

Abstract
Molecular and morphological data have important roles in illuminating evolutionary history. DNA data often yield well resolved phylogenies for living taxa, but are generally unattainable for fossils. A distinct advantage of morphology is that some types of morphological data may be collected for extinct and extant taxa. Fossils provide a unique window on evolutionary history and may preserve combinations of primitive and derived characters that are not found in extant taxa. Given their unique character complexes, fossils are critical in documenting sequences of character transformation over geologic time and may elucidate otherwise ambiguous patterns of evolution that are not revealed by molecular data alone. Here, we employ a methodological approach that allows for the integration of molecular and paleontological data in deciphering one of the most innovative features in the evolutionary history of mammals—laryngeal echolocation in bats. Molecular data alone, including an expanded data set that includes new sequences for the A2AB gene, suggest that microbats are paraphyletic but do not resolve whether laryngeal echolocation evolved independently in different microbat lineages or evolved in the common ancestor of bats and was subsequently lost in megabats. When scaffolds from molecular phylogenies are incorporated into parsimony analyses of morphological characters, including morphological characters for the Eocene taxa Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropteryx, the resulting trees suggest that laryngeal echolocation evolved in the common ancestor of fossil and extant bats and was subsequently lost in megabats. Molecular dating suggests that crown-group bats last shared a common ancestor 52 to 54 million years ago.
Both molecular and morphological data have important roles in elucidating evolutionary history and phylogeny. Advantages of molecular data include the large number of characters that are available (1) as well as sophisticated models of sequence evolution that may be used in phylogenetic analyses (2). However, in most instances it has not been possible to obtain DNA sequences for fossil taxa. In contrast, certain types of morphological data may be collected for both fossil and extant taxa. Fossils provide a unique window on evolutionary history and often preserve combinations of primitive and derived characters that are not found in extant taxa (3). Given their unique character complexes, fossils are critical in documenting sequences of character transformation over geologic time (4). Morphological evidence from fossils may reveal patterns of evolution that are not apparent from molecular phylogenies alone. Below, we illustrate a methodological approach that allows for the integration of molecular and paleontological data in deciphering one of the most innovative features in the evolutionary history of mammals—laryngeal echolocation in bats.
Interfamilial relationships among bats, as well as relationships of the order Chiroptera to other eutherian orders, have been investigated with both morphological and molecular data. Morphological data suggest that bats are in the superordinal group Archonta (5), which also includes primates, tree shrews, and flying lemurs. Within Archonta, the Volitantia hypothesis postulates a sister-group relationship between flying lemurs and bats (6). Although some authors have questioned bat monophyly based on evidence from the penis and nervous system (7, 8), the bulk of morphological data supports bat monophyly (9). Among living Chiroptera, morphological data provide strong support for the reciprocal monophyly of megachiropterans (Old World fruit bats), all of which lack laryngeal echolocation, and microchiropterans, all of which possess laryngeal echolocation (6, 10, 11). In a total evidence analysis that combined 180 morphological and 12 restriction site characters, Simmons (10) obtained 100% bootstrap support for the monophyly of extant microbats.
The oldest fossil bats are from early Eocene deposits in Africa, Australia, Europe, and North America. Of the eight currently recognized genera from the early Eocene, four (Icaronycteris, Archaeonycteris, Hassianycteris, Palaeochiropteryx) are known from relatively complete skeletons (6). Simmons and Geisler (6) expanded the data set of Simmons (10) to include 208 characters (195 morphological); they also added Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropteryx to this matrix. Based on an extensive set of phylogenetic analyses, they concluded that the Eocene taxa constitute a consecutive series of sister-taxa to a monophyletic crown-group Microchiroptera. Furthermore, the Eocene fossils exhibit anatomical features that suggest the occurrence of laryngeal echolocation in these taxa. Simmons and Geisler (6) thus concluded that laryngeal echolocation evolved only once in the evolutionary history of Chiroptera.
