

Abstract
Little is known about stem cell biology or the specialized environments or niches believed to control stem cell renewal and differentiation in self-renewing tissues of the body. Functional assays for stem cells are available only for hematopoiesis and spermatogenesis, and the microenvironment, or niche, for hematopoiesis is relatively inaccessible, making it difficult to analyze donor stem cell colonization events in recipients. In contrast, the recently developed spermatogonial stem cell assay system allows quantitation of individual colonization events, facilitating studies of stem cells and their associated microenvironment. By using this assay system, we found a 39-fold increase in male germ-line stem cells during development from birth to adult in the mouse. However, colony size or area of spermatogenesis generated by neonate and adult stem cells, 2–3 months after transplantation into adult tubules, was similar (∼0.5 mm2). In contrast, the microenvironment in the immature pup testis was 9.4 times better than adult testis in allowing colonization events, and the area colonized per donor stem cell, whether from adult or pup, was about 4.0 times larger in recipient pups than adults. These factors facilitated the restoration of fertility by donor stem cells transplanted to infertile pups. Thus, our results demonstrate that stem cells and their niches undergo dramatic changes in the postnatal testis, and the microenvironment of the pup testis provides a more hospitable environment for transplantation of male germ-line stem cells.
In the body, there are several types of self-renewing tissues, including bone marrow, epidermis, intestinal epithelium, and testis. These tissues contain stem cells that self-renew and produce daughter cells continuously to repopulate the tissue with progenitor and differentiated cells throughout life. Studies on stem cell biology have been difficult because they require a functional assay that exists only for hematopoietic stem cells (HSCs) and spermatogonial stem cells (1–4). Generally, a stem cell is present in a special cellular organization called a niche, which provides a microenvironment designed to maintain the proliferation or survival of stem cells (5). In a niche, the stem cell has a high probability of self-renewal and divides rarely. When a stem cell divides, it has been proposed that only one daughter can remain in the niche, and the other cell will be committed to differentiate unless another niche is available. The niche must provide factors that maintain stem cells and exclude factors that induce differentiation (5–8). By definition, if space with the necessary microenvironment were limited, the number of stem cells also would be limited (5). Although the niche theory was proposed originally to explain the behavior of transplanted hematopoietic stem cells (5), this theory now has been extended to stem cells in several organs (6–9), and the niche is thought to be involved in the transdifferentiation of stem cells (10). Thus, the niche in self-renewing tissues controls the proliferation and commitment of stem cells and plays a critical role in stem cell biology.
Currently, the important role of the niche has been studied most thoroughly for HSCs, for which partial in vitro reconstitution of the stem cell microenvironment is possible (11). It was shown that HSCs must be in direct contact with stromal cells to facilitate survival, growth, and differentiation when they are cultured in vitro (12). If they are prevented from attaching to stromal cells, they die. Thus, the survival and proliferation of stem cells depends on close association with specific support cells, and these in vitro studies have contributed to the current concept of the niche. Recent reports provide insight into the Drosophila germ cell niche, in vivo (6–8), but little is known about how stem cells interact with the niche in mammals. These investigations have been hampered in hematopoietic tissues because of the relative inaccessibility of the stem cell and its environment in vivo, and even less is known about the niche for other stem cell tissues for which a functional assay is not available.
Testis seminiferous tubule epithelium is the only other self-renewing tissue in which there is a functional assay for stem cell activity. In the mouse, spermatogenesis begins immediately after birth. The only germ cell in the newborn mouse testis is the prospermatogonia, which is located in the center of the seminiferous tubule (13, 14). By 6 days postpartum (PP), these cells migrate to the basement membrane and produce undifferentiated type A spermatogonia, which begin to differentiate in a stepwise manner (15). Spermatozoa are not found in the seminiferous tubules until ≈35 days PP. During this period, the testicular microenvironment also undergoes significant changes. The Sertoli cell, a major somatic cell type in the seminiferous tubule, is known to interact with germ cells to provide essential support for successful spermatogenesis (16). Soon after birth, immature Sertoli cells begin to proliferate and continue to divide until 10–12 days PP (16). At this time, proliferation ceases and tight junctions form between neighboring Sertoli cells, which constitute the blood–testis barrier and compartmentalize the seminiferous tubule into basal and apical regions (16). Thus, formation of the spermatogonial stem cell niche takes place during a very short postnatal period and is accessible for study. In contrast, hematopoiesis begins before birth, and interaction between stem cell and niche is established in the fetus, which complicates analysis of the process.
