymphocyte theory of schizophrenia: additional evidence. Medical Hypotheses. 1995;45(2):135–141. doi: 10.1016/0306-9877(95)90062-4. [DOI] [PubMed] [Google Scholar]
- 146.Vawter MP, Dillon-Carter O, Issa F, Wyatt RJ, Freed WJ. Transforming growth factors β1 and β2 in the cerebrospinal fluid of chronic schizophrenic patients. Neuropsychopharmacology. 1997;16(1):83–87. doi: 10.1016/S0893-133X(96)00143-1. [DOI] [PubMed] [Google Scholar]
- 147.Rajapakse R, Uring-Lambert B, Andarawewa KL, et al. 1,25(OH)2D3 inhibits in vitro and in vivo intracellular growth of apicomplexan parasite Toxoplasma gondii . Journal of Steroid Biochemistry and Molecular Biology. 2007;103(3-5):811–814. doi: 10.1016/j.jsbmb.2006.12.058. [DOI] [PubMed] [Google Scholar]
- 148.McGrath JJ, Eyles DW, Pedersen CB, et al. Neonatal vitamin D status and risk of schizophrenia: a population-based case-control study. Archives of General Psychiatry. 2010;67(9):889–894. doi: 10.1001/archgenpsychiatry.2010.110. [DOI] [PubMed] [Google Scholar]
- 149.Kanduc D, Stufano A, Lucchese G, Kusalik A. Massive peptide sharing between viral and human proteomes. Peptides. 2008;29(10):1755–1766. doi: 10.1016/j.peptides.2008.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Dietrich DE, Bode L. Human Borna disease virus-infection and its therapy in affective disorders. APMIS. 2008;116(124):61–65. doi: 10.1111/j.1600-0463.2008.00m10.x. [DOI] [PubMed] [Google Scholar]
- 151.Fazzini E, Fleming J, Fahn S. Cerebrospinal fluid antibodies to coronavirus in patients with Parkinson’s disease. Movement Disorders. 1992;7(2):153–158. doi: 10.1002/mds.870070210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Bagert BA. Epstein-Barr virus in multiple sclerosis. Current Neurology and Neuroscience Reports. 2009;9(5):405–410. doi: 10.1007/s11910-009-0059-9. [DOI] [PubMed] [Google Scholar]