

Abstract
Allergic asthma rates have increased steadily in developed countries, arguing for an environmental aetiology. To assess the influence of gut microbiota on experimental murine allergic asthma, we treated neonatal mice with clinical doses of two widely used antibiotics—streptomycin and vancomycin—and evaluated resulting shifts in resident flora and subsequent susceptibility to allergic asthma. Streptomycin treatment had little effect on the microbiota and on disease, whereas vancomycin reduced microbial diversity, shifted the composition of the bacterial population and enhanced disease severity. Neither antibiotic had a significant effect when administered to adult mice. Consistent with the ‘hygiene hypothesis’, our data support a neonatal, microbiota-driven, specific increase in susceptibility to experimental murine allergic asthma.
Keywords: antibiotics, asthma, gut, microbiota
Introduction
Allergic asthma affects over 100 million people worldwide and its prevalence is increasing, particularly among children in industrialized countries [1]. This growing epidemic has been attributed in part to improved sanitation methods and widespread antibiotic use, which have in turn reduced childhood exposures to microbial antigens critical for proper immune development [2]. Blaser and Falkow [3] have recently revisited the hygiene hypothesis, suggesting that it might not be a decline in childhood infections that is important, rather, it is how modern societal practices are causing the disappearance of ancestral species of our indigenous microbiota that might confer benefits beyond our current understanding. The loss of these ancestral species could be causing a rapid reorganization of the microbial hierarchy in our guts faster than our immune systems can adapt, promoting the emergence of dysregulated immunological disorders like allergic asthma.
Over the last several decades, evidence from human and mouse studies indicate that colonization of the gut early in life has a substantial role in directing immune system development. Disrupting this normal ‘immunological program’ could increase the likelihood of developing atopic disorders like asthma. Epidemiological data show gut microbiota differs between asthmatic and non-asthmatic infants [4], early life exposure to environmental microorganisms is protective [5] and early life as well as prenatal antibiotic exposures increase the risk of allergic asthma [6, 7, 8].
Antibiotic or probiotic administration as a means of microbiota manipulation in experimental models of murine allergic airways disease has provided further insights into what is now referred to as the ‘gut–lung axis’. Noverr et al [9, 10] describe how antibiotic treatment followed by oral administration of Candida albicans renders immunocompetent hosts susceptible to allergic airways disease. Similarly, oral administrations of Mycobacterium vaccae [11], Helicobacter pylori [12] as well as conventional probiotic strains [13, 14] have been shown to ameliorate symptoms of allergic airways disease in mice. Several of these studies highlight the necessity of neonatal administration rather than adult, emphasizing that immune modulation is most effective during the developmental period [12, 14]. Despite this growing body of evidence that supports a role for the gut microbiota in allergic disease, there is little known about how antibiotic-induced changes in microbial composition affect the immune status of the host and influence disease susceptibility.
The human gut is colonized by ∼1014 bacteria, and contains upwards of 1,000 bacterial species [15]. Specific subsets of the microbiota have been shown to differentially regulate immune function. For example, Bacteroides fragilis modulates T-helper-type 1/2 (Th1/Th2) balance [16], segmented filamentous bacteria direct Th17 cell differentiation [17] and Clostridium species induce regulatory T-cell (Treg) production [18]. Antibiotics have the ability to create global shifts in microbial communities, and are valuable tools for studying how changes in the gut microbiota influence disease states [19].
Here we characterize the effects of two antibiotics on resident murine gut flora and the subsequent susceptibility of these mice to experimental allergic asthma. Although it is known that this model only recapitulates some aspects of the human disease (particularly when mice are only given a single or limited number of antigen challenges) it does recapitulate the airway hyperresponsiveness (AHR) and some of the early phase responses [20]. It has therefore become a very popular model for evaluating susceptibility to Th2-driven allergic airways disease. Our data show that neonatal, but not adult, exposure to certain antibiotics promotes enhanced susceptibility to experimental allergic asthma.