Molecular data validate the conclusion that the order Chiroptera is monophyletic (12, 13). There are also unexpected results from molecular studies. First, bats are not closely related to other archontans (14–17). Rather, bats are members of the clade Laurasiatheria (16, 18), which also includes eulipotyphlan insectivores, pangolins, carnivores, perissodactyls, and cetartiodactyls. This implies that putative synapomorphies of Volitantia are instead homoplasic features in Chiroptera and Dermoptera. Molecular data also challenge microbat monophyly and instead suggest that microbats are paraphyletic, with rhinolophoids more closely related to megabats than to other microbats (15, 19, 20). New sequence data presented below provide increased support for this hypothesis. Microbat paraphyly implies that complex laryngeal echolocation either evolved independently in rhinolophoids and other microbats or evolved in the ancestor of Chiroptera with subsequent loss in megabats. Given these competing hypotheses, which are equally parsimonious in the context of molecular phylogenies that have only included living taxa, we investigated the origin and evolutionary history of laryngeal echolocation in microbats by using an integrated approach that used both molecular phylogenies and morphological data for living and extinct taxa. Molecular phylogenies were used to construct a backbone phylogenetic constraint, or scaffold, for a subset of extant bat families/subfamilies for which DNA phylogenies are available (see Methods). Parsimony analyses with the morphological data set of Simmons and Geisler (6), in conjunction with the molecular scaffolds, provide support for the hypothesis that laryngeal echolocation evolved in the common ancestor of Chiroptera and was subsequently lost in megabats.
Methods
New sequences for ≈1.3 kb of the A2AB gene were amplified and sequenced as described (16, 21) for Megaderma lyra (GenBank accession no. AF337537), Hipposideros commersoni (AF337538), Pteropus rayneri (AF337539), Myotis daubentoni (AF337540), Tonatia bidens (AF337541), Tadarida brasiliensis (AF337542), and Taphozous sp. (AF337543). Additional A2AB sequences were obtained from GenBank for one megabat (Cynopterus sphinx; AJ251181) and the outgroups Rattus norvegicus (M32061), Homo sapiens (M34041), Felis catus (AJ251174), and Cynocephalus variegatus (AJ251182). The outgroups included one laurasiatherian (Felis) and two archontans (Homo, Cynocephalus) based on alternate views that bats are either members of Laurasiatheria or Archonta. The A2AB sequences were aligned by using clustal w (22). Phylogenetic analyses were performed for A2AB sequences alone as well as for a concatenation of sequences that included published sequences from Teeling et al. (15) for four additional nuclear genes (RAG1, RAG2, vWF, BRCA1) and three mitochondrial genes (12S rRNA, tRNA valine, 16S rRNA) for the same taxa. Phylogenetic analyses included maximum likelihood (ML), minimum evolution (ME), and maximum parsimony (MP). Given uneven base composition and differences in transition and transversion rates, ML analyses were performed under the HKY85 model of sequence evolution; analyses were performed with and without an allowance for a gamma (Γ) distribution of rates. ML estimates of the transition to transversion ratio (ts/tv) and the shape parameter (α) of the Γ distribution were obtained from MP trees. For the A2AB data set, ts/tv was estimated at 1.89 without heterogeneity; with rate-heterogeneity, ts/tv and α were estimated at 2.26 and 0.40, respectively. For the 9427-bp data set, ts/tv was estimated at 2.16 without heterogeneity; with rate-heterogeneity, ts/tv and α were estimated at 2.44 and 0.52, respectively. ME analyses were performed with maximum likelihood (HKY85 model) and logdet distances. MP analyses were unweighted. In all analyses, we used tree-bisection and reconnection (TBR) branch-swapping. Bootstrap analyses used 500 replications for all methods. All phylogenetic analyses were performed with PAUP* 4.0 (23).
Given that all of our analyses supported microbat paraphyly, we used Monte Carlo simulations to investigate whether or not different phylogenetic methods will fail to recover traditional microbat monophyly because of systematic biases and/or taxonomic sampling. First, we determined the best likelihood tree for the 9427-bp data set, including branch lengths, given the constraint of microbat monophyly. Given this tree, 100 simulated data sets of length 9427 bp were generated by using Seq-Gen (24) under the HKY85 model of sequence evolution with empirical base frequencies (A = 0.294; C = 0.239; G = 0.243; T = 0.224), ts/tv = 2.44, and α = 0.52. We then analyzed each simulated data set with MP and ML to determine differences in tree length (for MP) and in likelihood (for ML) between the best trees with and without microbat monophyly.
Quartet dating (25) with a two-rate model was used to estimate the divergence time between megabats plus rhinolophoids versus nonrhinolophoid microbats. This method requires two pairs of sister taxa in conjunction with reliable divergence dates for each of these pairs. The method then estimates the timing of the split between these two groups. qdate can be used to estimate dates under one- and two-rate models. The former model assumes the same rate over the entire quartet, whereas the latter model allows different rates for each pair within the quartet. We used the two-rate model in all of our comparisons. Maximum likelihood estimates of ts/tv and α were obtained for the relevant quartets with paup* 4.0 (23). We obtained divergence estimates based on individual genes as well as a concatenation of all nuclear genes. Dates were rejected if the likelihood score for the quartet tree with the two-rate model was significantly worse than the likelihood score with a five-rate model following the recommendation of Rambaut and Bromham (25). Based on earliest fossil occurrences, we assumed divergence times of 43 million years for the split between megabats and rhinolophoids (6, 26), 43 million years for the split between emballonurids and noctilionoids (6, 26), and 51 million years for the split between emballonurids and vespertilionoids (6, 26, 27), respectively.