As an additional advantage to the study of spermatogenesis, an assay to assess stem cell number and activity of donor testis cell populations has been developed recently (3, 4). In this technique, donor cells are injected into the seminiferous tubule of an infertile recipient male that lacks endogenous spermatogenesis. Transplanted spermatogonial stem cells generate colonies of spermatogenesis in recipient seminiferous tubules, and the resulting spermatozoa transmit the donor haplotype to offspring (3, 17). By definition, stem cells are the only cell type that can produce this result. Each colony seems to represent proliferation and differentiation from a single stem cell (18), and the number of colonies produced is indicative of the number of stem cells in the donor population (19). A particular advantage of this technique is that transplanted cells are introduced directly into and restricted to the testis; thus, individual colonization events can be counted and the kinetics of colony expansion quantitated after transplantation. Therefore, this technique not only provides an assay system for the identification of spermatogonial stem cells, similar to the stem cell assay for HSCs (1), but also allows clonal analysis of each transplanted stem cell in association with its differentiated products (18, 19). Using the spermatogonial stem cell assay as a functional endpoint, we sought to investigate differences that might exist between stem cells in young and adult animals, and whether accessibility to the stem cell niche or microenvironment changes during postnatal development.
Materials and Methods
Donor Mice and Cell Collection.
Neonate (day 0–2 PP), pup (day 5–12 PP), and cryptorchid adult (14–20 wk of age) donor testis cells used for the first experiment were isolated from the transgenic mouse strain B6;SJL-TgN(c177lacZ)227Bri (The Jackson Laboratory), which contains a zinc finger (ZF) promoter coupled to the Escherichia coli lacZ (E. coli lacZ) structural gene. This mouse strain is designated ZFlacZ, and the transgene is expressed in round spermatids (20). Donor cells used for the second experiment were isolated from the Fn progeny (designated ROSA26) of B6.129S7-Gtrosa26 (The Jackson Laboratory) males and C57BL/6 females. These mice express the E. coli lacZ structural gene in all cells of the seminiferous tubule (14, 21). Transplanted donor cells that express β-galactosidase from both strains can be stained by using the substrate 5-bromo-4-chloro-3-indolyl β-d-galactoside (X-gal).
Cryptorchid testes were produced as described and lack all differentiated germ cells by the time of donor cell collection, 2–3 months after the cryptorchid operation (22). Single-cell suspensions from neonate, pup, and cryptorchid adult donor testes were produced by enzymatic digestion (3, 23, 24). Isolated cells were suspended at concentrations ranging from 2 to 30 × 106 cells per ml in Dulbecco-modified Eagle's medium, supplemented as described (24), and maintained at 5°C throughout the procedure. Cell concentration must be adjusted for each experiment so that individual colonies can be distinguished easily and then counted.
Recipient Mice and Transplantation Procedure.
In the first experiment, C57BL/6 × SJL F1 hybrid mice (B6/SJL), treated i.p. with busulfan (44 mg/kg) at 4–6 wk of age, were used as immunologically compatible recipients of ZFlacZ donor cells (3). In the second experiment, ROSA26 testis cells were transplanted into immunologically compatible W54/Wv or Wv/W pup and adult recipients, designated W (refs. 17 and 25; The Jackson Laboratory), that lack spermatogenesis because of a mutation in the gene for the c-Kit receptor (26). Adult recipient mice were anesthetized by Avertin injection (640 mg/kg, i.p.). Pup recipients were placed on ice to cause hypothermia-induced anesthesia (27). Approximately 10 μl of donor-cell suspension was introduced into each testis of adult recipients for experiment 1. For the W mice used in experiment 2, 4 μl were injected into each adult and 2 μl into each pup recipient testis. Two to three experiments were performed for each donor cell type, and at least seven recipient testes were analyzed per recipient group.
Analysis of Recipient Testes.
Recipient testes were collected 3 (first experiment) or 2 months (second experiment) after donor-cell transplantation, stained with X-gal to visualize donor-derived spermatogenesis (19, 28), and analyzed by a computer-assisted imaging system (18). A donor spermatogonial stem cell is defined by its ability to produce a blue-staining spermatogenic colony in a recipient testis. Other types of testis cells do not generate colonies of spermatogenesis, and endogenous recipient testis cells do not express the lacZ transgene. The ability to spread along the seminiferous tubule was determined by measuring the blue area (mm2) per colony (18). Colony number and colonized area were normalized to 105 cells injected.
The purpose of our analyses was to determine whether there were any statistically significant differences among neonate, pup, and adult donors, and between pup and adult recipients with respect to the number of colonies per 105 cells (Figs. 1B and 2A) and total colony area (mm2; Fig. 1B) or blue area per colony (mm2; Fig. 2B). To account for the repeated measures on each animal (from left and right testes), generalized estimating equations methodology was used to model each outcome (29). This methodology accounts for the correlation between the repeated measures within an animal and does not delete any animal having some missing repeats (as a repeated-measures ANOVA would do).
