

Abstract
We show that replication defective adenovirus can be used for localized overexpression of a chosen gene in Xenopus tadpoles. Xenopus contains two homologs of the Coxsackie and Adenovirus Receptor (xCAR1 and 2), both of which can confer sensitivity for adenovirus infection. xCAR1 mRNA is present from the late gastrula stage and xCAR2 throughout development, both being widely expressed in the embryo and tadpole. Consistent with the expression of the receptors, adenovirus will infect a wide range of Xenopus tissues cultured in vitro. It will also infect early embryos when injected into the blastocoel or archenteron cavities. Furthermore adenovirus can be delivered by localized injection to tadpoles and will infect a patch of cells around the injection site. The expression of GFP in infected cells persists for several weeks. This new gene delivery method complements the others that are already available.
Keywords: adenovirus, Xenopus, green fluorescent protein, Coxsackie and Adenovirus Receptor (CAR)
Introduction
Methods for introducing new genes or gene products into organisms represent an extremely valuable technology in developmental biology. Such methods, generally referred to as “overexpression”, or “ectopic expression”, can be used to establish the biological activity of gene products in vivo. They can also often be used to bring about ablation of a specific gene product, for example by expressing dominant negative versions of gene products (Lagna and Hemmati-Brivanlou, 1998) or RNAi constructs (Li and Rohrer, 2006). To enable these techniques to be used to their maximum effect it should be possible to overexpress a specific gene at any desired position and any desired stage of a developing organism. Overexpression techniques of various types exist for all of the main laboratory organisms for developmental biology research, although for each there are limitations on what is possible (Slack, 2005). Xenopus is one of these systems (Sive et al., 2000). In the past the main overexpression technique used has been the injection of synthetic mRNA into fertilised eggs or early cleavage stage blastomeres (Krieg and Melton, 1984). This has proved an extraordinarily valuable method and has contributed to countless studies of early developmental mechanisms in the last 20 years. But there are two limitations. Firstly, the mRNA starts being translated from the time of its injection and its effects on early developmental stages may compromise the embryo at later stages, although this can sometimes be overcome by using a fusion protein of the gene product of interest with the hormone binding domain of a nuclear hormone receptor (Kolm and Sive, 1995). Secondly, the mRNA has a limited lifetime and it will decay over a period of some hours or days, depending on its intrinsic stability to nuclease digestion. Since 1996, RNA based overexpression has been joined by some effective methods of transgenesis which enable DNA to be integrated into the entire organism. The original Kroll-Amaya method involves introduction of DNA into sperm, while the more recently published ones introduce DNA into fertilized eggs (Kroll and Amaya, 1996; Amaya, 1999; Allen and Weeks, 2005; Ogino et al., 2006; Pan et al., 2006). Transgenesis in Xenopus has overcome the limitations of mRNA stability and has opened up the phases of organogenesis, regeneration and metamorphosis to further study by specific gene overexpression. However, the preparation of transgenics is still demanding and to get long term survival of transgenic tadpoles requires very good egg quality and considerable skill on the part of the experimenter.
For many applications related to the study of late development it is required to introduce a gene or gene product just into a specific location at a specific time. Here we examine the feasibility of using adenoviral vector as a gene delivery system. Adenovirus is a nonenveloped, double-stranded DNA virus with an icosahedral coat composed mainly of three proteins called hexon, penton, and fiber. From the parent virus have been derived several generations of gene therapy vectors which have been widely used in mammalian systems and to some extent in human clinical trials (Gardlik et al., 2005; Verma and Weitzman, 2005). Adenovirus can infect either dividing or non-dividing cells. It introduces double stranded DNA into the infected cell which is expressed for some days or even weeks, but does not become integrated into the genome. The form of virus used for gene introduction is infectious but cannot replicate in the host cells because essential functions have been deleted from its genome. This is produced using a packaging cell line which contains the missing genes and can complement the defect, allowing for the production and release of active virus particles. The most commonly used adenoviral vector is human adenovirus serotype 5, which is rendered replication-defective by the deletion of the E1 and E3 genes. The E1 gene is essential for the assembly of infectious virus particles and is supplied by the packaging cell line. The E3 gene encodes proteins involved in evading host immunity and is dispensable. In addition to rendering the virus incapable of replicating itself, these deletions also create space for an insert of up to 7.5 kb. It has already been shown that adenovirus can infect Xenopus and introduce functional genes (Kawakami et al., 2006), but this study was in the context of many other experiments on several model organisms, and limited technical and background information was given for the use of adenovirus in Xenopus.
We show here that adenoviral vector is able to infect Xenopus and to express its transgene for a prolonged period in the host. It can be administered by localized injection and cause overexpression of the gene of interest in a small patch of tissue. For infection of tadpoles a temperature increase is needed during the infection phase but not thereafter. Despite the lack of integration into the nuclear genome, gene expression continues for several weeks following the initial infection. We believe that this technique will complement the existing methods of gene overexpression, such as RNA injection, transgenesis and electroporation, and will prove to be a convenient route for studying the effects of localized overexpression of tissue specific genes in Xenopus
Results
Identification of Two Xenopus Adenovirus Receptors
Initially we did not know whether adenovirus could infect amphibian cells. The normal mechanism of infection for mammalian cells involves binding of virus to the Coxsackie and Adenovirus Receptor (CAR) an immunoglobulin superfamily member that mediates homotypic cell adhesion and is located in epithelial tight junctions, although it is also present on non-epithelial cell types (Bergelson et al., 1997; Philipson and Pettersson, 2004). The existence of a zebrafish homolog of CAR (Petrella et al., 2002) made it likely that Xenopus would also contain one. Bioinformatic searching led to the identification of two CAR homologues now called xCAR1 and xCAR2. xCAR1 was identified from a cDNA clone derived from a stage 24 embryo EST library and was sequenced in full. The coding region was then amplified by PCR and was cloned into the expression vector pcDNA3.0. The other CAR homolog, xCAR2, was identified by blast sequence homology to human CAR from the NCBI Xenopus EST database. The coding sequence region was amplified by PCR from whole stage 20 embryo cDNA and cloned into pcDNA3.0.
