

Abstract
Dams designed for hydropower and other purposes alter the environments of many economically important fishes, including Chinook salmon (Oncorhynchus tshawytscha). We estimated that dams on the Rogue River, the Willamette River, the Cowlitz River, and Fall Creek decreased water temperatures during summer and increased water temperatures during fall and winter. These thermal changes undoubtedly impact the behavior, physiology, and life histories of Chinook salmon. For example, relatively high temperatures during the fall and winter should speed growth and development, leading to early emergence of fry. Evolutionary theory provides tools to predict selective pressures and genetic responses caused by this environmental warming. Here, we illustrate this point by conducting a sensitivity analysis of the fitness consequences of thermal changes caused by dams, mediated by the thermal sensitivity of embryonic development. Based on our model, we predict Chinook salmon likely suffered a decrease in mean fitness after the construction of a dam in the Rogue River. Nevertheless, these demographic impacts might have resulted in strong selection for compensatory strategies, such as delayed spawning by adults or slowed development by embryos. Because the thermal effects of dams vary throughout the year, we predict dams impacted late spawners more than early spawners. Similar analyses could shed light on the evolutionary consequences of other environmental perturbations and their interactions.
Keywords: anthropogenic change, Chinook salmon, dams, development, embryos, emergence, fry, selection gradients, temperature
Introduction
Although multiple factors have contributed to the decline of Pacific salmon (Oncorhynchus spp.) across portions of their natural range, dams have arguably played a major role in many locations (NRC 1996; Lichatowich 1999; Ruckelshaus et al. 2002). Large dams (>15 m tall)–designed to generate hydropower, control floods, or facilitate navigation – have had unintended side effects on the quantity and quality of habitat used by salmon. Specifically, these dams have decreased the area for spawning and rearing, and have altered flows, sediments, and temperatures downstream (PFMC 1979; Ward and Stanford 1979; Ligon et al. 1995).
The effects of large dams on thermal conditions have been documented worldwide, and include changes in the mean and variance of temperature at several temporal scales (Petts 1984; Crisp 1987; USACE 2000a; Preece and Jones 2002; Steel and Lange 2007). These changes have significant biologic consequences because temperature influences the growth and reproduction of aquatic organisms (Ward and Stanford 1979; Schlosser et al. 2000; Vinson 2001). For salmon, the spawning of adults, the emergence of fry, and the migration of smolts coincide with thermal cues in natal lakes and streams (e.g., Brett 1971; Brannon 1987; Roper and Scarnecchia 1999). Anthropogenic changes in temperature can induce mismatches between life histories and environmental conditions. Such maladaptations could threaten the persistence of populations.
Although changes in thermal regimes might have contributed to the declines of salmon (NRC 1996), we have much to learn about the underlying mechanisms (see reviews in McCullough 1999; and Richter and Kolmes 2005). Unusually high temperatures can delay migration (Hyatt et al. 2003; Goniea et al. 2006), increase mortality (Smith et al. 2003; Crozier and Zabel 2006; Gilhousen 1990), perturb energetics (Brett 1995; Rand and Hinch 1998; Macdonald et al. 2000), slow growth (Bisson and Davis 1976; Pearcy 1992), and create morbidity (Bartholomew 1998; Materna 2001). Indirect effects can also result from shifts in the timing of or phenotypes at life-history transitions (Quinn and Adams 1996). For example, high temperatures during embryonic development can lead to premature emergence, which could expose fry to peak flows, scarce resources, or greater predation (Brannon 1987; Murray and McPhail 1988; Jensen and Johnsen 1999; USACE 2000a).
Since the first large dams in the Pacific Northwest were built in the early 1900s, many generations of salmon have spawned, developed, emigrated, and returned to spawn in the waters downstream. Possibly, these fish have begun to adapt to the new thermal regimes caused by dams (Waples et al. 2008). Although we cannot infer adaptation from the available data, we can estimate the potential evolutionary impacts of dams and discuss the types of data that would be necessary to make such inferences. Under this premise, we present an exploratory analysis of Chinook salmon (Oncorhynchus tshawytscha) in four parts. First, we assess the historical changes in water temperatures downstream of large dams. Second, we review the effects of water temperature on physiological performance at specific life stages. Third, we estimate the degree to which changes in physiological performance would influence the fitness of genotypes. Finally, we assess the possibility of an evolutionary response to selective pressures imposed by dams.
Impacts of dams on water temperature
Quantifying changes in water temperature downstream of a dam requires several types of data, including the location of the dam, the date of construction, and records of temperature. Linking thermal changes to the performance of Chinook salmon, in particular, requires additional knowledge of the distribution of populations. We compiled these data from a variety of sources (Willingham 1983; Myers et al. 1998; USACE 2000b; StreamNet 2005; City of Tacoma 2005; Kimbrough et al. 2006; Herrett et al. 2006; USGS 2007), and then imposed five criteria to identify several dams for our analysis. First, the dam had to reside upstream of a gauge that recorded water temperatures. Second, the gauge’s period of record had to span multiple years before and after the construction of the dam. Third, the dam had to occur on a river or large creek that supported Chinook salmon in Washington, Oregon, California or Idaho (USA). Fourth, the distance between the dam and the gauge had to be <30 km, to minimize the attenuation of thermal effects downstream (USACE 2000a). Finally, no other large dam could exist upstream of the focal dam prior to its construction. Using these criteria, we selected four dams for analysis (Table 1, Fig. 1).
Table 1.
Four dams selected for analysis based, in part, on the existence of water temperature gauges downstream.
| Dam | Type | Location | State | Gauge number* | Dam-gauge distance (km) | Date of operation† | Beginning of record | End of record |
|---|---|---|---|---|---|---|---|---|
| Mayfield | ER | Cowlitz River | WA | 14238000 | 2.3 | 1962 | 1954 | 1982 |
| Fall Creek | HR | Fall Creek | OR | 14151000 | 1.8 | 10/1965 | 1951 | 2007 |
| Hills Creek | HR | Willamette River | OR | 14148000 | 17.9 | 1961 | 1950 | 1987 |
| Lost Creek | MR | Rogue River | OR | 14337600 | 4.5 | 2/1977 | 1970 | 2007 |
Names, types and locations of these dams are provided, along with the approximate dates when the dams became operational (i.e., the reservoirs upstream were created). Gauge identification numbers, distances between gauges and dams, and periods of temperature records are also provided.
ER, epilimnetic release; MR, multiple depth release; HR, hypolimnetic release.
For details, see USGS (2007).
The specific month of first operation was provided when known. If only the year was known, the month was assumed to be January. In all cases, the first day of the month was assumed to be the starting point of the dam’s operation.
Figure 1.
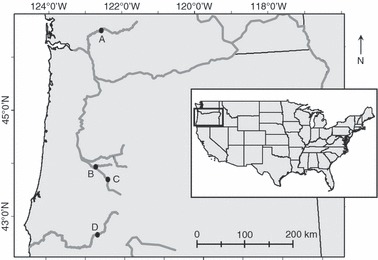
A map of Washington and Oregon, USA, depicting the locations of four dams selected for analysis: (A) Mayfield, (B) Fall Creek, (C) Hill Creek, and (D) Lost Creek.