Figure 1.

Determination of spermatogonial stem cell number in neonate, pup, and cryptorchid adult ZFlacZ donor testes by transplantation into busulfan-treated adult B6/SJL recipients. (A) Macroscopic (Top) and histological (Bottom) appearance of ZFlacZ donor testes. Neonate (day 0–2 PP), pup (day 6 PP), and cryptorchid adult testes are shown left, middle, and right, respectively. Neonate testes weighed 0.94 ± 0.1 mg, and 0.62 × 106 ± 0.14 cells were recovered per testis. Pup testes weighed 4.1 ± 0.1 mg, and 0.98 × 106 ± 0.17 cells were recovered per testis. Cryptorchid adult testes weighed 24.1 ± 0.9 mg, and 1.82 × 106 ± 0.04 cells were recovered per testis. There are many more interstitial cells in testes as they mature from neonate to adult that are removed during testis cell recovery (see Materials and Methods). Testis weight and cell recovery data were based on n ≥ 6. Testes from all donor ages were similarly devoid of differentiated germ cells and therefore show no X-gal staining. Arrows indicate germ cells. [Bars = 2 mm (Top) and 20 μm (Bottom Inset).] Testes were stained with X-gal, sectioned, and counterstained with nuclear fast red. (B) Colonization of recipient testes by donor testis cells from developmental ages in A. Approximately 1–3 × 105 cells were injected into each testis. Spermatogonial stem cell number and colony expansion are determined by the number of individual blue colonies of spermatogenesis (black bars) and total blue area (open bars) per 105 cells injected, respectively. The number of colonies produced per 105 cells from neonate, pup, and cryptorchid adult were 3.4 ± 0.4, 13.9 ± 2.8, and 46.1 ± 3.5, respectively, and blue area was 2.22 ± 0.30 mm2, 7.21 ± 1.95 mm2, and 22.34 ± 2.23 mm2, respectively (mean ± SEM; n = 22 for neonate and pup; n = 23 for cryptorchid adult).
Figure 2.

Evaluation of recipient environment (pup vs. adult) and its effect on donor spermatogonial stem cell colonization. Pup (day 5–12 PP) and cryptorchid adult ROSA26 donor testis cells were transplanted into pup or adult W recipients (Wv/W or W54/Wv). Donor pup testes weighed 2.34 ± 0.37 mg, and 1.18 × 106 ± 0.17 × 106 cells were recovered per testis. Cryptorchid adult donor testes weighed 22.18 ± 0.59 mg, and 1.15 × 106 ± 0.17 × 106 cells were recovered per testis. Testis weight and cell recovery data were based on n ≥ 4. Degree of colonization, 2 months after transplantation, is represented by number of individual blue colonies per 105 cells injected (A), and by blue area (mm2) per colony (B). Black bars represent pup donors and open bars represent adult donors. Note that regardless of donor, pup recipients had more colonies per 105 cells (A) and a greater area (mm2) per colony (B). Values are means ± SEM; n = 5–14.
To determine whether infertile recipient males could be made fertile as a result of the transplantation procedure, pup and adult W recipient testes were injected with adult cryptorchid ROSA26 donor cells (6 × 104 cells per testis) and mated with nontransgenic female mice. At least 3 litters and 26 progeny were produced from each fertile recipient mouse and were analyzed for β-galactosidase activity by staining with X-gal. Approximately 5 months after transplantation, all remaining breeding males were killed and their testes were analyzed histologically to determine the extent of spermatogenesis.
Results
Stem Cell Number Increases During Postnatal Development.
The first experiment was designed to evaluate the relative stem cell number from donor testes of three different developmental ages, neonate, pup, and adult. Donor testis cells from ZFlacZ transgenic mice express lacZ starting at the round spermatid stage of spermatogenesis and beyond. Importantly, Sertoli cells from ZFlacZ mice do not express the transgene (20). Therefore, if a few colonies of proliferating Sertoli cells from neonate and pup donors were formed, they would not be stained. Testes collected from ZFlacZ donor mice increased in size (Fig. 1A Top) and weight (Fig. 1A Legend) during development from neonate to pup to cryptorchid adult. However, the seminiferous tubules of all these ages were devoid of differentiated germ cells and, therefore, were enriched relatively for spermatogonial stem cells (Fig. 1A Bottom). The weight of testes and the number of cells that could be recovered per testis increased significantly during development (Fig. 1A Legend; P ≤ 0.001).