Total evidence has been suggested as one approach to integrate molecular and morphological data in phylogenetic analysis. One limitation of total evidence is that it has been confined to maximum parsimony, whereas other methods of analysis, such as maximum likelihood, may be more appropriate with molecular data (2). Another limitation, at least in cases that involve fossil taxa, is that molecular data are usually unattainable for fossils. We did not use the total evidence approach for these reasons and also because molecular and morphological data for taxa relevant to this study have been sampled at different taxonomic levels that cannot be readily combined. Instead, we used a new approach to analyze morphological data that incorporated important findings from molecules. Specifically, we used molecular scaffolds from molecular analyses in MP analyses with morphological data. The scaffold that we used includes clades that were strongly supported (i.e., bootstrap support >90%) by both concatenated DNA sequences (ref. 15 and Fig. 1) and single-copy DNA hybridization (20) and was as follows: (Outgroups, (Pteropodidae (Hipposiderinae, Megadermatidae)), (Molossinae, Myotinae, Emballonuridae, Phyllostomidae)). Other taxa, including the Eocene fossils, were not constrained by the molecular scaffold. The data set of Simmons and Geisler (6) includes 195 morphological characters. We analyzed three versions of this data set: (i) All characters. (ii) A reduced matrix (192 characters) that omitted three characters [character 26 (enlargement of cochlea), character 35 (orbicular process on malleus), and character 74 (cranial expansion of stylohyal)] that were mapped in subsequent parsimony reconstructions. It was necessary to remove these characters to generate trees that are independent of these same characters. (iii) Eighty-one of the 195 characters for which data were scored for at least one of the four fossil taxa. Characters were unordered following Simmons and Geisler (6). We used unweighted MP and successive approximations MP with reweighting according to the rescaled consistency index. Bootstrap analyses included 500 replications. All searches were heuristic with 1,000 random input orders per replication and tree-bisection and reconnection branch-swapping. Parsimony reconstructions of ancestral character states were obtained by using macclade (28) with both delayed transformations (Deltran) and accelerated transformations (Acctran) for all of the most parsimonious trees (unweighted and reweighted) that were obtained for the 195, 192, and 81 character data sets.
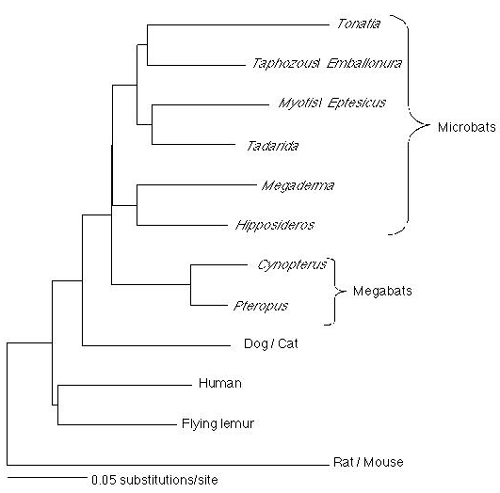
Figure 1.

Maximum likelihood phylogram (HKY85 model with a Γ distribution of rates) based on the 9427-bp data set (−ln likelihood = 48718.269). Scale bar corresponds to 5% sequence divergence. ML bootstrap support values are shown adjacent to or above branches. Support values with other methods of analysis are given in Table 1.
Results and Discussion
Molecular Phylogenies.
When A2AB sequences were analyzed alone, both the megabat-rhinolophoid and nonrhinolophoid microbat clades were recovered (Table 1). Support for the former clade ranged from 79 to 99%, whereas support for the latter clade ranged from 57 to 94%. Support for microbat monophyly ranged from 0 to 18%. These results provide independent support that corroborates earlier molecular studies (15, 19, 20) reporting a megabat-rhinolophoid association and contradicting the monophyly of Microchiroptera.
Table 1.
