

Abstract
Properties of thyme extracts from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis) were examined. Two oil fractions from each species were obtained by CO2 supercritical fluid extraction. Main compounds presented in the supercritical extracts of the three thyme varieties were 1,8 cineole, thymol, camphor, borneol, and carvacrol. As a cellular model of inflammation/atherogenesis, we use human macrophages derived from THP-1 monocytes and activated by oxidized LDLs. These cells were incubated with the thyme fraction oils, and the productions and gene expressions of the inflammatory mediators TNF-α, IL-1B, IL-6, and IL-10 were determined. Thyme extracts significantly reduced production and gene expression of the proinflammatory mediators TNF-α, IL-1B, and IL-6 and highly increased these parameters on the anti-inflammatory IL-10 cytokine. Changes on production and gene expressions were dose dependent and according to the thyme content of each species. Taken together, these results may suggest that thyme extracts could have anti-inflammatory effects.
1. Introduction
Cardiovascular disease, the leading cause of death in Western countries, is a preeminent health problem worldwide [1]. Atherosclerosis, a Chronic Inflammatory-Based Disease (CIBD), constitutes the single most important contributor for cardiovascular complications [2]. Mainly, atherosclerosis results from an immune response to oxidized low-density lipoproteins (LDLs) [3]. Induced by an atherogenic diet, monocytes are promoted to adhesion into the artherial endothelium and intimae (diapedesis). Once in the intimae, monocytes differentiate into macrophages and then modified lipoproteins (such as oxidized LDLs) are accumulated as cytoplasmatic droplets. This process becomes macrophage into foam cells that characterize the early atherosclerotic lesion [4]. Macrophages and foam cells produce inflammatory mediators responsible for the lesion growth. Some of these mediators are involved in proatherogenic processes, such as interleukin- (IL-) 1, IL-6, and Tumor Necrosis Factor alpha (TNF-α) that involve in the upregulation of the molecular adhesion on the endothelial cells. Others have demonstrated to have antiatherogenic properties such as IL-10 that involve in the attenuation of the monocyte differentiation into macrophages [3].
The use of medicinal plants, or extracts from them, has been traditionally practiced worldwide in the prevention and treatment of several chronic diseases such as cardiovascular diseases, intestinal inflammatory diseases, inflammatory bowel disease, arthritis, diabetes, allergies, multiple sclerosis, Parkinson's and Alzheimer's diseases, and others [5]. A Mediterranean aromatic plant, thyme, has been frequently used for those purposes. Extracts from thyme have been used in traditional medicine for the treatment of several respiratory diseases like asthma and bronchitis [6] and for the treatment of other pathologies thanks to several properties such as antiseptic, antispasmodic, antitussive antimicrobial, antifungal, antioxidative, and antiviral [7, 8]. Thyme oils have also been described as a strong bactericide against Gram-positive and Gram-negative bacteria and also as a bronchospasmolytic [9, 10]. However, too many little is known about thyme anti-inflammatory activity. For example, it has been reported that thyme oil reduces NO production in J774A.1 murine macrophages [11]. The biological activity of medicinal plants strongly depends on their composition. Major bioactive compounds of the extracts from thyme are carvacrol and thymol [12, 13]. Thymol exhibits multiple biological activities including anti-inflammatory [14], immunomodulating [15], antioxidant [16], antibacterial [17], antifungal [18], and free radical scavenging properties [19]. Carvacrol also possesses antimicrobial, antifungal, and antioxidant activities [20–22], as well as antimutagenic and anticarcinogenic effects [23]. There have been demonstrated effects on the treatment of colitic mice with essential oils of thyme and oregano containing thymol and carvacrol as their principal bioactive compounds, decreasing levels of the proinflammatory cytokines IL-1β, IL-6, and TNF-α. However, mechanisms mediating these suppressive effects are unclear [8]. Borneol, another compound present in thyme, has been also described as an anti-inflammatory since its dietary supplementation significantly decreases the concentration of the proinflammatory cytokines IL-1β and IL-6 in mice [5].
Nowadays medicinal therapies for CIBDs involve treatment with nonsteroidal anti-inflammatory drugs, antibiotics, corticosteroids, and immunosuppressant, but the application of these drugs is limited due to their toxicity and side effects [24]. Therefore, there is an increasing interest in finding alternative treatments with fewer side effects.
Supercritical fluid extraction (SFE) is considered an attractive extraction method when compared to conventional techniques such as steam distillation or Soxhlet extraction because it avoids solute contamination with solvent residues and the degradation of thermolabile compounds [25]. In this sense, SFE with CO2 is in increasing demand to produce high-quality essential oils from plant material with medicinal properties [26].
During our on-going screening program, designed to identify natural compounds with anti-inflammatory potential, we have studied thyme oils from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis). To determinate whether thyme oils could have immunomodulation properties and could mediate in inflammatory cytokines regulation, we study the effect of our extracts on ox-LDL-activated THP-1 macrophages, measuring the expression and release of several inflammatory mediators.
2. Results and Discussion
CO2 SFE oil fractions composition from three different species of thyme (Thymus vulgaris, Thymus zygis, and Thymus hiemalys) were determined by GC-MS (See Table 1). Main compounds present were thymol, 1,8-cineole, carvacrol, and borneol. Chemical structures of these compounds are shown in Figure 1.
Table 1.
Composition of the supercritical extracts of the three thyme varieties obtained in separators 1 and 2, and the contribution of each compound to the total chromatographic area. N-I: nonidentified compound; R.I.: linear retention index; n.d.: non detected.