The results shown in Fig. 1B indicate a significant stepwise increase in stem cell concentration (colonies per 105 cells injected) and colonized area (blue area, mm2) during testis development. The number of functional spermatogonial stem cells per testis (colonies per 105 cells × cells per testis, legends Fig. 1 A and B) increased 39-fold during development (P < 0.005) from neonate (21.3) to pup (136.2) to adult (839.6). Although neonate stem cells produced larger colonies (0.63 ± 0.04 mm2 per colony, mean ± SEM, P < 0.001) than pup (0.44 ± 0.04 mm2 per colony) or adult (0.46 ± 0.02 mm2 per colony), all these colony sizes fell within the range (0.35–0.65 mm2 per colony) found in previous studies (28, 30).
Niche Accessibility in Seminiferous Tubules Decreases During Postnatal Development.
In the second experiment, a two by two factorial design was used in which donor cells from pup or cryptorchid adult ROSA26 testes were transplanted into pup or adult W recipients. The W recipient provides the best model to compare pup to adult recipients because they lack all stages of differentiating germ cells, but they are capable of generating spermatogenesis and offspring from transplanted testis cells (17). Germ cells in wild-type pups cannot be destroyed totally by busulfan treatment; therefore, pup recipients would develop endogenous spermatogenesis and confound the transplantation results. ZFlacZ donor testis cells are not immunologically compatible with the W recipients; therefore, ROSA26 donors were used. In contrast to the neonate, pup donor-testis cells produce few Sertoli cell colonies, and they generally can be distinguished by morphology.
Stem cell-derived spermatogenic colonies from ROSA26 adult donor testes were 2.0 times (107.5 vs. 53.6 colonies per 105 cells) more frequent than those formed from pup donor testes in pup recipients and 2.8 times (13.8 vs. 4.9 colonies per 105 cells) more frequent in adult recipients (Fig. 2A). A similar relationship exists (2.0-fold in pup recipients and 2.74-fold in adult recipients) between the number of spermatogonial stem cells per testis (colonies per 105 cells × cells per testis, Fig. 2A) from adult and pup donors, because the cell recovery for these two donor ages was almost identical in this mouse strain. Blue area formed per colony (a measure of colony size) from ROSA26 pup stem cells was 1.3 times (2.42 vs. 1.88 mm2) larger than that formed from adult donor cells in pup recipient testes and 1.2 times (0.59 vs. 0.50 mm2) larger in adult recipient testes (Fig. 2B). In contrast to these modest differences between donor cell types, host environment (recipient age) had a dramatic impact on colonization by spermatogonial stem cells, regardless of donor age (Fig. 2 A and B). ROSA26 pup donor cells colonized pup testes 10.9 times (53.6 vs. 4.9 colonies per 105 cells) more frequently than adult testes, and adult donor cells colonized pup testes 7.8 times (107.5 vs. 13.8 colonies per 105 cells) more frequently than adult testes. The blue area formed per colony from ROSA26 pup stem cells was 4.1 times (2.42 vs. 0.59 mm2) larger in pup than adult recipients, and the blue area from adult cryptorchid donor cells was 3.8 times (1.88 vs. 0.50 mm2) larger in pup than adult recipients (Fig. 2B). Because the statistical analysis indicated no significant interaction between donor cell type and recipient environment type, the data for donors (pup and adult) and recipients (pup and adult) could be combined to determine the significance of differences. The number of colonies formed from adult cryptorchid donor cells (combining recipient types) was 2.4 times (mean of 2.0 and 2.8) more than that formed from pup cells (P = 0.125); and the blue area per colony from pup donor cells (combining recipient types) was 1.25 times (mean of 1.3 and 1.2) greater than that for adult cryptorchid (P = 0.091). In contrast, the number of colonies formed in pup recipients was 9.4 times (mean of 10.9 and 7.8) that in adults (P = 0.007), and the blue area per colony in pup recipients was 4.0 times (mean of 4.1 and 3.8) that in adults (P < 0.001). Taken together, these data on number of colonies and blue area per colony demonstrate that the pup seminiferous tubule environment provides more colonization foci (niches) and allows greater colony expansion than the adult tubule.
W Pup Recipients Become Fertile from Donor Stem Cells.
