

Abstract
Memory of past experience is essential for guiding goal-related behavior. Being able to control accessibility of memory through modulation of retrieval enables humans to flexibly adapt to their environment. Understanding the specific neural pathways of how this control is achieved has largely eluded cognitive neuroscience. Accordingly, in the current paper I review literature that examines the overt control over retrieval in order to reduce accessibility. I first introduce three hypotheses of inhibition of retrieval. These hypotheses involve: i) attending to other stimuli as a form of diversionary attention, ii) inhibiting the specific individual neural representation of the memory, and iii) inhibiting the hippocampus and retrieval process more generally to prevent reactivation of the representation. I then analyze literature taken from the White Bear Suppression, Directed Forgetting and Think/No-Think tasks to provide evidence for these hypotheses. Finally, a neuroanatomical model is developed to indicate three pathways from PFC to the hippocampal complex that support inhibition of memory retrieval. Describing these neural pathways increases our understanding of control over memory in general.
Keywords: Anatomy, Hippocampus, Inhibition, Memory, Neuroimaging, Retrieval
1. Introduction
One of the quintessential aspects of being human is reflected in the connection to one’s past, supported by our memories. Memory, whether procedural or declarative, governs and facilitates countless behavioral phenomena during life. It places us in our current context, reminds us of where we are going, and where we have come from. Yet we are not slaves to our memory, as we exhibit the ability to control various aspects of it. Control over memory, in particular, may exist as a cognitive mechanism that influences our awareness of memories in a flexible manner. Such control mechanisms may have evolved to influence older brain circuitry, such as modulating the memory processes of the non-neocortical hippocampus, which may be referenced or used in on-going behavior. Clearly, when working towards a goal, there are countless occurrences in which some memory information must be facilitated, while other irrelevant memory information must be inhibited. Being able to accomplish this feat clearly enables us to be more flexible when interacting with our environment.
Our control over memory becomes most apparent when we are confronted with memories that we wish to avoid thinking about, such as those that are emotionally painful or associated with traumatic events. In some cases reliving such affective experiences may be beneficial to a point, as in bereavement (Malkinson, 2001). However, if not eventually brought under control, such memories may repeatedly exceed conscious thresholds, becoming intrusive and ruminative in nature. In some conditions, such as post-traumatic stress disorder (PTSD), retrieval of such memories may cause serious distress and impairment.
In view of their importance for behavior, processes that are involved in the creation and use of memory, such as encoding, maintenance and retrieval have been extensively researched. However, one memory phenomena that continues to be contentious is the question of whether we can inhibit the retrieval of specific memories. While the last century has brought about a significant increase in our understanding of how memory information becomes less accessible, exemplified in numerous theoretical accounts of how forgetting occurs, it has focused mainly on processes that are automatic (e.g., interference, decay). Only more recently has cognitive psychology and neuroscience probed into whether controlled aspects of memory apply to the inhibition of retrieval. Consequently, there is little consensus on the specific mechanisms by which control may be exerted to inhibit memory retrieval.
The following integrative review attempts to understand and model mechanistically the ability to direct control over memory in order to inhibit retrieval at both the level of cognitive (e.g., long-term and working memory) and neural (e.g., hippocampus, PFC) systems. In the review I attempt to clearly illustrate how inhibition of retrieval may involve alterations of encoding or alternatively learning to avoid retrieval. This perhaps occurs by cognitive/inhibitory control, reflected as increased activation of PFC areas to down-regulate hippocampal/MTL regions. Specific anatomical PFC-hippocampal/MTL pathways and their associated processes will be discussed in regard to dopamine and neural oscillations that may affect (i) pattern completion, (ii) activity in posterior cortex (sensory processing and multimodal association areas), and (iii) information gated through the basal ganglia to be accessed or maintained by WM, all of which are likely to affect the amount of information recalled at any given moment. The subsequent discussion to illustrate such control over memory processes proceeds with the following sections: First, in view of the current focus on inhibition of retrieval, the basic nature of encoding and retrieval processes in memory are described. Second, three of the most relevant hypotheses regarding inhibition of retrieval in the literature are discussed and illustrated. Next, the behavioral and neuroimaging literatures examining the inhibition of retrieval are then analyzed to assess which of these three hypotheses is most strongly supported. Finally, a neuroanatomical model of inhibition of retrieval is presented that incorporates most findings in the literature.