| Compound | Ret. time (min) | R.I. | Thymus hyemalis | Thymus zygis | Thymus vulgaris | |||
|---|---|---|---|---|---|---|---|---|
| Separator 1 | Separator 2 | Separator 1 | Separator 2 | Separator 1 | Separator 2 | |||
| % Area | % Area | % Area | % Area | % Area | % Area | |||
| P-cymene | 12.93 | 1022 | 2.97 | 2.77 | 4.77 | 2.98 | 4.90 | 5.57 |
| 1,8 cineole | 13.30 | 1029 | 20.97 | 19.39 | 1.03 | n.d. | 1.29 | 0.95 |
| Gamma-terpinene | 14.89 | 1056 | 0.81 | 0.67 | 0.7 | 0.5 | 1.14 | 1.11 |
| Sabinene | 15.37 | 1065 | 2.52 | 1.97 | n.d. | 0.45 | 1.37 | 0.92 |
| Linalool | 17.39 | 1099 | 3.73 | 1.57 | 6.30 | 4.22 | 6.61 | 3.72 |
| Camphor | 19.74 | 1140 | 27.44 | 19.63 | 1.47 | 1.04 | 1.49 | 0.83 |
| Borneol | 21.03 | 1163 | 8.33 | 8.43 | 4.71 | 4.01 | 3.35 | 2.60 |
| Alpha-terpineol | 22.51 | 1188 | 3.96 | 3.80 | 0.66 | 0.82 | 0.81 | 0.45 |
| Bornyl acetate | 28.18 | 1282 | 1.67 | 1.24 | n.d. | 0.71 | 0.81 | 0.75 |
| Thymol | 28.63 | 1290 | 10.05 | 23.01 | 70.86 | 75.04 | 65.43 | 71.15 |
| Carvacrol | 29.21 | 1299 | 0.64 | 1.34 | 4.20 | 4.61 | 4.26 | 4.30 |
| Isobornyl propionate | 34.62 | 1372 | 1.21 | 0.87 | n.d. | n.d. | n.d. | n.d. |
| E-caryophyllene | 37.78 | 1412 | 4.05 | 3.04 | 2.01 | 1.25 | 4.51 | 2.22 |
| N-I | 39.63 | 1433 | 0.09 | n.d. | 1.84 | 2.98 | 2.63 | 4.49 |
| Germacrene-D | 43.13 | 1474 | 1.73 | 1.26 | n.d. | n.d. | n.d. | n.d. |
| Gamma-elemene | 44.52 | 1490 | 1.97 | 1.55 | n.d. | n.d. | n.d. | n.d. |
| Elemol | 49.37 | 1552 | 1.15 | 1.46 | n.d. | n.d. | n.d. | n.d. |
| Spathulenol | 51.21 | 1576 | 2.42 | 2.85 | n.d. | n.d. | n.d. | n.d. |
| Caryophyllene oxide | 51.42 | 1579 | 1.34 | 0.90 | 1.4 | 1.04 | 1.33 | 0.87 |
| N-II | 56.52 | 1683 | 2.77 | 4.15 | n.d. | 0.28 | n.d. | n.d. |
Figure 1.
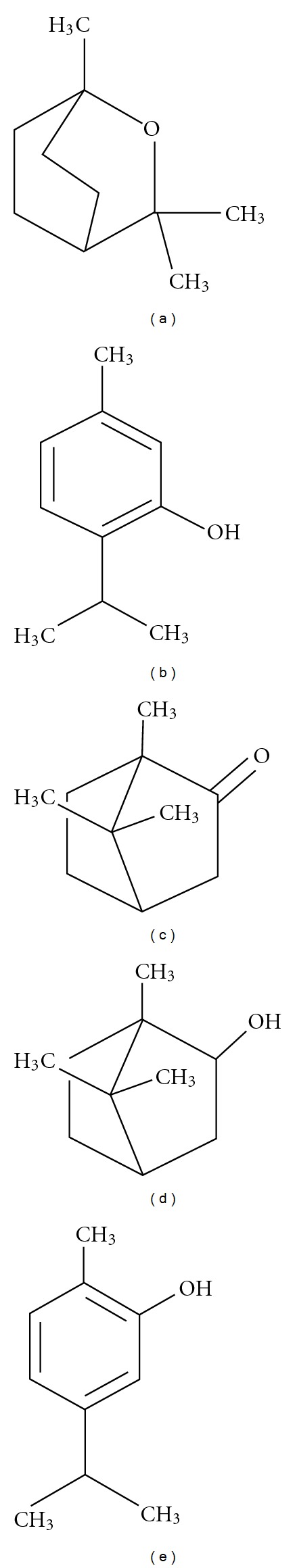
Chemical structures of the main compounds present in the supercritical extracts of the three thyme varieties: (a) 1,8 cineole, (b) thymol, (c) camphor, (d) borneol, and (e) carvacrol.
The cytotoxicity effect on THP-1 macrophages of S1 and S2 CO2 supercritical fluid extracts from thyme leaves was evaluated before the bioactivity study. MTT assay was performed for periods of 24 of incubation (Figure 2). After 24 hours of incubation neither extracts reduced significantly cell viability for concentrations from 5 to 25 μg/mL.
Figure 2.
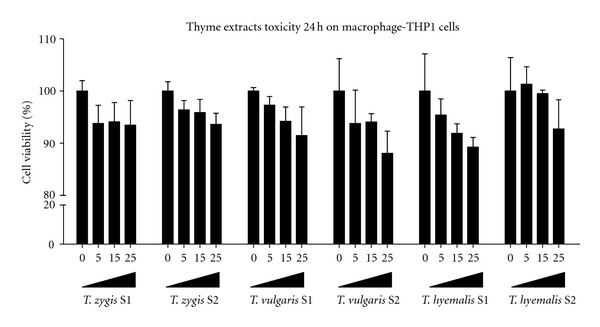
Effects of thyme extracts from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis), on macrophage-THP-1 cell viability. Cells were treated with increasing concentrations of thyme extracts, from 0 to 25 μg/mL for 24 h. Cell viability was determined by the MTT assay. Values represent the mean ± SEM of six independent experiments. No statistic signification in P values lower than 0.05 was found.
To activate the THP-1 macrophage, Cu2+-oxidized LDLs (ox-LDL) were added to the incubation medium. These ox-LDL-treated cells showed an increase in total protein secreted (data not shown). The increase in protein secretion was used as an indicator for macrophage activation [3]. Treatment of activated cells with thyme fractions results in an overall reduction of proinflammatory cytokines release, TNFα, IL-1β, and IL-6 (Figures 3 and 4), in a dose-dependent manner. To test anti-inflammatory effects of oil extracts, one group was treated with a small amount of diclofenac (5 μg/mL). It was carried out as positive standard control [27, 28]. Extracts in general show a better anti-inflammatory effect than diclofenac at this small concentration, diminishing more the proinflammatory cytokines (TNF-α and IL-1β) than diclofenac did and inducing anti-inflammatory release of IL-10 that was not observed with diclofenac.
Figure 3.