Morphological examination of W pup and adult recipient testes 5 months after transplantation of cryptorchid adult ROSA26 donor cells (Fig. 3, Table 1) demonstrated that pup recipient testes had complete spermatogenesis in about 77% of seminiferous tubules. In contrast, the adult recipients had complete spermatogenesis in only 31% of tubules, and 53% of tubules were completely devoid of germ cells. Spermatogenesis in the recipient testes could come only from donor stem cells because the W recipients are incapable of generating spermatogenesis from the defective endogenous stem cells. Four pup and six adult W recipient mice were housed with wild-type B6/SJL females to determine whether fertility would result from transplantation of ROSA26 cryptorchid adult testis cells. Three pup recipients sired offspring within 84 d posttransplantation (Table 1), including two pup recipients that had spermatogenesis in only one testis (Table 1, 4819 and 4870). One pup recipient was found to have a hernia and abnormal testes; therefore, it was excluded from consideration. None of the adult recipient animals were fertile up to the time of analysis, at least 133 d posttransplantation. At the time of sacrifice, ≈5.5 months after transplantation, fertile pup recipient testes were significantly larger (61.1 ± 9.6 mg) than adult recipient testes (25.3 ± 1.4 mg, P < 0.0001), reflecting the greater degree of donor-derived spermatogenesis in pup recipients (Fig. 3, Table 1). The lacZ transgene was transmitted to 44% (41/93, not shown) of the F1 progeny, approximately the percentage predicted by normal Mendelian genetics (50%), because the donor ROSA26 mice are hemizygous for the lacZ transgene.
Figure 3.

Macroscopic and histological comparison of pup and adult W recipient testes after transplantation of adult cryptorchid ROSA26 donor cells. (Top) Donor cells were transplanted into the testes of 5–9-day-old pup W recipients. (Bottom) Donor cells were transplanted into the testes of adult W recipients. Recipient testes stained with X-gal 2 months after transplantation of donor cells are shown (Left). Blue color represents donor-derived spermatogenesis. Testes stained with X-gal ≈5.5 months after donor cell transplantation are shown (Center). Note greater X-gal staining and degree of donor-derived spermatogenesis in pup recipient. (Bar = 2 mm.) Histological sections from recipient testes ≈5.5 months after transplantation are displayed (Right). Insets: Note normal appearance and organization of germ cells and the presence of spermatozoa in the center of a pup seminiferous tubule, and the absence of spermatogenesis in a tubule from an adult. Stain, Nuclear Fast red. (Bar = 50 μm.)
Table 1.
Evaluation of spermatogenesis and fertility in W pup and adult recipient mice
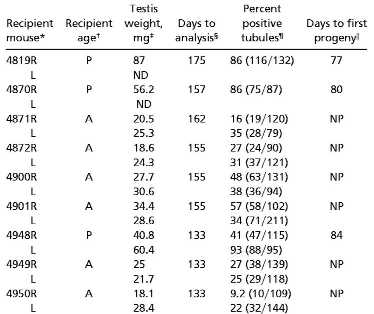 |
R, right testis; L, left testis.
P, pup; A, adult.
Average weight of W pup recipient testes (61.1 ± 9.6 mg) was significantly greater than adult recipient testes (25.3 ± 1.4 mg, P < 0.0001). ND, not determined; 4819L was fibrous and could not be analyzed; 4870R was not injected with donor cells.
Days from transplantation to analysis.
Seminiferous tubule cross sections with sperm heads were considered to be positive for complete spermatogenesis. Pup recipients had complete spermatogenesis in 77% of tubules compared to 31% for adult recipients. Values in parentheses are the number of cross sections with complete spermatogenesis over the total examined.
Days from transplant to first progeny (fetuses or pups); NP, no progeny.
Discussion
Studies with donors of different developmental ages demonstrated a remarkable 39-fold increase in spermatogonial stem cell number in the testis from newborn to adult. The number of stem cells in the adult represents 5 to 6 doublings of the original population in the neonate, and stem cells from a single adult ZFlacZ cryptorchid testis produced 840 colonies. Previous studies have demonstrated that cryptorchid and wild-type testes contain a similar number of stem cells (22), and morphological studies of adult wild-type mouse testes identify about 2 × 104 putative stem cells per testis (31, 32). Assuming 100% harvesting efficiency, only about 4.2% of this number generates colonies of spermatogenesis in the transplantation assay. These data suggest that only 1 in 20 adult stem cells colonize a recipient, which is in agreement with our previous estimates (30). In hematopoiesis, it has been estimated that between 1 in 5 and 1 in 20 available stem cells may function in reconstitution assays (33).