Bootstrap support for select clades with the A2AB and 9427-bp data sets
| Method of analysis | Clade
|
|||
|---|---|---|---|---|
| Laurasia. | Rhino. + megabats | Nonrhino. microbats | Microbat monophyly | |
| MP | 84 /98 | 89 /98 | 90 /100 | 7 /1 |
| TVP | 59 /100 | 93 /100 | 57 /100 | 0 /0 |
| ME-Logdet | 83 /100 | 79 /99 | 77 /100 | 18 /1 |
| ME-ML | 77 /100 | 88 /100 | 76 /100 | 8 /0 |
| ML w/o Γ | 90 /100 | 97 /100 | 92 /100 | 2 /0 |
| ML w/Γ | 92 /100 | 99 /100 | 94 /100 | 0 /0 |
The first bootstrap support value in each cell is for the A2AB data set; the second value is for the concatenated 9427-bp data set. Laurasia., Laurasiatheria; Nonrhino., nonrhinolophoid microbats; Rhino., rhinolophoid microbats (Megaderma and Hipposideros); ME, minimum evolution; TVP, transversion parsimony.
Fig. 1 shows a maximum likelihood tree for the concatenated data set (9427 bp) that includes A2AB and the data of Teeling et al. (15). Bootstrap support values are also shown on this tree. The basal split among bats is between megabats plus rhinolophoids versus nonrhinolophoid microbats. Both the megabat-rhinolophoid and nonrhinolophoid microbat clades are supported at or above the 98% bootstrap support level with diverse phylogenetic methods (Table 1). Support for microbat monophyly was always low (≤1%) in analyses with the 9427-bp data set (Table 1). Kishino–Hasegawa tests rejected microbat monophyly (MP: Δ in # of steps = 18; P = 0.0201; ML without Γ: Δ in ln likelihood = 53.208; P = 0.0013; ML with Γ: Δ in ln likelihood = 32.836; P = 0.0026). Monte Carlo simulations provide no support for the possibility that failure to recover microbat monophyly results from systematic biases associated with MP and/or ML (see supplemental data, which is published on the PNAS web site, www.pnas.org).
In contrast to traditional classifications for Chiroptera, which divide bats into the suborders Microchiroptera and Megachiroptera, molecular results presented here and elsewhere (15, 20) suggest the need for a new classification in which Rhinolophidae and Megadermatidae are placed in the same suborder as Pteropodidae. Also, we propose including Emballonuridae in Yangochiroptera. Our proposed classification for bats included in our molecular study is presented in Table 2.
Table 2.
Traditional and proposed classifications for bats included in this study
| Simmons (10) | This study |
|---|---|
| ORDER CHIROPTERA | ORDER CHIROPTERA |
| Suborder Megachiroptera | Suborder Yinpterochiroptera |
| Family Pteropodidae (Cynopterus, Pteropus) | Superfamily Pteropodoidea |
| Suborder Microchiroptera | Family Pteropodidae (Cynopterus, Pteropus) |
| Infraorder incertae sedis | Superfamily Rhinolophoidea |
| Superfamily Emballonuroidea | Family Megadermatidae (Megaderma) |
| Family Emballonuridae (Emballonura, Taphozous) | Family Rhinolophidae (Hipposideros) |
| Infraorder Yinochiroptera | Suborder Yangochiroptera |
| Superfamily Rhinolophoidea | Superfamily Emballonuroidea |
| Family Megadermatidae (Megaderma) | Family Emballonuridae (Emballonura, Taphozous) |
| Family Rhinolophidae (Hipposideros) | Superfamily Noctilionoidea |
| Infraorder Yangochiroptera | Family Phyllostomidae (Tonatia) |
| Superfamily Noctilionoidea | Superfamily Vespertilionoidea |
| Family Phyllostomidae (Tonatia) | Family Vespertilionidae (Myotis, Eptesicus) |
| Superfamily Vespertilionoidea | Family Molossidae (Tadarida) |
| Family Vespertilionidae (Myotis, Eptesicus) | |
| Family Molossidae (Tadarida) |
The proposed suborder Yinpterochiroptera is based on the conjunction of “yin” and “ptero” to reflect the composition of this new taxon. The family Emballonuridae is now included in Yangochiroptera.
Molecular Dates.
Quartet dating results for the most recent common ancestor of crown-group bats were obtained for RAG1 (53.2 million years), vWF (53.0 million years), A2AB (48.3 million years), and a concatenation of all RAG1, RAG2, vWF, A2AB, and BRCA1 (53.4 million years). Estimates based on other genes were rejected because of significant differences in likelihood scores in comparisons between two-rate versus five-rate models. In cases where estimates were accepted, the overlapping range of dates that is consistent with all of the 95% confidence intervals was 51.6 to 53.7 million years (Table 3).