Nucleotide sequence analysis and amino acid prediction were performed using the standard bioinformatics tool set through the NCBI database. Amino acid alignment of both xCAR1 and xCAR2 with other vertebrates demonstrated low level sequence homology across species with an overall identity of xCAR1 to human CAR of 38% and of xCAR2 of 53% at the protein level (Figure 1A). This relatively low identity would not in itself imply that the proteins function as adenoviral receptors. However structural studies have shown that the virus binds via its fiber protein to the N-terminal domain of the CAR (Bewley et al., 1999). Fig.1B depicts the contact surface of the CARs deduced from structural analysis of the human molecule, with Xenopus residues shown color coded for similarity to the human. In this critical region amino acid conservation of xCAR1 (66%) and xCAR2 (70%) is very high and it therefore seemed likely that both of the xCAR molecules would be able to support infection of Xenopus cells by adenovirus.
Figure 1. Sequence analysis.

A. Predicted amino acid sequences of the Xenopus laevis Coxsackievirus and Adenovirus Receptors (xCAR1 and 2). The xCAR sequences are shown aligned with CAR proteins from mouse, human and zebrafish. The shaded residues are identical in all these sequences.
B. Diagram of the binding site for virus on the CAR surface, based on the structure of hCAR in (He et al., 2001). The numbering is for the Xenopus sequences and the color depicts the relatedness to the human sequence: bright green identical, yellow conserved, brown unrelated. (structural biology method adapted from (Petrella et al., 2002)).
In order to confirm the predicted biological activity of the two receptors, we transiently transfected the full length xCAR1 and xCAR2 into CHO cells, which have no CAR of their own (Graham and Prevec, 1992), and then tested for infectability with adeno-GFP at various multiplicities of infection. Green cells became visible in all the wells transfected with xCAR1 and xCAR2, but not in controls, at a frequency similar to the overall transfection frequency, indicating that both xCARs can support virus entry to cells (Fig.2). Analysis of the cells by flow cytometry showed that the transfection efficiency is about 5% and that essentially all of the cells transfected with xCARs became infected by the adeno- GFP.
Figure 2. Evidence that xCARs function as adenovirus receptors.

The lower panels show GFP fluorescence in the cells, the upper show flow cytometry of the above-threshold cells by GFP intensity (vertical axis) and cell size (horizontal axis). The number of GFP positive cells in the presence of xCAR is similar to the transfection efficiency (about 5%), indicating that all or most cells transfected with xCAR become infected. All cell nuclei are stained blue with DAPI.
A. CHO cells treated with lipofectamine alone and infected with adeno-GFP (negative control).
B. CHO cells transfected with pcDNA3-GFP (transfection control).
C. CHO cells transfected with xCAR1, and then infected with adeno-GFP.
D. CHO cells transfected with xCAR2, and then infected with adeno-GFP. Scale bars 5 μm.
An expression analysis of xCAR1 and xCAR2 was conducted by RT-PCR (Fig.3A). This showed that xCAR1 mRNA is expressed from mid-gastrulation onwards, while xCAR2 mRNA is present at all stages, presumably therefore being both maternal and zygotic. Tissues dissected from stage 54-56 tadpoles all express xCAR1 and xCAR2, suggesting that susceptibility to adenovirus expression is widespread (Fig.3B). Wholemount in situ hybridization of embryos also confirmed widespread expression (Fig.3C-F). The pattern seen is typical of ubiquitously expressed genes, although there is some minor additional expression of xCAR2 in the pronephros and in the neural part of the tail bud.
Figure 3. Expression of xCAR mRNA in embryonic and tadpole stages.

A. RT-PCR of xCAR1 and xCAR2 in mRNA extracted from whole embryos at the developmental stages indicated. xCAR1 is expressed from gastrulation onwards, while xCAR2 is expressed both maternally and zygotically.
B. RT-PCR of xCAR1 and xCAR2 in mRNA extracted from stage 54-56 tadpole tissues. M muscle, Li Liver, H head, S skin.
C-H. Wholemount in situ hybridizations. C, D show no probe controls at stage 25 and 31; E, F show xCAR1 at stage 25 and 31; G, H show xCAR2 at stage 25 and 31. White arrowheads in G and H indicate expression of xCAR2 in dorsal tailbud and in pronephros and nephric duct, which are greater than seen for xCAR1.
Infection of In Vitro Cultures
Because in vivo administration of gene therapy vectors is often limited by access to the virus receptors on cell surfaces, we first tested for infectability of Xenopus cells in vitro. The cells were grown from explanted tissues cultured in L15 medium, either on fibronectin coated plastic or on Millipore filters. We found that a wide range of tissues could be infected. The GFP expression became visible from about the 3rd day and persisted for at least 7 days after this. In Fig.4 is shown infection of cell types characteristic of all three germ layers, including cells with morphology corresponding to neurons, fibroblasts and epithelial cells. The time to appearance of the GFP is about twice what would be expected in mammalian cells, a difference probably attributable to the lower temperature used for the Xenopus cultures (28°C instead of 37°C).
Figure 4. Adeno-GFP infection of Xenopus cells grown in-vitro.