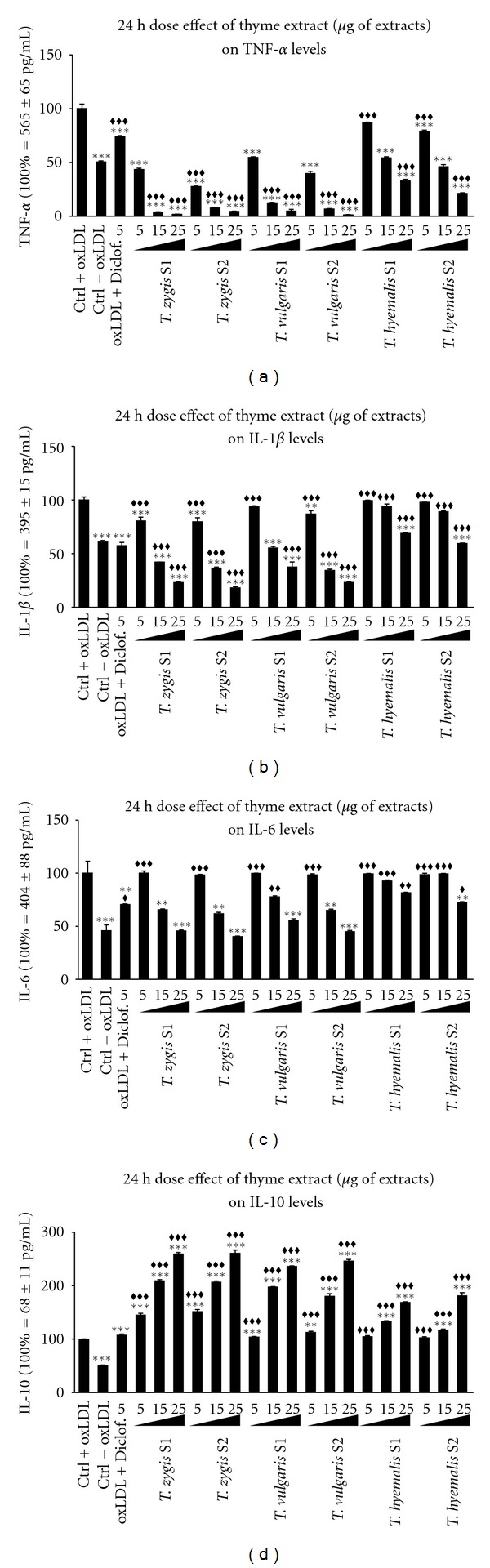
Dose effect of 5, 15, and 25 μg/mL of thyme extracts S1 and S2 from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on the production and secretion of TNF-α (a), IL-1β (b), IL-6 (c), and IL-10 (d) on macrophage-THP-1 cells. Cells were differentiated with (PMA) 100 ng/mL for 24 h and afterwards treated as described in Section 3 for 24 hours. Data represent means ± SEM calculated from six independent experiments with 3 replications for each treatment. Statistic Dunnett's multiple comparison test versus Ctrl +oxLDL signification is represented by *P values less than 0.05, significant; **P values less than 0.01, very significant; and ***P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control. Statistic Bonferroni multiple comparison test versus Ctrl −oxLDL signification is represented by ♦♦P values less than 0.01, very significant and ♦♦♦P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control.
Figure 4.
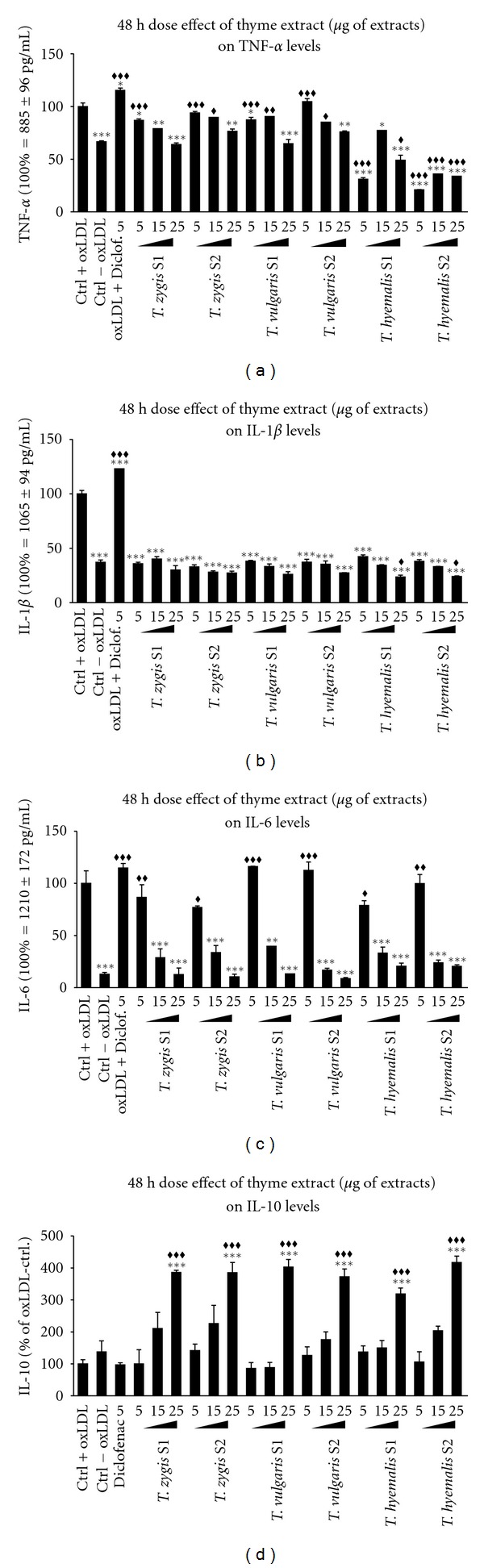
Dose effect of 5, 15, and 25 μg/mL of thyme extracts S1 and S2 from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on the production and secretion of TNF-α (a), IL-1β (b), IL-6 (c), and IL-10 (d) on macrophage-THP-1 cells. Cells were differentiated with (PMA) 100 ng/mL for 24 h and afterwards treated as described in Section 3 for 48 hours. Data represent means ± SEM calculated from six independent experiments with 3 replications for each treatment. Statistic Dunnett's multiple comparison test versus Ctrl +oxLDL signification is represented by *P values less than 0.05, significant; **P values less than 0.01, very significant; and ***P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control. Statistic Bonferroni multiple comparison test versus Ctrl −oxLDL signification is represented by ♦♦P values less than 0.01, very significant; and ♦♦♦P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control.
After 24 hours of incubation, activated cells treated with any of the thyme fractions showed a very significant decrease in TNF-α release when compared with nontreated cells. Both fractions of Thymus zygis and Thymus vulgaris had similar effects on TNF-α secretion. For fraction concentrations of 15 μg/mL and higher, the reduction of TNF-α was such that these cytokine levels were much lower than the nonactivated basal levels. For Thymus hyemalis, TNF-α secretion was lower than the nonactivated controls for fraction concentrations of 25 μg/mL. Despite the large reduction of TNF-α secretion at 24 hours treatments, treatment with fractions of Thymus zygis and Thymus vulgaris for 48 hours showed a lesser reduction of TNF-α. For these fractions, only concentrations of 25 μg/mL induced a very significant reduction of TNF-α levels, equal to nonactivated basal levels. At 48 hours, Thymus hyemalis induced larger TNF-α release inhibition, with all fractions concentration showing a decrease under the nonactivated basal levels.
Regarding IL-1β release at 24 hours, treatment of activated cells with any of the thyme fractions induced a decrease of this cytokine secretion in a dose-dependent manner (Figure 3(b)). Both fractions of Thymus zygis or Thymus vulgaris induced similar reduction of IL-1β. 24-hour treatment with 15 μg/mL of these fractions reduced IL-1β secretion significantly to the nonactivated cells basal secretion. After 48-hour incubation with these thyme fractions, IL-1β concentration in the medium was the same as the basal secretions of nonactivated cells. IL-1β concentration did not decrease when thyme fraction concentrations were increased. Regarding Thymus hyemalis, at 24-hour treatments, both fractions of this species induced similar cell responses. There is a decrease in IL-1β secretion with the increase of fraction concentration, although this IL-1β decrease is only significant till 25 μg/mL incubations. After 48 hours, incubations with these two fractions reduced IL-1β secretion to nonactivated basal levels in the same manner as Thymus zygis and Thymus vulgaris did.