Results
Antibiotic treatment increases asthma severity
To determine whether antibiotic perturbations of intestinal microbiota affect asthma, experimental murine allergic asthma was induced in mice exposed to vancomycin or streptomycin as neonates (including exposure in utero) or only as adults. Ovalbumin (OVA)-challenged mice treated early in life with vancomycin, but not streptomycin, resulted in increased total inflammatory cell infiltrates in the bronchoalveolar lavage (BAL) relative to OVA-challenged controls (Fig 1A). Treatment of adult mice with the same antibiotics had no effect on asthma induction (Fig. 1A and supplementary Fig S1 online). Neonatal vancomycin treatment also increased eosinophils in the BAL (Fig 1B). Antigen-specific immunoglobulin-E (IgE) was also significantly higher in the vancomycin-treated animals, consistent with exacerbated disease (Fig 1C). Antibiotic treatment alone (PBS-sensitized and challenged) had no effect on BAL counts (Fig 1A), eosinophil numbers or OVA-specific IgE (data not shown as there was none detected).
Figure 1.

Neonatal vancomycin treatment exacerbates allergic asthma. (A) Total cellular infiltrates from bronchoalveolar lavage (BAL) of control or antibiotic-treated mice challenged with ovalbumin (OVA) or PBS. (B) Eosinophil numbers in the BAL quantified by cytospin. (C) Serum OVA-specific IgE responses measured by enzyme-linked immunosorbent assay. (D) Airway hyperresponsiveness, measured by changes in resistance (R) in response to increasing doses of methacholine administered intravenously. (E) Total pathological scores and representative haematoxylin- and eosin-stained lung sections. Scale bar, 300 μm. All assessments were made on day 26. The data are shown as means of 5–7 mice per group±s.e.m. and represent at least two independent experiments. Statistics shown are based on comparisons to OVA-challenged controls. Antibiotic-treated animals in B–E were treated neonatally. *P<0.05, **P<0.01; Abx, antibiotics; ND, none detected; NS, not significant; Strep, streptomycin; Vanco, vancomycin.
Airway hyperresponsiveness, measured by changes in airway resistance (AR) in response to methacholine (MCh), is another defining feature of asthma. Both OVA-challenged control and streptomycin-treated mice exhibited increased AR over naive animals (Fig 1D); however, those treated with vancomycin showed markedly higher AHR compared to the other groups.
We examined lungs of mice from all treatment groups for histopathology to confirm allergic disease. Pathological scores were consistently higher in the vancomycin-treated animals (Fig 1E). Pathology in streptomycin-treated, OVA-challenged mice was comparable to control OVA-challenged mice. PBS-treated control mice were also compared to PBS-treated vancomycin and streptomycin-treated animals and no differences were detected (supplementary Fig S2 online). In summary, on the basis of an evaluation of BAL infiltrates, OVA-specific IgE, AHR and histopathology, we found that neonatal vancomycin treatment enhances the severity of experimental murine allergic asthma.
Vancomycin treatment profoundly alters gut flora
We next sought to correlate increased sensitivity to asthma to alterations in bacterial communities in the intestine. Total bacteria in stool pellets from antibiotic-treated mice were moderately reduced compared with the controls (Fig 2A). This reduction, however, was less pronounced than the 100-fold reduction previously observed in animals treated with antibiotics that greatly exceed therapeutic levels [21].
Figure 2.

Antibiotic treatment alters gut bacterial communities. (A) Total bacteria in faeces of control or antibiotic-treated mice quantitated by SYBR green. These data are shown as means of 3–5 mice per group±s.e.m. and represent two independent experiments. *P<0.05 relative to control; NS, not significant. (B) Bacterial communities from ileum and faeces of naive or ovalbumin (OVA)-challenged control and antibiotic-treated mice were compared using principal coordinate analysis (PCO). (C) Family level phylogenetic classification of 16S rRNA gene frequencies in faeces collected from naive or OVA-challenged control and antibiotic-treated animals. Those indicated with a classification level other than family level (f) could only be identified confidently to the level indicated. Classification scheme: k, kingdom; p, phylum; c, class; o, order; f, family. Each bar represents one mouse. Rare taxa were removed from the legend, but still included in the graph. Antibiotic-treated animals in A and B were treated neonatally. Abx, antibiotics; Strep, streptomycin; Vanco, vancomycin.