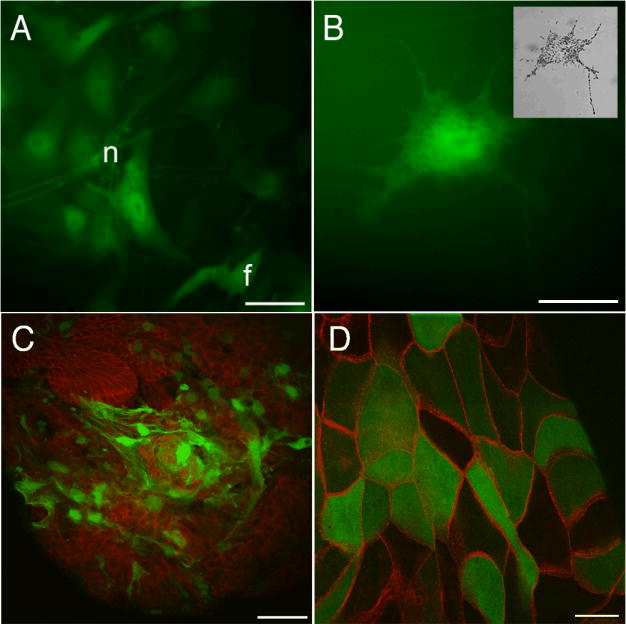
A. Fibroblasts (f) and neuron-like cells (n) in outgrowths from explants of stage 40-44 tadpole neural tubes.
B. A melanophore migrating from stage 40-44 tadpole neural tube explant (brightfield view in insert).
C. Stage 45 tadpole liver explant. Mesenchymal cells express GFP while the underlying epithelium, stained for E-cadherin (red), does not.
D. Stage 45 gall bladder. E-cadherin is shown in red.
Scale bars are 20 μm in A, 30 μm in B, 50 μm in C and 20 μm in D.
Infection of Embryos and Tadpoles
Initial in vivo experiments were conducted by injecting the virus into the blastocoel or archenteron of early stage embryos. The injections were not associated with any noticeable mortality or morbidity and GFP fluorescence was seen to develop about 3 days later. It was present in many tissues at many positions within the later tadpoles (Fig.5). The fluorescence persisted for at least 10 days without diminution of intensity. In Table 1 is shown the percentages of tadpoles labeled in different tissues following injection at different stages. High proportions of muscle labeling arise from injection of stage 8 blastulae, high proportions of intestinal labeling from injection of gastrulae, and overall infection at early neurula stage seemed less effective. These differences probably reflect the accessibility of the prospective regions for the tissues in question rather than any major difference in the susceptibility of different tissues. In the blastula stage many cells lining the blastocoel will later contribute to trunk and tail muscles (Dale and Slack, 1987). In the gastrula, the cells lining the archenteron will later end up in the intestine (Chalmers and Slack, 2000). In the neurula stage the archenteron cavity is connected to the exterior by a patent blastopore, so some of the injected virus may leak out, reducing the effective dose. In these experiments no labeling was seen in the central nervous system. This is probably due to limited access at the stages of injection as the in vitro experiments indicate that neurons can be infected.
Figure 5. Adeno-GFP infection of Xenopus early embryos.

Embryos were injected with adeno-GFP at the indicated early stage, raised to tadpoles, and photographed for GFP fluorescence at stage 45.
A. Trunk and tail muscle, injected stage 8.
B. Scattered cells in liver, injected stage 8.
C. Small intestine loop, injected stage 14. Bright patch on left is autofluorescence of gall bladder.
D. Jaw muscles, injected stage 10.5.
E. Confocal view of flattened small intestine epithelium, injected stage 14.
F. Transverse confocal view of GFP (green) and smooth muscle actin (red) from small intestine, injected stage 14.
G. RT-PCR of GFP mRNA. Samples are from embryos injected with GFP RNA at 2 cells (top row); embryos injected with adeno-GFP as blastulae (second row); control embryos (third row); loading control (bottom row).
Scale bars indicate 100μm in A-D and 50μm in E-F
Table 1. % of tadpoles at stage 45 showing GFP in particular tissues following injection of early stage embryos with adeno-GFP.
| Stage injected | Jaw muscle | Brain | Heart | Liver | Stomach | Small intestine | Large intestine | Trunk/tail muscle | No GFP |
|---|---|---|---|---|---|---|---|---|---|
| Stage 8 (n=30) | 6.7 | 6.7 | 30 | 13.3 | 0 | 16.7 | 0 | 90 | 3.3 |
| Stage 10.5 (n=108) | 8.3 | 0.9 | 6.5 | 3.3 | 0 | 73.5 | 1.9 | 12.0 | 16.6 |
| Stage 12 (n= 73) | 0 | 0 | 1.4 | 0 | 0 | 80.1 | 5.4 | 5.4 | 11 |
| Stage 14 (n = 74) | 8.5 | 7 | 2.8 | 0 | 12.7 | 19.7 | 9.9 | 11.3 | 74.6 |
To find whether the long lifetime of the fluorescence was due to persistence of active virus or simply to the stability of the GFP protein, we compared the persistence over 10 days of GFP RNA and fluorescence following either injection of RNA to cleaving embryos, or injection of adeno-GFP to blastulae (Table 2 and Fig.5 G). The RNA injections show, as expected, that the RNA is mostly gone by 3d, while the GFP fluorescence persists in over half the embryos for 7d. By contrast the virus injections show that both RNA and protein persist at a high level for at least 10d. This shows that the virus remains active for at least this period, and probably for much longer.
Table 2.
Longevity of GFP RNA and protein: comparison of injected RNA and injected adenovirus. % indicates percentage of embryos in the sample.