IL-6 secretion was reduced significantly when activated cells were incubated with any of the fractions of either Thymus zygis or Thymus vulgaris at a concentration of 15 μg/mL or higher. This reduction on IL-6 secretion was dose dependent and was verified on both 24 and 48 hours of incubations. Incubations with 25 μg/mL of these extracts reduced IL-6 secretion to nonactivated cells basal levels. 48-hour treatment with Thymus hyemalis, induced the same decrease as Thymus zygis and Thymus vulgaris, whereas at 24 hours, there was no significant reduction of IL-secretion at the concentrations used.
Thyme had opposite effects on the anti-inflammatory cytokine IL-10. 24-hour treatment with any of the fractions induced an increase on IL-10 secretion in a dose-dependent manner; higher fraction concentration induced higher IL-10 secretion. Again, Thymus zygis and Thymus vulgaris were more effective than Thymus hyemalis. Thymus zygis and Thymus vulgaris increased significantly IL-10 secretion of activated cells, with both fractions and at any of the concentrations used. As for Thymus hyemalis, the increase in IL-10 secretion was lower and only significant when the fractions were 15 μg/mL or higher concentrations. 48-hour treatments with higher concentrations of the fractions induced an increase on IL-10 secretion; the rest concentrations did not induce significant increases.
In all experiments, Thymus hyemalis had shown to be less effective than Thymus vulgaris and Thymus zygis, either in increasing or reducing cytokine release. A dose-dependent effect was observable in treatments with 24 and 48 hours of incubation. Changes on production were dose dependentent and according to the thymol content of each species (Table 1).
Relative quantification (RQ) determinates the change in expression of a nucleic acid sequence relative to a control. RQ values represented in Figures 5 and 6 are equivalent to mRNA expression. Endogenous control represented in this figures was 18S rRNA; similar results were showed using GAPDH (data not shown).
Figure 5.
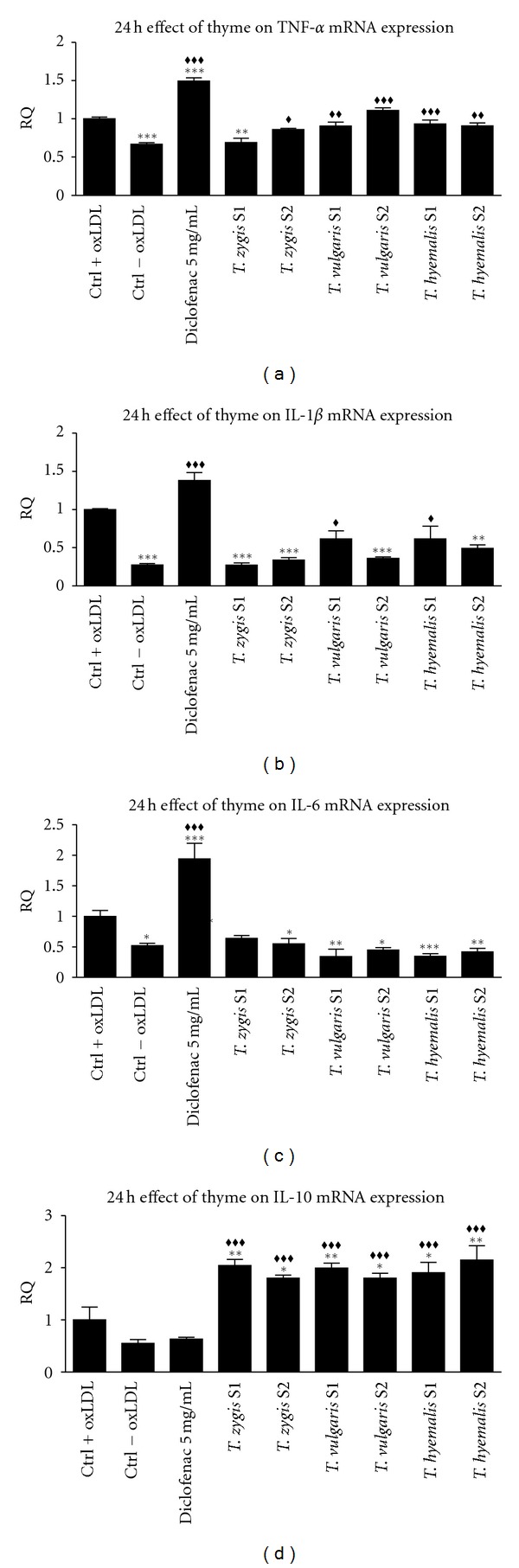
Effect of 25 μg/mL of thyme extracts S1 and S2 from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on the relative quantification (RQ) of TNF-α gene expression (a), IL-1β (b), IL-6 (c), and IL-10 (d) on macrophage-THP-1 cells. Cells were differentiated with (PMA) 100 ng/mL for and afterwards treated as described in Section 3 for 24 hours. Data represent means ± SEM calculated from six independent experiments with 3 replications for each treatment. Statistic Dunnett's multiple comparison test versus Ctrl +oxLDL signification is represented by *P values less than 0.05, significant; **P values less than 0.01, very significant; and ***P values less than 0.001, extremely significant. Statistic Bonferroni multiple comparison test versus Ctrl −oxLDL signification is represented by ♦♦P values less than 0.01, very significant; and ♦♦♦P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control.
Figure 6.