Changes in gut community composition after antibiotic treatment were assessed by pyrosequencing the V1–V2 regions of bacterial 16S rRNA genes. A total of 68,014 quality reads were obtained from 69 samples (the data set is available in the Sequence Read Archive (SRA) of NCBI, accession no. SRA050145). Principle coordinate analysis (PCO) showed that antibiotic treatment had the greatest effect on community composition, irrespective of sampling site (ileum/colon) or asthma induction (Fig 2B). In fact, 46.5% of the variability was explained on axis PCO1, emphasizing the large shift in community composition that resulted after vancomycin treatment compared with animals treated with streptomycin. Permutational multivariate analysis of variance indicated that antibiotic treatment significantly affected community composition (F=65.1, P<0.0004), and confirmed that the effect of vancomycin was much greater than that of streptomycin (Bray-Curtis similarity index: control versus streptomycin, 51.66; control versus vancomycin, 1.39). Estimated diversity was similar in control and streptomycin-treated communities, but significantly lower in those treated with vancomycin (Simpson's diversity indices: 0.90, 0.81 and 0.33, respectively). Strikingly, the average number of operational taxonomic units per sample dropped from 54 in the control to 48 in those treated with streptomycin, to only 6 after vancomycin treatment. The 16S rRNA sequences were taxonomically identified at the genus level using the Greengenes database [22] and a naive Bayesian classifier. The 16S rRNA genes and their frequencies classified at the family level (for simplicity) in faeces from control, streptomycin- or vancomycin-treated animals further emphasizes how significantly the bacterial community in the gut changed after antibiotic treatment, particularly in the animals that received vancomycin, irrespective of exposure type (Fig 2C).
Both neonatal and adult antibiotic treatment significantly altered microbial communities in the gut; however, these changes were not identical. The neonatal vancomycin-treated animals possessed a microbiota that was completely different from animals treated with vancomycin only as adults (Bray-Curtis similarity index: 34.7). Interestingly, communities from mice treated with vancomycin only as adults had greater diversity and were less distinct from the other treatments than the communities from mice treated with vancomycin as neonates (Simpson's diversity index: 0.67 and 0.18, respectively). When neonatal and adult antibiotic treatments were compared using PCO, it was clear that the two different exposures produced distinct community structures (supplementary Fig S3 online). Importantly, these data represent two independent experiments, in which cage effects, parent litter and age are all factors that affect microbiota composition, and could contribute to the significant differences observed between even the untreated controls in these experiments.
Antibiotic-specific microbial indicators of asthma
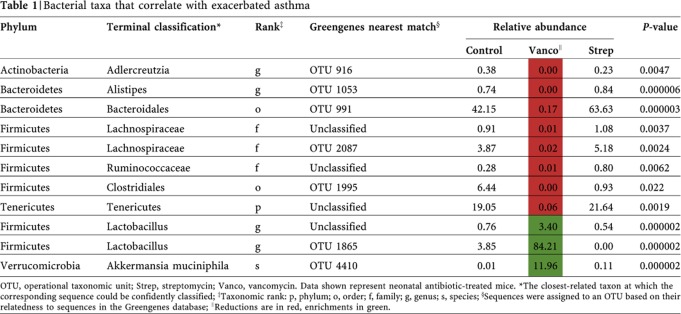
Given that streptomycin-treated animals had comparable disease to controls, bacteria that were depleted or enriched after vancomycin, but not streptomycin, were identified as candidates that might affect disease susceptibility (Table 1). This list highlights bacterial taxa that were significantly depleted or enriched in the vancomycin-treated animals based on the point biserial correlation coefficient (supplementary methods online). For sequence information on these indicators refer to supplementary methods, Data Set S1 online.
Table 1. Bacterial taxa that correlate with exacerbated asthma.
Antibiotics alter CD4+CD25+Foxp3+ Tregs in the colon
Several members of the indigenous gut microbiota have been implicated in the induction of Treg cells in the gut [18, 23]. To provide insight into how vancomycin-treated microbiota might predispose mice to exacerbated experimental asthma, we compared levels of CD4+CD25+Foxp3+ T cells (a differentiation antigen expression phenotype typical of Treg cells) in the colon of untreated, vancomycin- and streptomycin-treated animals. We found colonic CD4+CD25+Foxp3+ T cells to be strikingly reduced in the vancomycin-treated animals (Fig 3A), while their frequency in streptomycin-treated animals were comparable to controls. This reduction in phenotypic Tregs was not detected in the lungs of vancomycin-treated animals (Fig 3B). There is increasing evidence to suggest that mucosal sites function together as a system-wide organ [24]. The gut–lung axis, a proposed mechanism of probiotic action, is an extension of this idea. Probiotic studies indicate that tolerogenic responses from the gut via gut-associated lymphoid tissue spread to the airways only in response to immune challenge and inflammation [25]. The data shown represent Treg populations from naive, unchallenged mice. Hence, further studies are required to determine whether a key orchestrator of tolerogenic mucosal immune responses is altered in vancomycin-treated animals.
