

Abstract
Nucleolar size and appearance correlate with ribosome biogenesis and cellular activity. The mechanisms underlying changes in nucleolar appearance and regulation of nucleolar size that occur during differentiation and cell cycle progression are not well understood. Caenorhabditis elegans provides a good model for studying these processes because of its small size and transparent body, well-characterized cell types and lineages, and because its cells display various sizes of nucleoli. This paper details the advantages of using C. elegans to investigate features of the nucleolus during the organism's development by following dynamic changes in fibrillarin (FIB-1) in the cells of early embryos and aged worms. This paper also illustrates the involvement of the ncl-1 gene and other possible candidate genes in nucleolar-size control. Lastly, we summarize the ribosomal proteins involved in life span and innate immunity, and those homologous genes that correspond to human disorders of ribosomopathy.
1. Introduction
Caenorhabditis elegans lives freely in soil. The organism occurs naturally in two sexes; both sexes have five pairs of autosomes [1]. However, hermaphrodites possess one pair of X chromosomes (XX), while males have a single X chromosome (XO). Mature adults are about 1 mm in length and 80 μm in diameter; they contain approximately 1,000 somatic cells comprising the animal's minimal systems, including a hypodermis, muscular system, nervous system, digestive organ, and reproductive organ (Figure 1(a)). The life cycle of C. elegans takes about 3 days to complete at 25°C and comprises an embryonic stage, four larval stages (L1 to L4), and adulthood [2]. Adult hermaphrodites can produce about 300 genetically identical progeny by self-fertilization. Progeny carrying various genetic combinations are obtainable by crossing hermaphrodites and males from two genetically different backgrounds [3].
Figure 1.
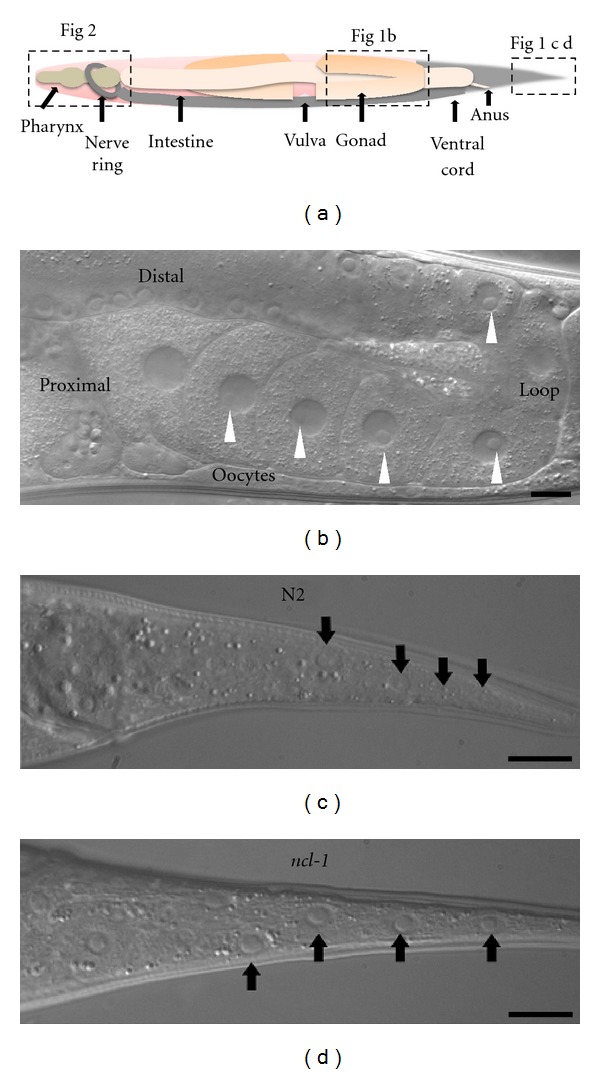
Diagram and Nomarski micrographs of adult worm structures. (a) An illustration of the hermaphrodite adult worm showing the major systems and organs, not in precise proportions. Rectangular boxes indicate parts taken using Nomarski optics as shown in (b) (gonad), (c, d) (tail), and Figure 2 (head region). (b) A section of a gonad arm under the light microscope. Relative positions in the gonad are indicated as distal, loop, and proximal. White arrowheads indicate the nucleoli in the corresponding cells, germ cells, and oocyte. Note that the nucleolus in the -1 oocyte is absent, and that the diameter ratio of nucleolus to nucleus decreases from germ cells to oocytes. The scale bar represents 10 μm; (c, d) the tail section showing hypodermal cell nucleoli (indicated by arrows). N2 is a wild-type worm and Ncl-1 is a mutant with loss of NCL-1 function. Scale bars indicate 10 μm.
The C. elegans genome was fully sequenced in 1998 [4] and seven more complete nematode genomes (including Caenorhabditis briggsae, Caenorhabditis angaria, Pristionchus pacificus, and Trichinella spiralis) are now available [5, 6], providing an excellent resource for comparative genomic studies. Genetics continues to be an important tool for studying gene function in C. elegans. Many genetic mutants and knockout strains are available from the Caenorhabditis Genetics Center (CGC, University of Minnesota) for gene function and genetic epistasis experiments [7]. Additionally, whole-genome RNAi (RNA interference) clone libraries are available, providing a convenient and efficient way to study gene function genome wide [8]. Although C. elegans is a simple organism, more than 50% of its genes have human homologues, and many essential biological activities are conserved between humans and C. elegans [9]. Internet resources for worm research are available, including WormBase (http://www.wormbase.org/), WormBook (http://www.wormbook.org/), and WormAtlas (http://www.wormatlas.org/).
The invariant cell linage and the transparent body of C. elegans allows viewing of all 959 somatic cells of hermaphrodites, using Nomarski differential interference contrast (DIC) optics with a light microscope; fluorescent signals are readily detected with a fluorescence microscope for a reporter gene (green fluorescence protein, gfp) observations. The nucleolus is an easily recognizable subcompartment within the nucleus of germline cells and tail hypodermal cells (Figures 1(b)–1(d)). While electron microscopic studies of whole worm sections have been carried out (see WormAtlas), it remains unclear whether the worm nucleolus contains a fibrillar center (FC), a dense fibrillar center (DFC), and a granular component (GC) as discreet regions, as found in higher eukaryotes. In the C. elegans genome, there are approximately 1400 genes known to produce functional noncoding RNA (ncRNA) transcripts. These include about 275 ribosomal RNA (rRNA) genes and around 100 small nucleolar RNA (snoRNA) genes [4, 10–12]. Approximately 55 copies of a 7.2 kb tandem repeat of rDNA is located at the end of chromosome I, which are transcribed to pre-rRNA and then processed into primary 18S, 5.8S, and 26S rRNA subunits. About 110 copies of a 1 kb rDNA unit located on chromosome V are transcribed into a 5S rRNA [4, 13–17].