2. The Basic Nature of Encoding and Retrieval Processes
Because the focus of this paper involves inhibition of retrieval, it is important to outline what we know about the retrieval process in memory. Of course, the amount of information retrieved is also dependent on previous attempts to encode information, so it is necessary to also consider the encoding process. Because these two processes of memory are examined by multiple lines of research in the fields of cognitive psychology and neuroscience, there is significant debate on the specific details of each component. The specific distinctions within each of these processes are beyond the scope of our discussion. Yet, their general aspects, defined by recent consensus in cognitive neuroscience, are important to outline, because their neural counterparts and putative functionality will later serve to illustrate how mechanisms of inhibition of retrieval may interact with each process.
First, concerning memory encoding, the medial temporal lobe (MTL) is a major terminus of the various sensory processing pathways. As the neuronal firing of these sensory features is maintained by focused attention on the representation of a stimulus, a binding process occurs within the hippocampus proper and its surrounding MTL cortices (i.e., parahippocampal and entorhinal cortices) to conjoin the different perceptual features of the event in a conjunctive manner (Jonides et al., 2008; O’Reilly & Norman, 2002). Within the hippocampus, this process occurs in a sparse neuronal firing pattern that is thought to index and represent the original neural representations of the event that occurred in various sensory processing regions. By encoding and binding these conjunctive representations into a sparse representation in the hippocampus, the likelihood of interference from other events is decreased. Furthermore, the hippocampus proper exhibits a fast learning rate, which also minimizes interference that may occur during the time between perception and encoding (O’Reilly & Norman, 2002; O’Reilly & McClelland, 1994). As the hippocampus starts binding these different neural sensory representations of the external event, plasticity (both Hebbian and non-Hebbian LTP) occurs. As plasticity takes place, it causes increased communication between pre- and post-synaptic cells associated with the sparse representation. This process allows the linkage between the different neural sensory component representations of the specific event within the hippocampus proper via enhanced protein synthesis (Wixted, 2004; Jonides et al., 2008). The plasticity process provides the neural mechanisms of consolidation, in which a particular external event becomes a lasting memory, which is the basis for long-term memory (LTM).
If encoding occurs with sufficient activation during a specific event, a memory may become established and durable. If so, then a particular internal or external sensory cue may allow for retrieval. The memory of the initial event is retrieved through activation of one of the component representations in the neural network that represents the entire composite memory representation. Specifically, if a component representation is activated and attention is focused on it, the representation may then continue to activate in a feed-forward manner to the hippocampus. If this occurs, activation of any linked component representation of the original event in sensory regions may feed-forward to the hippocampus, causing the sparse representation indexed within the hippocampus to pattern complete (Rolls, 1989; O’Reilly & McClelland, 1994). Pattern completion then reactivates all the specific component representations of the original event through feed-back activation from the hippocampus to sensory regions that originally encoded the event. As more attention is focused on the emerging component representations of the features of the original memory event, the specific representations which were first engaged in perception are reactivated to some degree. Attentional resources (e.g., prefrontal and parietal regions) then maintain the firing of these representations as a composite memory (Jonides et al., 2008).
The maintenance of these retrieved memories is thought to be governed by a cortical-basal ganglia-thalamo-cortical loop. In this loop sensory representations in posterior cortex are actively maintained by the prefrontal cortex (PFC) through a gating mechanism in the basal ganglia and thalamus that allows the PFC to communicate with posterior cortex (Frank et al., 2001; Hazy et al., 2006). This gating mechanism is controlled by dopaminergic input to the basal ganglia and signals when to open and close the gate so that PFC can flexibly maintain the firing and updating of memory representations. During maintenance, the flow of information is inhibited so that current representations that are actively firing in PFC can continue to do so. Conversely, when updating occurs this loop becomes uninhibited, so that new information can transfer to the PFC for subsequent maintenance (for a review see O’Reilly & Frank, 2006). The selective maintenance and updating of these representations is the basis for which information is accessible to working memory (WM) and ultimately what is recalled.