Figure 3.

Vancomycin treatment reduces CD4+CD25+Foxp3+ regulatory T-cell accumulation in the colon but not in the lung. At 7 weeks of age, the percentage of CD25+Foxp3+ cells within the CD45+CD4+ cell population in the colon (A) or lung (B) of naive mice treated with neonatal antibiotics was analysed. The data shown are means of three mice per group±s.e.m. and represent three independent experiments. Vanco, vancomycin; Strep, streptomycin. **P<0.01.
Discussion
Over the last several decades, substantial effort has been focused on identifying bacterial populations that correlate with the development of allergy-related disorders [4, 25]. The results differ significantly depending on the study, which likely stems from differences in sample populations and methods of microbiota analysis. Nevertheless, most of these studies point towards the first 6 months of life as a ‘critical window’, during which the infant gut microbiota might influence key immunological events that alter allergic sensitization [4]. In the current study we have addressed the same question using a different approach: by creating defined shifts in the microbiota with antibiotics early in life, we have identified members of the microbiota that differ between more and less susceptible individuals.
In this study, animals treated neonatally with vancomycin showed increases in all major indicators of experimental allergic asthma. Correspondingly, vancomycin treatment markedly shifted the composition of the gut microbiota, compared with both streptomycin-treated and control populations. Adult vancomycin treatment also altered the microbiota, however, to a different extent, which could be why differences in disease susceptibility ensued. Because our neonatal antibiotic model encompassed several critical time points (that is, prenatal, perinatal, weaning), it will be important to look at how antibiotic treatment at each time point individually affects disease outcome.
Microbial diversity was also reduced after vancomycin, but not streptomycin treatment, despite having similar numbers of total bacteria. There is evidence that reduced diversity in the infant gut microbiota correlates with increased risk of allergic disorders [26]. On the basis of the extent of microbial reorganization in the gut after neonatal antibiotic treatment, several bacterial populations were positively and negatively associated with disease severity.
Vancomycin-shifted microbiota could alter host immune development in several ways such as inducing Th2 hyperreactivity or reducing key tolergenic regulatory mechanisms. Consistent with our report, Atarashi et al [18] observed that vancomycin treatment reduces colonic CD4+Foxp3+ Tregs. They suggest that this observation is due to a reduction in Clostridium species (clusters IV and XIVa) that they identify as potent inducers of Tregs. We also found that depletion of Clostridiales with vancomycin correlated with a reduction in cells expressing a CD4+CD25+Foxp3+ Treg phenotype; however, vancomycin treatment also reduced other bacterial phyla. Unlike the colon, we were not able to detect significant changes in CD4+CD25+Foxp3+ Treg numbers in the lung. Perhaps a more detailed look at Treg populations in the lung and mediastinal lymph nodes over the course of disease induction, measuring Treg suppressive capacity or analysing other regulatory cell populations (for example, interleukin 10 producing Tr1 cells), may provide new insights.
Surprisingly, Bacteroidetes were almost entirely depleted after vancomycin treatment, despite not being directly targeted by the antibiotic; they were replaced by an overgrowth of Lactobacilli. Several Bacteroides strains have been implicated in Treg differentiation [18, 23] and their depletion might also explain the altered susceptibility to experimental allergic asthma. Epidemiological data indicate that antibiotic use in early infancy as well as infants born by caesarean section show similar decreases in Bacteroides species [27], both of which are factors associated with increased childhood asthma [8, 28]. Lactobacilli were negatively correlated with Treg abundance in this study, despite their previous associations with Treg induction in probiotic studies [29]. These regulatory effects are likely strain-dependent, and perhaps there are other species or strains of Lactobacilli that have reciprocal effects that remain uncharacterized. Interestingly, Lactobacilli actually only represent a small proportion of total bacteria in the healthy mammalian intestine [21] despite their potential benefits, which suggests that an overgrowth of particular Lactobacilli species might do more harm than good. Colonizing gnotobiotic animals with subsets of the bacteria discussed here might help identify whether they have specific roles in the development of experimental allergic asthma.