C. elegans has approximately 75 genes coding for the small and large subunits of ribosomal proteins, and hundreds of genes coding for nonribosomal proteins that either play a role in ribosome biogenesis, or are of unknown function in the nucleolus. C. elegans contains more than 1,000 operons, which are transcribed into polycistronic mRNA and trans-spliced into two to eight mature mRNAs for translation [18]. Genes arranged in operons are ubiquitous, and may be transcribed in all types of cells, explaining why a high percentage of operons is involved in ribosome biogenesis [19]. For example, the operon CEOP5428 encodes fibrillarin (FIB-1) and RPS-16 [20]. FIB-1 is a methyltransferase for pre-rRNA processing and modification, and RPS-16 is a ribosomal protein of the small subunit. Many genes coding important nucleolar proteins found in humans are also conserved in worms, though they are not arranged in operons, examples are Nopp140 (dao-5) and nucleostemin (nst-1). Nopp140 serves as a scaffold protein, binding to rDNA and acrocentric α-satellite DNA in the human nucleolus; it is required for maintenance of nucleolus integrity [21]. Loss of dao-5 in worms results in growth retardation (C. C. Lee and S. J. Lo, unpublished data). The NST-1 protein belongs to a conserved family of nucleolar GTPases and functions to export pre-60S ribosomal subunits from the nucleolus [22, 23]. Global loss of NST-1 results in a larval arrest phenotype; while loss of NST-1 in germline results in animals displaying germline stem cell proliferation arrest [22]. A few other genes that encode nucleolar or nucleolar-associated proteins in humans, such as B23 (nucleophosmin) and coilin (a hallmark protein of the Cajal body), are not found in worms.
Many human cancer cells exhibit an increase in both nucleolar size and number because of the high demand for ribosomes in the rapidly dividing cells. This hallmark feature provides a marker for pathological diagnosis [24, 25]. Mutations in the cMyc oncogene and tumor suppressor genes p53 and Rb are associated with alteration of nucleolar structure [26, 27]. The ncl-1 gene (abnormal NuCLeolus) was found to regulate nucleolar size in C. elegans, functioning similarly to the p53 and Rb tumor suppressors in vertebrates [28]. The ncl-1 gene is a useful tool for mosaic analysis because of the enlarged nucleolus in the ncl-1 mutant, and because it acts cell autonomously [29]. In worms, NCL-1 is a homologue of Brat (brain tumor) in Drosophila [30]. Absence or alteration of the brat gene causes overproliferation of neuron cells and enlarged nucleoli in flies [31]. However, while ncl-1 mutations in worms cause enlargement of the nucleoli in all cells, mutations do not induce proliferation of neurons [28].
The association of nucleolar size with genetic content has long been observed in tomato plants [32], and nucleolar-size changes occur in response to seasonal environmental cues and nutrition uptake in fish and experimental rats [33, 34]. In mammalian cells, the target of the rapamycin (TOR) pathway plays a nutrition-sensing role by coupling growth factors and nutrients to protein homeostasis [35, 36]. TOR kinase (let-363) in C. elegans is a homologue of TOR complex 1. Mango and colleagues previously demonstrated that let-363 and ruvb-1, members of the TOR pathway, control nucleolar size and promote localization of box C/D snoRNPs to nucleoli [37].
The following sections describe the morphology and variation in nucleoli size in different cells, dynamic patterns of FIB-1 as visualized by fluorescence microscopy, and functionality of nucleoli in C. elegans.
2. Morphology of C. elegans Nucleoli
2.1. Visualization of C. elegans Nucleoli by Nomarski and Fluorescence Microscopy
Nomarski optics (DIC microscopy) provides high-quality images and is commonly used to observe individual cells of C. elegans [38]. The high refractive index provided by DIC microscopy provides micrographs that clearly reveal the nucleoli of germ cells and growing oocytes in a gonad (Figure 1(b)). An adult hermaphrodite gonad is composed of two U-shaped arms classified as distal and proximal ends, based on their position relative to the uterus (Figure 1(a)). The distal arm contains germ cells in the mitotic zone, and cells in various stages of meiosis. The proximal arm contains growing, mature oocytes, and the spermatheca. The germ cells in the syncytium exhibit a high ratio of nucleolus-to-nucleus diameter; the oocytes have a complete plasma membrane with an enlarged nucleus and exhibit a decrease in the ratio of nucleolus to nucleus diameter (Figure 1(b)). The nucleolar size of germ cells, oocytes, and intestinal nucleoli (see below) are relatively large, with diameters ranging from 3 to 7 μm in worms, while the nucleoli of oocytes, adjacent to the spermatheca (designated as -1 oocyte), are undetectable (Figure 1(b)).
The nuclei and nucleoli of the hypodermis at the hermaphrodite tail are easily observable under a DIC microscope (Figures 1(c) and 1(d)) because it is the only region of the worm with a single layer of cells [38]. The nucleoli of these hypodermal cells are intermediate sizes, ranging from 2 μm to 3 μm in diameter. As shown in Figures 1(c) and 1(d), nucleolar size differences in hypodermal cells between wild-type (N2) and ncl-1 mutant worms are easily identifiable [38]. Although NCL-1 is a negative regulator for rRNA biosynthesis [28], it remains unclear how the protein regulates nucleolus size (see below). The ncl-2 (e1896) gene, a mutant with defects in germline nucleoli, has not yet been fully characterized.
Observing and determining the size of the nucleoli of neuronal cells, particularly those within the nerve ring—the equivalent to the brain of higher animals is more difficult because neuronal cells are found in a multiple layer zone and are the smallest nucleoli in worms (Figure 2(a)). The use of transgenic worms that express fibrillarin, one of the most conserved nucleolar proteins, fused with green fluorescence protein FIB-1::GFP facilitates detection of neuronal nucleoli under a fluorescence microscope (Figure 2(b)). Compared with hypodermal cell nucleoli, the nucleoli of neurons in the nerve ring are smaller and less easily distinguished in DIC micrographs (Figures 2(a) and 2(c)). By contrast, the nucleoli of neurons expressing FIB-1::GFP are instantly recognizable under a fluorescence microscope, despite their being less than one micron in diameter (Figure 2(b)). Nucleolar size usually reflects the cell activity of ribosome biogenesis [24, 25]. We speculate that germ cells and intestinal cells may exhibit greater ribosome production activities than neuronal cells do, presumably because neuron function has less need for protein translation than intestinal cells have.
Figure 2.

Comparison of nucleolar appearance of cells in the head region in Nomarski and fluorescence micrographs. A transgenic worm expressing nucleolar protein FIB-1::GFP was photographed using Nomarski and fluorescent microscopy. An arrow indicates the nucleolus of neuronal cells located at the nerve ring (as indicated) while arrowheads indicate the nucleoli of the hypodermis in (a) Nomarski micrograph, (b) fluorescence micrograph, and (c) merged (a, b) images. The scale bar indicates 20 μm.
2.2. ncl-1 in Nucleolar-Size Control in Worms
Transgenic worms expressing FIB-1::GFP in wild-type (N2) and ncl-1 background worms are easily distinguishable because in the former, nucleoli appear as weak GFP foci and are few in number, while the latter have more foci and produce a stronger GFP signal (Figure 3(a)). Absence of NCL-1 causes an increase in FIB-1 expression and an enlargement of nucleoli in the cell; however, the magnitudes of these effects are not the same for each cell (compare the cells indicated by a double arrowhead and single arrowheads in Figure 3(a)). There may be different levels of ncl-1 expression in various tissues, for example, the precursor cells of the intestine are the first cell type to lose NCL-1 during embryogenesis [28]. Another interesting feature is the difference in nucleolar size seen in the hypodermis, adjacent to five ventral neurons in worm larva (Figure 3(b), lower panel). The five ventral cord neurons share the same hypodermis precursor cell (Figure 3(b), upper panel). It would be interesting to know whether the hypodermis displays a decrease of NCL-1, or if the ventral neuron expresses greater levels of NCL-1 immediately after precursor cells have completed asymmetric cell division. A recent study on the ASE gustatory neurons revealed that asymmetric ASE cell size, in which the ASER soma is larger than that of ASEL, is under developmental control [39]. ASER and ASEL neuron nucleoli size and number are also different. These are affected by the fib-1 gene, and directly or indirectly are regulated by die-1, which encodes a Zn finger transcription factor [39]. These findings support the hypothesis that asymmetric cell division during development, may alter expression of regulator factors that control nucleolar size.