Prospermatogonia of the neonate testis, identified by their morphology and position in the center of the seminiferous tubule, demonstrated an even lower ability than adult stem cells to colonize recipient testes. The neonatal mouse testis contains ≈20,000 prospermatogonia (34). Because 21.3 colonies resulted from the cells of one neonatal testis, only about 0.1% of prospermatogonia produced colonies. Thus, these putative stem cells are much less efficient in a functional assay than adult stem cells. Previous studies have indicated a high level of apoptosis among prospermatogonia and undifferentiated spermatogonia in neonatal testes (35–38), and some of the cells not undergoing apoptosis are thought to develop directly into differentiating spermatogonia (39, 40). On the basis of these earlier studies, it seems that at least 50% of the prospermatogonia are lost (35). Our studies indicate that a very low percentage of the remaining cells can function as stem cells, and that the number of functional stem cells increases about 6.4-fold by 5–12 days PP, and another 6.2-fold in the adult, on a per-testis basis. On a per-cell basis, the number of functional stem cells increases 13.4-fold from neonate to adult (Fig. 1B). This increase is in contrast to hematopoietic tissues where it is considered that long-term reconstituting stem cells are almost 7 times more frequent in fetal liver than in adult bone marrow (33).
Similar to the results of the first experiment, we observed a developmental increase (2.74-fold) in the number of spermatogonial stem cells per testis between pup and adult ROSA26 donors, based on colonization data in adult W recipients. This increase is somewhat less than the 6.2-fold increase observed for ZFlacZ mice, perhaps reflecting donor and recipient differences in these experiments. Spermatogenic colony size (blue area per colony) from ROSA26 pup donors was slightly, but not significantly, larger (1.2-fold) than adult donors. ZFlacZ pup and adult donors also produced colonies of similar size.
Although ROSA26 adult and pup donor cell populations demonstrated a 2.4-fold difference in stem cell concentration (combining data from adult and pup recipients) and a 1.25-fold difference in colony size (blue area per colony), there was a dramatic change in the seminiferous tubule environment between pup and adult W recipients. About 9.4 times as many colonies were found in pup testes as adult testes, and the area per colony was 4.0 times larger in pups. This relationship between pup and adult recipients was true regardless of donor age, demonstrating that there is no advantage to transplanting donor cells into a host of similar age. The observed increase in colony number in pup vs. adult recipients may result from the absence of Sertoli cell junctions before 10–12 days PP (16), allowing the stem cell easier migratory access to the seminiferous tubule basement membrane. The greater area of spermatogenesis per colony may be influenced by the increase in seminiferous tubule length that occurs as the testis becomes larger, as well as niche proliferation, possibly resulting from multiplication of Sertoli cells during maturation and growth of the testis. Other factors facilitating colony formation and spread, such as hormonal and growth factor differences, probably are involved also in producing a unique growth environment in the prepubertal testis (41). In hematopoietic tissues, similar factors must regulate stem cells and their interaction with niche, but the systems may not respond identically. In hematopoiesis, the relationship between stem cells and niche may depend more on developmental age. Hematopoietic reconstitution studies, using W fetuses as recipients, demonstrated that adult bone marrow HSCs continued to express adult-stage-specific genes and had a generally lower capacity for self-renewal than did fetal liver HSCs (42). Moreover, early yolk sac cells were found to reconstitute newborn animals but not adults (43, 44). Although these findings suggest that a unique microenvironment or niche, perhaps related to developmental age, is required for stem cell function in hematopoietic tissues, the results of our second experiment indicate that spermatogonial stem cells are more flexible in their niche requirement.
The superiority of pup compared with adult recipients was confirmed in a remarkable manner when the animals were maintained for a longer period and mated with females. The 3 pup recipients sired progeny in less than 3 months, whereas none of the 6 adult recipients became fertile. The speed as well as the efficiency of offspring production from pup recipients is notable. The production of progeny in less than 3 months is comparable to the age at which control wild-type males first produce offspring. It is remarkable that only a few hundred stem cells, less than 2% of the adult number, were required to produce fertile males when the recipients were W pups. A similarly small number of stem cells from testes of adult Steel mice, which are devoid of differentiating germ cells (17, 22), could produce fertility in about 50% of adult W recipients (17). However, in that study, the weights of recipient testes were significantly lower than observed in the present study, and fertility occurred only after 5 months.
The testis is an ideal model to study stem cell–niche interaction for several reasons. First, a stem cell transplantation technique is available that allows identification of individual colonies arising from single stem cells. Second, the kinetics of colony differentiation can be determined independently for each stem cell (clonal analysis). Third, the microenvironment or niche, including the external hormonal balance, can be manipulated easily. No other self-renewing tissue has such favorable characteristics to permit study of stem cell–niche interaction and the formation of differentiating cell populations. We took advantage of these unique characteristics to address fundamental questions of stem cell biology during development. We demonstrated a dramatic increase in the number of spermatogonial stem cells per testis during postnatal development, but found little change in colony size resulting from stem cells derived from young and adult mice. In the second experiment, a unique recipient (W) was used to allow assessment of donor and recipient age effects. The seminiferous tubule microenvironment of the immature pup was dramatically better than the adult in terms of the number of colonization foci and the expansion of each colony, regardless of donor age.