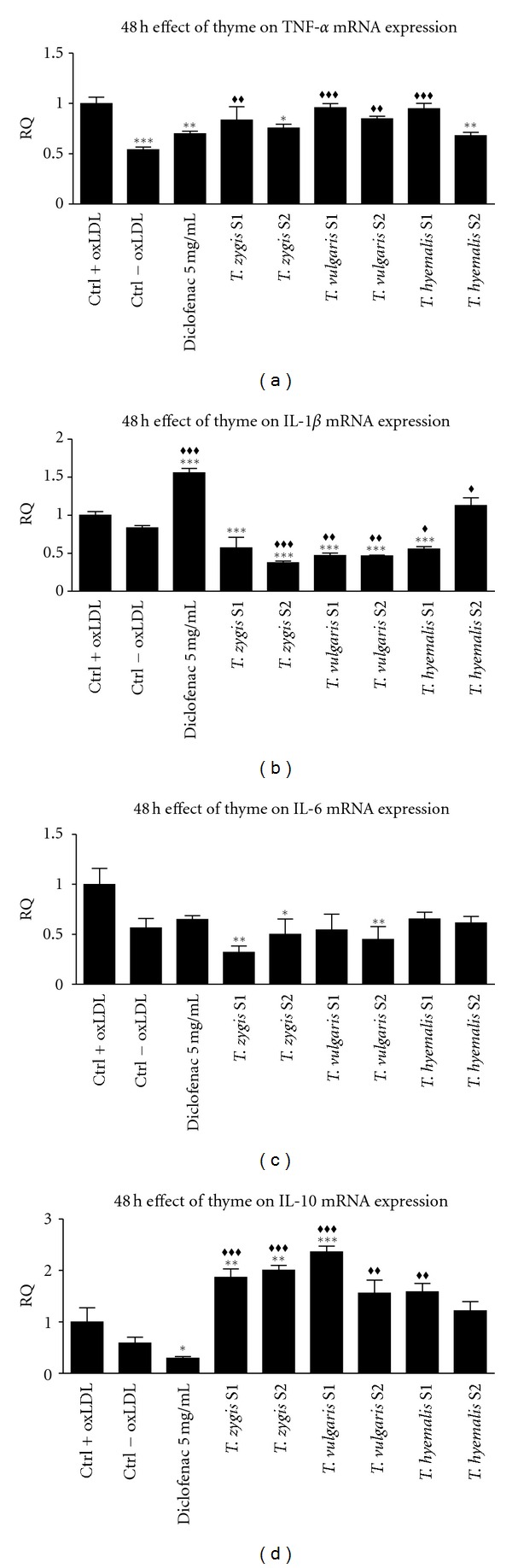
Effect of 25 μg/mL of thyme extracts S1 and S2 from three different species (Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on the relative quantification (RQ) of TNF-α gene expression (a), IL-1β (b), IL-6 (c), and IL-10 (d) on macrophage-THP-1 cells. Cells were differentiated with (PMA) 100 ng/mL for 24 h and afterwards treated as described in Section 3 for 48 hours. Data represent means ± SEM calculated from six independent experiments with 3 replications for each treatment. Statistic Dunnett's multiple comparison test versus Ctrl +oxLDL signification is represented by *P values less than 0.05, significant; **P values less than 0.01, very significant; and ***P values less than 0.001, extremely significant. Statistic Bonferroni multiple comparison test versus Ctrl −oxLDL signification is represented by ♦♦P values less than 0.01, very significant; and ♦♦♦P values less than 0.001, extremely significant. One group was treated with a small concentration of diclofenac (5 μg/mL) as positive standard control.
Similar to the observed cytokine release, gene expression of analyzed cytokines, but not IL-10, increased in oxLDL-activated cells compared with nonactivated cells in treatments during 24 hours. 48-hour treatments caused an increase only in TNFα expression on oxLDL-activated cells compared with nonactivated cells.
TNFα gene expression decreases only in cells treated with Thymus zygis S1 at 24 hours of incubation; other treatments did not change expression of this gene with respect to oxLDL-activated cells. Similar to 24 hours of incubation, at 48 hours TNF-α gene expression did not change with extract treatments; only Thymus hyemalis S2 caused significant reduction of this cytokine gene expression.
Expression of IL-1β decreased in cells treated with all thyme extracts with respect to activated cells at 24 hours of incubation. At 48 hours of incubation, all treatments, except Thymus hyemalis S2, caused reduction in gene expression. For Thymus zygis S2, Thymus vulgaris S1, and S2 and Thymus hyemalis S1, expression of IL-1β was lower than the nonactivated controls cells.
IL-6 gene expression at 24-hour treatment with any of thyme extracts was reduced until level of nonactivated control cells which expression was decreased to half compared to activated cells. In contrast, at 48-hour treatment, expression was reduced using only Thymus zygis S1 and S2 and Thymus vulgaris S2 extracts with respect to activated cells.
IL-10 expression gene increased twice with all thyme extracts at 24 hours of treatment compared with oxLDL-activated cells. At 48 hours, gene expression in cells treated with Thymus zygis S1 and S2 and Thymus vulgaris S1 extracts increased significantly, but Thymus vulgaris S2 and Thymus hyemalis did not change expression of IL-10 compared to oxLDL-activated cells.
Cytokines are considered to be key players in the inflammatory response involved in atherosclerosis and other Chronic Inflammatory-Based Diseases (CIBDs). Among these, Interleukin (IL)-1β, IL-6, IL-10, and Tumor Necrosis Factor-α (TNF-α) are expressed in atherosclerotic lesions by endothelium cells, macrophages, and smooth muscle cells [29, 30]. Some of them are involved in proatherogenic processes, such as upregulation of adhesion molecules on the endothelial cells, while others were proved to have an antiatherogenic role like attenuating the differentiation of monocytes in macrophages. Both IL-1 and IL-6 enhance the inflammatory process. The imbalanced expression of cytokines has been implicated in the progression of many diseases including CIBDs [31]. Cytokines exhibit both beneficial and pathologic effects on their target cells and are produced by many cell types [32]. Several natural compounds are known for their beneficial properties to some diseases or their derived complications and particularly concerning anti-inflammatory effects.
In our experiments, were observed significant cellular responses elicited by the treatment of THP-1 cells with thyme fractions. The action of thyme fractions appears to involve the expression of the proinflammatory cytokine: TNF-α, IL-6, and IL-1β and the anti-inflammatory cytokine IL-10. The most interesting observation made during these studies was that treatment of oxLDL-activated THP-1 cells with thyme oils had different effects on proinflammatory and anti-inflammatory expression: TNF-α, IL-6, and IL-1β expressions were inhibited while IL-10 expression was enhanced. Proinflammatory cytokines levels decreased in a dose-dependent manner with any thyme fraction from Thymus vulgaris, Thymus zygis, or Thymus hyemalis used (after 24 or 48 h of incubation). These results were in agreement with the ones regarding the expression of cytokines genes at 24 hours of incubation. Other authors have previously reported an increment in cytokine secretion in activated macrophages treated with ox-LDL [3, 33]. Similar results have been described for essential oils extracted from Cinnamomum osmophloeum, a herb traditionally used in Asia as food and as a medicine, which contains cinnamaldehyde. Murine macrophages were treated with essential oils from this plant; anti-inflammatory effects by decreasing TNF-α, IL-6, and IL-1β secretions were reported [33].
Main compounds present in supercritical thyme extract were thymol, 1,8 cineole, camphor, borneol, and carvacrol. Anti-inflammatory effect of thymol has been reported on human neutrophiles incubated with 10 or 20 μg/mL of this compound [14]. Mice edema has been reported to be reduced with a topical application of 100 μg/cm2 of carvacrol [34]. Moreover, antioxidant properties of thymol and carvacrol have been demonstrated in several studies, suggesting their use as nutraceutical ingredients in the development of novel functional foods. Derivatives of thymol and carvacrol have been described as antioxidant according to the DPPH radical scavenging method [35–37]. Essential oils of thyme and their components carvacrol and thymol inhibited 3-nitrotyrosine formation, biomarker of the oxidative stress, supporting the nutraceutical value of thyme and the potential of thymol and carvacrol in preventing the formation of toxic products by the action of reactive nitrogen species [36]. Also, thymol and carvacrol prevent autoxidation of lipids [37].
In the same way, the inhibitory activity of 1,8-cineol (eucalyptol) on cytokine production in cultured human lymphocytes and monocytes has been described [38]: particularly, in monocytes, inhibition of TNF-α, IL-1β, IL-6, and IL-8 was 99, 84, 76, and 65%, respectively, when monocytes were treated with 0.15 μg/mL of 1,8-cineol.