Figure 3.
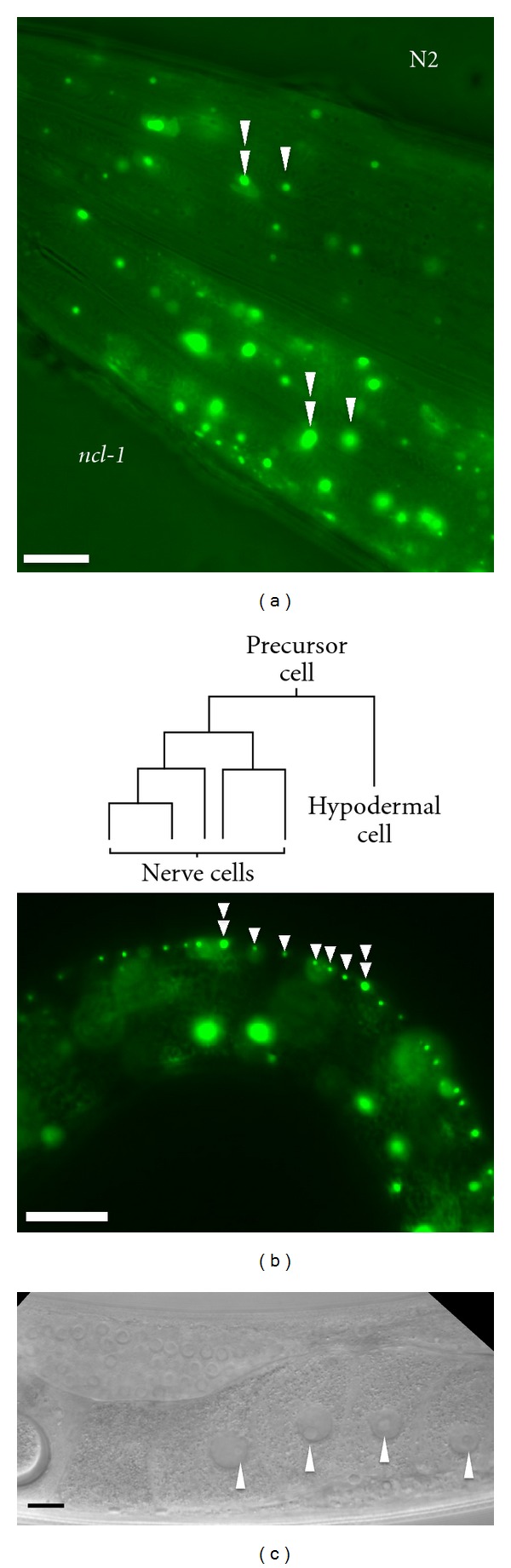
Comparison of the nucleolar size in ncl-1 and N2 worms. (a) N2 and ncl-1 background transgenic worms (as indicated) side by side and photographed under a fluorescence microscope to show the head region. The heads of both worms are facing left. Double arrowheads indicate nucleoli where the nucleolar protein (FIB-1::GFP) expression was less affected by NCL-1 than nucleoli marked by a single arrow. (b) Schematic illustration of ventral cord neurons and hypodermal cell lineages (upper panel). The fluorescence micrograph shows that the hypodermis nucleolar size (indicated by double arrowheads) is larger than nucleoli in neuronal cells (indicated by single arrowheads). The worm head is facing left and the ventral side is at the top. (c) A worm gonad arm-section treated with RNAi against the ncl-1 gene shows a similar appearance to the -1 oocyte nucleolus (arrowhead) of ncl-1 mutants. Scale bars indicate 20 μm.
RNAi gene silencing is performed by feeding worms E. coli, which produces a double-stranded RNA corresponding to the target gene in the worms to be knocked down [8, 40]. N2 worms in which ncl-1 has been silenced by RNAi show enlarged nucleoli and bear the appearance of a nucleolus of the -1 oocytes (Figure 3(c), also compare with Figure 1(b)), a phenotype similar to the ncl-1 mutant animals. The presence of a nucleolus in the -1 oocyte indicates that NCL-1 controls nucleolar size in somatic cells, and regulates nucleolar formation in the -1 oocyte. Since nucleolar size decreases on moving from the -2 oocyte, to the -1 oocyte, and to the ncl-1 worm blastomeres, it is very likely that at least one other gene, besides ncl-1, is a negative regulator of nucleolar size in growing oocytes and blastomeres.
2.3. Dynamic Pattern Changes of FIB-1::GFP in Transgenic Worms
Transgenic worms expressing FIB-1::GFP also provide a tool to study nucleolar pattern changes during embryogenesis and aging. In the 40–60 embryo cell stage, FIB-1::GFP appears as one or two small bright dots (less than 0.2 μ m diameter) in the nucleolar region, and as a faint distribution in the nucleoplasm (Figure 4(a)). This feature is also present in embryos expressing GFP::NST-1 [22], a nucleolar protein involved in maintenance of worm germ cells. Cell division and nucleogenesis are continuous when the embryo is in the active mitotic stage, and so an electron microscopy investigation is needed to determine if the small diameter foci seen for FIB-1::GFP and GFP::NST-1 represent prenucleolar bodies or small nucleoli.
Figure 4.
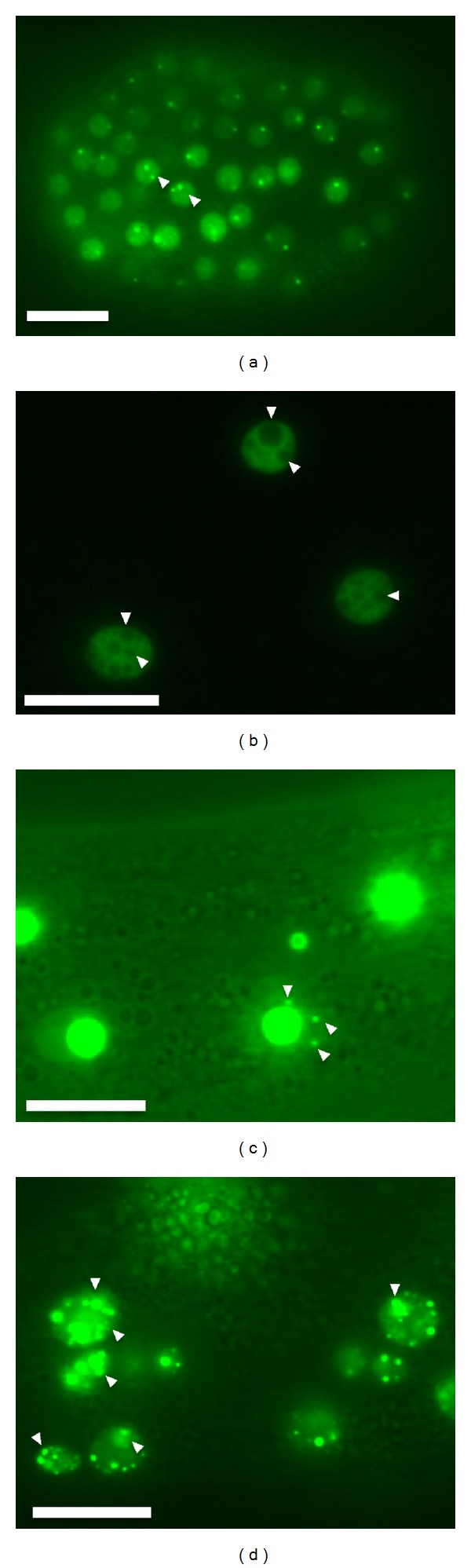
FIB-1::GFP dynamic pattern changes during development. (a) An embryo at the 40–50 cell stage, arrowheads indicate strong dots of nucleoli. Faint GFP outlines are the nucleus. (b) Intestinal cells at the larval stage, arrowheads indicate circles that may represent a GC region. (c) Intestinal cells at the young adult stage, arrowheads indicate GFP foci close to the nucleoli. These may be like the Cajal body or perinucleolar compartment (PNC) in human cells. (d) Intestinal cells of an aged worm, arrowheads indicate FIB-1::GFP fragments, possibly a sign that ribosome biogenesis was aberrant in these cells. The scale bar represents 10 μm.