Table 3.
Molecular divergence estimates for the base of crown-group Chiroptera
| Gene | Quartet | Ts/tv | α | Date (m.y.) | 95% C.I. |
|---|---|---|---|---|---|
| A2AB | (Tap,Ton),(Meg,Cyn) | 2.27 | 0.36 | 48.3 | 44.6–53.7 |
| RAG1 | (Tap,Myo),(Hip,Cyn) | 2.76 | 0.16 | 53.2 | 51.0–61.0* |
| vWF | (Tap,Tad),(Hip,Cyn) | 1.87 | 0.34 | 53.0 | 51.0–59.5* |
| All nuclear | (Tap,Tad),(Hip,Pte) | 2.37 | 0.59 | 53.4 | 51.6–55.5 |
Estimates were obtained with qdate (24). Tap, Taphozous; Ton, Tonatia; Meg, Megaderma; Cyn, Cynopterus; Myo, Myotis; Hip, Hipposideros; Pte, Pteropus; α, shape parameter of the gamma distribution with four discrete rate-categories; m.y., millions of years; C.I., confidence interval.
Lower confidence interval limited by a specified fossil date.
Analyses with Morphological Data.
MP analyses with three different variations of the morphological data set all resulted in trees on which the basal split among chiropterans is between Icaronycteris and all other taxa (Fig. 2). Furthermore, bootstrap support for the clade containing crown-group bats, Palaeochiropteryx, Hassianycteris, and Archaeonycteris, to the exclusion of Icaronycteris, was robust and ranged from 91 to 97% in six different MP analyses. Notably, these analyses were performed with and without characters that could be scored for the Eocene taxa; thus, the same placement was obtained for Icaronycteris whether we included or excluded characters that were coded as missing for all of the fossils.
Figure 2.

Strict consensus of three equally most parsimonious trees (309 steps) for the 81 character data set with unweighted parsimony and the molecular scaffold. Bootstrap percentages are shown in bold for clades supported above 50%. The hypothesis that laryngeal echolocation evolved in the common ancestor of bats, with subsequent loss in megabats, requires one evolutionary gain (1G) and one evolutionary loss (1L). The competing hypothesis, that laryngeal echolocation evolved independently in different microbats, requires four evolutionary gains on the branches labeled 2G. Other analyses also produced trees on which the Eocene fossils constitute a paraphyletic assemblage, either as shown in Fig. 3 or entirely at the base of Chiroptera. Taxa shown with asterisks were constrained by the following molecular scaffold: (Outgroups, (Pteropodidae, (Hipposiderinae, Megadermatidae)), (Molossinae, Myotinae, Emballonuridae, Phyllostomidae)). Other taxa, including the Eocene fossils, were not constrained. Color designations for taxa are as follows: outgroups, black; early Eocene fossils, green; emballonuroids, red; yangochiropterans, blue; yinochiropterans, purple; megabats, orange.
In the tree shown in Fig. 2, the Eocene fossils constitute a paraphyletic assemblage that is distributed at the base of Chiroptera as well as at the base of a clade that includes Yangochiroptera and Emballonuridae [sensu Simmons (10)]. Other methods also produced trees on which the Eocene fossils constitute a paraphyletic assemblage, either as shown in Fig. 2 or entirely at the base of Chiroptera. Among living bats, our analyses recovered both Yangochiroptera and Yinochiroptera of Simmons (10), although not with high bootstrap support. In contrast to our results, the analyses of Simmons and Geisler (6) produced trees on which the Eocene taxa were consecutive sister-taxa to Microchiroptera only. The placement of early Eocene fossils in Fig. 2 is reminiscent of Van Valen's (29) paraphyletic Eochiroptera, although on Van Valen's (29) tree extant megabats and microbats are each monophyletic groups.