In our study, two events were required to significantly affect the outcome of experimental asthma: (1) global restructuring of the intestinal microbiota with vancomycin and (2) altering the microbiota during infancy, a time point when key immunological thresholds are established. There is substantial evidence that microbial signals are required for lymphoid tissue development and maintenance in the gut (reviewed in Hill and Artis [30][30]). Vancomycin-shifted microbiota likely produces an entirely different set of microbial signals compared with healthy controls that could alter intestinal epithelial cell signalling cascades and dysregulate innate and adaptive immune responses. It is already known that mice treated with vancomycin have fewer Th17 cells in the small intestine [31] and fewer CD4+ Treg cells in the colon [18]. Studies in germ-free animals show that the regulation of immune cell subsets (monocyte/macrophages, B cells, CD4+Treg/Th17 and CD8+ T cells) by gut microbes is not restricted to the intestinal compartment [30]. Similarly, mice deficient in the innate recognition receptors TLR4 or GPR43 show exacerbated asthma responses [32, 33]. Insights from antibiotic use in other disease models suggest mechanisms mediated by skewed Th2 immune responses [34] and elevated serum IgE levels [35]. Dendritic cells have also been implicated in the pathogenesis of asthma [36], and are thought to be involved in bridging the gut–lung axis [37], however whether or not signals from intestinal bacteria influence systemic dendritic cell populations remains to be tested directly. Although the link between gut and lung mucosal immune responses remains unclear, we propose that vancomycin administered early in life selects for a community of microbes that disrupt the balance of proinflammatory and regulatory immune responses occurring at local and distant mucosal sites.
In summary, using relevant doses of antibiotics we have found that neonatal, but not adult, treatment with select antibiotics can have profound effects on susceptibility to experimental murine allergic asthma. While antibiotic treatment did not substantially deplete bacterial numbers, it resulted in profound shifts in microbial composition. This was correlated with the loss of a key regulatory subset of immune cells in the colon that could, by way of the gut–lung axis, promote enhanced susceptibility to inflammatory diseases in the lung by a mechanism that remains undefined. This work provides a new framework for interrogating the altered communities responsible for heightened Th2 immune responses in human disease, including asthma.
Methods
Mice. C57BL/6J mice (Jackson Laboratories, Bar Harbor, ME) were bred and maintained in a specific pathogen-free facility at The Biomedical Research Center. All experiments were in accordance with the University of British Columbia Animal Care Committee guidelines.
Antibiotic treatment. C57BL/6J breeding pairs were given vancomycin or streptomycin (Sigma-Aldrich, St Louis, MI) at 200 mg l−1 in drinking water. Pups born from respective breeding pairs were reared on antibiotic-treated water with their littermates for the duration of the experiment. Hence, our term ‘neonatal exposure’ refers to antibiotic exposure both in utero and after birth. Alternatively, 7-week-old C57BL/6J female mice were given vancomycin or streptomycin (Sigma) at 200 mg l−1 in drinking water for 2 days before sensitization with OVA, hence ‘adult exposure’.
Vancomycin, an antibiotic that directly targets Gram-positive bacteria, and streptomycin, an antibiotic that directly targets both Gram-positive and Gram-negative bacteria, were chosen because both are used in clinical practice and are poorly absorbed when given orally, minimizing the risk of systemic effects on the host. The rationale behind the clinically relevant concentrations used has been described in detail previously [38].
Ovalbumin model of experimental murine allergic asthma. Experimental murine allergic asthma was induced as previously described [39] with minor modifications. Although this model does not fully recapitulate the phenotype of human allergic asthma (particularly after limited numbers of antigen challenge [20]), it is a useful model for evaluating many aspects of this Th2-driven lung inflammatory disease. Mice were sensitized intraperitoneally with 200 μg OVA and 1.3 mg alum (both from Sigma) on days 0 and 7. On days 21, 22, 23 and 25, mice were challenged intranasally with 1 mg OVA in PBS. On day 26, mice were anaesthetized with 200 mg kg−1 ketamine per 10 mg kg−1 xylazine and blood was collected by cardiac puncture. After killing, BALs were performed by 3 × 1 ml washes with PBS. Total BAL cells were counted by hemocytometer and eosinophils were quantified from cytospins (Thermo Shandon, Pittsburg, PA) stained with HemaStain (Fisher Scientific), based on standard morphological criteria.
Determination of serum IgE. OVA-specific IgE in serum was measured by enzyme-linked immunosorbent assay (Chondrex, Redmond, WA).
Airway hyperresponsiveness. Airway hyperresponsiveness was measured on day 26. Mice were anaesthetized, tracheotomized, and intubated with a cannula. Following paralysis with pancuronium bromide (0.5 mg kg−1), AR was measured with a flexiVent apparatus (SCIREQ, Montreal, QC, Canada). Increasing concentrations of MCh (0–1.5 mg kg−1) were administered via the jugular vein. MCh response curves were generated by calculating the change in AR relative to baseline (PBS) for each MCh dose.