After the completion of mitotic division, larval intestinal cells appear as a ring structure with FIB-1::GFP occupying a few empty holes (Figure 4(b)). An electron micrograph displaying a similar ring feature, shows a higher intensity of granules in the periphery than at the center (see WormAtlas). Since FIB-1 is located at the fibrillar center (FC) and dense fibrillar center (DFC) but not the granular component (GC) in other eukaryotes, the non-GFP region is probably a GC in worms. In adults, the FIB-1::GFP occupies the entire nucleolus, and occasionally, a small green dot appears near the nucleolus (Figure 4(c)). This small substructure may correspond to the Cajal body or the perinucleolar compartment (PNC) of higher organisms [41–43], although worms do not have a gene homolog for coilin. It is probable that the small focus expressing the FIB-1::GFP marker protein is a Cajal body based on two observations; fibroblast cells derived from the coilin-gene knock-out mouse have Cajal bodies [44], and fibrillarin and Nopp140 are present in the coilin-null Cajal body [45].
Fragmented and irregularly shaped FIB-1::GFP is common in the intestinal nucleoli of aged worms (Figure 4(d)), possibly reflecting reduced ribosome biogenesis activity in the intestinal cells of aged worms. This suggestion is supported by reports that aged worms exhibit decreased expression of lamin A, and that lamin A maintains the functional plasticity of nucleoli [46–48]. Additionally, mammalian cells display fragmented fibrillarin distribution when treated with inhibitors, such as actinomycin D and Aapatone [49, 50].
2.4. Similar Components for Localization of Nucleolar Proteins in Worm and Mammalian Cells
Proteomic analysis reveals that human nucleoli contain at least 700 proteins, most of which are conserved across species [51] (and references therein). To investigate the similarities in molecular function of worm and mammalian cell nucleoli, one can reciprocally introduce nucleolar genes into worms or mammalian cells. Worms transformed by dao-5, a human homologue of Nopp140, fused with GFP show fluoresce in the nucleoli of most cells (Figure 5(a)) [52]. Conversely, the DAO-5 GFP signal produced by HeLa cells transfected with the dao-5::GFP gene is colocalized with fibrillarin in the nucleoli (Figures 5(b)–5(d)), indicating that the DAO-5 nucleolar localization signal sequence (NoLS) is recognizable by both worms and mammalian factors.
Figure 5.
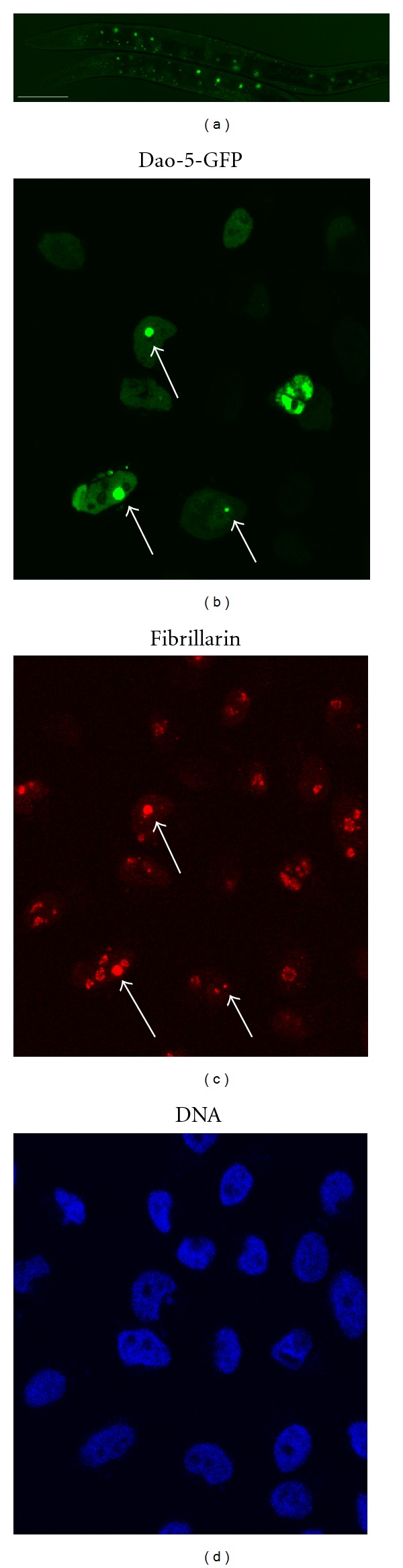
GFP::DAO-5 expression in worms and HeLa cells. (a) A transgenic worm expressing GFP::DAO-5 shows an oval distribution of green fluorescence representing cell nucleoli. (b and c) HeLa cells transfected by plasmids containing gfp::dao-5 and co-stained with anti-fibrillarin antibody show the location of nucleoli expressing GFP fusion protein (arrows). (d) The same field of cells stained by a DNA dye and visualized through a UV fluorescence microscope. The scale bar in (a) indicates 100 μm.
Human hepatitis D virus (HDV) is the smallest known RNA virus. HDV has a genome of approximately 1700 nucleotides that encodes two antigen isoforms named SDAg and LDAg, for the small and large delta antigens, respectively [53]. SDAg is required for genome replication while the LDAg is used for virion packaging [54]. Both antigens can target the nucleoli of human cells, although they also shuttle to other locations when posttranslationally modified [55–58]. LDAg is detectable at the nucleoli of transfected human hepatoma cells by using GFP as a reporter of gene expression (Figures 6(a)–6(c)). When transgenic worms coexpress FIB-1::GFP and RFP::LD using a bicistronic vector [59], LDAg is localized at the nucleoli of most cells by FIB-1::GFP (Figures 6(d)–6(f)). Interestingly, in the presence of FIB-1::GFP, RFP::LD is found at the perimeter of the nucleolus in intestinal cells (Figures 6(g)–6(i)). The localization of LDAg at the edge of the nucleolus is uncommon in mammalian cells, which implies that worm intestinal cells have different components to those of mammals. These examples indicate that both organisms' nucleolar components recognize LDAg and DAO-5 NoLS.
Figure 6.