Given the topologies that resulted from our analyses, we obtained parsimony character state reconstructions for features that are indicative of laryngeal echolocation. All living bats with a stylohyal that is expanded cranially, a large orbicular process on the malleus, and a moderately enlarged cochlea are capable of laryngeal echolocation (6). Simmons and Geisler (6) inferred that Icaronycteris was also capable of primitive laryngeal echolocation based on the occurrence of these three features in this taxon. Other authors (30, 31) have also suggested that Icaronycteris was a laryngeal echolocating bat. In every case, our reconstructions suggest that the common ancestor of Icaronycteris and other bats exhibited a stylohyal that was expanded at the cranial tip, a large orbicular process on the malleus, and a moderately enlarged cochlea. Based on these reconstructions, we suggest that the most recent common ancestor of living chiropterans, Icaronycteris, Archaeonycteris, Palaeochiropteryx, and Hassianycteris was a bat with laryngeal echolocating capabilities. Furthermore, the most parsimonious reconstruction for the evolution of laryngeal echolocation is that this feature evolved only once (1G, Fig. 2) and was subsequently lost in megabats (1L, Fig. 2). The alternate hypothesis, that laryngeal echolocation evolved independently in different lineages and was not lost in the ancestry of megabats, is less parsimonious and requires four to six separate origins given the trees that resulted from different parsimony analyses (four for the trees shown in Fig. 2; six for trees where the Eocene taxa form a consecutive series of sister-taxa to all extant chiropterans).
The Evolution of Echolocation.
“Flight-first” (32), “echolocation-first” (33), and “tandem evolution” (34) models have been proposed to account for the evolution of powered flight and laryngeal echolocation in Chiroptera. If we accept the monophyly of Chiroptera and Microchiroptera, respectively, it follows that the flight-first hypothesis is most parsimonious with flight evolving in the common ancestor of Chiroptera and laryngeal echolocation evolving in the common ancestor of Microchiroptera. The echolocation-first and tandem evolution hypotheses become even more difficult if we accept the monophyly of Archonta because the latter disconnects the ancestry of bats from other mammalian taxa that echolocate.
In contrast, our results suggest that laryngeal echolocation, like flight, evolved before the most recent common ancestor of living and fossil taxa. Given the deployment of both flight and laryngeal echolocation to deep levels in the chiropteran tree, an additional implication is that the flight-first, echolocation-first, and tandem evolution hypotheses all remain viable. Furthermore, molecular data suggest that bats are members of the superordinal clade Laurasiatheria (16), a group that also includes eulipotyphlan insectivores such as moles and shrews. Both Eulipotyphla and Chiroptera may be early offshoots within Laurasiatheria (16, 18). Notably, some shrews in Eulipotyphla are capable of echolocation (6). The ancestry of bats may thus have a more proximal link to other mammalian taxa that echolocate. Discrimination between the flight-first, echolocation-first, and tandem evolution hypotheses may only be possible with future fossil discoveries. In any case, it now appears that the first bats were much more like Icaronycteris than previously believed.
Molecular dates presented here suggest that crown-group bats last shared an ancestor in the range of 52 to 54 million years based on the overlap of confidence intervals for individual genes and concatenations thereof. These estimates, of course, assume that the fossil calibration points are reliable. If we accept the fossil calibrations, then our molecular dates imply that key transitional fossils documenting the origin of flight and echolocation in the ancestry of living bats should be older than 52 to 54 million years. Similarly, molecular dates that were calculated by Nikaido et al. (17) suggest that flight and echolocation evolved during a 25 million year window extending from ≈83 million years to 58 million years.
Although laryngeal echolocation may have evolved only once in the evolutionary history of Chiroptera, there have been numerous modifications once the basic system was in place. Given the moderate size of the cochlea in the common ancestor of crown group plus fossil chiropterans, echolocation was probably a primitive type of low-duty cycle echolocation that was used for orientation and obstacle detection. Subsequent enlargement of the cochlea in Hassianycteris, Palaeochiropteryx, and living microbats resulted in sophisticated laryngeal echolocation and permitted aerial hawking rather than gleaning from a perch.
There are observations that are consistent with the hypothesis that laryngeal echolocation was lost in megabats. First, the moderately enlarged cochlea in some nonecholocating megachiropterans, which overlaps in size with the cochlea in Icaronycteris, Archaeonycteris, and some living microchiropterans (e.g., Megaderma lyra, Phyllostomus hastatus) (6, 30), may be a residual feature from this earlier stage of evolution. Second, even though living megabats do not have an expanded stylohyal, they are capable of emitting short broadband or multiharmonic FM calls in social situations such as precopulation and hostile male–male interactions (33); these calls are similar to those that microbats sometimes use in comparable social situations (33). Our reconstructions suggest that broadband or multiharmonic FM calls in megabats trace back to an ancestry in which similar calls were used in echolocation. Finally, the highly developed visual system in megabats has been viewed as a primitive feature of Chiroptera given the widespread occurrence of highly developed visual systems in other archontans including tree shrews, flying lemurs, and primates. However, molecular data consistently dissociate Chiroptera from other archontans (14–17), which suggests that the highly developed visual orientation system in megabats is an autapomorphy for this group. If the protobat did not have an enhanced visual system, as in megabats, then constraints related to brain-size (6) would have been less likely to preclude echolocation in the earliest bats.