The gauges downstream of these dams contained two types of gaps in their records of temperature. The first type comprised days when the minimal and maximal temperatures were recorded but the mean temperature was not recorded. For these days, we estimated the mean temperature by averaging the minimal and maximal temperatures. The second type comprised days when both the mean temperatures and extreme temperatures were missing. In these cases, we estimated the means by regression or interpolation (see Appendix A).
We assessed the changes in temperature associated with each dam by comparing daily mean temperatures, averaged among the years before and after construction of the dam. Based on our findings, the effect of a dam on water temperature depends on the time of year (Fig. 2). During summer, water was generally cooler after a dam was built. Conversely, during the fall and early winter, water was generally warmer in the presence of a dam. This reversal–from summer cooling to fall warming–was evident as early as August on the Rogue River and as late as October on the Cowlitz River (Fig. 2).
Figure 2.
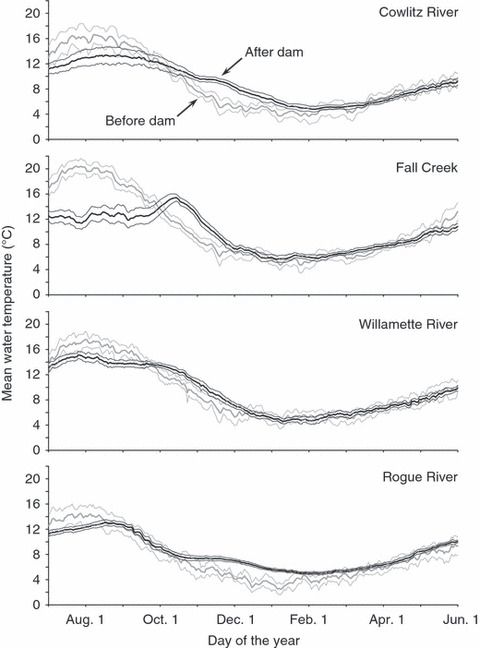
Mean water temperatures before and after dam construction on the Cowlitz River, Fall Creek, the Middle Fork of the Willamette River, and the Rogue River. Mean values were generated by averaging daily mean temperatures among years. Bold lines depicting the mean water temperatures are surrounded by thinner lines depicting the upper and lower bounds of the 95% confidence intervals.
In theory, the effect of a dam on water temperature should depend not only on the time of year, but also on thermal stratification upstream and the depth of water released at the dam (Ward 1982; Crisp 1987; Poff and Hart 2002). If the reservoir upstream is thermally stratified and the dam draws water from below the thermocline (i.e., the hypolimnetic layer), the water downstream of the dam should be unseasonably cold during summer and unseasonably warm during fall and winter. If the dam draws water from above the thermocline (i.e., the epilimnetic layer), the water downstream should be unusually warm during the summer. Other dams draw water from multiple depths, enabling finer control over temperatures downstream. Given that our sample contained three types of dams (Table 1), we were surprised to discover these dams affected water temperatures similarly (Fig. 2).
Consequences of dams for the performance of salmon
A rich literature describes the influence of water temperature on the development, growth, and survival of salmon at various life stages (reviewed by McCullough 1999). Generally, embryonic development proceeds more rapidly at higher temperatures. In Chinook salmon, a doubling of water temperature within tolerable limits will halve the time between the fertilization of an egg and the emergence of a fry (Beacham and Murray 1990). Embryonic survivorship remains high throughout a wide range of temperatures, but decreases precipitously outside that range. For example, the survivorship of Chinook embryos exceeds 90% between 5 and 15°C; however, below 2°C and above 18°C the rate drops to <5% (Fig. 3, see also Tang et al. 1987; Velsen 1987; Beacham and Murray 1989). Once salmon emerge from the gravel, their growth and survival continue to depend on temperature. For example, the growth rate of fry increases between 10 and 15°C (Banks et al. 1971), but survivorship decreases at higher temperatures (Fig. 3, see also Olson et al. 1970). Because these data pertain to salmon raised at constant temperatures, we must assume variation in temperature does not alter these conclusions–a dubious assumption. Some salmon tolerate acute exposure to temperatures that would prove lethal during prolonged exposure (Murray and Beacham 1987; Tang et al. 1987; Campbell et al. 1998), but we need more research to define the general limits of tolerance in variable environments.
Figure 3.
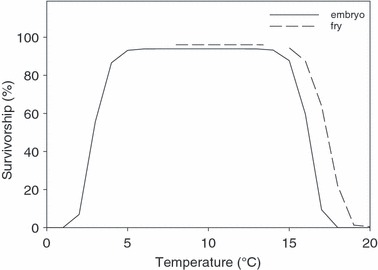
Embryos and fry of Chinook salmon survive well over a range of approximately 10°C. Temperatures >15°C lead to very high mortality. This conclusion was based on models that best fit published data for embryos (Velsen 1987; Beacham and Murray 1989) and fry (Olson et al. 1970). Model selection was performed according to Angilletta (2006); details are provided in Appendix B.
The thermal sensitivities of physiological performances can mediate the direct and indirect effects of dams on the fitness of salmon. Because temperatures in the fall can approach those that stress Chinook salmon (e.g., see Fall Creek in Fig. 2), warming by a dam could directly increase the mortality of embryos. In the laboratory, brief exposure to 17°C during embryonic development caused more than 98% mortality between the fertilization and emergence (Geist et al. 2006). More likely, however, dams indirectly influence fitness by advancing the timing of emergence. Emergence describes a developmental transition in which fry leave the interstitial gravel and enter the water column, thereby initiating a sustained, free-swimming residence in a stream or river. At this point in development, the energy stored as yolk has been virtually exhausted and the juvenile must begin to feed exogenously. Early or late emergence could lead to mortality from excessive flows, abundant predators, or insufficient resources (Jensen and Johnsen 1999; Einum and Fleming 2000). The warming caused by a dam in fall and winter would accelerate the development of embryos, leading to early emergence. Thus, a dam could disrupt a match between the actual and the optimal dates of emergence.
Adaptation of behavior or physiology would ameliorate the thermal effects of a dam. If salmon below the dam spawned later in the year, the temperatures experienced by their offspring would be lower because of seasonal cooling (see Fig. 2). Consequently, development would proceed more slowly and the timing of emergence would correspond to that before the dam’s influence. Alternatively, an evolutionary decrease in developmental rate would enable salmon to delay emergence despite the warm water below the dam. Whether spawning behavior or developmental physiology would experience greater selective pressure depends on the cost of each strategy and the impact on other stages of the life cycle.