Histology. Lungs were collected and fixed in 10% formalin, embedded in paraffin, cut longitudinally into 5-μm sections and stained with haematoxylin and eosin. Inflammation was blindly assessed from five fields per section, each graded on a scale of 0–5 (0=no signs of disease, 5=severe disease) for each of the following parameters: (1) peribronchial infiltration, (2) perivascular infiltration, (3) parenchymal infiltration and (4) epithelium damage for a maximum score of 20.
Microbial analysis. Bacteria in stool pellets of 7-week-old naive mice were stained with SYBR green and counted as previously described [40]. For composition analyses, stool pellets and ileal contents from naive or OVA-challenged mice were homogenized using a bead-beating method (FastPrep instrument, MP Biomedicals, Solon, OH) and total DNA was extracted (Ultra Clean Fecal DNA kit, Mo Bio Laboratories, Carlsbad, CA). 16S rRNA gene fragments were amplified using 33 nucleotide-bar-coded primer pairs (27F; 5′-AGAGTTTGATCMTGGCTCAG-3′), (519R; 5′-GWATTACCGCGGCKGCTG-3′). PCR products were gel-purified (QIAquick gel extraction kit, Qiagen, Valencia, CA). Each amplicon (100 ng) was pooled and pyrosequenced using a 454 Titanium platform (Roche, Branford, CT).
Bioinformatics. High-quality sequence reads were determined using MOTHUR[41] and V-XTRACTOR [42]. Each read was assigned to the GREENGENES database [22] using a naive Bayesian classifier [43]. Global community structure comparisons from faecal and ileal samples were made using PCO [44], permutational multivariate analysis of variance [45] and Simpson's diversity index [46] implemented in PRIMER6+ [47]. Taxa-treatment association analysis [48] in conjunction with additional statistical analyses (supplementary methods online) were performed to determine antibiotic-related indicator species (faecal samples only). Raw, unfiltered sequences were submitted to the SRA of NCBI.
Isolation of lymphocytes and flow cytometry. Colon and lung tissues were collected from 7-week-old naive mice and cells were isolated as previously described [49]. Cells were stained with fluorochrome-conjugated antibodies against CD45, CD4, CD25 and Foxp3 (BD Biosciences, Franklin Lakes, NJ). Flow cytometry was performed using an LSR II (BD Biosciences) and data were analysed with FlowJo 8.7 software (TreeStar, Ashland, OR).
Statistics. Differences between control and experimental groups were compared using an unpaired Student's t-test to calculate statistical significance (GraphPad Prism software, version 4.0, San Diego, CA).
Supplementary information is available at EMBO reports online (http://www.emboreports.org).
Supplementary Material
Acknowledgments
We thank Michael Hughes, Jami Bennett, Roland Scholz and Marie-Claire Arrieta for their helpful advice; Kay Jian for her technical support, Pedro Dimitriu for pyrosequencing support; and members of The Biomedical Research Center Animal Facility for expert animal care. This work was funded by a Canadian Institutes of Health Research (CIHR) Catalyst Grant and a CIHR Emerging Team Grant in partnership with Genome BC and the AllerGen NCE.
Author contributions: S.L.R: study concept and design, data acquisition and analysis; M.J.G.: experimental asthma data acquisition and analysis; M.H.: microbial ecology-related bioinformatics and statistical analyses; B.P.W.: intellectual content, study design, data acquisition and analysis; L.T., M.W., N.G.: intellectual content, data acquisition and analysis; M.-R.B.: study design; W.W.M.: intellectual content; K.M.M. and B.B.F.: supervision, study design and interpretation.
Footnotes
The authors declare that they have no conflict of interest.