Acknowledgments
We thank Drs. R. Behringer and E. Sandgren for helpful comments on the manuscript. We appreciate the assistance of C. Freeman and R. Naroznowski with animal maintenance and experimentation, C. Brensinger for statistical assistance, and J. Hayden for help with photography. T.S. was supported by the Japan Society for Promotion of Science. Microscopic sections were produced in the Institute for Human Gene Therapy, Cellular Morphology Core (5-P30-DK-47747-07). Financial support for the research was from the National Institute of Child Health and Human Development Grant 36504, the Commonwealth and General Assembly of Pennsylvania, and the Robert J. Kleberg, Jr. and Helen C. Kleberg Foundation.
Abbreviations
- HSC
hematopoietic stem cell
- PP
postpartum
- X-gal
5-bromo-4-chloro-3-indolyl β-d-galactoside
- ZF
zinc finger
References
- 1.Harrison D E. Blood. 1980;55:77–81. [PubMed] [Google Scholar]
- 2.Till J E, McCulloch E A. Radiat Res. 1961;14:213–222. [PubMed] [Google Scholar]
- 3.Brinster R L, Avarbock M R. Proc Natl Acad Sci USA. 1994;91:11303–11307. doi: 10.1073/pnas.91.24.11303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brinster R L, Zimmermann J W. Proc Natl Acad Sci USA. 1994;91:11298–11302. doi: 10.1073/pnas.91.24.11298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schofield R. Blood Cells. 1978;4:7–25. [PubMed] [Google Scholar]
- 6.Xie T, Spradling A C. Science. 2000;290:328–330. doi: 10.1126/science.290.5490.328. [DOI] [PubMed] [Google Scholar]
- 7.Tran, J., Brenner, T. J. & DiNardo, S. (2 000) Nature (London)407, 754–757. [DOI] [PubMed]
- 8.Kiger A A, White-Cooper H, Fuller M T. Nature (London) 2000;407:750–754. doi: 10.1038/35037606. [DOI] [PubMed] [Google Scholar]
- 9.Williams E D, Lowes A P, Williams D, Williams G T. Am J Pathol. 1992;141:773–776. [PMC free article] [PubMed] [Google Scholar]
- 10.Bjornson C R, Rietze R L, Reynolds B A, Magli M C, Vescovi A L. Science. 1999;283:534–537. doi: 10.1126/science.283.5401.534. [DOI] [PubMed] [Google Scholar]
- 11.Dexter T M, Allen T D, Lajtha L G. J Cell Physiol. 1977;91:335–344. doi: 10.1002/jcp.1040910303. [DOI] [PubMed] [Google Scholar]
- 12.Dexter T M, Spooncer E. Annu Rev Cell Biol. 1987;3:423–441. doi: 10.1146/annurev.cb.03.110187.002231. [DOI] [PubMed] [Google Scholar]
- 13.Clermont I, Perey B. Am J Anat. 1957;100:241–260. doi: 10.1002/aja.1001000205. [DOI] [PubMed] [Google Scholar]
- 14.Nagano M, Brinster R L. APMIS. 1998;106:47–57. doi: 10.1111/j.1699-0463.1998.tb01318.x. [DOI] [PubMed] [Google Scholar]
- 15.McCarrey J R. In: Cell and Molecular Biology of the Testis. Desjardins C, Ewing L L, editors. New York: Oxford Univ. Press; 1993. pp. 58–89. [Google Scholar]
- 16.de Kretser D M, Kerr J B. In: The Physiology of Reproduction. Knobil E, Neill J D, editors. New York: Raven; 1994. pp. 1177–1290. [Google Scholar]
- 17.Ogawa T, Dobrinski I, Avarbock M R, Brinster R L. Nat Med. 2000;6:29–34. doi: 10.1038/71496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dobrinski I, Ogawa T, Avarbock M R, Brinster R L. Mol Reprod Dev. 1999;53:142–148. doi: 10.1002/(SICI)1098-2795(199906)53:2<142::AID-MRD3>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- 19.Nagano M, Avarbock M R, Brinster R L. Biol Reprod. 1999;60:1429–1436. doi: 10.1095/biolreprod60.6.1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zambrowicz B P, Zimmermann J W, Harendza C J, Simpson E M, Page D C, Brinster R L, Palmiter R D. Development (Cambridge, UK) 1994;120:1549–1559. doi: 10.1242/dev.120.6.1549. [DOI] [PubMed] [Google Scholar]