Borneol, one of the major compounds present in essential oils from sage, has been described as anti-inflammatory since its dietary supplementation significantly decreases the concentration of the proinflammatory cytokines IL-1β and IL-6 in mice [5].
Results observed in this work suggest that supercritical thyme S1 and S2 fraction oils from Thymus vulgaris, Thymus zygis, and Thymus hyemalis may act as effective inhibitors of oxLDL-induced proinflammatory cytokines (TNF-α, IL-1β, and IL-6) secretion, and also as enhancers of the anti-inflammatory cytokine IL-10 secretion, in macrophage THP-1 cells. In summary, CO2 supercritical thyme extracts showed anti-inflammatory properties by (a) reducing the release of proinflammatory cytokines, and (b) increasing the anti-inflammatory secretion in activated macrophages. These results may suggest that essential oils from thyme extracts could be used as novel options for treatment of chronic diseases based on inflammatory processes. However, numerous and in-depth studies should be carried out for this purpose.
3. Experimental Section
3.1. Plant Materials and Preparation of the CO2 Supercritical Fluid Extraction Fraction
3.1.1. Extraction of Plant Material
Dried and cryogenic grinded leaves from three varieties of thymes (Thymus hyemalis, Thymus zygis, and Thymus vulgaris) were subjected to supercritical fluid extraction with CO2. The supercritical extractions were carried out in a pilot-plant-scale supercritical fluid extractor (Thar Technology, Pittsburgh, PA, USA, model SF2000) of 2 L capacity using pure supercritical CO2 at a pressure of 300 bar and a temperature of 40°C. Extracts from the three thyme species were fractionated using a two-cascade depressurized system and samples were collected in each of the two (separators 1 and 2) separators. Fractionation conditions were as follows: separator 1 was kept at constant pressure and temperature of 15 MPa and 40°C, respectively, whereas separator 2 was maintained at a pressure of 2 MPa and a temperature of 40°C.
3.1.2. GC-MS Analysis
Composition of the supercritical thyme extracts was carried out by a GC-2010 (Shimadzu, Japan), equipped with a split/splitless injector, electronic pressure control, AOC-20i autoinjector, GCMS-QP2010 Plus mass spectrometer detector, and a GCMS Solution software. The column used was a ZB-5 (Zebron) capillary column, 30 m × 0.32 mm I.D, and 0.25 μm phase thickness. Helium, 99.996%, was used as a carrier gas at a flow of 1 mL/min. Oven temperature programming was 60°C isothermal for 4 min, increased to 64°C at 1°C/min, and then increased to 106°C at 2.5°C/min. Oven temperature was then increased from 106°C to 130°C at 1°C/min, then to 200°C at 5°C/min, and then to a final temperature of 250°C/min at 8°C/min which was kept constant for 10 min. Sample injections (1 μL) were performed in split mode (1 : 20). The inlet pressure of the carrier gas was 57.5 KPa. Injector temperature was of 250°C and MS ion source and interface temperatures were 230 and 280°C, respectively. The mass spectrometer was used in TIC mode, and samples were scanned from 40 to 500 m/z units. Compounds thymol, carvacrol, borneol, and linalool were identified by comparison with standard mass spectra obtained in the same conditions and compared with the mass spectra from library Wiley 229. Rests of the compounds were identified by comparison with the mass spectra from Wiley 229 library and by their linear retention index. The chromatographic method was to be based on the previously described by Jordán et al. [39].
3.1.3. Cell Culture
Human THP-1 monocyte cell lines (American Type Culture Collection, ATCC) were maintained in suspension in RPMI 1640 culture medium (ATCC) supplemented with 10% FBS (GIBCO), 100 U/mL penicillin (GIBCO), 100 mg/mL streptomycin (GIBCO), 0.05 mM β-mercaptoethanol (Sigma-Aldrich), and 2 mM L-glutamine (GIBCO), at a density of 3–9 × 105 cells/mL at 37°C in 5% air 95% CO2. Cells were discarded and replaced by frozen stocks every 15 passages.
3.1.4. Cell Differentiation
Cells were pelleted via centrifugation and assessed for viability using the Trypan-blue exclusion method. Viable cells were plated at a density of 5 × 105 cells/mL in 24 wells plates (100 μL and 1 mL, resp.) and incubated with Phorbol 12-Myristate, 13-Acetate (PMA) 100 ng/mL (Sigma-Aldrich) for 48 h in FBS-free medium. Afterwards, the wells were washed with PBS and the treatment initiated.
3.1.5. Cytotoxicity Assay
Oil extracts toxicity was assessed using the mitochondrial-respiration-dependent 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT) reduction method. THP-1 cells were plated in 96-well plates, differentiated and incubated with different concentrations of extract for 24 and 48 hours at 37°C in 5% CO2. After treatment, the cells were washed with PBS and incubated with MTT 1 mg/mL in PBS for 2 hours at 37°C in 5% CO2. Afterwards, formazan crystals produced from MTT by the mitochondrial hydrolase of the viable cells were solubilized in lysis buffer (10% SDS in 50% dimetilformamida pH = 7). The absorbance of each well was then read at 540 nm using a microplate reader (Sunrise Remote, Tecan). The optical density of formazan formed in control cells (without treatment with extract) was taken as 100% viability.
3.1.6. Bioactivity Assay
Oil thyme extracts were dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) to stock concentration of 10 mg/mL determined as the maximum doses not cytotoxic by the cell viability assays. THP-1 cells were differentiated in 24-well plates. After differentiation, the cells were washed with PBS and treated with or without Cu2+ oxidized LDLs to activated or not activated them. Then, cells were incubated with the corresponding thyme extract diluted in FBS-free medium, for 24 or 48 hours at 37°C in 5% CO2. Afterwards, the supernatant was frozen and RNA isolated. Aliquots were analyzed to determine secreted cytokines.
3.1.7. Quantification of Cytokines
Supernatants were centrifuged at 12,000 rpm to remove debris and then stored at −80°C until cytokine analysis. IL-10, IL-1β, IL-6, and TNF-α were quantified using ELISA kits from BD Biosciences, according to the manufacturer's instructions. 100 μL of 1 : 10 diluted medium was added to anticytokine antibody-coated polystyrene wells and incubated for 2 h. After washing, the plates were incubated with biotin-labeled secondary antibody for 1 hour. The plates were washed and incubated for 30 min in the dark with substrate solution. Stop solution was added and the absorbance read at 450 nm with λ correction at 570 nm using a microplate reader (Sunrise Remote, Tecan Austria GmbH, Grödig, Austria).