In general, it is worth noting that inhibition of retrieval can be influenced by differences in the nature of processing i.e., automatic vs. controlled processes. While both automatic and controlled processes can be seen as essential for proper memory function, they likely involve different neural components. Whereas automatic processes occur outside of awareness or cognitive influence, controlled processes are cognitively directed. Automatic processes are more akin to conceptions of standard processes of forgetting (e.g., proactive/retroactive interference), whereas inhibition of retrieval implies a cognitively directed mechanism to lessen the accessibility of memory information. However, the difference between these processes likely occurs in impetus or motivation, and does not necessitate that every component by which these processes function are distinctly separable (discussed later). Because the focus of this paper concerns cognitive attempts during inhibition of retrieval, only cognitive memory paradigms that overtly require individuals to block or interfere with retrieval are analyzed.1
3. Hypotheses of Inhibitory Modulation of Memory Retrieval
Although the term inhibition is widely used in psychology and neuroscience to describe a range of different phenomena (e.g., cognitive, behavioral, neural), it is often vaguely conceived and consequently leads to debate (Munakata et al. in press). Therefore, in order to provide a framework in which findings can be reviewed and interpreted, this section proposes three hypotheses that define the manner in which inhibition of retrieval may occur (illustrated in Figure 1). Although these hypotheses have not been fully specified in mechanistic detail in the literature, they do encompass the major competing views about how inhibitory modulation may reduce or lessen memory retrieval.
Figure 1.
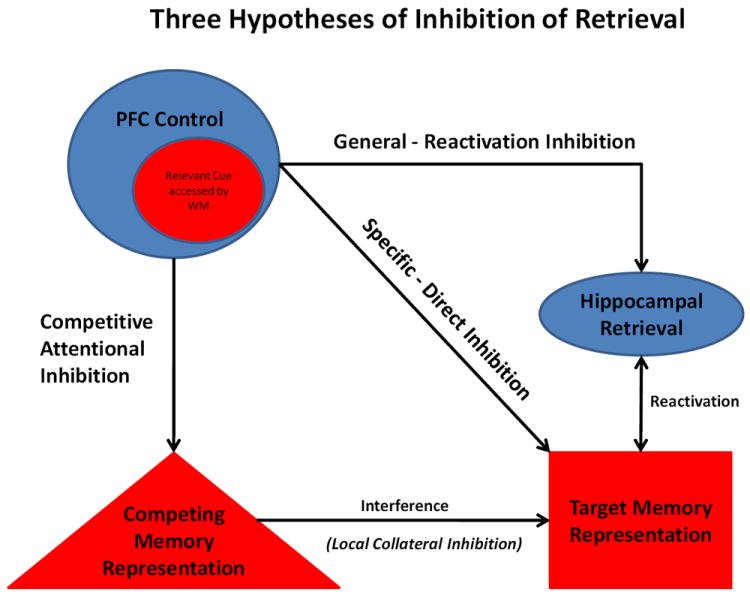
Three hypotheses of how inhibition of memory retrieval may function, including: direct inhibition (PFC specific inhibition of a single neural representation), reactivation inhibition (PFC general inhibition of the retrieval process) and competitive attentional inhibition (diversionary attentional distraction from a specific memory).
3.1 Direct Inhibition
This hypothesis, often referred to as retrieval inhibition or directed suppression, concerns inhibition over a specific, singular memory representation (Gieselman et al., 1983; Bjork, 1989; Bjork et al., 1998; Anderson & Green, 2001; Levy & Anderson, 2002). According to this hypothesis, as cues relevant to stored memories are accessed or maintained by working memory (WM), the cues initiate cognitive control processes exerted by prefrontal cortex (PFC). PFC is hypothesized to send an inhibitory signal directed at the specific representation of the target memory, stored as a long-term memory that has been associated with the cue, thereby preventing retrieval. 2
3.2 Reactivation Inhibition
This hypothesis proposes that cognitive control processes of the PFC initiate inhibition that modulates memory retrieval processes more generally rather than a specific singular memory representation. In this case, inhibitory signals affect the memory retrieval process, a process which is hypothesized to involve a hippocampal/MTL mechanism that reactivates the perceptual representations of the original memory by pattern completion (discussed above). Under this hypothesis, stopping the reactivation process from occurring would involve increased PFC control that induces the reduction of information output from the hippocampus to surrounding MTL cortices (Depue et al., 2007; 2010; 2011). Decreased output of the hippocampus/MTL would, in turn, reduce pattern completion of the memory, which subsequently stops the reactivation of the full composite memory ensemble in supporting cortices. The end result is insufficient information for processes of WM to represent and maintain the memory (Depue et al., 2007; 2010; 2011). The major distinction between this hypothesis and the Direct Inhibition hypothesis is that, in the former, inhibition is invoked as a general control mechanism over a process or brain region rather than a specific memory item or representation, as in the latter.