-,+,++,+++,++++ indicate visual scores of GFP fluorescence in positive cases, and of RNA band intensity.
| Days post-injection | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 10 | |||||||||||
| Reagent | n | Stage injected | RNA | %GFP+ | GFP intensity | RNA | %GFP+ | GFP intensity | RNA | %GFP+ | GFP intensity | RNA | %GFP+ | GFP intensity |
| Water | 60 | 2 cell intracellular | - | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - |
| GFP RNA | 120 | 2 cell intracellular | ++++ | 96 | ++++ | + | 95 | +++ | - | 68 | ++ | - | 28 | + |
| Ad-GFP | 120 | Stage 8 blastocoel | - | 0 | - | ++ | 92 | +++ | ++ | 91 | +++ | ++ | 89 | +++ |
Initial trials of localized injections into tadpoles gave no GFP expression at all. After some trial and error, success was obtained by keeping the tadpoles at an elevated temperature of 34°C overnight (16 hours), immediately following the virus injection. Although adenovirus usually functions at 37°C, we know that virus infection can occur at lower temperatures as shown by the early embryo experiments. The successful infection of tadpoles may require the higher temperature both to facilitate the virus-cell interaction and to expose the xCAR on cell surfaces. The overnight elevated temperature did not cause any deaths or obvious abnormalities among the tadpoles, nor was there any visible toxicity or damage arising from the presence of the virus at the injection site. Fig.6 and Table 3 show localized injection of both limb buds and of the tail. The muscle fibers are obviously infected as shown by the wholemount view. The sections of the limbs show that connective tissue cells are also infected although the cartilages are not. This may be because of access problems as the cartilages are surrounded by dense connective tissue sheaths. Following these localized injections into tadpoles the GFP typically takes about 5 days to appear, and it then persists for at least 4 weeks.
Figure 6. Adeno-GFP infection of Xenopus tadpoles at stage 54-56.

A. GFP fluorescence visible in muscle and limb five days after injection (muscle and limb injected separately).
B. Higher power of another specimen showing GFP in a limb (outlined).
C. Distribution of GFP-expressing cells in the limb shown in B by anti-GFP antibody (green) and DAPI (blue) staining.
D. Tissue distribution of positive cells shown by staining with anti-GFP antibody (green) and 12/101 (anti-muscle antibody, red). Some GFP positive cells are muscle fibers and others are connective tissue cells.
Scale bars: 500 μm.
Table 3.
Percentage of tadpoles positive for GFP expression in the specified position following injection with adeno-GFP at stage 54-56.
| Injection site | Number of injected tadpoles | Number of GFP expressing tadpoles |
|---|---|---|
| tail muscle | 27 | 25 (92.6%) |
| limb | 27 | 11 (40.7%) |
Discussion
Although mostly associated in the past with studies of early development, Xenopus has recently also become a useful model organism for investigations of organogenesis, regeneration and metamorphosis (Sive et al., 2000; Brown and Cai, 2007; Slack et al., 2008). This means that there will be an increasing demand for methods of gene introduction at specific times and positions chosen by the experimenter. Injection of RNA or antisense morpholinos into specific blastomeres of early embryos have proved extremely useful for the study of early development, but inevitably the injected materials decay, and after 3 or more days of development their limited persistence becomes an issue. At present the methods available for localized overexpression are the use of specific promoters for transgenesis, the performance of grafts of transgenic tissue to wild type hosts, and electroporation.
Spatial control of transgene expression can be achieved by the use of tissue specific promoters, which need not be of Xenopus origin (Beck and Slack, 1999). Temporal control can be achieved by the use of the hsp70 promoter, which becomes active at 34°C and provides a tight regulation between active and inactive states (Wheeler et al., 2000). But it has not so far proved possible to combine these techniques to achieve tissue specific induction at a desired time. Some studies have shown the feasibility of binary transgenic techniques previously used in other model organisms. The Cre-lox system in its gene-activating mode has been adapted from mice (Werdien et al., 2001; Ryffel et al., 2003; Gargioli and Slack, 2004; Waldner et al., 2006). Two constructs need to be introduced. One carries the gene for the Cre recombinase driven by a promoter of choice. The other carries the gene of interest whose expression will be activated by Cre activity. The Gal4 system has been adapted from its original use in Drosophila (Brand and Perrimon, 1993) to Xenopus (Chae et al., 2002; Hartley et al., 2002). Again, two constructs are introduced, a specific promoter driving production of the yeast transcription factor Gal4, and the gene of interest driven by the specific upstream activating sequence (UAS) which is recognized and bound by Gal4. Either of these systems can be enhanced by the use of hormone inducible forms of the Cre (Hayashi and McMahon, 2002) or Gal4 (Chae et al., 2002) which confers inducibility by the addition of the appropriate hormone. The fusion protein is sequestered in the cytoplasm by endogenous heat shock proteins and thereby kept in an inactive configuration. When the hormone is added to the embryos it binds to the receptor, liberates the fusion protein from the complex, and enables it to perform its function (Kolm and Sive, 1995). The tetracycline system (Gossen and Bujard, 1992; Urlinger et al., 2000) has also been applied to Xenopus (Das and Brown, 2004). The version used is the Tet-on system where addition of doxycycline activates transgene expression. All three of these binary systems show promise, but they do not necessarily function well with new gene combinations, and further development work is needed before they become generally accepted.
Spatial regulation of overexpression can also be achieved by using grafts from donors that are transgenic for ubiquitously expressed transgenes, or those driven by the hsp70 promoter. This has been used for the study of cell lineage in tail regeneration (Gargioli and Slack, 2004; Lin et al., 2007) but there are limits on what is possible. Because of the physical properties of the tissues, it is much easier to make grafts at embryonic than at tadpole stages, and so the shape and size of the final genetically modified domain is not easy to control.
The transgenic methods are all demanding because they require complex interventions at early embryonic stages, followed by successful cultivation of tadpoles to the required stage for the experiments. For this reason, electroporation of plasmid DNA has been used in some studies, mostly involving the central nervous system (Eide et al., 2000; Haas et al., 2002; Echeverri and Tanaka, 2003; Falk et al., 2007). Like virus infection this has the potential advantage that it can be carried out on normal tadpoles that are taken from a stock population and have had no previous treatments. But a recognised problem with electroporation is that it can cause instability of cell differentiation state (Atkinson et al., 2006). Also we have noticed a tendency for tadpoles to die of stopped heart when the electrodes are in the vicinity of the heart.