Among megabats, a different type of echolocation, based on tongue-clicks, has been demonstrated in Rousettus (35). Given the hypothesis that laryngeal echolocation was lost in megabats, this raises the following question: Does Rousettus represent an intermediate stage in the loss of laryngeal echolocation? Molecular evidence places Rousettus as sister to an endemic African clade of megabats rather than at the base of the megabat radiation (36, 37). This phylogenetic position for Rousettus argues that echolocation was secondarily gained in this genus and is not an intermediate stage in the loss of laryngeal echolocation.
Molecular data have provided a new perspective on relationships among living bat families that has important consequences for understanding the origins of flight and echolocation in bats (15, 20). Fossil data are also of fundamental importance in reconstructing early stages in bat evolution and we agree with Simmons and Geisler (6) that Icaronycteris, Archaeonycteris, Palaeochiropteryx, and Hassianycteris provide “an unprecedented view of steps leading to a major adaptive radiation of mammals.” However, consideration of these fossils jointly with molecular phylogenies alters our view of these steps. Instead of a phylogenetic placement that is basal to living and fossil microbats, Icaronycteris is basal to fossil forms as well as all extant bats, megabats included. Furthermore, Icaronycteris, Archaeonycteris, Palaeochiropteryx, and Hassianycteris constitute a paraphyletic assemblage at the base of Chiroptera. This phylogenetic arrangement, in conjunction with reconstructions for key characters in the echolocation apparatus, implies that megabats evolved from echolocating microbat ancestors and have secondarily lost laryngeal echolocation. Phylogenetic evidence supporting the hypothesis that laryngeal echolocation evolved once in bats and was subsequently lost in megabats has only emerged from a combined analysis that integrates fossils with molecular phylogenies. The incorporation of molecular phylogenetic constraints into investigations of evolutionary history should be applicable in other cases where a taxonomic group includes fossils and living taxa.
Supplementary Material
Acknowledgments
We thank Drs. John Kirsch, Michael Woodburne, and four anonymous reviewers for helpful comments on earlier drafts of this manuscript. This work was supported by the National Science Foundation (DEB-9903810 to M.S.S.) and the Training and Mobility of Researchers program of the European Commission (to M.J.S. and W.W.d.J.).
Abbreviations
- ML
maximum likelihood
- MP
maximum parsimony
- ts/tv
transition to transversion ratio
Footnotes
References
- 1.Hillis D M. Annu Rev Ecol Syst. 1987;18:23–42. [Google Scholar]
- 2.Swofford D L, Olsen G J, Waddell P J, Hillis D M. In: Molecular Systematics. 2nd Ed. Hillis D M, Moritz C, Mable B K, editors. Sunderland, MA: Sinauer; 1996. pp. 407–514. [Google Scholar]
- 3.O'Leary M A, Geisler J H. Syst Biol. 1999;48:455–490. doi: 10.1080/106351599260102. [DOI] [PubMed] [Google Scholar]
- 4.Donoghue M J, Doyle J A, Gauthier J, Kluge A G, Rowe T. Annu Rev Ecol Syst. 1989;20:431–460. [Google Scholar]
- 5.Novacek M J. Nature (London) 1992;356:121–125. doi: 10.1038/356121a0. [DOI] [PubMed] [Google Scholar]
- 6.Simmons N B, Geisler J H. Bull Am Mus Nat Hist. 1998;235:1–182. [Google Scholar]
- 7.Smith J D, Madkour G. In: Proceedings of the Fifth International Bat Research Conference. Wilson D E, Gardner A L, editors. Lubbock, TX: Texas Tech Press; 1980. pp. 347–365. [Google Scholar]
- 8.Pettigrew J D. Science. 1986;231:1304–1306. doi: 10.1126/science.3945827. [DOI] [PubMed] [Google Scholar]
- 9.Simmons N B. Am Mus Novit. 1994;3103:1–54. [Google Scholar]