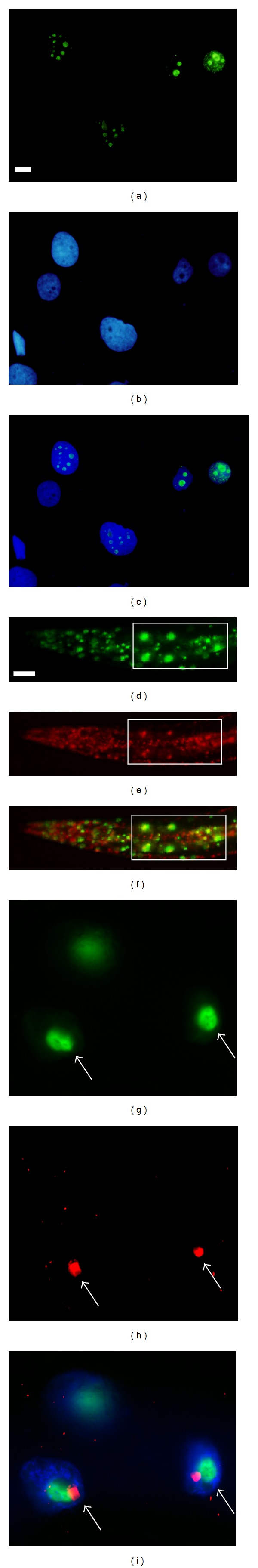
Nucleolar localization of the large delta antigen of human hepatitis D virus in human hepatoma cells and in worms. Human hepatoma cells transfected with plasmids expressing GFP::LDAg, (a) visualized by GFP, (b) stained with DNA dye, and (c) the merged picture. (d–i) A transgenic worm carrying a bicistronic vector coexpressing FIB-1::GFP and RFP::LD. A low magnification of worms photographed via (d) the FITC channel to visualize FIB-1::GFP, (e) the rhodamine channel to visualize RFP::LD, and (f) the merged picture. The rectangles in panels (d–f) show four intestinal cells in the same worm. A high magnification of intestinal cells shows (g) FIB-1::GFP, (h) RFP::LD, and (i) the merged picture. Arrows indicate the nucleoli. Note that RFP::LD and FIB-1::GFP are not perfectly colocalized. Scale bars indicate 10 μm.
3. Functionality of Nucleoli in C. elegans
3.1. Ribosome Biogenesis
As with other eukaryotes, C. elegans nucleolus' main function is as a site for ribosome production, including pre-rRNA transcription, processing, modification, and ribosome assembly [52, 62]. Many methods exist to measure and quantify rRNA biosynthesis activity in cells or organisms. Isotope pulse and chase labeling, and Northern blot analysis are commonly used to reveal dynamic changes during progression of pre-rRNA into mature rRNA. These methods are not applicable at the level of the single cell because of physical limits of detection; in situ demonstration of ribosome biogenesis can be carried out by isotope or BrUTP incorporation to visualize rRNA synthesis activity in individual cells. For example, the signal from BrUTP incorporation into a worm gonad demonstrated that germ cells in the pachytene zone during meiotic division have greater rRNA synthesis activity than germ cells exhibit during the mitotic phase or in the maturing oocyte (Figure 7). Increased rates of ribosome biogenesis occur in the germ cells of many animals in preparation for fast protein-translation during embryogenesis. Currently, the rRNA synthesis activity of individual muscular and neuronal cells in worms remains unclear because of the difficulty of the required analytical techniques.
Figure 7.
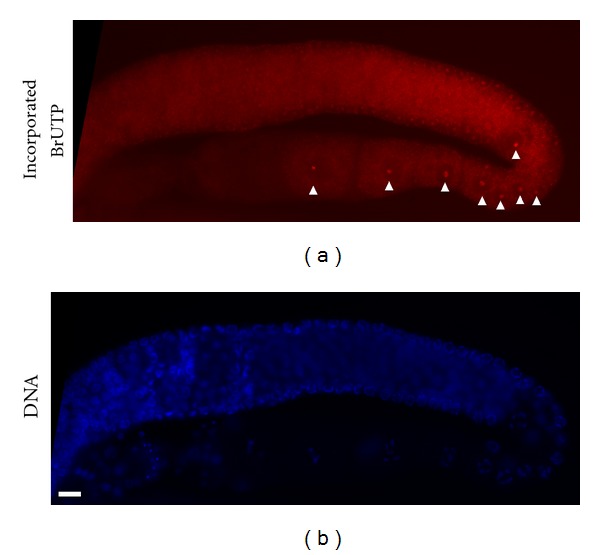
In situ BrUTP incorporation in a gonad to show rRNA synthesis activity. (a) Incorporation of BrUTP was detected by an anti-BrUTP antibody conjugated with fluorescence dye Alexa 594. Arrowheads indicate the signal from BrUTP incorporation. (b) DNA staining of the same gonad. Note the increased BrUTP incorporation in germ cells at the pachytene zone of meiosis. The scale bar indicates 10 μm.
3.2. Worm Ribosome Biogenesis Deficiency Phenotypes
Killian and Hubbard reported the C. elegans “proximal tumor” phenotype, exhibited by the hermaphrodite gonad, this was the first phenotype associated with loss of nucleolar integrity and impaired ribosome biogenesis function [63]. Hubbard mapped this unusual phenotype to pro genes (pro-1, pro-2, and pro-3), which encode factors involved in ribosome biogenesis [63, 64]. RNAi screening against other factors involved in ribosome biogenesis revealed a similar gonadogenesis phenotype, suggesting that such phenotypes can result from inefficient ribosome biogenesis.
In addition to being a ribosome factory, nucleoli have important roles in other cellular processes such as the cell cycle, stress response, and coordination of the biogenesis of other RNPs species [65, 66]. Ribosomal proteins may act independently to interact with other cellular proteins, for example, RPS-16 and RPL-6 interact with LET-756, one of two C. elegans fibroblast growth factors (EGFs), for the regulation of ribosome biogenesis [67]. Interestingly, a mutation in nol-6, which encodes a nucleolar RNA-associated protein, was found to enhance worm innate immunity against bacterial infection [68]. In mammals, the disruption of ribosomal proteins or nucleolar proteins elicits a p53-mediated response [65]. RNAi screening revealed that knock down of any one of 20 small ribosomal proteins results in increased resistance to p53/CEP-1-dependent bacterial infection [68]. Germ cells have been demonstrated to control both innate immunity and lifespan through distinct signaling pathways [69]. It is unclear whether germ cell nucleoli are involved in these activities, nevertheless, depletion of several ribosomal proteins such as RPS-3, RPS-8, and RPS-16 at the postembryonic stage extended worm life expectancy [70, 71].
3.3. Worm Genes Corresponding to Ribosomopathy-Associated Human Disease
Impaired ribosome biogenesis resulting from loss of nucleolar integrity or disruption of rRNA biosynthesis has been described as “nucleolus stress” or “ribosomal stress” [65]. Ribosomopathiesis a clinical pathological term defined as “a collection of disorders in which genetic abnormalities cause impaired ribosome biogenesis and function, resulting in specific clinical phenotypes” [61, 72]. These disorders include Bowen-Conradi syndrome [60], cartilage-hair hypoplasia (CHH) [73], dyskeratosis congenital (DC) [74], Diamond-Blackfan anemia (DBA) [75, 76], Shwachman-Diamond syndrome (SDS) [77], and Treacher-Collin syndrome [78, 79]. Human genes associated with ribosomopathy are also present in C. elegans (Table 1). Most genes encode ribosomal proteins, although some are nucleolar proteins involved in pre-rRNA processing, for example, mrpr-1 encodes for a noncoding RNA. Correlating phenotype with mutations in worms can aid our understanding of the mechanisms of human diseases and so inform drug development for new treatments in the future.