The response to selection depends on the heritabilities of spawning behavior and embryonic physiology, and their genetic covariance. The relative timing of spawning seems highly heritable in salmon, with estimates of narrow-sense heritabilities ranging from 0.4 to 1.3 (e.g., see Smoker et al. 1998; Quinn et al. 2000; Dickerson et al. 2005; Neira et al. 2006). We know less about the heritability of thermal physiology in salmon, but additive genetic variance for developmental rate exists under some thermal conditions (Hebert et al. 1998; Kinnison et al. 1998). These data suggest behavior, physiology, or both could evolve in response to thermal change. The genetic covariance between spawning date and developmental rate could enhance or constrain adaptation, depending on its sign and magnitude (Lande and Arnold 1983).
Despite the ideas discussed above, current models of thermal adaptation cannot generate deep insights about the responses of salmon to dams. First, these models focus on physiological responses to environmental temperature (reviewed by Angilletta et al. 2002; Kingsolver and Gomulkiewicz 2003), ignoring the potential for behavioral responses that seem relevant to salmon (e.g., timing of spawning, placement of nests). Second, these models assume that temperature influences either survivorship (Lynch and Gabriel 1987) or fecundity (Gilchrist 1995) uniformly throughout the life of the organism. Because both the thermal impacts of dams and the thermal tolerances of salmon vary throughout the life cycle, simple models will not accurately predict the strength of selection. In the next section, we use an age-structured model to explore the impact of physiological performance on the fitness of a genotype. This approach should yield a better understanding of the selective pressures imposed by dams.
Modeling selective pressures imposed by dams
Matrix models of population growth provide a useful way to quantify the ecological and evolutionary consequences of environmental changes (Caswell 2001), such as the thermal changes caused by dams. Salmon and most other organisms have life cycles with distinct stages that occur in a characteristic seasonal cycle. Dams and other anthropogenic factors typically affect some life stages more than others. Matrix projections enable us to explore how a change in performance at a specific stage impacts the mean fitness of a population.
Matrix models have been applied to salmon by classifying the life cycle in terms of either stage or age (e.g., Kareiva et al. 2000; Greene and Beechie 2004; Scheuerell et al. 2006; Zabel et al. 2006). Ratner et al. (1997) developed an age-classified model for the spring run of Chinook salmon from the South Fork of the Umpqua River (Oregon, USA). In this population, 90% of juveniles migrate to the ocean during their first spring (Ratner et al. 1997). Therefore, the survivorship from oviposition to age 1 equals the product of survivorships for the embryo, fry, and smolt. Adults return to breed at age 3, 4 or 5. Thus, the matrix form of the model for a female is as follows:
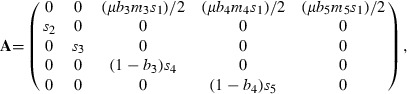 |
where sx equals the survivorship from age x-1 to age x, bx equals the propensity to breed at age x, mx equals the fecundity at age x, and μ equals the survivorship of adults during their migration to the spawning area.
The leading eigenvalue of this matrix (λ) represents the long-term rate of population growth (Caswell 2001). For the case represented by matrix A, we cannot solve explicitly for λ, but we can calculate this rate numerically given a set of parameter values. Using values for Chinook salmon of the Umpqua River (see Table 2 of Ratner et al. 1997), we computed a value of λ = 1.17, which indicates a growing population (λ > 1).
Table 2.
Predicted dates of emergence for a range of spawning dates at each of four locations impacted by dams.
| Location and date of spawning | Emergence date before dam | Emergence date after dam | Difference | F | P-value |
|---|---|---|---|---|---|
| Cowlitz River | |||||
| August 1 | 61.1 (7.0) | 73.2 (14.6) | 12.1 | 4.3 | 0.05 |
| August 15 | 82.3 (8.6) | 89 (13.8) | 6.7 | 1.4 | 0.24 |
| September 1 | 120.3 (11.6) | 110.6 (12.2) | −9.7 | 3.3 | 0.08 |
| September 15 | 166.3 (20.3) | 132.8 (9.9) | −33.5 | 32.4 | <0.0001 |
| October 1 | 212.9 (20.7) | 165.2 (11.3) | −47.7 | 57.1 | <0.0001 |
| October 15 | 238.4 (17.9) | 196.3 (15.3) | −42.1 | 34.9 | <0.0001 |
| November 1 | 260.3 (15.7) | 228.9 (14.8) | −31.4 | 22.2 | <0.0001 |
| November 15 | 272.4 (14.6) | 250.4 (12.7) | −22.0 | 14.2 | 0.0009 |
| Fall Creek | |||||
| August 1 | 49.4 (4.1) | 69.6 (8.1) | 20.2 | 68 | <0.0001 |
| August 15 | 69.8 (5.1) | 81.2 (8.8) | 11.4 | 17.8 | 0.0001 |
| September 1 | 104.4 (7.3) | 98.3 (9.6) | −6.1 | 3.6 | 0.06 |
| September 15 | 137.8 (10.6) | 116.7 (8.5) | −21.2 | 47.2 | <0.0001 |
| October 1 | 179.5 (14.4) | 147.2 (12) | −32.3 | 57.6 | <0.0001 |
| October 15 | 207.2 (13.2) | 182 (13.7) | −25.2 | 30.4 | <0.0001 |
| November 1 | 234.2 (10.9) | 218.9 (12.9) | −15.3 | 13.2 | 0.0007 |
| November 15 | 248.9 (9.9) | 239.5 (12.2) | −9.4 | 5.8 | 0.02 |
| Willamette River | |||||
| August 1 | 57 (4.8) | 64.2 (4.2) | 7.2 | 13.2 | 0.001 |
| August 15 | 78.2 (5.4) | 80.2 (3.6) | 2.0 | 1.0 | 0.32 |
| September 1 | 111 (8.0) | 102.4 (5.3) | −8.6 | 13.1 | 0.001 |
| September 15 | 151.7 (11.0) | 127.4 (10.8) | −24.3 | 34.1 | <0.0001 |
| October 1 | 189.7 (17.3) | 167.3 (16.4) | −22.4 | 10.6 | 0.003 |
| October 15 | 219.6 (17.2) | 203 (15.8) | −16.6 | 5.2 | 0.03 |
| November 1 | 245.4 (16.5) | 236.4 (15.0) | −9.0 | 1.9 | 0.20 |
| November 15 | 261 (14.6) | 255.5 (13.2) | −5.5 | 0.76 | 0.38 |
| Rogue River | |||||
| August 1 | 84.5 (16.7) | 82.7 (8.5) | −1.8 | 0.2 | 0.64 |
| August 15 | 122.3 (25.2) | 105.9 (7.3) | −16.4 | 9.8 | 0.004 |
| September 1 | 176.7 (25.7) | 138.6 (11.4) | −38.1 | 34.2 | <0.0001 |
| September 15 | 215.8 (19.8) | 168.6 (13.7) | −47.2 | 50.8 | <0.0001 |
| October 1 | 244.6 (16.1) | 199.2 (13.3) | −45.4 | 54.4 | <0.0001 |
| October 15 | 261.2 (14.8) | 220.5 (12.0) | −40.7 | 53.2 | <0.0001 |
| November 1 | 275.3 (11.8) | 241.7 (10.0) | −33.6 | 52.9 | <0.0001 |
| November 15 | 285.7 (12.3) | 257 (8.8) | −28.7 | 46.1 | <0.0001 |
Emergence dates were predicted from water temperature recorded before and after the construction of dams. These dates are reported on a relative scale (days after August 1), along with the standard deviation of inter-annual variation in parentheses. Analysis of variance was used to infer significant differences between predicted emergence dates before and after the construction of dams.