References
- Lai CK, Beasley R, Crane J, Foliaki S, Shah J, Weiland S (2009) Global variation in the prevalence and severity of asthma symptoms: phase three of the International Study of Asthma and Allergies in Childhood (ISAAC). Thorax 64: 476–483 [DOI] [PubMed] [Google Scholar]
- Strachan DP (1989) Hay fever, hygiene, and household size. Br Med J 299: 1259–1260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blaser MJ, Falkow S (2009) What are the consequences of the disappearing human microbiota? Nat Rev Microbiol 7: 887–894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penders J, Stobberingh EE, van den Brandt PA, Thijs C (2007) The role of the intestinal microbiota in the development of atopic disorders. Allergy 62: 1223–1236 [DOI] [PubMed] [Google Scholar]
- Ege MJ, Mayer M, Normand AC, Genuneit J, Cookson WO, Braun-Fahrlander C, Heederik D, Piarroux R, von Mutius E (2011) Exposure to environmental microorganisms and childhood asthma. N Engl J Med 364: 701–709 [DOI] [PubMed] [Google Scholar]
- Marra F, Marra CA, Richardson K, Lynd LD, Kozyrskyj A, Patrick DM, Bowie WR, Fitzgerald JM (2009) Antibiotic use in children is associated with increased risk of asthma. Pediatrics 123: 1003–1010 [DOI] [PubMed] [Google Scholar]
- Martel MJ, Rey E, Malo JL, Perreault S, Beauchesne MF, Forget A, Blais L (2009) Determinants of the incidence of childhood asthma: a two-stage case-control study. Am J Epidemiol 169: 195–205 [DOI] [PubMed] [Google Scholar]
- Murk W, Risnes KR, Bracken MB (2011) Prenatal or early-life exposure to antibiotics and risk of childhood asthma: a systematic review. Pediatrics 127: 1125–1138 [DOI] [PubMed] [Google Scholar]
- Noverr MC, Falkowski NR, McDonald RA, McKenzie AN, Huffnagle GB (2005) Development of allergic airway disease in mice following antibiotic therapy and fungal microbiota increase: role of host genetics, antigen, and interleukin-13. Infect Immun 73: 30–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noverr MC, Noggle RM, Toews GB, Huffnagle GB (2004) Role of antibiotics and fungal microbiota in driving pulmonary allergic responses. Infect Immun 72: 4996–5003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt JR, Martinelli R, Adams VC, Rook GA, Brunet LR (2005) Intragastric administration of Mycobacterium vaccae inhibits severe pulmonary allergic inflammation in a mouse model. Clin Exp Allergy 35: 685–690 [DOI] [PubMed] [Google Scholar]
- Arnold IC, Dehzad N, Reuter S, Martin H, Becher B, Taube C, Muller A (2011) Helicobacter pylori infection prevents allergic asthma in mouse models through the induction of regulatory T cells. J Clin Invest 121: 3088–3093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forsythe P, Inman MD, Bienenstock J (2007) Oral treatment with live Lactobacillus reuteri inhibits the allergic airway response in mice. Am J Respir Crit Care Med 175: 561–569 [DOI] [PubMed] [Google Scholar]
- Lyons A et al. (2011) Bacterial strain-specific induction of Foxp3+ T regulatory cells is protective in murine allergy models. Clin Exp Allergy 40: 811–819 [DOI] [PubMed] [Google Scholar]
- Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI (2005) Host-bacterial mutualism in the human intestine. Science 307: 1915–1920 [DOI] [PubMed] [Google Scholar]
- Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL (2005) An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122: 107–118 [DOI] [PubMed] [Google Scholar]
- Ivanov II et al. (2009) Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139: 485–498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atarashi K et al. (2011) Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331: 337–341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willing BP, Russell SL, Finlay BB (2011) Shifting the balance: antibiotic effects on host-microbiota mutualism. Nat Rev Microbiol 9: 233–243 [DOI] [PubMed] [Google Scholar]
- Zosky GR, Larcombe AN, White OJ, Burchell JT, Janosi TZ, Hantos Z, Holt PG, Sly PD, Turner DJ (2008) Ovalbumin-sensitized mice are good models for airway hyperresponsiveness but not acute physiological responses to allergen inhalation. Clin Exp Allergy 38: 829–838 [DOI] [PubMed] [Google Scholar]
- Hill DA, Hoffmann C, Abt MC, Du Y, Kobuley D, Kirn TJ, Bushman FD, Artis D (2010) Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal Immunol 3: 148–158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeSantis TZ et al. (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol 72: 5069–5072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Round JL, Mazmanian SK (2010) Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci USA 107: 12204–12209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill N, Wlodarska M, Finlay BB (2010) The future of mucosal immunology: studying an integrated system-wide organ. Nat Immunol 11: 558–560 [DOI] [PubMed] [Google Scholar]