- 10.Simmons N B. In: Bat Biology and Conservation. Kunz T H, Racey P A, editors. Washington, DC: Smithsonian Inst. Press; 1998. pp. 3–26. [Google Scholar]
- 11.Simmons N B. In: Ontogeny, Functional Ecology, and Evolution of Bats. Adams R A, Pedersen S C, editors. Cambridge, U.K.: Cambridge Univ. Press; 2000. pp. 9–58. [Google Scholar]
- 12.Bailey W J, Slightom J L, Goodman M. Science. 1992;256:86–89. doi: 10.1126/science.1301735. [DOI] [PubMed] [Google Scholar]
- 13.Stanhope M J, Czelusniak J, Si J-S, Nickerson J, Goodman M. Mol Phylogenet Evol. 1992;1:148–160. doi: 10.1016/1055-7903(92)90026-d. [DOI] [PubMed] [Google Scholar]
- 14.Pumo D E, Finamore P S, Franek W R, Phillips C J, Tarzami S, Balzarano D. J Mol Evol. 1998;15:709–717. doi: 10.1007/pl00006430. [DOI] [PubMed] [Google Scholar]
- 15.Teeling E C, Scally M, Kao D J, Romagnoli M L, Springer M S, Stanhope M J. Nature (London) 2000;403:188–192. doi: 10.1038/35003188. [DOI] [PubMed] [Google Scholar]
- 16.Madsen O, Scally M, Kao D J, DeBry R W, Douady C J, Adkins R, Amrine H, Stanhope M J, de Jong W W, Springer M S. Nature (London) 2001;409:610–614. doi: 10.1038/35054544. [DOI] [PubMed] [Google Scholar]
- 17.Nikaido M, Harada M, Cao Y, Hasegawa M, Okada N. J Mol Evol. 2000;51:318–328. doi: 10.1007/s002390010094. [DOI] [PubMed] [Google Scholar]
- 18.Waddell P J, Okada N, Hasegawa M. Syst Biol. 1999;48:1–5. [PubMed] [Google Scholar]
- 19.Porter C A, Goodman M, Stanhope M J. Mol Phylogenet Evol. 1996;5:89–101. doi: 10.1006/mpev.1996.0008. [DOI] [PubMed] [Google Scholar]
- 20.Hutcheon J M, Kirsch J A W, Pettigrew J D. Philos Trans R Soc London B. 1998;353:607–617. doi: 10.1098/rstb.1998.0229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Springer M S, Cleven G C, Madsen O, de Jong W W, Waddell V G, Amrine H M, Stanhope M J. Nature (London) 1997;388:61–64. doi: 10.1038/40386. [DOI] [PubMed] [Google Scholar]
- 22.Thompson J D, Higgins G D, Gibson T J. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Swofford D L. PAUP* Phylogenetic Analysis Using Parsimony (* and Other Methods) Sunderland, MA: Sinauer; 1998. , Version 4. [Google Scholar]
- 24.Rambaut A, Grassly N C. Comput Appl Biosci. 1997;13:303–306. doi: 10.1093/bioinformatics/13.3.235. [DOI] [PubMed] [Google Scholar]
- 25.Rambaut A, Bromham L. Mol Biol Evol. 1998;15:442–448. doi: 10.1093/oxfordjournals.molbev.a025940. [DOI] [PubMed] [Google Scholar]
- 26.Beard K C, Sige B, Krishtalka L. C R Acad Sci. 1992;314:735–741. [Google Scholar]
- 27.McKenna M C, Bell S K. Classification of Mammals Above the Species Level. New York: Columbia Univ. Press; 1997. [Google Scholar]
- 28.Maddison W P, Maddison D R. MACCLADE: Analysis of Phylogeny and Character Evolution. Sunderland, MA: Sinauer; 1992. [DOI] [PubMed] [Google Scholar]
- 29.Van Valen L. Evol Theory. 1979;4:104–121. [Google Scholar]
- 30.Habersetzer J, Storch G. Naturwissenchaften. 1992;79:462–466. [Google Scholar]
- 31.Fenton M B, Audet D, Obrist M K, Rydell J. Paleobiology. 1995;21:229–242. [Google Scholar]
- 32.Norberg U M. In: Ecological Morphology: Integrative Organismal Biology. Wainwright P C, Reilly S M, editors. Chicago: Univ. Chicago Press; 1994. pp. 205–239. [Google Scholar]
- 33.Fenton M B. Communication in the Chiroptera. Bloomington, IN: Indiana Univ. Press; 1985. [Google Scholar]
- 34.Speakman J R. Symp Zool Soc London. 1993;65:39–63. [Google Scholar]
- 35.Griffin D R, Novick A, Kornfield M. Biol Bull. 1958;115:107–113. [Google Scholar]
- 36.Kirsch J A W, Flannery T F, Springer M S, Lapointe F-J. Aust J Zool. 1995;43:395–428. [Google Scholar]
- 37.Romagnoli M L, Springer M S. J Mammal Evol. 2000;7:259–284. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}