Assessing the effects of environmental change on population dynamics requires two steps. In the first step, we must relate changes in the environment to changes in life-history traits (sx, mx, bx or μ). In the second step, we must relate changes in life-history traits to changes in mean fitness (λ). The latter relationship is referred to as the demographic sensitivity (Sp):
where p equals a life-history trait. For example, the demographic sensitivity for s1, the survivorship from oviposition to age 1, depends on dA/ds1:
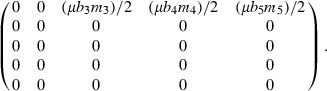 |
Using estimates of these parameters for Chinook salmon in the Umpqua River, the demographic sensitivity of s1 (123.23) greatly exceeds the sensitivities of all other survivorships in the matrix (0.12–0.35).
In this framework, the impact of a change in water temperature on fitness equals the product of the demographic and thermal sensitivities:
Because survivorship changes nonlinearly with temperature, the effect on λ depends on the mean temperature, as well as the change in temperature. Although dams have elevated the temperatures experienced by embryos and fry, temperatures during fall and winter usually lie within the tolerable limits (compare Figs 2 and 3). Therefore, the warming caused by dams likely imposes little direct thermal stress on embryos or fry. Nevertheless, the winter warming caused by dams could have indirect effects on s1, such as those believed to stem from early emergence (see previous section).
How much could early emergence alter the fitness of Chinook salmon? A matrix model can help to answer this question. Assume λ approximates the fitness of a genotype with the life cycle and vital rates described by the matrix A. Then, we need only two pieces of data to connect early emergence to a decrement in fitness. First, we must know the degree to which a dam advances the date of emergence. As a first approximation, we can calculate this advance from records of water temperature and studies of developmental rate. Second, we must know the relationship between the timing of emergence and the survivorship of juveniles (s1). Unfortunately, this relationship has not been resolved for any population of Chinook salmon, let alone the populations that spawn near our four dams. Therefore, we conducted a sensitivity analysis by exploring a set of plausible relationships between the date of emergence and s1.
As a first step in our analysis, we calculated emergence dates in the Rogue River for a set of potential spawning dates. We focused on the Rogue River because it lies close to the Umpqua River and the Chinook salmon in these two rivers undergo similar life cycles (Myers et al. 1998; ODFW 2003); thus, we felt comfortable using the parameter values reported by Ratner et al. (1997) for our modeling exercise. For each year for which we had thermal data, we used a degree-day model to estimate the timing of emergence associated with each spawning date (ranging from August 1 to January 1). We summed thermal units over successive days until we reached an empirically estimated threshold for emergence (889 units; Wedemeyer 2001). Using this procedure, we calculated emergence dates during the periods before and after the construction of a dam. Although water temperatures recorded by the gauges (Fig. 2) probably differ from those experienced by embryos in the gravel, we assumed a similar relationship between recorded temperatures and actual temperatures existed before and after the construction of the dam.
As a second step, we used predicted dates of emergence to estimate the potential impact on fitness. We used hypothetical functions describing the relationship between the date of emergence and s1; these functions differed in the optimal date of emergence and the strength of selection on the timing of emergence (Fig. 4). Optimal emergence dates were chosen by assuming development under predam conditions for three spawning dates: September 1, October 1, and November 1. These hypothetical spawning dates span the temporal window of spawning within the Rogue River (Myers et al. 1998; ODFW 2003). Three strengths of selection were chosen based on a recent study of the empirical relationship between emergence date and juvenile survivorship in Atlantic salmon (Einum and Fleming 2000). For all functions, the maximal value of s1 was set to 0.00267 (Ratner et al. 1997). For each function, we computed λ for a genotype spawning before and after the construction of the dam by inserting s1 in the matrix model of Ratner et al. (1997).
Figure 4.
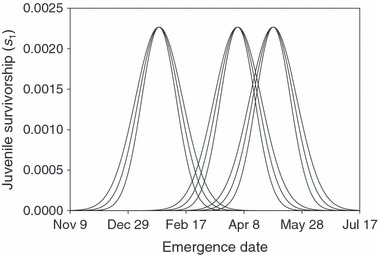
Nine hypothetical relationships between the timing of emergence and the survivorship of juveniles (s1) used in our sensitivity analysis. These relationships vary in the optimal date of emergence (early, intermediate, and late) and the strength of selection for the optimal date (strong, intermediate, and weak).
Two insights emerged from this sensitivity analysis. First, the water temperatures imposed by dams appear to influence emergence dates in a complex manner (Table 2). If salmon spawn in August, their fry would emerge at approximately the same time or slightly later than they would have emerged before the dam existed. However, if salmon spawn in September or later, their fry would emerge much earlier than they would have before the dam. Because the thermal effects of dams change seasonally, the results of our sensitivity analysis depend strongly on the optimal date of emergence. In general, the presence of a dam shifts the optimal spawning date to a later time, such that offspring would emerge on the historical date despite their accelerated development. However, the optimal spawning date shifts more for salmon that historically spawned in October or November than it does for salmon that historically spawned in September (Fig. 5).
Figure 5.
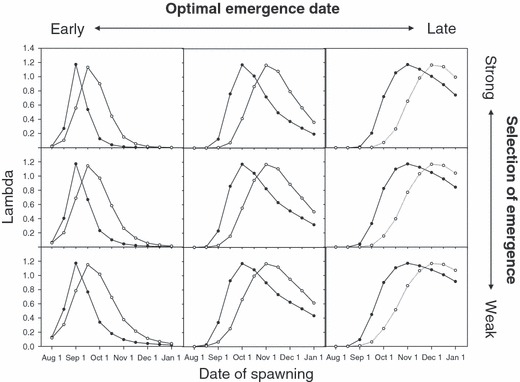
Predicted fitnesses (λ) of Chinook salmon before (filled symbols) and after (open symbols) the construction of the Lost Creek Dam on the Rogue River. Each plot corresponds to one of the relationships between emergence date and juvenile survivorship depicted in Fig. 4. From left to right, plots show earlier to later optimal dates of emergence. From top to bottom, plots show decreasing strengths of selection for emergence on the optimal date.
Within the range of conditions we considered, the dam’s influence on developmental rate should have caused the natural selection of spawning date. To illustrate this point, consider a hypothetical population in the Rogue River whose optimal spawning date was September 1 before the construction of the Lost Creek Dam. Suppose that immediately after the dam began to influence water temperature, the mean spawning date was September 1 with a standard deviation of 12.4 days (based on Beechie et al. 2006). Figure 6 illustrates the predicted relationships between the spawning date and the relative fitness for the conditions of our sensitivity analysis. The slope of each relationship represents a linear selection gradient (β), which quantifies the strength of directional selection (range = 0.024–0.036 day−1).