Table 1.
Mutated human genes involved in the diseases of ribosomopathies and their C. elegans homologs.
| Disease | Defective gene* | C. elegans homolog | Chromosomal location | Product and function | Operon |
|---|---|---|---|---|---|
| Bowen-Conradi syndrome | EMG1 | Y39A1A.14 | III | Methyltransferase | |
| Treachere-Collins syndrome | TCOF1 | K06A9.1 | X | Three predicated proteins, containing 825 aa, 1032 aa, and 2232 aa | |
| 5q-syndrome | RPS14 | rps-14 | III | Small ribosomal subunit S14 protein | |
| Cartilage hair hypoplasia | RMRP | mrpr-1 | II | RNA component of the endoribonuclease RNase MRP | |
| Shwachman-Diamond syndrome | SBDS | W06E11.4 | III | Nucleolar protein required for maturation of 60S ribosomal subunits | T19C3.7 |
| Dyskeratosis congenita | DKC1 | K01G5.5 | III | A predicated protein containing 445 aa | |
| Diamond-Blackfan anemia | RPS19, RPS24, RPS17, RPL35, RPL5, RPL11, RPS7, RPL36, RPS15, RPS27A | rps-19 | I | Small ribosomal subunit S19 protein |
4. Conclusion and Prospects
This review illustrates that C. elegans is a useful animal for studying nucleolus biology. C. elegans has a short life cycle and is very easy to handle in the laboratory. Different cell types become available throughout the worms various stages of development from embryo to adult and during aging, and during the various phases of the cell cycle, including meiotic and mitotic division, asymmetric cell division, and postmitotic stages. Nomarski optics allow the study and analysis of changes in nucleolar size, and used in combination with GFP tagging of nucleolar proteins, transgenic worms can provide information about nucleolar activity. RNAi silencing of specific genes is straightforward to perform in worms and many mutant worms are now available, facilitating the study of nucleoli functionality in worms. C. elegans is a good choice to study nucleolus biology during development, and many important problems remain unresolved, for example, investigating how ncl-1 is regulated temporally and spatially, and determining the functions of the Cajal-like bodies that lack coilin, in worms.
Acknowledgments
The authors thank Yi-Chun Wu (National Taiwan University) for assistance in worm research and supplying materials at the beginning period, Hong Zhang (National Institute of Biological Sciences at Beijing) for use of his worm facility during SJL sabbatical at his laboratory, and Scott C. Schuyler (Chang Gung University) for critical reading of the paper. They also thank two anonymous reviewers for their suggestions on the paper, and Nova Medical Editing for proofreading services. Special thanks are due to Po-chiang Chen and Heuy-Jen Lai for providing Figures 3(c), 5(b) and 5(d), respectively, and to CGC (Caenorhabditis Genetics Center) for supplying various worm strains. This paper was supported by Grants from the Chang Gung Memorial Hospital (CMRPD32037I-III and CMRPD18001-3) and the Taiwan Ministry of Education (EMRPD1A0431 and EMRPD1A0881).
References
- 1.Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77(1):71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Riddle DL, Blumenthal T, Meyer BJ, et al. Introduction to C. elegans. 1997. [PubMed] [Google Scholar]
- 3.Fay D. Genetic mapping and manipulation: chapter 1-introduction and basics. WormBook. 2006:1–12. doi: 10.1895/wormbook.1.90.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.The C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: a platform for investigating biology. Science. 1998;282(5396):2012–2018. doi: 10.1126/science.282.5396.2012. [DOI] [PubMed] [Google Scholar]
- 5.Shaye DD, Greenwald I. Ortholist: a compendium of C. elegans genes with human orthologs. PLoS One. 2011;6(5) doi: 10.1371/journal.pone.0020085. Article ID e20085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Blaxter M. Nematodes: the worm and its relatives. PLoS Biology. 2011;9(4) doi: 10.1371/journal.pbio.1001050. Article ID e1001050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Huang LS, Sternberg PW. Genetic dissection of developmental pathways. WormBook. 2006:1–19. doi: 10.1895/wormbook.1.88.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kamath RS, Ahringer J. Genome-wide RNAi screening in Caenorhabditis elegans. Methods. 2003;30(4):313–321. doi: 10.1016/s1046-2023(03)00050-1. [DOI] [PubMed] [Google Scholar]
- 9.Williams CH, Hong CC. Multi-step usage of in vivo models during rational drug design and discovery. International Journal of Molecular Sciences. 2011;12(4):2262–2274. doi: 10.3390/ijms12042262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stricklin SL, Griffiths-Jones S, Eddy SR. C. elegans noncoding RNA genes. WormBook. 2005:1–7. doi: 10.1895/wormbook.1.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hokii Y, Ushida C. Regulation of Gene Expression by Small RNAs. CRC Press; 2009. Isolation and characterization of small RNAs in Caenorhabditis elegans; pp. 101–121. [Google Scholar]
- 12.Wang PPS, Ruvinsky I. Computational prediction of Caenorhabditis box H/ACA snoRNAs using genomic properties of their host genes. RNA. 2010;16(2):290–298. doi: 10.1261/rna.1876210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Albertson DG. Localization of the ribosomal genes in Caenorhabditis elegans chromosomes by in situ hybridization using biotin-labeled probes. The EMBO Journal. 1984;3(6):1227–1234. doi: 10.1002/j.1460-2075.1984.tb01957.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ellis RE, Sulston JE, Coulson AR. The rDNA of C. elegans: sequence and structure. Nucleic Acids Research. 1986;14(5):2345–2364. doi: 10.1093/nar/14.5.2345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Saijou E, Fujiwara T, Suzaki T, Inoue K, Sakamoto H. RBD-1, a nucleolar RNA-binding protein, is essential for Caenorhabditis elegans early development through 18S ribosomal RNA processing. Nucleic Acids Research. 2004;32(3):1028–1036. doi: 10.1093/nar/gkh264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nelson DW, Honda BM. Two highly conserved transcribed regions in the 5S DNA repeats of the nematodes Caenorhabditis elegans and Caenorhabditis briggsae. Nucleic Acids Research. 1989;17(21):8657–8667. doi: 10.1093/nar/17.21.8657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sulston JE, Brenner S. The DNA of Caenorhabditis elegans. Genetics. 1974;77(1):95–104. doi: 10.1093/genetics/77.1.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Blumenthal T, Evans D, Link CD, et al. A global analysis of Caenorhabditis elegans operons. Nature. 2002;417(6891):851–854. doi: 10.1038/nature00831. [DOI] [PubMed] [Google Scholar]
- 19.Blumenthal T, Gleason KS. Caenorhabditis elegans operons: form and function. Nature Reviews Genetics. 2003;4(2):112–120. doi: 10.1038/nrg995. [DOI] [PubMed] [Google Scholar]
- 20.Pi H, Lee LW, Lo SJ. New insights into polycistronic transcripts in eukaryotes. Chang Gung Medical Journal. 2009;32(5):494–498. [PubMed] [Google Scholar]