Given the problems with electroporation, we think that there is also a role for the use of viral vectors in overexpression experiments. Adenovirus is easy to prepare and can be stored at -80°C before use. Its administration requires no equipment other than that already present in Xenopus laboratories. We have noticed that the development of GFP fluorescence is quite slow following virus infection. This probably indicates introduction of a relatively small number of DNA molecules per cell, and suggests that the virus method is a benign one that does not cause damage to infected cells. In fact we have not noticed any ill effects or deaths among virus-infected tadpoles. Despite the non-integration into the genome the duration of adenoviral mediated overexpression is very long and is likely to be long enough for any experiment on Xenopus tadpoles. If integration into genomic DNA were essential for a particular experiment then adenovirus would not be suitable and an integrating viral vector such as lentivirus would be necessary. So far there are only a few studies on the use of lentivirus in Xenopus, and they are all in vitro rather than on whole embryos or tadpoles (Gatlin et al., 2001; Sugiyama et al., 2005). But these studies do show that Xenopus cells can be infected and suggest that lentivirus may also prove useful for localized overexpression at late stages.
Until a few years ago the preparation and use of viral gene therapy vectors such as adeno- or lentivirus, was a specialized activity that had to be carried out by virology labs. Today however there are kits available to construct and expand the viruses, and their use requires just normal molecular and cell biology skills. There are also companies that will perform the construction and amplification of adenovirus for a fee. These technical advances mean that this new method of specific gene overexpression should be readily available to developmental biology labs in future.
Experimental Procedures
Virus Preparation
A first-generation, replication-defective, recombinant adenoviral vector was used in these experiments. Ad-RSV-GFP (here called adeno-GFP) was propagated in the E1-containing human embryonic kidney (HEK) cell line 293 (Graham and Prevec, 1992). HEK 293 cells were maintained in Dulbecco's Modified Eagle Medium (Invitrogen) supplemented with 10% heat-inactivated fetal calf serum (Sigma). The virus was grown to high titer, released from the cells by rapidly freeze/thawing three times and then purified twice by cesium chloride density ultra-centrifugation. The buffer was exchanged with 10mM Tris.HCl pH 7.5, 1mM MgCl2 on a PD-10 Sephadex column (Amersham Biosciences), before the virus was passed through a sterile 0.22μm Millex filter (Millipore) and frozen in 10μl aliquots using dry ice. The viral stocks were stored at -80°C. The titer of the vector was 3×1010 infectious units/ml, which was calculated using the Adeno-X™ Rapid Titer Kit (Stratagene). Virus was prepared and used under BSL2 safety conditions.
xCAR Cloning
Two homologues (xCAR1 and xCAR2) of the human coxsackie and adenovirus receptor were identified as cDNA clones from NCBI gene database with accession number AW782510 and AW638467, respectively. Clone AW782510 was obtained from the RZPD (Deutsches Ressourcenzentrum für Genomforschung) and sequenced in full. A 1158 bp xCAR1 coding sequence was amplified by PCR using Vent polymerase (New England Biolabs) using 5′primer (5′-CGAAGATGGAAGAACGGAGGATCTC-3′) and 3′primer (5′-CGCTCGAGCCTGGATCTAAGGACTATC -3′). The entire 1044 bp coding sequence for xCAR2 was amplified from whole stage 20 embryo cDNA using the 5′primer (5′-CCCAAGCTTTACACCCGGAGCTTAGCACT-3′) and the 3′primer (5′- CCGCTCGAGGGTCAACGAGGCACTTTCAT -3′). The resulting PCR fragments were digested with HindIII and XhoI and inserted into the mammalian expression vector pcDNA3 (Invitrogen) and confirmed by sequencing.
CHO Cell Culture and Adeno-GFP Infection
CHO-K-1 cells were purchased from the ATCC and grown in DMEM supplemented with 10% FBS, penicillin and streptomycin at 37°C in 5% CO2. Cells were plated at 4×105 cells per well of a 12 well plate and transfected with xCAR1 or xCAR2 using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturers instructions, using 2μg plasmid DNA per well. Adeno-GFP was added to the medium 24 hours after transfection at a multiplicity of about 25 virus particles per cell, and incubated for 12 hours. GFP positive cells were examined and photographed 72 hours after virus infection. To quantify the result, the cell populations were analyzed by flow cytometry using a FACS Calibur. Cells were stained with Propidium Iodide (PI) and the threshold GFP fluorescence was determined for the untreated CHO cells. Then cells were counted for low PI (to confirm viability) and GFP fluorescent intensity.
Embryos and Tadpoles
Xenopus laevis embryos were obtained by in vitro fertilization and staged according to the Nieuwkoop and Faber (NF) tables (Nieuwkoop and Faber, 1967). Embryos were dejellied with 2% cysteine (Sigma), pH 7.8, and then cultured in 0.1×NAM. From stage 46, they were fed on tadpole diet (NASCO).
Early stage Xenopus embryos were arranged in a slit cut in a thin layer of agar in a 90mm petri dish and all medium was removed. Adeno-GFP + 1% phenol red was front loaded into glass microinjection needles using a Nanoject II microinjector (Drummond) and known volumes injected directly into individual embryos. Capped nGFP mRNA, used for controls on mRNA and protein stability, was in vitro transcribed using T7 mMessage mMachine kit (Ambion) according to the manufacturer's instructions.
Late stage tadpoles (around stage 54 to stage 56) were anesthetized in 0.02% MS222 and kept in the anesthetic solution during adenovirus injection. Adeno-GFP was injected into the tail muscle (approx. 1 μl) and limbs (9.2 nl) of each tadpole using a Nanoject injector. After the injection, the tadpoles were incubated in 0.1× MMR at 34°C for 16 hours and then transferred to a 25 °C incubator.