- Forsythe P (2011) Probiotics and lung diseases. Chest 139: 901–908 [DOI] [PubMed] [Google Scholar]
- Bisgaard H, Li N, Bonnelykke K, Chawes BL, Skov T, Paludan-Muller G, Stokholm J, Smith B, Krogfelt KA (2011) Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J Allergy Clin Immunol 128: 646–652, e645 [DOI] [PubMed] [Google Scholar]
- Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, van den Brandt PA, Stobberingh EE (2006) Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 118: 511–521 [DOI] [PubMed] [Google Scholar]
- van Nimwegen FA et al. (2011) Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J Allergy Clin Immunol 128: 948–955, e941–943 [DOI] [PubMed] [Google Scholar]
- Karimi K, Inman MD, Bienenstock J, Forsythe P (2009) Lactobacillus reuteri-induced regulatory T cells protect against an allergic airway response in mice. Am J Respir Crit Care Med 179: 186–193 [DOI] [PubMed] [Google Scholar]
- Hill DA, Artis D (2010) Intestinal bacteria and the regulation of immune cell homeostasis. Annu Rev Immunol 28: 623–667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanov II, Frutos Rde L, Manel N, Yoshinaga K, Rifkin DB, Sartor RB, Finlay BB, Littman DR (2008) Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 4: 337–349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dabbagh K, Dahl ME, Stepick-Biek P, Lewis DB (2002) Toll-like receptor 4 is required for optimal development of Th2 immune responses: role of dendritic cells. J Immunol 168: 4524–4530 [DOI] [PubMed] [Google Scholar]
- Maslowski KM et al. (2009) Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461: 1282–1286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oyama N, Sudo N, Sogawa H, Kubo C (2001) Antibiotic use during infancy promotes a shift in the T(H)1/T(H)2 balance toward T(H)2-dominant immunity in mice. J Allergy Clin Immunol 107: 153–159 [DOI] [PubMed] [Google Scholar]
- Bashir ME, Louie S, Shi HN, Nagler-Anderson C (2004) Toll-like receptor 4 signaling by intestinal microbes influences susceptibility to food allergy. J Immunol 172: 6978–6987 [DOI] [PubMed] [Google Scholar]
- Lambrecht BN, Hammad H (2003) Taking our breath away: dendritic cells in the pathogenesis of asthma. Nat Rev Immunol 3: 994–1003 [DOI] [PubMed] [Google Scholar]
- Noverr MC, Huffnagle GB (2005) The ‘microflora hypothesis’ of allergic diseases. Clin Exp Allergy 35: 1511–1520 [DOI] [PubMed] [Google Scholar]
- Sekirov I, Tam NM, Jogova M, Robertson ML, Li Y, Lupp C, Finlay BB (2008) Antibiotic-induced perturbations of the intestinal microbiota alter host susceptibility to enteric infection. Infect Immun 76: 4726–4736 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchet MR, Maltby S, Haddon DJ, Merkens H, Zbytnuik L, McNagny KM (2007) CD34 facilitates the development of allergic asthma. Blood 110: 2005–2012 [DOI] [PubMed] [Google Scholar]
- Lupp C, Robertson ML, Wickham ME, Sekirov I, Champion OL, Gaynor EC, Finlay BB (2007) Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2: 204. [DOI] [PubMed] [Google Scholar]
- Schloss PD et al. (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75: 7537–7541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann M, Howes CG, Abarenkov K, Mohn WW, Nilsson RH (2010) V-Xtractor: an open-source, high-throughput software tool to identify and extract hypervariable regions of small subunit (16S/18S) ribosomal RNA gene sequences. J Microbiol Methods 83: 250–253 [DOI] [PubMed] [Google Scholar]
- Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73: 5261–5267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gower JC (1966) Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 53: 325–32& [Google Scholar]
- Anderson MJ (2001) Permutation tests for univariate or multivariate analysis of variance and regression. Can J Fish and Aquat Sci 58: 626–639 [Google Scholar]
- Simpson EH (1949) Measurement of diversity. Nature 163: 688–688 [Google Scholar]
- Clarke KR, Gorley RN (2006) PRIMER v6, User Manual/Tutorial. PRIMER-E, Plymouth (http://www.primer-e.com/Primary_papers.htm)
- De Caceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecology 90: 3566–3574 [DOI] [PubMed] [Google Scholar]
- Wlodarska M, Willing B, Keeney KM, Menendez A, Bergstrom KS, Gill N, Russell SL, Vallance BA, Finlay BB (2011) Antibiotic treatment alters the colonic mucus layer and predisposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect Immun 79: 1536–1545 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.