- 21.Tsai YT, Lin CI, Chen HK, et al. Chromatin tethering effects of hNopp140 are involved in the spatial organization of nucleolus and the rRNA gene transcription. Journal of Biomedical Science. 2008;15(4):471–486. doi: 10.1007/s11373-007-9226-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kudron MM, Reinke V. C. elegans nucleostemin is required for larval growth and germline stem cell division. PLoS Genetics. 2008;4(8) doi: 10.1371/journal.pgen.1000181. Article ID e1000181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Romanova L, Grand A, Zhang L, et al. Critical role of nucleostemin in pre-rRNA processing. Journal of Biological Chemistry. 2009;284(8):4968–4977. doi: 10.1074/jbc.M804594200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Derenzini M, Trere D, Pession A, Govoni M, Sirri V, Chieco P. Nucleolar size indicates the rapidity of cell proliferation in cancer tissues. Journal of Pathology. 2000;191(2):181–186. doi: 10.1002/(SICI)1096-9896(200006)191:2<181::AID-PATH607>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- 25.Zink D, Fischer AH, Nickerson JA. Nuclear structure in cancer cells. Nature Reviews Cancer. 2004;4(9):677–687. doi: 10.1038/nrc1430. [DOI] [PubMed] [Google Scholar]
- 26.Koh CM, Gurel B, Sutcliffe S, et al. Alterations in nucleolar structure and gene expression programs in prostatic neoplasia are driven by the MYC oncogene. American Journal of Pathology. 2011;178(4):1824–1834. doi: 10.1016/j.ajpath.2010.12.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Trere D, Ceccarelli C, Montanaro L, Tosti E, Derenzini M. Nucleolar size and activity are related to pRb and p53 status in human breast cancer. Journal of Histochemistry and Cytochemistry. 2004;52(12):1601–1607. doi: 10.1369/jhc.4A6454.2004. [DOI] [PubMed] [Google Scholar]
- 28.Frank DJ, Roth MB. ncl-1 is required for the regulation of cell size and ribosomal RNA synthesis in Caenorhabditis elegans. Journal of Cell Biology. 1998;140(6):1321–1329. doi: 10.1083/jcb.140.6.1321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hedgecock EM, Herman RK. The ncl-1 gene and genetic mosaics of Caenorhabditis elegans. Genetics. 1995;141(3):989–1006. doi: 10.1093/genetics/141.3.989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Frank DJ, Edgar BA, Roth MB. The Drosophila melanogaster gene brain tumor negatively regulates cell growth and ribosomal RNA synthesis. Development. 2002;129(2):399–407. doi: 10.1242/dev.129.2.399. [DOI] [PubMed] [Google Scholar]
- 31.Arama E, Dickman D, Kimchie Z, Shearn A, Lev Z. Mutations in the β-propeller domain of the Drosophila brain tumor (brat) protein induce neoplasm in the larval brain. Oncogene. 2000;19(33):3706–3716. doi: 10.1038/sj.onc.1203706. [DOI] [PubMed] [Google Scholar]
- 32.Lesley MM. The relation between satellite size and nucleolus size in three races of Solanum lycopersicum. Genetics. 1938;23(5):485–493. doi: 10.1093/genetics/23.5.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lewis CG, Winick M. Studies on ribosomal RNA synthesis in vivo in rat liver during short-term protein malnutrition. Journal of Nutrition. 1978;108(3):329–340. doi: 10.1093/jn/108.3.329. [DOI] [PubMed] [Google Scholar]
- 34.Alvarez M, Quezada C, Molina A, Krauskopi M, Vera MI, Thiry M. Ultrastructural changes of the carp (Cyprinus carpio) hepatocyte nucleolus during seasonal acclimatization. Biology of the Cell. 2006;98(8):457–463. doi: 10.1042/BC20060006. [DOI] [PubMed] [Google Scholar]
- 35.Martin DE, Powers T, Hall MN. Regulation of ribosome biogenesis: where is TOR? Cell Metabolism. 2006;4(4):259–260. doi: 10.1016/j.cmet.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 36.Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell. 2006;124(3):471–484. doi: 10.1016/j.cell.2006.01.016. [DOI] [PubMed] [Google Scholar]
- 37.Mango SE, Sheaffer KL, Updike DL. The target of rapamycin pathway antagonizes pha-4/FoxA to control development and aging. Current Biology. 2008;18(18):1355–1364. doi: 10.1016/j.cub.2008.07.097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yochem JK. Nomarski images for learning the anatomy, with tips for mosaic analysis. WormBook. 2006:1–47. doi: 10.1895/wormbook.1.100.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Goldsmith AD, Sarin S, Lockery S, Hobert O. Developmental control of lateralized neuron size in the nematode Caenorhabditis elegans. Neural Development. 2010;5(1, article 33) doi: 10.1186/1749-8104-5-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Timmons L, Court DL, Fire A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene. 2001;263(1-2):103–112. doi: 10.1016/s0378-1119(00)00579-5. [DOI] [PubMed] [Google Scholar]
- 41.Morris GE. The Cajal body. Biochimica et Biophysica Acta. 2008;1783(11):2108–2115. doi: 10.1016/j.bbamcr.2008.07.016. [DOI] [PubMed] [Google Scholar]
- 42.Gall JG. Cajal bodies: the first 100 years. Annual Review of Cell and Developmental Biology. 2000;16:273–300. doi: 10.1146/annurev.cellbio.16.1.273. [DOI] [PubMed] [Google Scholar]
- 43.Pollock C, Huang S. The perinucleolar compartment. Journal of Cellular Biochemistry. 2009;107(2):189–193. doi: 10.1002/jcb.22107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tucker KE, Berciano MT, Jacobs EY, et al. Residual Cajal bodies in coilin knockout mice fail to recruit Sm snRNPs and SMN, the spinal muscular atrophy gene product. Journal of Cell Biology. 2001;154(2):293–307. doi: 10.1083/jcb.200104083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Verheggen C, Mouaikel J, Thiry M, et al. Box C/D small nucleolar RNA trafficking involves small nucleolar RNP proteins, nucleolar factors and a novel nuclear domain. The EMBO Journal. 2001;20(19):5480–5490. doi: 10.1093/emboj/20.19.5480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gruenbaum Y, Lee KK, Liu J, Cohen M, Wilson KL. The expression, lamin-dependent localization and RNAi depletion phenotype for emerin in C. elegans. Journal of Cell Science. 2002;115(part 5):923–929. doi: 10.1242/jcs.115.5.923. [DOI] [PubMed] [Google Scholar]
- 47.Haithcock E, Dayani Y, Neufeld E, et al. Age-related changes of nuclear architecture in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(46):16690–16695. doi: 10.1073/pnas.0506955102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Martin C, Chen S, Maya-Mendoza A, Lovric J, Sims PFG, Jackson DA. Lamin B1 maintains the functional plasticity of nucleoli. Journal of Cell Science. 2009;122(part 10):1551–1562. doi: 10.1242/jcs.046284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chen M, Jiang P. Altered subcellular distribution of nucleolar protein fibrillarin by actinomycin D in HEp-2 cells. Acta Pharmacologica Sinica. 2004;25(7):902–906. [PubMed] [Google Scholar]