Figure 6.
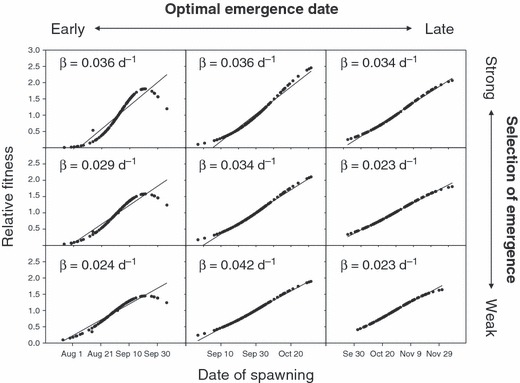
Predicted relationships between spawning date and relative fitness (λ, standardized to a mean of 1) following the construction of the Lost Creek Dam on the Rogue River. The slope of each relationship estimates the strength of directional selection (β). Each plot corresponds to one of the relationships between emergence date and juvenile survivorship depicted in Fig. 4. From left to right, plots show earlier to later optimal dates of emergence. From top to bottom, plots show decreasing strengths of selection for emergence on the optimal date.
We can use these selection gradients in two ways. First, we can standardize the selection gradients by the standard deviation of spawning date; a standardized selection gradient (β) provides a useful metric to compare the strength of selection among traits and populations (Lande and Arnold 1983; Arnold and Wade 1984). In our analysis, standardized selection gradients ranged from 0.29 to 0.52, indicating strong selection compared to values reported for other natural populations (Kingsolver et al. 2001). Second, with knowledge of the phenotypic variance and the narrow-sense heritability of spawning time, we can predict the evolutionary response to selection (i.e., the change in the mean phenotypic value per generation). Using a conservative estimate of the heritability of spawning date (0.2; Hard 2004), we predict an evolutionary change in the mean spawning time of 0.7–1.1 days per generation, depending on the relationship between emergence date and juvenile survivorship (see Fig. 4). This calculation suggests the mean spawning date would approach the new optimum within 20 generations (∼85 years). Although the dam might have caused a substantial loss in fitness (Fig. 5), evolution of the spawning date might have restored some of this loss during the last three decades and may continue to do so in the future.
Conclusions and future directions
Although dams have been altering the environments of salmon for decades, we still have much to learn about their ecological and evolutionary impacts. We explored some potential impacts by quantifying the thermal effects of dams and their consequences for the performance of Chinook salmon. This exercise indicated that dams impose substantial loss of fitness and strong selection of phenotypes. Given the high heritabilities of traits such as spawning date (Dickerson et al. 2005; Neira et al. 2006), the evolutionary responses to this selection should be rapid. Understanding the process of adaptation could help us to manage populations affected by dams. Furthermore, this framework for analyzing physiological, ecological, and evolutionary responses applies to other kinds of environmental change experienced by salmon.
Although our analysis offers insights on the potential for adaptation, we do not know whether Chinook salmon have actually adapted to the thermal changes caused by dams. Our calculations involved many assumptions to simplify the population dynamics of Chinook salmon. For example, we assume juvenile survivorship depends only on emergence date (i.e., no density-dependence), spawning behavior does not influence other traits, and genetic variance remains constant during adaptation. More complex models could relax some of these assumptions to yield deeper insights. For example, we might also consider processes that would ameliorate a negative effect of early emergence on juvenile survivorship. A phenological advance of the entire aquatic community could prevent a mismatch between emerging fry and their prey. Furthermore, poor survivorship of embryos could lead to better survivorship of fry, through known mechanisms of density dependence (Einum et al. 2008). Furthermore, we could consider the interaction of spawning date and developmental rate during adaptation. Our focus on spawning behavior simply reflects the fact that heritabilities of phenological traits tend to exceed those of physiological traits (reviewed by Carlson and Seamons 2008). Nonetheless, a change in the thermal sensitivity of developmental rate could also help salmon populations recover the loss of fitness caused by dams. Indeed, physiological adaptation might proceed more rapidly than behavioral adaptation if genetic variance in spawning date erodes over time or if adults incur substantial costs of delayed spawning. For all of these reasons, one should interpret our analysis as a means of generating testable hypotheses about the impacts of dams and as a guide for identifying crucial data to confirm these impacts.
Based on our analysis, we should improve predictions about the impacts of dams by resolving certain empirical relationships. Specifically, three factors contributed to the range of selective pressures reported in Fig. 6: (i) the thermal effects of dams; (ii) the historical date of spawning; and (iii) the relationship between emergence and survivorship. To precisely define selective pressures on salmon, all three factors must be quantified empirically for the population of interest. Moreover, a general understanding of these selective pressures requires that these analyses be carried out for multiple dams. Furthermore, additional data on the genetic variances of spawning date and developmental rate would provide better estimates of the response to selection. Finally, we can use experimental approaches to establish whether populations have already responded to the selective pressure predicted by our analysis. Common garden experiments, using eggs from populations affected by dams and populations unaffected by dams, could demonstrate whether developmental rate has adapted to the changes in thermal regimes. Studies of spawning date in a common garden would also be informative yet more challenging, because of the need to rear large numbers of salmon to adulthood. Still, only field studies can reveal how the thermal impacts of dams interact with predation risk, food availability, and hydrological conditions.
Finally, large dams do more than alter water temperatures. Reservoirs upstream of dams impact the timing of migration by juveniles, and reduced flows downstream of dams prolong the migration of both juveniles and adults (Waples et al. 2008). In some cases, dams prevent adults from reaching their historical spawning grounds, reducing the extent and variability of habitats that might favor one life-history type over another (Sheer and Steel 2006). Changes in physical habitats both upstream and downstream of dams also create new opportunities for predators and competitors. (Petersen and Kitchell 2001; Levin et al. 2002). Dams are often mitigated by hatcheries, which present another source of competitors and an additional source of selection (Ford et al. 2006). Construction of large dams has also facilitated massive withdrawals of water for agricultural and municipal purposes. Runoff from these areas increases the concentrations of nutrients, contaminants, and salts in downstream reaches (NRC 1996). These changes in water quality adversely affect the development, growth, health, and survival of salmon (Regetz 2003; Loge et al. 2005). Given this combination of stressors, our analysis likely represents an overly optimistic outlook on the ability of salmon to adapt to the environmental changes imposed by dams.
Acknowledgments
We thank Jim Myers (NOAA Fisheries) for his help with data sleuthing, and Greg Taylor, Chuck Willis, and Mindy Simmons (Army Corps of Engineers) for their insights. We also thank Sally Otto for assistance in developing the ecological and evolutionary models in Mathematica. Tom Quinn, Robin Waples, Mike Ford, Jeff Hard and an anonymous reviewer provided valuable suggestions for improving the manuscript. The views expressed herein are those of the authors and do not reflect necessarily the views of NOAA or its agencies.