In-vitro Cell and Explant Organ Culture
Embryonic X laevis neural tube explants were performed by dissecting the neural tube from embryos prior to neural crest migration and plating them onto fibronectin coated plastic dishes. Endodermal organ cultures were performed following dissection of the relevant structures from tadpoles, washing them extensively in 70% L15 media supplemented with gentamycin (2.5% of Signma stock) before placing them on permeable CM membrane culture filters (Millipore) in dishes of culture media. The culture conditions were 70% L15 media supplemented with 5% fetal bovine serum, 1% penicillin/streptomycin, 1% amphotericin and 2.5% gentamycin. The explants were grown at 28°C in air and the medium was changed every other day. Adeno-GFP infection was performed by adding virus diluted in culture medium directly to the organs in culture.
Gene Expression Analysis
RNA was isolated from stage 2 to 45 whole embryos or from dissected stage 54/56 Xenopus organs using TriReagent (Sigma) according to the manufacturer's instructions. 1 μg DNAseI treated total RNA was reverse transcribed using SuperScriptII (Invitrogen). PCR was performed with primers specific for xCAR1 [RSD1061 5′- GAGGCGATTGTCATTCAGT -3′and RSD1062 5′- CCCATTTGTAACTGAGAG -3′(Tm 56)], xCAR2 [RSD1059 5′- GTCTCCAATGACCCCAAGAC -3′ and RSD1060 5′- CACCTCGTTTTTGCTGGTTT-3′(Tm 58)], and xEf1α [RSD1065 5′- CAGATTGGTGCTGGATAT -3′ and RSD1066 5′- ACTGCCTTGATGACTCCTAG-3′ (Tm60)] for 30 cycles.
Whole-mount in situ hybridization was performed according to the standard protocol (Harland, 1991). Both pcDNA3-xCAR1 and pcDNA3-xCAR2 were linearized with HindIII and transcribed in the presence of DIG-UTP with SP6 RNA polymerase to produce full length DIG labeled antisense probes followed by hydrolysis to increase probe penetration. Embryos were incubated overnight at 58°C in 1μg of probe. Probes were visualized using an anti-DIG-AP Fab fragment antibody (Roche;1/2000) incubated overnight at 4°C and developed with BM purple reagent (Roche).
The stability of GFP transcripts was determined by RT-PCR on mRNA obtained from stage 2 GFP-mRNA (1ng) injected or stage 8 blastocoel Ad-GFP (2.3nl) injected embryos at 1, 3, 7 and 10 days post injection. RNA was isolated from whole embryos or tadpoles using TriReagent (Sigma) according to the manufacturer's instructions. 1 μg DNAseI treated total RNA was reverse transcribed using SuperScriptII (Invitrogen). PCR was performed with primers specific for GFP [GFP-C 5′- CATGGTCCTGCTGGAGTTCGTG -3′ and GFP-N 5′-CGTCGCCGTCCAGCTCGACCAG -3′ (Tm 56)] and xEf1α [RSD1065 5′- CAGATTGGTGCTGGATAT -3′ and RSD1066 5′- ACTGCCTTGATGACTCCTAG-3′ (Tm60)] for 25 cycles. GFP fluorescence intensity was assessed in-vivo using the GF2 filter on a Leica dissecting microscope and acquired with a Qimaging Retiga 2000R camera. Percent GFP positive numbers were determined from the number of embryos or tadpoles with observable GFP signal when photographed using the same acquisition parameters on all observation days. Both RNA and GFP signal intensity were ranked on a scale of not detectable (-) to maximum signal detected (++++).
Antibody Staining and Microscopy
Cells and tissue explants were fixed in Zamboni's fixative (40mM Na2HPO4, 120mM NaH2PO4, 2% paraformaldehyde and 0.1 % saturated picric acid) for 20 minutes at room temperature, washed with PBS and incubated in PBS with 1% Triton-X100 for 30 minutes. After blocking in 1X blocking reagent (Roche) dissolved in PBS 0.1% Triton X-100 for 1 hour at room temperature the primary antibody was applied in blocking solution overnight at 4°C. Secondary antibodies were obtained from Molecular Probes and incubated for 60 minutes at room temperature. Primary antibodies used were: rat anti-E-cadherin (Transduction Laboratories 1:400 dilution) and mouse monoclonal anti-smooth muscle actin (Sigma, 1:100).
The limbs with GFP expression were fixed in Zamboni's fixative and then sectioned transversely with a cryostat. The sections were double immunostained with anti-GFP (Abcam) antibody at 1:500 dilution and 12/101 antibody (muscle-specific, Developmental Studies Hybridoma Bank, 1:100 dilution. Their secondary antibodies were fluorescein-conjugated anti-rabbit IgG (Vector labs) and Alexa Fluor 594 anti–mouse IgG (Invitrogen), respectively.
GFP expression in tadpoles and organ explants was observed using a Leica MZ16F microscope. Pictures were taken with a Qimaging Retiga 2000R camera and presented using Photoshop (Adobe). Some in vitro cultures were also photographed using a Zeiss LSM510 confocal microscope.
Acknowledgments
This work was supported by the Wellcome Trust and the University of Minnesota. RSD is supported by Training Grant NIH T32 AR050938 “Musculoskeletal Training Grant”. We are grateful to Dr Mark Hornsey for preparing the virus and to Genya Gekker for assistance with flow cytometry.