3.3 Competitive Attentional Inhibition
This hypothesis proposes that retrieval of a target memory is reduced by directing attention to something else, that is, by diversion from the present memory. Subsequently, the diversionary process is initiated when a target-relevant cue is accessed or maintained by WM. At that point, attention is then focused on an alternative memory representation other than the target memory, which then becomes associated with the cue. As continued attention to the diversionary memory representation strengthens its association with the cue, it may inhibit the original target representation through competitive collateral inhibition, normally associated with stored representations in LTM. I would suggest that this process is achieved, not by competing representations in LTM, but rather through WM processes of PFC. In order for local collateral competition to inhibit competing representations in LTM, the neural representations must be spatially close in proximity for collateral inhibition to transfer from one representation to another. Therefore, it would seem unlikely that the content and neural representation of diversionary memories would always meet this spatial requirement. One possibility is that competition occurs between more abstract tags associated with WM maintenance of both the diversionary and target memory located spatially close within PFC, which may induce local collateral inhibition. In this view, Competitive Attentional Inhibition involves a reduction or resolution of interference among various memory representations competing for expression (Miller & Cohen, 2001; Wimber et al., 2008). Therefore, this hypothesis can also be regarded as engaging automatic inhibitory processes that likely function in standard conceptions of forgetting (e.g., retro/proactive interference; Konishi et al., 2011).
4. Empirical Findings on Inhibitory Modulation of Memory Retrieval
Because the focus of this review concerns cognitive attempts at inhibition of retrieval, only memory paradigms that overtly require individuals to stop thinking, not think, forget, or control their thoughts about specific memories are considered. The three most relevant paradigms that have been used to examine the inhibition of retrieval are: i) White Bear Suppression task, ii) Directed Forgetting task, and iii) Think/No-Think task. Findings related to these three paradigms are discussed in two major sections, one focusing on behavioral studies, and the other focused on neuroimaging studies. Although debate over the contribution of mechanisms described in the three previous hypotheses exists, it is indisputable that these tasks examine controlled processes in efforts to reduce retrieval and hence, awareness of memory information.
4.1 Behavioral Studies
4.1.1 White Bear (WB) Experiments
A wave of interest in the control over memory began with seminal work by Wegner and colleagues in the WB experiments (Wegner et al. 1987; Wenzlaff et al.; 1988; 1991; Wegner & Erber, 1992; Wegner, 1992). During the task, participants are told that in one portion of the study they are going to be asked to “not think of a white bear”, referred to as the suppression period, and in another portion to engage in free thought, known as the expression period. The order in which these two portions occur is counterbalanced across individuals so that some have the expression period initially and others the suppression period initially. Each period usually lasts five minutes, and participants are asked to ring a bell when they encounter the target thought (i.e., white bear). Results have indicated two major effects: 1) participants in the suppression initial period have a higher rate of target thoughts than those in expression initial period; and 2) participants in the suppression initial period, have a higher rate of target thoughts during the subsequent expression period. Wegner termed this latter finding as post-suppression rebound (Wegner, 1994), a paradoxical shift that occurs when participants are asked to control memory information.
In some versions of the task, participants are instructed to think of a diversionary thought/memory during the suppression period. During some experiments using this method, no evidence of rebound effects was observed (Wegner et al., 1987, experiment 2). Thus, it was concluded that, if individuals have an alternate stimulus to which they divert their attention while attempting to control the memory of the white bear, the probability of retrieval decreased. In contrast, if no diversionary stimulus was provided, retrieval increased. Therefore, successfully inhibiting retrieval in some of these studies has been attributed to diversionary mechanisms that are specifically termed environmental cueing (Wegner, 1994). Environmental cueing suggests that, as an individual attempts to control memory information by focusing on a diversionary stimulus, these stimuli become associated with the original target memory (i.e., white bear). Therefore, repeated attempts to control the target memory by diversionary tactics associate multiple alternate memories with the target stimulus, making retrieval of the target less probable (Wegner, 1989; 1994).
Such diversionary accounts were incorporated into a more formal Dual-Process Theory consisting of: 1) the operating process, which involves consciously searching for diversionary stimuli on which to focus attention, and 2) the unconscious monitoring process, which is sensitive to the intrusions of the target memory under control. It was proposed that when the two processes become imbalanced, intrusions occur more frequently than diversionary stimuli and increased conscious detection of the unwanted target occurs. This explanation suggests that, when diversionary attempts fail, individual’s become aware of the target memory. This was later termed the Ironic Process Theory (Wegner, 1992; 1994; 1997; Wegner & Wenzlaff, 1996).