3.1.8. Total RNA Isolation
Total RNA from THP-1 cells was isolated using the Trizol reagent from Invitrogen. 5 × 105 cells were homogenized in 200 μL of Trizol reagent and, if necessary, stored at −80°C. Following homogenization, samples were left to rest at room temperature for 5 minutes. After 40 μL of chloroform was added, the tubes were vigorously shaken for 15 seconds and let rest at room temperature for 5 minutes. Tubes were then centrifuged at 12000 g, 4°C for 15 minutes. The aqueous (upper and colorless) phase was transferred to a new tube. 100 μL of isopropyl alcohol was added to the aqueous phase; the tube was then gently mixed and incubated at room temperature for 10 minutes. After incubation, samples were centrifuged at 12000 g, 4°C for 10 minutes. A gel-like pellet was formed and the isopropyl alcohol was removed. The pellet was washed with 200 mL of 75% Ethanol in DEPC-treated H2O and centrifuged at 7600, 4°C for 5 min. The ethanol was then removed and the pellet was let dry until colorless. Total RNA was then dissolved in 15 μL of DEPC H2O, incubated at 55°C for 10 minutes, and stored at −80°C for future use.
3.1.9. Gene Expression Quantification
Total RNA isolated from THP-1 cells was quantification of IL-1β, IL-6, IL-10, TNF-α, 18sRNA, and GAPDH gene expression using real-time PCR. 10 ng/μL total RNA was used as template for cDNA synthesis using the High-Capacity Archive Kit from Applied Biosystems, according to the manufacturer's instructions. Real-time PCR was performed using Taqman Probes (Applied Biosystems) following the manufacturer's recommendations in an AB7900 HT Fast Real-Time PCR system (Applied Biosystems). The Taqman probes used were Hs99999029_m1 for IL-1β, Hs00174131_m1 for IL-6, Hs999999035_m1 for IL-10, Hs00174128_m1 for TNF-α, Hs99999901_s1 for 18S rRNA, and Hs99999905_m1 for GAPDH. Gene expression quantification was determined using delta-delta CT method with correction for values of amplification efficiency and normalized to 18S rRNA expression. SDS 2.2.2 software was used. Comparison with GAPDH as endogenous worked out similar results (data not shown).
3.1.10. Statistical Analysis
All data were expressed as the mean ± SEM. For single variable comparisons, Student's t-test was used. For multiple variable comparisons, data were analyzed by one-way analysis of variance (ANOVA) followed by Dunnett's multiple comparison test and Bonferroni test when necessary using the GraphPad Prism statistical software (GraphPad Software Inc. Windows Version 5). P values less than 0.05 were considered significant.
Abbreviations
- oxLDLs:
Oxidized low-density lipoproteins
- TNF:
Tumor necrosis factor.
References
- 1.World-Healt-Organization WHO Cardiovascular diseases. 2009, http://www.who.int/cardiovascular_diseases/en/
- 2.Ross R. Mechanisms of disease: atherosclerosis—an inflammatory disease. The New England Journal of Medicine. 1999;340(2):115–126. doi: 10.1056/NEJM199901143400207. [DOI] [PubMed] [Google Scholar]
- 3.Østerud B, Bjørklid E. Role of monocytes in atherogenesis. Physiological Reviews. 2003;83(4):1069–1112. doi: 10.1152/physrev.00005.2003. [DOI] [PubMed] [Google Scholar]
- 4.Levula M, Jaakkola O, Luomala M, Nikkari ST, Lehtimäki T. Effects of oxidized low- and high-density lipoproteins on gene expression of human macrophages. Scandinavian Journal of Clinical and Laboratory Investigation. 2006;66(6):497–508. doi: 10.1080/00365510600836651. [DOI] [PubMed] [Google Scholar]
- 5.Juhás S, Cikos S, Czikková S, et al. Effects of borneol and thymoquinone on TNBS-induced colitis in mice. Folia Biologica. 2008;54(1):1–7. doi: 10.14712/fb2008054010001. [DOI] [PubMed] [Google Scholar]
- 6.Alonso JR. In: Tratado de Fitomedicina. Bases Clínicas y Farmacológicas. Alonso JR, editor. Buenos Aires, Argentina: Isis Ediciones; 1998. pp. 670–677. [Google Scholar]
- 7.Soliman KM, Badeaa RI. Effect of oil extracted from some medicinal plants on different mycotoxigenic fungi. Food and Chemical Toxicology. 2002;40(11):1669–1675. doi: 10.1016/s0278-6915(02)00120-5. [DOI] [PubMed] [Google Scholar]
- 8.Bukovska A, Cikos S, Juhas S, Il'kova G, Rehak P, Koppel J. Effects of a combination of thyme and oregano essential oils on TNBS-induced colitis in mice. Mediators of Inflammation. 2007;2007:p. 9. doi: 10.1155/2007/23296. Article ID 23296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Van Den Broucke CO, Lemli JA. Spasmolytic activity of the flavonoids from Thymus vulgaris. Pharmaceutisch Weekblad. 1983;5(1):9–14. doi: 10.1007/BF01959645. [DOI] [PubMed] [Google Scholar]
- 10.Essawi T, Srour M. Screening of some Palestinian medicinal plants for antibacterial activity. Journal of Ethnopharmacology. 2000;70(3):343–349. doi: 10.1016/s0378-8741(99)00187-7. [DOI] [PubMed] [Google Scholar]
- 11.Vigo E, Cepeda A, Gualillo O, Perez-Fernandez R. In-vitro anti-inflammatory effect of Eucalyptus globulus and Thymus vulgaris: nitric oxide inhibition in J77RA.1 murine macrophages. Journal of Pharmacy and Pharmacology. 2004;56(2):257–263. doi: 10.1211/0022357022665. [DOI] [PubMed] [Google Scholar]
- 12.Hudaib M, Speroni E, Di Pietra AM, Cavrini V. GC/MS evaluation of thyme (Thymus vulgaris L.) oil composition and variations during the vegetative cycle. Journal of Pharmaceutical and Biomedical Analysis. 2002;29(4):691–700. doi: 10.1016/s0731-7085(02)00119-x. [DOI] [PubMed] [Google Scholar]
- 13.Tian H, Lai D-M. Analysis on the volatile oil in Origanum vulgare. Zhong Yao Cai. 2006;29(9):920–921. [PubMed] [Google Scholar]
- 14.Braga PC, Dal Sasso M, Culici M, Bianchi T, Bordoni L, Marabini L. Anti-inflammatory activity of thymol: inhibitory effect on the release of human neutrophil Elastase. Pharmacology. 2006;77(3):130–136. doi: 10.1159/000093790. [DOI] [PubMed] [Google Scholar]
- 15.Suzuki Y, Furuta H. Stimulation of guinea pig neutrophil superoxide anion-producing system with thymol. Inflammation. 1988;12(6):575–584. doi: 10.1007/BF00914319. [DOI] [PubMed] [Google Scholar]