Grant support: Wellcome Trust 60082
NIH Training Grant T32 AR050938 (RSD)
References
- Allen BG, Weeks DL. Transgenic Xenopus laevis embryos can be generated using phi C31 integrase. Nature Methods. 2005;2:975–979. doi: 10.1038/nmeth814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amaya E, Kroll KL. A method for generating transgenic frog embryos. Methods Mol Biol. 1999;97:393–414. doi: 10.1385/1-59259-270-8:393. [DOI] [PubMed] [Google Scholar]
- Atkinson DL, Stevenson TJ, Park EJ, Riedy MD, Milash B, Odelberg SJ. Cellular electroporation induces dedifferentiation in intact newt limbs. Developmental Biology. 2006;299:257–271. doi: 10.1016/j.ydbio.2006.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck C, Slack JMW. Gut specific expression using mammalian promoters in transgenic Xenopus laevis. Mech Dev. 1999 doi: 10.1016/s0925-4773(99)00217-8. in press. [DOI] [PubMed] [Google Scholar]
- Bergelson JM, Cunningham JA, Droguett G, KurtJones EA, Krithivas A, Hong JS, Horwitz MS, Crowell RL, Finberg RW. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. doi: 10.1126/science.275.5304.1320. [DOI] [PubMed] [Google Scholar]
- Bewley MC, Springer K, Zhang YB, Freimuth P, Flanagan JM. Structural Analysis of the Mechanism of Adenovirus Binding to Its Human Cellular Receptor, CAR. Science. 1999;286:1579–1583. doi: 10.1126/science.286.5444.1579. [DOI] [PubMed] [Google Scholar]
- Brand AH, Perrimon N. Targeted gene-expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- Brown DD, Cai LQ. Amphibian metamorphosis. Developmental Biology. 2007;306:20–33. doi: 10.1016/j.ydbio.2007.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chae J, Zimmerman LB, Grainger RM. Inducible control of tissue-specific transgene expression in Xenopus tropicalis transgenic lines. Mechanisms of Development. 2002;117:235–241. doi: 10.1016/s0925-4773(02)00219-8. [DOI] [PubMed] [Google Scholar]
- Chalmers AD, Slack JMW. The Xenopus tadpole gut: fate maps and morphogenetic movements. Development. 2000;127:381–392. doi: 10.1242/dev.127.2.381. [DOI] [PubMed] [Google Scholar]
- Dale L, Slack JMW. Fate map for the 32 cell stage of Xenopus laevis. Development. 1987;99:527–551. doi: 10.1242/dev.99.4.527. [DOI] [PubMed] [Google Scholar]
- Das B, Brown DD. Controlling transgene expression to study Xenopus laevis metamorphosis. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:4839–4842. doi: 10.1073/pnas.0401011101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Echeverri K, Tanaka EM. Electroporation as a tool to study in vivo spinal cord regeneration. Developmental Dynamics. 2003;226:418–425. doi: 10.1002/dvdy.10238. [DOI] [PubMed] [Google Scholar]
- Eide FF, Eisenberg SR, Sanders TA. Electroporation-mediated gene transfer in free-swimming embryonic Xenopus laevis. FEBS Letters. 2000;486:29–32. doi: 10.1016/s0014-5793(00)02124-4. [DOI] [PubMed] [Google Scholar]
- Falk J, Drinjakovic J, Leung KM, Dwivedy A, Regan AG, Piper M, Holt CE. Electroporation of cDNA/morpholinos to targeted areas of embryonic CNS in Xenopus. BMC Developmental Biology. 2007;7 doi: 10.1186/1471-213X-7-107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardlik R, Palffy R, Hodosy J, Lukacs J, Turna J, Celec P. Vectors and delivery systems in gene therapy. Medical Science Monitor. 2005;11:RA110–RA121. [PubMed] [Google Scholar]
- Gargioli C, Slack JMW. Cell lineage tracing during Xenopus tail regeneration. Development. 2004;131:2669–2679. doi: 10.1242/dev.01155. [DOI] [PubMed] [Google Scholar]
- Gatlin J, Unett DJ, Lerner MR, Garcia JV. Efficient, long-term transgene expression in Xenopus laevis dermal melanophores. Pigment Cell Research. 2001;14:275–282. doi: 10.1034/j.1600-0749.2001.140408.x. [DOI] [PubMed] [Google Scholar]
- Gossen M, Bujard H. Tight Control of Gene-Expression in Mammalian-Cells by Tetracycline-Responsive Promoters. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:5547–5551. doi: 10.1073/pnas.89.12.5547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham FL, Prevec L. Adenovirus-based expression vectors and recombinant vaccines. Biotechnology. 1992;20:363–390. doi: 10.1016/b978-0-7506-9265-6.50022-1. [DOI] [PubMed] [Google Scholar]
- Haas K, Jensen K, Sin WC, Foa L, Cline HT. Targeted electroporation in Xenopus tadpoles in vivo - from single cells to the entire brain. Differentiation. 2002;70:148–154. doi: 10.1046/j.1432-0436.2002.700404.x. [DOI] [PubMed] [Google Scholar]
- Harland RM. In situ hybridization: an improved whole-mount method for Xenopus embryos. Methods Cell Biol. 1991;36:685–695. doi: 10.1016/s0091-679x(08)60307-6. [DOI] [PubMed] [Google Scholar]
- Hartley KO, Nutt SL, Amaya E. Targeted gene expression in transgenic Xenopus using the binary Gal4-UAS system. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:1377–1382. doi: 10.1073/pnas.022646899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi S, McMahon AP. Efficient Recombination in Diverse Tissues by a Tamoxifen-Inducible Form of Cre: A Tool for Temporally Regulated Gene Activation/Inactivation in the Mouse. Developmental Biology. 2002;244:305–318. doi: 10.1006/dbio.2002.0597. [DOI] [PubMed] [Google Scholar]