- 21.Zambrowicz B P, Imamoto A, Fiering S, Herzenberg L A, Kerr W G, Soriano P. Proc Natl Acad Sci USA. 1997;94:3789–3794. doi: 10.1073/pnas.94.8.3789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shinohara T, Avarbock M R, Brinster R L. Dev Biol. 2000;220:401–411. doi: 10.1006/dbio.2000.9655. [DOI] [PubMed] [Google Scholar]
- 23.Bellvé A R, Cavicchia J C, Millette C F, O'Brien D A, Bhatnagar Y M, Dym M. J Cell Biol. 1977;74:68–85. doi: 10.1083/jcb.74.1.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ogawa T, Aréghaga J M, Avarbock M R, Brinster R L. Int J Dev Biol. 1997;41:111–122. [PubMed] [Google Scholar]
- 25.Silvers W K. The Coat Colors of Mice. New York: Springer; 1979. pp. 206–241. [Google Scholar]
- 26.Geissler E N, Ryan M A, Housman D E. Cell. 1988;55:185–192. doi: 10.1016/0092-8674(88)90020-7. [DOI] [PubMed] [Google Scholar]
- 27.Suckow M A, Danneman P, Brayton C. The Laboratory Mouse. Boca Raton, FL: Cancer Research Campaign; 2001. [Google Scholar]
- 28.Shinohara T, Avarbock M R, Brinster R L. Proc Natl Acad Sci USA. 1999;96:5504–5509. doi: 10.1073/pnas.96.10.5504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zeger S L, Liang K Y. Biometrics. 1986;42:121–130. [PubMed] [Google Scholar]
- 30.Shinohara T, Orwig K E, Avarbock M R, Brinster R L. Proc Natl Acad Sci USA. 2000;97:8346–8351. doi: 10.1073/pnas.97.15.8346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Meistrich M L, van Beek M E A B. In: Cell and Molecular Biology of the Testis. Desjardins C, Ewing L L, editors. New York: Oxford Univ. Press; 1993. pp. 266–295. [Google Scholar]
- 32.Tegelenbosch R A J, de Rooij D G. Mutat Res. 1993;290:193–200. doi: 10.1016/0027-5107(93)90159-d. [DOI] [PubMed] [Google Scholar]
- 33.Morrison S J, Hemmati H D, Wandycz A M, Weissman I L. Proc Natl Acad Sci USA. 1995;92:10302–10306. doi: 10.1073/pnas.92.22.10302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Vergouwen R P, Huiskamp R, Bas R J, Roepers-Gajadien H L, Davids J A, de Rooij D G. J Reprod Fertil. 1993;99:479–485. doi: 10.1530/jrf.0.0990479. [DOI] [PubMed] [Google Scholar]
- 35.Beaumont H M, Mandl A M. J Embryol Exp Morph. 1963;11:715–740. [PubMed] [Google Scholar]
- 36.Huckins C, Clermont Y. Arch Anat Histol Embryol. 1968;51:341–354. [PubMed] [Google Scholar]
- 37.Roosen-Runge E C, Leik J. Am J Anat. 1968;122:275–299. doi: 10.1002/aja.1001220208. [DOI] [PubMed] [Google Scholar]
- 38.Wang R A, Nakane P K, Koji T. Biol Reprod. 1998;58:1250–1256. doi: 10.1095/biolreprod58.5.1250. [DOI] [PubMed] [Google Scholar]
- 39.Kluin P M, de Rooij D G. Int J Androl. 1981;4:475–493. doi: 10.1111/j.1365-2605.1981.tb00732.x. [DOI] [PubMed] [Google Scholar]
- 40.de Rooij D G. Int J Exp Pathol. 1998;79:67–80. doi: 10.1046/j.1365-2613.1998.t01-1-00057.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Orth J M. In: Cell and Molecular Biology of the Testis. Desjardins C, Ewing L L, editors. New York: Oxford Univ. Press; 1993. pp. 3–42. [Google Scholar]
- 42.Fleischman R A, Mintz B. J Exp Med. 1984;159:731–745. doi: 10.1084/jem.159.3.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Müller A M, Medvinsky A, Strouboulis J, Grosveld F, Dzierzak E. Immunity. 1994;1:291–301. doi: 10.1016/1074-7613(94)90081-7. [DOI] [PubMed] [Google Scholar]
- 44.Yoder M C, Hiatt K, Mukherjee P. Proc Natl Acad Sci USA. 1997;94:6776–6780. doi: 10.1073/pnas.94.13.6776. [DOI] [PMC free article] [PubMed] [Google Scholar]