While studies examining the WB task yield varied results regarding the efficacy of inhibiting retrieval, the majority of the findings seem to show ineffectual control of memory as measured by intrusion rates, although there is some evidence to suggest that providing specific diversionary tactics may be more successful than other attempts to inhibit retrieval. Thus, some evidence from the WB task appears to support the mechanism of Competitive Attentional Inhibition hypothesis of inhibition of retrieval.
4.1.2 Directed Forgetting (DF) Experiments
One of the most investigated approaches to inhibition of retrieval has used the DF task (Muther, 1965; Bjork et al., 1968; Bjork, 1970; Bjork & Woodward, 1973). Because more than 600 studies exist using the DF task, only an overview of the findings and the mechanisms/processes that have been hypothesized to be involved in inhibition of retrieval is presented (for a comprehensive reviews see Wilson & Kipp, 1998; Macleod, 1999). In general, there are two different versions of this task, each involving different cueing methods: 1) the item- or single-item cueing method, where a forget or remember cue is presented immediately prior, during, or after presentation of a target stimulus, and then testing for recall and/or recognition is performed after presentation of all stimuli (Muther, 1965; Weiner & Reed, 1969; Geiselman & Bagheri, 1985; Lehman & Bovasso, 1993); and 2) the list- or blocked-cueing method, where a list or partial subset of the list of words is studied by the participant, after which they are instructed to forget the list or partial subset of the list and remember a subsequently presented list (Bjork, 1970; Bruce & Papay, 1970).
Many of these studies demonstrate a reduced recollection or recognition of to-be-forgotten (TBF) items, as compared to to-be-remembered (TBR) items, indicating control over memory information. Key differences between the two methodologies have elicited different mechanistic explanations of DF task findings. In general, results from the item method are more frequently attributed to mechanisms occurring during encoding as opposed to retrieval processes. The item method yields differences in recognition performance, such that TBR items show increased recognition compared to TBF items. Recognition performance is postulated to be sensitive to encoding, such that increased recognition indicates increased access to memory information (Wilson & Kipp, 1998). Therefore, the major theory concerning mechanisms engaged during the item method of the DF task is selective or elaborative rehearsal (Bjork & Geiselman, 1978; Block 1971; Elmes, Adams, & Roediger, 1970; Geiselman, Bjork, & Fishman, 1983, Wilson & Kipp, 1998). Selective or elaborative rehearsal simply means that TBR items receive more rehearsal than TBF items, perhaps because the cue presentation occurs in close temporal proximity to the target (Wilson & Kipp, 1998). This indicates that TBR items may be more strongly encoded, and subsequently show increased recognition making inferences about retrieval processes difficult. Because the mechanisms that are thought to inhibit retrieval during the item method are linked to encoding processes, they do not fall under the outlined hypotheses of inhibition of retrieval. However, they still indicate why the retrieval of memory information may be altered and will be revisited in the neuroimaging and model sections below.
In contrast to the item method, results from the list method are used to support mechanisms occurring during retrieval processes. In general, findings from the list method yield equal recognition of TBR and TBF items, indicating similar amounts of rehearsal and encoding. Furthermore, recollection is usually greater for TBR items, leading some to suggest that inhibitory control over memory occurs during retrieval processes (Elmes et al., 1970; Bjork, 1970; Block, 1971; Bjork & Woodward, 1973). The major account of these findings usually termed retrieval inhibition is suggested to act as an inhibitory mechanism affecting the actual representation of TBF lists or items in LTM. Although specific details are vague, some explanations of this hypothesis suggest that the inhibitory process targets the specific memory representation and decreases the likelihood of its retrieval (Bjork, 1989; Bjork et al., 1998). Because the reductions of retrieval in the behavioral studies of the DF task do not indicate where inhibition is focused (i.e., the actual specific representation of items in the list or the retrieval process in general), the mechanisms attributed to the list method fall under both the Direct and Reactivation Inhibition hypotheses (Figure 1).
4.1.3 Think/No-Think (TNT) Experiments
More recent investigations that examine inhibition of retrieval use the influential TNT task designed to investigate the influence of repetitive attempts at inhibiting retrieval (Anderson & Green, 2001). The TNT task involves the learning of pairs of stimuli (cue-target) to a prescribed level of accuracy, ensuring that the cue-target pairings have been encoded into LTM. Once learned, only the cue is presented, and participants are instructed in one condition to “think” or in another condition to “not think” of previously associated targets. These cues are then shown a number of times (e.g., 12-16) to allow for multiple opportunities to exert control over memory information. Some stimuli, not presented during the experimental phase, serve as a baseline memory condition to which Think (T) and No-Think (NT) trials can be compared. Accuracy of memory is then assessed by cued recall.