Supporting information
Literature cited
- Achord S, Zabel RW, Sandford BP. Migration timing, growth, and estimated parr-to-smolt survival rates of wild Snake River spring-summer Chinook salmon from the Salmon River basin, Idaho, to the Lower Snake River. Transactions of the American Fisheries Society. 2007;136:142–154. [Google Scholar]
- Angilletta MJ. Estimating and comparing thermal performance curves. Journal of Thermal Biology. 2006;31:541–545. [Google Scholar]
- Angilletta MJ, Niewiarowski PH, Navas CA. The evolution of thermal physiology in ectotherms. Journal of Thermal Biology. 2002;27:249–268. [Google Scholar]
- Arnold SJ, Wade MJ. On the measurement of natural and sexual selection: theory. Evolution. 1984;38:709–719. doi: 10.1111/j.1558-5646.1984.tb00344.x. [DOI] [PubMed] [Google Scholar]
- Banks JL, Fowler LG, Elliott JW. Effects of rearing temperature on growth, body form, and hematology of fall chinook fingerlings. Progressive Fish Culturist. 1971;33:20–26. [Google Scholar]
- Bartholomew JL. Host resistance to infection by the myxosporean parasite Ceratomyxa shasta: a review. Journal of Aquatic Animal Health. 1998;10:112–120. [Google Scholar]
- Beacham TD, Murray CB. Temperature, egg size, and development of embryos and alevins of five species of Pacific salmon: a comparative analysis. Transactions of the American Fisheries Society. 1990;119:927–945. [Google Scholar]
- Beacham TD, Murray CB. Variation in developmental biology of sockeye salmon (Oncorhynchus nerka) and Chinook salmon (O. tshawytscha) in British Columbia. Canadian Journal of Zoology. 1989;67:2081–2089. [Google Scholar]
- Beechie T, Buhle E, Ruckelshaus M, Fullerton A, Holsinger L. Hydrologic regime and the conservation of salmon life history diversity. Biological Conservation. 2006;130:560–572. [Google Scholar]
- Bisson PA, Davis GE. Production of juvenile chinook salmon, Oncorhynchus tshawytscha, in a heated model stream. Fishery Bulletin. 1976;74:763–774. [Google Scholar]
- Brannon EL. Mechanisms stabilizing salmonid fry emergence timing. In: Smith HD, Margolis L, Wood CC, editors. Canadian Special Publication of Fisheries and Aquatic Sciences. Vol. 96. 1987. pp. 102–124. Sockeye Salmon (Oncorhynchus nerka) Population Biology and Future Management. [Google Scholar]
- Brett JR. Energetic responses of salmon to temperature: a study of some thermal relations in the physiology and freshwater ecology of sockeye salmon (Oncorhynchus nerka. American Zoologist. 1971;11:99–113. [Google Scholar]
- Brett JR. Energetics. In: Groot C, Margolis L, Clarke WC, editors. Physiological Ecology of Pacific Salmon. Vancouver: UBC Press; 1995. pp. 3–68. [Google Scholar]
- Campbell WB, Emlen JM, Hershberger WK. Thermally induced chronic developmental stress in coho salmon: integrating measures of mortality, early growth, and developmental instability. Oikos. 1998;81:398–410. [Google Scholar]
- Carlson SM, Seamons TR. A review of quantitative genetic components of fitness in salmonids: implications for adaptation to future change. Evolutionary Applications. 2008;1:222–238. doi: 10.1111/j.1752-4571.2008.00025.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caswell H. Matrix Population Models: Construction, Analysis, and Interpretation. Sunderland: Sinauer Associates; 2001. [Google Scholar]
- City of Tacoma. 2005. Cowlitz river project history. http://www.ci.tacoma.wa.us/Power/parksandpower/hydro_power/cowlitz/project_history.htm (accessed on 1 November 2007)
- Crisp DT. Thermal ‘resetting’ of streams by reservoir releases with special reference to effects on salmonid fishes. In: Craig JF, Kemper JB, editors. Regulated Streams: Advances in Ecology. New York: Plenum Press; 1987. pp. 163–182. [Google Scholar]
- Crozier L, Zabel RW. Climate impacts at multiple scales: evidence for differential population responses in juvenile Chinook salmon. Journal of Animal Ecology. 2006;75:1100–1109. doi: 10.1111/j.1365-2656.2006.01130.x. [DOI] [PubMed] [Google Scholar]
- Dickerson BR, Willson MF, Bentzen P, Quinn TP. Heritability of life history and morphological traits in a wild pink salmon population assessed by DNA parentage analysis. Transactions of the American Fisheries Society. 2005;134:1323–1328. [Google Scholar]
- Einum S, Fleming IA. Selection against late emergence and small offspring in Atlantic salmon (Salmo salar. Evolution. 2000;54:628–639. doi: 10.1111/j.0014-3820.2000.tb00064.x. [DOI] [PubMed] [Google Scholar]
- Einum S, Robertsen G, Fleming IA. Adaptive landscapes and density-dependent selection in declining salmonid populations: going beyond numerical responses to human disturbance. Evolutionary Applications. 2008;1:239–251. doi: 10.1111/j.1752-4571.2008.00021.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ford MJ, Fuss H, Boelts B, LaHood E, Hard J, Miller J. Changes in run timing and natural smolt production in a naturally spawning coho salmon (Oncorhynchus kisutch) population after 60 years of intensive hatchery supplementation. Canadian Journal of Fisheries and Aquatic Sciences. 2006;63:2343–2355. [Google Scholar]
- Geist DR, Abernethy CS, Hand KD, Cullinan VI, Chandler JA, Groves PA. Survival, development, and growth of fall Chinook salmon embryos, alevin, and fry exposed to variable thermal and dissolved oxygen regimes. Transactions of the American Fisheries Society. 2006;135:1462–1477. [Google Scholar]
- Gilchrist GW. Specialists and generalists in changing environments. I. Fitness landscapes of thermal sensitivity. American Naturalist. 1995;146:252–270. [Google Scholar]
- Gilhousen P. Prespawning mortalities of sockeye salmon in the Fraser River system and possible causal factors. International Pacific Salmon Fisheries Commission Bulletin. 1990;26:1–58. [Google Scholar]
- Goniea TM, Keefer ML, Bjornn TC, Peery CA, Bennett DH, Stuehrenberg LC. Behavioral thermoregulation and slowed migration by adult fall Chinook salmon in response to high Columbia River water temperatures. Transactions of the American Fisheries Society. 2006;135:408–419. [Google Scholar]
- Greene CM, Beechie TJ. Consequences of potential density-dependent mechanisms on recovery of ocean-type Chinook salmon (Oncorhynchus tshawytscha. Canadian Journal of Fisheries and Aquatic Sciences. 2004;61:590–602. [Google Scholar]