- He Y, Chipman PR, Howitt J, Bator CM, Whitt MA, Baker TS, Kuhn RJ, Anderson CW, Freimuth P, Rossmann MG. Interaction of coxsackievirus B3 with the full length coxsackievirusadenovirus receptor. Nature Structural Biology. 2001;8:874–878. doi: 10.1038/nsb1001-874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawakami Y, Esteban CR, Raya M, Kawakami H, Marti M, Dubova I, Belmonte JCI. Wnt/beta-catenin signaling regulates vertebrate limb regeneration. Genes & Development. 2006;20:3232–3237. doi: 10.1101/gad.1475106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolm PJ, Sive HL. Efficient hormone-inducible protein function in Xenopus-laevis. Developmental Biology. 1995;171:267–272. doi: 10.1006/dbio.1995.1279. [DOI] [PubMed] [Google Scholar]
- Krieg PA, Melton DA. Functional messenger RNAs are produced by Sp6 in vitro transcription of cloned c. DNAs Nucleic Acid Res. 1984;12:7057–7070. doi: 10.1093/nar/12.18.7057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroll KL, Amaya E. Transgenic Xenopus embryos from sperm nuclear transplantations reveal FGF signaling requirements during gastrulation. Development. 1996;122:3173–3183. doi: 10.1242/dev.122.10.3173. [DOI] [PubMed] [Google Scholar]
- Lagna G, Hemmati-Brivanlou A. Use of dominant negative constructs to modulate gene expression. Curr Top Dev Biol. 1998;36:75–98. doi: 10.1016/s0070-2153(08)60496-2. [DOI] [PubMed] [Google Scholar]
- Li M, Rohrer B. Gene silencing in Xenopus laevis byDNA vector-based RNA interference and transgenesis. Cell Research. 2006;16:99–105. doi: 10.1038/sj.cr.7310013. [DOI] [PubMed] [Google Scholar]
- Lin G, Chen Y, Slack JMW. Regeneration of melanophores and other neural crest derivatives in the Xenopus tadpole tail. BMC Developmental Biology. 2007;7:56. doi: 10.1186/1471-213X-7-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieuwkoop PD, Faber J. Normal table of Xenopus laevis (Daudin) Amsterdam: North-Holland: 1967. [Google Scholar]
- Ogino H, McConnell WB, Grainger RM. Highly efficient transgenesis in Xenopus tropicalis using I-SceI meganuclease. Mechanisms of Development. 2006;123:103–113. doi: 10.1016/j.mod.2005.11.006. [DOI] [PubMed] [Google Scholar]
- Pan FC, Chen YL, Loeber J, Henningfeld K, Pieler T. I-SceI meganuclease-mediated transgenesis in Xenopus. Developmental Dynamics. 2006;235:247–252. doi: 10.1002/dvdy.20608. [DOI] [PubMed] [Google Scholar]
- Petrella J, Cohen CJ, Gaetz J, Bergelson JM. A zebrafish coxsackievirus and adenovirus receptor homologue interacts with coxsackie B virus and adenovirus. Journal of Virology. 2002;76:10503–10506. doi: 10.1128/JVI.76.20.10503-10506.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Philipson L, Pettersson RF. The coxsackie-adenovirus receptor - A new receptor in the immunoglobulin family involved in cell adhesion. Adenoviruses: Model and Vectors in Virus-Host Interactions. 2004:87–111. doi: 10.1007/978-3-662-05599-1_3. [DOI] [PubMed] [Google Scholar]
- Ryffel GU, Werdien D, Turan G, Gerhards A, Goosses S, Senkel S. Tagging muscle cell lineages in development and tail regeneration using Cre recombinase in transgenic Xenopus. Nucleic Acids Research. 2003;31:1044–1051. doi: 10.1093/nar/gng044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sive HL, Grainger RM, Harland RM. Early Development of Xenopus laevis: A Laboratory Manual. Cold Spring Harbor Laboratory Press; 2000. [Google Scholar]
- Slack JMW. Essential Developmental Biology. Oxford: Blackwell Science; 2005. [Google Scholar]
- Slack JMW, Lin G, Chen Y. The Xenopus tadpole - a new model for regeneration research. Cellular and Molecular Life Sciences. 2008;65:54–63. doi: 10.1007/s00018-007-7431-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugiyama S, Miyoshi H, Yamauchi K. Characteristics of a thyroid hormone responsive reporter gene transduced into a Xenopus laevis cell line using lentivirus vector. General and Comparative Endocrinology. 2005;144:270–279. doi: 10.1016/j.ygcen.2005.06.003. [DOI] [PubMed] [Google Scholar]
- Urlinger S, Baron U, Thellmann M, Hasan MT, Bujard H, Hillen W. Exploring the sequence space for tetracycline-dependent transcriptional activators: Novel mutations yield expanded range and sensitivity. Proceedings of the National Academy of Sciences. 2000;97:7963–7968. doi: 10.1073/pnas.130192197. %R10.1073/pnas.130192197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma IM, Weitzman MD. Gene therapy: Twenty-first century medicine. Ann Rev Biochem. 2005;74:711–738. doi: 10.1146/annurev.biochem.74.050304.091637. [DOI] [PubMed] [Google Scholar]
- Waldner C, Sakamaki K, Ueno N, Turan G, Ryffel GU. Transgenic Xenopus laevis strain expressing Cre recombinase in muscle cells. Developmental Dynamics. 2006;235:2220–2228. doi: 10.1002/dvdy.20880. [DOI] [PubMed] [Google Scholar]
- Werdien D, Peiler G, Ryffel GU. FLP and Cre recombinase function in Xenopus embryos. Nucleic Acids Res. 2001;29:e53. doi: 10.1093/nar/29.11.e53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler GN, Hamilton FS, Hoppler S. Inducible gene expression in transgenic Xenopus embryos. Current Biology. 2000;10:849–852. doi: 10.1016/s0960-9822(00)00596-0. [DOI] [PubMed] [Google Scholar]