Multiple behavioral studies indicate that NT items are recalled less than T items and, crucially, less than baseline items that assess normal memory. Reductions from baseline in recall for NT items suggest that cognitive attempts to inhibit retrieval actually reduce the accessibility of NT items (Anderson & Green, 2001; Salame & Danion, 2007; Wessel et al., 2005; Depue et al., 2006, Paz-Alonso et al., 2009; Lambert et al., 2010; Norby et al., 2009). Although many studies show reduced retrieval for NT items, it must be noted that some have failed to replicate these results (Bulevich et al., 2006, Bergstrom et al., 2009; Mecklinger et al., 2009). Thus, it remains a question as to what role demand characteristics, as well as strategy and participant instruction play on behavioral results of the TNT task. Future studies examining these issues are of high interest.
Importantly, some studies include an independent probe (IP) condition during cued recall testing. The IP condition uses an alternative cue, not presented previously in the experiment, to assess cued recall (see Anderson & Green, 2001). For example, if the original studied NT cue-target pairing were the words ordeal+roach, and the cue “ordeal” was used during the experimental phase, the IP cued recall for the pair would be bug+r_____. If recall of “roach” is reduced below baseline levels in the IP condition, then the results cannot be explained by an alteration of the cue-target relationship in memory (e.g., associating another word other than roach with “ordeal”). Rather, the actual memory representation of the target must have been inhibited.
Recently, the results from the IP cued recall technique have been challenged by showing that using an interference condition (pressing a button to some cues) is enough to cause reduced retrieval even when tested by alternate cues (Tomlinson et al., 2009). To explain these findings, Tomlinson and colleagues associated their findings with a two-stage Global Memory Model, in which recall involves a sampling stage and a recovery stage. For instance, in the previous example of ordeal+roach, during retrieval at the sampling stage, the cue “ordeal” is used to sample only a partial representation of “roach” (i.e., r_ac_). If the cue “ordeal” has been linked to other diversionary targets during repeated exposures, then interference may occur during sampling. Using alternate cues, as in the IP condition, circumvents this sampling interference: that is, bug+r_____ cues “roach” with no interference caused by associated diversionary representations learned in connection with “ordeal”. Once again, because IP cued recall using bug+r____ do show decreased retrieval, it has been taken to suggest that the actual representation of “roach” has been inhibited.
The alternative explanation by Tomlinson and colleagues is that interference may also occur during the recovery stage. Such interference would occur when the repeated exposure of the cue “ordeal” induces partial sampling of “roach” (r_ac_). During this partial sampling a diversionary representation (thought, action) may then get associated not only to “ordeal” but also to “roach”. Consequently, during IP cued recall, full recovery of “roach” would suffer from competing diversionary representations associated with “roach,” regardless of the cue used. Therefore, their findings suggest that the individual representation may not need to be directly targeted for inhibition and that interference may be an alternate explanation to IP findings.
From these various studies, two main mechanisms for behavioral TNT results have been proposed. The first is directed suppression, which suggests that cognitive or inhibitory control is directed towards the specific neural representation of the target memory itself (Levy & Anderson, 2002). This theory is largely based on the findings of the IP condition, and posits that, if cued recall using alternative cues is below baseline recall, then inhibition over NT items occurs not by simply removing cue-target associations or by increasing interference, but rather by affecting the specific target representation itself (Anderson & Green, 2001; Levy & Anderson, 2002, Anderson, 2003). This mechanism serves as the basis for the Direct Inhibition hypothesis of inhibition of retrieval in Figure 1.
A second mechanism is a diversionary account, where control of memory information is achieved by thinking of alternative, distracting memories or thoughts (Hertel & Calcaterra 2005). When associated with cues of NT items, these alternative memories subsequently create interference or competition during cued recall. Supporting this account, Hertel & Calcaterra (2005) included an aided condition in which participants were supplied with substitution stimuli to think of when an NT trial occurred. This aided condition decreased the recall of NT items over general NT instructions.
Related to this diversionary account are the results of Tomlinson and colleagues (2009) discussed above with use of a button press as an interference condition. Results from the standard and IP cued recall conditions indicate that both NT and interference (button-press) items were decreased below baseline. This finding suggests that pressing a button creates enough interference to induce NT effects, even when tested with IP cues, and may be an alternative mechanism to Direct Suppression (Tomlinson et al., 2009). Because these diversionary and interference manipulations of the TNT task involve focusing attention on other stimuli or actions, which may cause interference or competition during retrieval, they fall under the Competitive Attentional Inhibition hypothesis of inhibition of retrieval.