- 50.Jamison JM, Gilloteaux J, Perlaky L, et al. Nucleolar changes and fibrillarin redistribution following apatone treatment of human bladder carcinoma cells. Journal of Histochemistry and Cytochemistry. 2010;58(7):635–651. doi: 10.1369/jhc.2010.956284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ahmad Y, Boisvert FM, Gregor P, Cobley A, Lamond AI. NOPdb: nucleolar proteome database—2008 update. Nucleic Acids Research. 2009;37(1):D181–D184. doi: 10.1093/nar/gkn804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lo SJ, Lee CC, Lai HJ. The nucleolus: reviewing oldies to have new understandings. Cell Research. 2006;16(6):530–538. doi: 10.1038/sj.cr.7310070. [DOI] [PubMed] [Google Scholar]
- 53.Lai MMC. The molecular biology of hepatitis delta virus. Annual Review of Biochemistry. 1995;64:259–286. doi: 10.1146/annurev.bi.64.070195.001355. [DOI] [PubMed] [Google Scholar]
- 54.Taylor JM. Hepatitis delta virus. Virology. 2006;344(1):71–76. doi: 10.1016/j.virol.2005.09.033. [DOI] [PubMed] [Google Scholar]
- 55.Lai MMC. RNA replication without RNA-dependent RNA polymerase: surprises from hepatitis delta virus. Journal of Virology. 2005;79(13):7951–7958. doi: 10.1128/JVI.79.13.7951-7958.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tan KP, Shih KN, Lo SJ. Ser-123 of the large antigen of hepatitis delta virus modulates its cellular localization to the nucleolus, SC-35 speckles or the cytoplasm. Journal of General Virology. 2004;85(part 6):1685–1694. doi: 10.1099/vir.0.19690-0. [DOI] [PubMed] [Google Scholar]
- 57.Shih KN, Lo SJ. The HDV large-delta antigen fused with GFP remains functional and provides for studying its dynamic distribution. Virology. 2001;285(1):138–152. doi: 10.1006/viro.2001.0845. [DOI] [PubMed] [Google Scholar]
- 58.Huang WH, Chen CW, Wu HL, Chen PJ. Post-translational modification of delta antigen of hepatitis D virus. Current Topics in Microbiology and Immunology. 2006;307:91–112. doi: 10.1007/3-540-29802-9_5. [DOI] [PubMed] [Google Scholar]
- 59.Lee LW, Lo HW, Lo SJ. Vectors for co-expression of two genes in Caenorhabditis elegans. Gene. 2010;455(1-2):16–21. doi: 10.1016/j.gene.2010.02.001. [DOI] [PubMed] [Google Scholar]
- 60.Armistead J, Khatkar S, Meyer B, et al. Mutation of a gene essential for ribosome biogenesis, EMG1, causes Bowen-Conradi syndrome. American Journal of Human Genetics. 2009;84(6):728–739. doi: 10.1016/j.ajhg.2009.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Narla A, Ebert BL. Ribosomopathies: human disorders of ribosome dysfunction. Blood. 2010;115(16):3196–3205. doi: 10.1182/blood-2009-10-178129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Boisvert FM, van Koningsbruggen S, Navascues J, Lamond AI. The multifunctional nucleolus. Nature Reviews Molecular Cell Biology. 2007;8(7):574–585. doi: 10.1038/nrm2184. [DOI] [PubMed] [Google Scholar]
- 63.Killian DJ, Hubbard EJA. C. elegans pro-1 activity is required for soma/germline interactions that influence proliferation and differentiation in the germ line. Development. 2004;131(6):1267–1278. doi: 10.1242/dev.01002. [DOI] [PubMed] [Google Scholar]
- 64.Voutev R, Killian DJ, Hyungsoo Ahn J, Hubbard EJA. Alterations in ribosome biogenesis cause specific defects in C. elegans hermaphrodite gonadogenesis. Developmental Biology. 2006;298(1):45–58. doi: 10.1016/j.ydbio.2006.06.011. [DOI] [PubMed] [Google Scholar]
- 65.Boulon S, Westman BJ, Hutten S, Boisvert FM, Lamond AI. The nucleolus under stress. Molecular Cell. 2010;40(2):216–227. doi: 10.1016/j.molcel.2010.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pederson T, Tsai RYL. In search of nonribosomal nucleolar protein function and regulation. Journal of Cell Biology. 2009;184(6):771–776. doi: 10.1083/jcb.200812014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Popovici C, Fallet M, Marguet D, Birnbaum D, Roubin R. Intracellular trafficking of LET-756, a fibroblast growth factor of C. elegans, is controlled by a balance of export and nuclear signals. Experimental Cell Research. 2006;312(9):1484–1495. doi: 10.1016/j.yexcr.2006.01.012. [DOI] [PubMed] [Google Scholar]
- 68.Fuhrman LE, Goel AK, Smith J, Shianna KV, Aballay A. Nucleolar proteins suppress Caenorhabditis elegans innate immunity by inhibiting p53/CEP-1. PLoS Genetics. 2009;5(9) doi: 10.1371/journal.pgen.1000657. Article ID e1000657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Alper S, McElwee MK, Apfeld J, Lackford B, Freedman JH, Schwartz DA. The Caenorhabditis elegans germ line regulates distinct signaling pathways to control lifespan and innate immunity. The Journal of Biological Chemistry. 2010;285(3):1822–1828. doi: 10.1074/jbc.M109.057323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hansen M, Taubert S, Crawford D, Libina N, Lee SJ, Kenyon C. Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans. Aging Cell. 2007;6(1):95–110. doi: 10.1111/j.1474-9726.2006.00267.x. [DOI] [PubMed] [Google Scholar]
- 71.Curran SP, Ruvkun G. Lifespan regulation by evolutionarily conserved genes essential for viability. PLoS Genetics. 2007;3(4, article e56) doi: 10.1371/journal.pgen.0030056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Khan S, Pereira J, Darbyshire PJ, et al. Do ribosomopathies explain some cases of common variable immunodeficiency? Clinical and Experimental Immunology. 2011;163(1):96–103. doi: 10.1111/j.1365-2249.2010.04280.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Williams MS, Hermanns P. Analysis of RPS19 in patients with cartilage-hair hypoplasia and severe anemia: preliminary results. American Journal of Medical Genetics. 2005;138(1):66–67. doi: 10.1002/ajmg.a.30903. [DOI] [PubMed] [Google Scholar]
- 74.Heiss NS, Knight SW, Vulliamy TJ, et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nature Genetics. 1998;19(1):32–38. doi: 10.1038/ng0598-32. [DOI] [PubMed] [Google Scholar]
- 75.Lipton JM, Ellis SR. Diamond Blackfan anemia 2008-2009: broadening the scope of ribosome biogenesis disorders. Current Opinion in Pediatrics. 2010;22(1):12–19. doi: 10.1097/MOP.0b013e328334573b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.McGowan KA, Li JZ, Park CY, et al. Ribosomal mutations cause p53-mediated dark skin and pleiotropic effects. Nature Genetics. 2008;40(8):963–970. doi: 10.1038/ng.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Boocock GRB, Morrison JA, Popovic M, et al. Mutations in SBDS are associated with Shwachman-Diamond syndrome. Nature Genetics. 2003;33(1):97–101. doi: 10.1038/ng1062. [DOI] [PubMed] [Google Scholar]
- 78.Wise CA, Chiang LC, Paznekas WA, et al. TCOF1 gene encodes a putative nucleolar phosphoprotein that exhibits mutations in Treacher Collins Syndrome throughout its coding region. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(7):3110–3115. doi: 10.1073/pnas.94.7.3110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Dixon J, Edwards SJ, Gladwin AJ, et al. Positional cloning of a gene involved in the pathogenesis of Treacher Collins syndrome. Nature Genetics. 1996;12(2):130–136. doi: 10.1038/ng0296-130. [DOI] [PubMed] [Google Scholar]