- 16.Aeschbach R, Löliger J, Scott BC, et al. Antioxidant actions of thymol, carvacrol, 6-gingerol, zingerone and hydroxytyrosol. Food and Chemical Toxicology. 1994;32(1):31–36. doi: 10.1016/0278-6915(84)90033-4. [DOI] [PubMed] [Google Scholar]
- 17.Didry N, Dubreuil L, Pinkas M. Activity of thymol, carvacrol, cinnamaldehyde and eugenol on oral bacteria. Pharmaceutica Acta Helvetiae. 1994;69(1):25–28. doi: 10.1016/0031-6865(94)90027-2. [DOI] [PubMed] [Google Scholar]
- 18.Segvi Klari M, Kosalec I, Masteli J, Piecková E, Pepeljnak S. Antifungal activity of thyme (Thymus vulgaris L.) essential oil and thymol against moulds from damp dwellings. Letters in Applied Microbiology. 2007;44(1):36–42. doi: 10.1111/j.1472-765X.2006.02032.x. [DOI] [PubMed] [Google Scholar]
- 19.Fujisawa S, Kadoma Y. Effect of phenolic compounds on the polymerization of methyl methacrylate. Dental Materials. 1992;8(5):324–326. doi: 10.1016/0109-5641(92)90108-o. [DOI] [PubMed] [Google Scholar]
- 20.Bozin B, Mimica-Dukic N, Samojlik I, Jovin E. Antimicrobial and antioxidant properties of Rosemary and Sage (Rosmarinus officinalis L. and Salvia officinalis L., Lamiaceae) essential oils. Journal of Agricultural and Food Chemistry. 2007;55(19):7879–7885. doi: 10.1021/jf0715323. [DOI] [PubMed] [Google Scholar]
- 21.Horosova K, Bujnakova D, Kmet V. Effect of oregano essential oil on chicken Lactobacilli and E. coli. Folia Microbiologica. 2006;51(4):278–280. doi: 10.1007/BF02931812. [DOI] [PubMed] [Google Scholar]
- 22.Alma MH, Mavi A, Yildirim A, Digrak M, Hirata T. Screening chemical composition and in vitro antioxidant and antimicrobial activities of the essential oils from Origanum syriacum L. growing in Turkey. Biological and Pharmaceutical Bulletin. 2003;26(12):1725–1729. doi: 10.1248/bpb.26.1725. [DOI] [PubMed] [Google Scholar]
- 23.Arcila-Lozano CC, Loarca-Pina G, Lecona-Uribe S. Oregano: properties, composition and biological activity. Archivos Latinoamericanos de Nutricion. 2004;54(1):100–111. [PubMed] [Google Scholar]
- 24.Martínez-Montiel MP, Muñoz-Yagüe MT. Biologic therapies for chronic inflammatory bowel disease. Revista Espanola de Enfermedades Digestivas. 2006;98(4):265–291. doi: 10.4321/s1130-01082006000400006. [DOI] [PubMed] [Google Scholar]
- 25.Almeida PFS. Crossover pressure for SFE of spearmint (Mentha spicata L.) essential oil with pure CO2 and with CO2 plus ethanol. In: Proceedings of the 1st Iberoamerican Conference on Supercritical Fluids (Prosciba '07); 2007. [Google Scholar]
- 26.Mukhopadhyay M. Natural Extracts Using SupercriticalCarbon Dioxide. Boca Raton, Fla, USA: CRC Press; 2000. p. 3. [Google Scholar]
- 27.Kim SK, Kim HJ, Choi SE, Park KH, Choi HK, Lee MW. Anti-oxidative and inhibitory activities on nitric oxide (NO) and prostaglandin E2(COX-2) production of flavonoids from seeds of Prunus tomentosa Thunberg. Archives of Pharmacal Research. 2008;31(4):424–428. doi: 10.1007/s12272-001-1174-9. [DOI] [PubMed] [Google Scholar]
- 28.Berg J, Fellier H, Christoph T, Grarup J, Stimmeder D, et al. The analgesic NSAID lornoxicam inhibits cyclooxygenase (COX)-1/-2, inducible nitric oxide synthase (iNOS), and the formation of interleukin (IL)-6 in vitro. Inflammation Research. 1999;48(7):369–379. doi: 10.1007/s000110050474. [DOI] [PubMed] [Google Scholar]
- 29.Ikeda U, Ikeda M, Seino Y, Takahashi M, Kano S, Shimada K. Interleukin 6 gene transcripts are expressed in atherosclerotic lesions of genetically hyperlipidemic rabbits. Atherosclerosis. 1992;92(2-3):213–218. doi: 10.1016/0021-9150(92)90280-t. [DOI] [PubMed] [Google Scholar]
- 30.Uyemura K, Demer LL, Castle SC, et al. Cross-regulatory roles of interleukin (IL)-12 and IL-10 in atherosclerosis. Journal of Clinical Investigation. 1996;97(9):2130–2138. doi: 10.1172/JCI118650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Arend WP, Gabay C. Cytokines in the rheumatic diseases. Rheumatic Disease Clinics of North America. 2004;30(1):41–67. doi: 10.1016/S0889-857X(03)00115-7. [DOI] [PubMed] [Google Scholar]
- 32.Feghali CA, Wright TM. Cytokines in acute and chronic inflammation. Frontiers in Bioscience. 1997;2:12–26. doi: 10.2741/a171. [DOI] [PubMed] [Google Scholar]
- 33.Chao LK, Hua KF, Hsu HY, Cheng SS, Liu JY, Chang ST. Study on the antiinflammatory activity of essential oil from leaves of cinnamomum osmophloeum. Journal of Agricultural and Food Chemistry. 2005;53(18):7274–7278. doi: 10.1021/jf051151u. [DOI] [PubMed] [Google Scholar]
- 34.Sosa S, Altinier G, Politi M, Braca A, Morelli I, Della Loggia R. Extracts and constituents of Lavandula multifida with topical anti-inflammatory activity. Phytomedicine. 2005;12(4):271–277. doi: 10.1016/j.phymed.2004.02.007. [DOI] [PubMed] [Google Scholar]
- 35.Mastelić J, Jerković I, Blažević I, et al. Comparative study on the antioxidant and biological activities of carvacrol, thymol, and eugenol derivatives. Journal of Agricultural and Food Chemistry. 2008;56(11):3989–3996. doi: 10.1021/jf073272v. [DOI] [PubMed] [Google Scholar]
- 36.Prieto JM, Iacopini P, Cioni P, Chericoni S. In vitro activity of the essential oils of Origanum vulgare, Satureja montana and their main constituents in peroxynitrite-induced oxidative processes. Food Chemistry. 2007;104(3):889–895. [Google Scholar]
- 37.Yanishlieva NV, Marinova EM, Gordon MH, Raneva VG. Antioxidant activity and mechanism of action of thymol and carvacrol in two lipid systems. Food Chemistry. 1999;64(1):59–66. [Google Scholar]
- 38.Juergens UR, Engelen T, Racke K, Stober M, Gillissen A, Vetter H. Inhibitory activity of 1,8-cineol (eucalyptol) on cytokine production in cultured human lymphocytes and monocytes. Pulmonary Pharmacology and Therapeutics. 2004;17(5):281–287. doi: 10.1016/j.pupt.2004.06.002. [DOI] [PubMed] [Google Scholar]
- 39.Jordán MJ, Martínez RM, Goodner KL, Baldwin EA, Sotomayor JA. Seasonal variation of Thymus hyemalis Lange and Spanish Thymus vulgaris L. essential oils composition. Industrial Crops and Products. 2006;24(3):253–263. [Google Scholar]