- Hard JJ. Evolution of chinook salmon life history under size-selective harvest. In: Hendry A, Stearns S, editors. Evolution Illuminated: Salmon and Their Relatives. Oxford: Oxford University Press; 2004. pp. 315–337. [Google Scholar]
- Hebert KP, Goddard PL, Smoker WW, Gharrett AJ. Quantitative genetic variation and genotype by environment interaction of embryo development rate in pink salmon (Oncorhynchus gorbuscha. Canadian Journal of Fisheries and Aquatic Sciences. 1998;55:2048–2057. [Google Scholar]
- Herrett TA, Hess GW, Stewart MA, Ruppert GP, Courts M. 2006. Water Resources Data – Oregon Water Year 2005. Prepared by the U.S. Geological Survey, in cooperation with federal, state, local, and tribal agencies, Portland, OR. Water Data Report OR-05-1. http://pubs.usgs.gov/wdr/#OR (accessed on 1 November 2007)
- Hyatt KD, Stockwell MM, Rankin DP. Impact and adaptation responses of Okanagan River sockeye salmon (Oncorhynchus nerka) to climate variation and change effects during freshwater migration: stock restoration and fisheries management implications. Canadian Water Resources Journal. 2003;28:689–713. [Google Scholar]
- Jensen AJ, Johnsen BO. The functional relationship between peak spring floods and survival and growth of juvenile Atlantic salmon (Salmo salar) and brown trout (Salmo trutta. Functional Ecology. 1999;13:778–785. [Google Scholar]
- Kareiva P, Marvier M, McClure M. Recovery and management options for spring/summer Chinook salmon in the Columbia River Basin. Science. 2000;290:977–979. doi: 10.1126/science.290.5493.977. [DOI] [PubMed] [Google Scholar]
- Kimbrough RA, Ruppert GP, Wiggins WD, Smith RR, Kresch DL. 2006. Water Resources Data – Washington Water Year 2005. U.S. Geological Survey, Tacoma, WA. Water Data Report WA-05-1. http://pubs.usgs.gov/wdr/#WA (accessed on 1 November 2007)
- Kingsolver JG, Gomulkiewicz R. Environmental variation and selection on performance curves. Integrative and Comparative Biology. 2003;43:470–477. doi: 10.1093/icb/43.3.470. [DOI] [PubMed] [Google Scholar]
- Kingsolver JG, Hoekstra HE, Hoekstra JM, Berrigan D, Vignieri SN, Hill CE, Hoang A, et al. The strength of phenotypic selection in natural populations. American Naturalist. 2001;157:245–261. doi: 10.1086/319193. [DOI] [PubMed] [Google Scholar]
- Kinnison MI, Unwin MJ, Hershberger WK, Quinn TP. Egg size, fecundity, and development rate of two introduced New Zealand chinook salmon (Oncorhynchus tshawytscha) populations. Canadian Journal of Fisheries and Aquatic Sciences. 1998;55:1946–1953. [Google Scholar]
- Lande R, Arnold SJ. The measurement of selection on correlated characters. Evolution. 1983;37:1210–1226. doi: 10.1111/j.1558-5646.1983.tb00236.x. [DOI] [PubMed] [Google Scholar]
- Levin PS, Achord S, Feist BE, Zabel RW. Non-indigenous brook trout and the demise of Pacific salmon: a forgotten threat. Proceedings of the Royal Society of London. Series B. 2002;269:1663–1670. doi: 10.1098/rspb.2002.2063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichatowich J. Salmon without Rivers: A History of the Pacific Salmon Crisis. Washington, DC: Island Press; 1999. [Google Scholar]
- Ligon FK, Dietrich WE, Trush WJ. Downstream ecological effects of dams. BioScience. 1995;45:183–192. [Google Scholar]
- Loge FJ, Arkoosh MR, Ginn TR, Johnson LL, Collier TK. Impact of environmental stressors on the dynamics of disease transmission. Environmental Science & Technology. 2005;39:7329–7336. doi: 10.1021/es0481934. [DOI] [PubMed] [Google Scholar]
- Lynch M, Gabriel W. Environmental tolerance. American Naturalist. 1987;129:283–303. [Google Scholar]
- Macdonald JS, Foreman GG, Farrell T, Williams IV, Grout J, Cass A, Woodey JC, et al. The influence of extreme water temperatures on migrating Fraser River sockeye salmon (Oncorhynchus nerka) during the 1998 spawning season. Canadian Technical Report of Fisheries and Aquatic Sciences. 2000;2326:1–117. [Google Scholar]
- Materna E. Seattle, WA: U.S. Fish and Wildlife Service; 2001. Temperature Interaction. Issue Paper 4, Temperature Water Quality Criteria Guidance Development Project EPA-910-D-01-004, Environmental Protection Agency Region 10. [Google Scholar]
- McCullough DA. A Review and Synthesis of Effects of Alterations to the Water Temperature Regime on Freshwater Life Stages of Salmonids, with Special Reference to Chinook Salmon. Region 10, Seattle, WA, USA: Environmental Protection Agency; 1999. [Google Scholar]
- Murray CB, Beacham TD. The development of Chinook (Oncorhynchus tshawytscha) and chum salmon (Onchorhynchus keta) embryos and alevins under varying temperature regimes. Canadian Journal of Zoology. 1987;65:2672–2681. [Google Scholar]
- Murray CB, McPhail JD. Effect of incubation temperature on the development of five species of Pacific salmon (Oncorhynchus) embryos and alevins. Canadian Journal of Zoology. 1988;66:266–273. [Google Scholar]
- Myers JM, Kope RG, Bryant GJ, Teel D, Leirheimer LJ, Wainwright TC, Grant WS, et al. Status Review of Chinook Salmon from Washington, Idaho, Oregon, and California. Washington, DC: U.S. Department of Commerce; 1998. NOAA Technical Memorandum NMFS-NWFSC-35. [Google Scholar]
- National Research Council (NRC) Upstream: Salmon and Society in the Pacific Northwest. Washington, DC: National Academy Press; 1996. [Google Scholar]
- Neira R, Diaz NF, Gall GAE, Gallardo JA, Lhorente JP, Alert A. Genetic improvement in coho salmon (Oncorhynchus kisutch). II: Selection response for early spawning date. Aquaculture. 2006;257:1–8. [Google Scholar]
- Olson PA, Nakatani RE, Meekin T. Effects of Thermal Increments on Eggs and Young of Columbia River Fall Chinook. Richland, WA: Pacific Northwest Laboratories; 1970. [Google Scholar]
- Oregon Department of Fish and Wildlife (ODFW) 2003. Natural resources information management program: Timing tables. http://nrimp.dfw.state.or.us/nrimp/default.aspx?pn=timingtables (accessed on 1 November 2007)
- Pacific Fishery Management Council (PFMC) 1979. Freshwater habitat, salmon produced, and escapements for natural spawning along the Pacific coast of the United States. Report prepared by the Anadromous Salmonid Environmental Task Force of the PFMC, Portland, OR.