4.2 Integration of Behavioral Studies
This section provides a brief integration of behavioral results and how they relate to the three previously outlined hypotheses of inhibition of retrieval. Results supporting the Competitive Attentional Inhibition hypothesis come from the WB and TNT tasks. Specifically, the best supporting evidence comes from studies that directly include a diversionary stimulus or action for individuals to divert their attention (Wegner et al., 1987; Hertel & Calcaterra, 2006; Tomlinson et al., 2009). When these conditions are included, retrieval of target memories is reduced below baseline and sometimes below normal NT conditions. The explanation provided for these findings is that the diversionary stimulus or action becomes associated with the original cue and interferes with the target during retrieval attempts. This idea directly constitutes the Competitive Attentional Inhibition hypothesis in that interference may result in the diversionary stimulus or action to outcompete the target stimulus, presumably resulting in collateral inhibition of the target’s representation. Therefore, the efficacy of retrieval is based on the strength of the diversionary representation to compete with the original target representation. This competition is exemplified best in the case where individuals are given a specific stimulus or action to divert their attention, which likely induces repetition and subsequent increases in the strength of the diversionary stimulus’ representation (Wegner et al., 1987; Hertel & Calcaterra, 2006; Tomlinson et al., 2009).
The Competitive Attentional Inhibition hypothesis may also gain support from the fact that ineffectual inhibition of retrieval is widely observed in the WB task when no overt diversionary stimulus is employed. One explanation for these findings is that the task instruction is directly associated with the target stimulus of control: “Don’t Think of a White Bear”. Because the target stimulus is combined with the overt task goal (likely maintained by WM), the resultant representation of the White Bear may be extremely salient and strong, making competition of diversionary representations ineffectual. The Competitive Attentional Inhibition hypothesis is concordant with some notions of interference between LTM representations, although it is more plausible that interference occurs between more abstract WM tags within PFC that may access the diversionary and original target representations. This idea is based on the importance of a mechanism involving collateral inhibition, such that inhibition is caused automatically by spatially close representations competing for expression.
Several lines of evidence support the Direct Inhibition hypothesis, which has also been referred to as retrieval inhibition (Bjork, 1989) and directed suppression (Levy & Anderson, 2002). However, these same results also provide support for the Reactivation Inhibition hypothesis as well. The differentiation of these two hypotheses lays in the proposal that inhibition either: (a) directly inhibits a specific target memory representation, or (b) modulates more generally the components of the retrieval process, respectively. This distinction is largely based on evidence from neuroimaging and neuroanatomy, so the behavioral evidence does not yet distinguish one from the other. Therefore, results supporting both hypotheses come from the DF and TNT tasks (Bjork et al., 1989; Anderson & Green, 2001; Levy & Anderson, 2002). As the item method of the DF task yields differing levels of recognition between TBR and TBF items suggesting differential encoding for the two types of items, only results of the list method DF studies are immediately relevant, as they relate more to control of retrieval processes, per se. Because TBR and TBF stimuli in the list method of DF task show equivalent recognition rates (Wilson & Kipp, 2002), and since all stimuli in the TNT task are learned to a criterion (Anderson & Green, 2001; Depue et al., 2006), both tasks provide evidence that encoding processes have been engaged to a similar degree. Thus, reductions in recall for TBF and NT items most likely occur during the retrieval process. One manner in which this may happen is that the specific LTM memory representation of the target stimulus has been weakened by inhibitory control by the PFC (Direct Inhibition hypothesis). Another manner in which this may occur is that the PFC communicates with the hippocampus to generally modulate its output (Reactivation Inhibition hypothesis). To illustrate that the actual representation of the target memory has been affected, the results from TNT studies using IP cued recall are commonly used (Anderson & Green, 2001), although Tomlinson and colleagues have suggested that TNT results from IP cued recall can also be attributed to increased interference.
These differing accounts of IP cued recall findings can be more simply illustrated by questioning where interference and competition occurs in regard to the memory representation. If diversionary representations are linked only in direct relation to the cue, then it is possible that the target representation has been inhibited. Alternatively, if the diversionary representations are linked to the target as well, then interference or competition may affect the target, and direct inhibition of the representation may not be a necessary mechanism for reductions of retrieval.