- Pearcy WG. Ocean Ecology of North Pacific Salmonids. Seattle, WA: University of Washington Press; 1992. [Google Scholar]
- Petersen JH, Kitchell JF. Climate regimes and water temperature changes in the Columbia River: bioenergetic implications for predators of juvenile salmon. Canadian Journal of Fisheries and Aquatic Sciences. 2001;58:1831–1841. [Google Scholar]
- Petts GE. Impounded Rivers: Perspectives for Ecological Management. New York: John Wiley and Sons; 1984. [Google Scholar]
- Poff NL, Hart DD. How dams vary and why it matters for the emerging science of dam removal. BioScience. 2002;52:659–668. [Google Scholar]
- Preece RM, Jones HA. The effect of Keepit Dam on the temperature regime of the Namoi River, Australia. River Research and Applications. 2002;18:397–414. [Google Scholar]
- Quinn TP, Adams DJ. Environmental changes affecting the migratory timing of American shad and sockeye salmon. Ecology. 1996;77:1151–1162. [Google Scholar]
- Quinn TP, Unwin MJ, Kinnison MT. Evolution of temporal isolation in the wild: genetic divergence in timing of migration and breeding by introduced chinook salmon populations. Evolution. 2000;54:1372–1385. doi: 10.1111/j.0014-3820.2000.tb00569.x. [DOI] [PubMed] [Google Scholar]
- Rand PS, Hinch SG. Swim speeds and energy use of up-river migrating sockeye salmon: simulating metabolic power and assessing risk of energy depletion. Canadian Journal of Fisheries and Aquatic Sciences. 1998;55:1832–1841. [Google Scholar]
- Ratner S, Lande R, Roper BB. Population viability analysis of spring chinook salmon in the South Umpqua River, Oregon. Conservation Biology. 1997;11:879–889. [Google Scholar]
- Regetz J. Landscape-level constraints on recruitment of Chinook salmon (Oncorhychus tshawytscha) in the Columbia River basin, USA. Aquatic Conservation: Marine and Freshwater Ecosystems. 2003;13:35–49. [Google Scholar]
- Richter A, Kolmes SA. Maximum temperature limits for chinook, coho, and chum salmon, and steelhead trout in the Pacific Northwest. Reviews in Fisheries Science. 2005;13:23–49. [Google Scholar]
- Roper BB, Scarnecchia DL. Emigration of age-0 chinook salmon (Onchorhynchus tshawytscha) smolts from the upper South Umpqua River basin, Oregon, U.S.A. Canadian Journal of Fisheries and Aquatic Sciences. 1999;56:939–946. [Google Scholar]
- Ruckelshaus MR, Levin P, Johnson JB, Kareiva PM. The Pacific salmon wars: what science brings to the challenge of recovering species. Annual Review of Ecology and Systematics. 2002;33:665–706. [Google Scholar]
- Scheuerell MD, Hilborn R, Ruckelshaus MH, Bartz KK, Lagueux K, Haas A, Rawson K. The Shiraz model: a tool for incorporating anthropogenic effects and fish–habitat relationships in conservation planning. Canadian Journal of Fisheries and Aquatic Sciences. 2006;63:1596–1607. [Google Scholar]
- Schlosser IJ, Johnson JD, Knotek WL, Lapinska M. Climate variability and size-structured interactions among juvenile fish along a lake-stream gradient. Ecology. 2000;81:1046–1057. [Google Scholar]
- Sheer MB, Steel EA. Lost watersheds: barriers, aquatic habitat connectivity, and species persistence in the Willamette and Lower Columbia basins. Transactions of the American Fisheries Society. 2006;135:1654–1669. [Google Scholar]
- Smith SG, Muir WD, Hockersmith EE, Zabel RW, Graves RJ, Ross CV, Connor WP, et al. Influence of river conditions on survival and travel time of Snake River subyearling fall chinook salmon. North American Journal of Fisheries Management. 2003;23:939–961. [Google Scholar]
- Smoker WW, Gharrett AJ, Stekoll MS. Genetic variation of return date in a population of pink salmon: a consequence of fluctuating environment or dispersive selection? Alaska Fishery Research Bulletin. 1998;5:46–54. [Google Scholar]
- Steel EA, Lange IA. Alteration of water temperature regimes at multiple scales: effects of multi-purpose dams in the Willamette River basin. River Research and Applications. 2007;23:351–359. [Google Scholar]
- StreamNet. 2005. Anadromous fish distribution. http://www.streamnet.org/online-data/GISData.html#anad (accessed on 1 November 2007)
- Tang J, Bryant MD, Brannon EL. Effect of temperature extremes on the mortality and development rates of coho salmon embryos and alevins. The Progressive Fish-Culturist. 1987;49:167–174. [Google Scholar]
- United States Army Corps of Engineers (USACE) Biological Assessment of the Effects of the Willamette River Basin Flood Control Project on Listed Species under the Endangered Species Act. Portland, OR: National Marine Fisheries Service and the US Fish and Wildlife Service; 2000a. [Google Scholar]
- United States Army Corps of Engineers (USACE) National Inventory of Dams. Washington, DC: United States Army Corps of Engineers in cooperation with the Federal Emergency Management Agency’s National Dam Safety Program; 2000b. [Google Scholar]
- United States Geological Survey (USGS) 2007. National water information system. http://waterdata.usgs.gov/usa/nwis/nwis (accessed on 1 November 2007)
- Velsen FPJ. Temperature and incubation in Pacific salmon and rainbow trout: compilation of data on median hatching time, mortality and embryonic staging. Canadian Data Report of Fisheries and Aquatic Sciences. 1987;626:1–58. [Google Scholar]
- Vinson MR. Long-term dynamics of an invertebrate assemblage downstream form a large dam. Ecological Applications. 2001;11:711–730. [Google Scholar]
- Waples RS, Zabel RW, Scheuerell MD, Sanderson BL. Evolutionary responses by native species to major anthropogenic changes to their ecosystems: Pacific salmon in the Columbia River hydropower system. Molecular Ecology. 2008;17:84–96. doi: 10.1111/j.1365-294x.2007.03510.x. [DOI] [PubMed] [Google Scholar]
- Ward JV. Ecological aspects of stream regulation: responses in downstream lotic reaches. Water Pollution and Management Reviews (New Dehli) 1982;2:1–26. [Google Scholar]
- Ward JV, Stanford JA, editors. The Ecology of Regulated Streams. New York: Plenum Press; 1979. [Google Scholar]
- Wedemeyer GA. Fish Hatchery Management. Bethesda: American Fisheries Society; 2001. [Google Scholar]
- Willingham WF. Army Engineers and the Development of Oregon: A History of the Portland District. Washington, DC: U.S. Army Corps of Engineers Government Printing Office; 1983. [Google Scholar]
- Zabel RW, Scheuerell MD, McClure MM, Williams JG. The interplay between climate variability and density dependence in the population viability of Chinook salmon. Conservation Biology. 2006;20:190–200. doi: 10.1111/j.1523-1739.2005.00300.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.