I would suggest that this question may seem futile, because the construct of cue and target are in essence artificial concepts that investigators have created to study memory phenomena. It may be more worthwhile to discuss each as component representations of the larger composite memory representation. If so, then IP effects would require that new learning of a diversionary representation would be associated to only one component (the cue) of the composite representation, where the cue and target are likely highly associated. In order for this to occur, learning would have to only associate the diversionary representation to the cue with no reactivation and association to the target component, which seems unlikely. Thus, if any new learning occurs in connection with a component, it likely is associated with the composite memory representation as well. Because this idea is highly speculative, more investigation is required. Therefore, the results from IP cued recall can currently be associated with both Competitive Attentional Inhibition and Direct Inhibition hypotheses.
In sum, the behavioral results from WB, DF and TNT tasks can be used to support some aspects of all three hypotheses of inhibition of retrieval. Because the behavioral evidence does not elucidate the finer-grained mechanisms that each hypothesis may employ, evidence has yet to support one hypothesis over another. Therefore, examination of the results from neuroimaging studies using the same three tasks above should provide a better understanding of the mechanisms of inhibition of retrieval.
4.3 Neuroimaging Studies
In parallel with the behavioral studies section, the neuroimaging data are discussed with regard to the specific task used (i.e., WB, DF, and TNT). Results from fMRI and event-related potential (ERP) studies are analyzed in terms of contrast activity, such that increases in BOLD activation or ERP signal in one condition is compared in relation to other conditions. In general, findings are placed in the context of greater activation or signal in the condition that requires attempts to inhibit retrieval as compared to conditions that (i) increase retrieval (i.e., TNT; NT>T), (ii) simply retrieve memory information (i.e., DF TBF>TBR), or (iii) think freely (i.e., WB; Suppress>Think). Exceptions to this general format of contrasts will be noted for individual studies.
While fMRI evidence is more intuitive because of distinct regional brain activations indicating possible functionality, ERP evidence requires a brief introduction. When discussing ERP evidence, findings are presented in terms of “ERP components or effects,” which are distinct waveforms that indicate the spatial and temporal properties of neural processes (i.e., attention, memory). These ERP components have long-standing terminological conventions in relation to retrieval processes. Four main ERP components will be discussed: i) the FN400 or frontally mediated old/new effect, which is elicited during recognition memory. This early ERP component (~300-500 msec post-stimulus) localized to midline frontal electrodes shows greater amplitude for stimuli that have been previously memorized than to new stimuli. This component is perhaps indicative of facilitated access to information (Rugg &Curran, 2007); ii) the parietal old/new effect, which is elicited during recollection memory. This later ERP component (~400-800 msec post-stimulus) localized to parietal and temporal electrodes shows greater amplitude for stimuli that have been previously memorized than to new stimuli. This component is perhaps indicative of facilitated retrieval of information (Rugg & Curran, 2007); iii) the right frontal old/new effect, which is elicited post retrieval. Also a later ERP component (~500-1000 msec post-stimulus) localized to right frontal electrodes that shows greater amplitude for successfully than unsuccessfully retrieved stimuli. This component is perhaps indicative of the reconstruction and/or organization of retrieved memory (Rugg &Curran, 2007); and iv) the N2, which is elicited during tasks that require inhibition of a prepotent response (e.g., Stop-Signal, Go-No-Go).This is an early ERP component (~200 msec post-stimlus) localized to midline or right lateralized frontal electrodes that shows greater amplitude for stimuli requiring inhibition than stimuli with no inhibitory requirement. Thus, this component is perhaps indicative of inhibitory processes of cognitive control (Ullsperger et al., 2000).
In order to evaluate the neuroimaging results and illustrate support for each hypothesis, I will first illustrate hypothetical fMRI brain activations and ERP components that may be expected for each hypothesis (Table 1). Direct Inhibition postulates that the specific, individual neural representation of single target memory is affected by inhibitory control by the PFC. Thus, fMRI results may indicate increased activations of the lateral prefrontal cortex (LPFC), associated with inhibitory control [e.g., specifically right inferior frontal gyrus (rIFG) and right middle frontal gyrus (rMFG)] would be expected. Because inhibitory control is specifically postulated to target a single memory representation, it is unlikely that the sensitivity of fMRI would be able to dissociate such inhibition of a single representation from surrounding noise. Therefore, it is unclear what activations would be indicated, beyond the LPFC. ERP results may be expected to show decreased signal in the FN400 underlying recognition, because the actual representation has been inhibited and subsequent access to it should be reduced. Similarly, decreased signal in parietal and right frontal old/new effects would be expected, because this would indicate retrieval or reconstruction processes of specific memories have been reduced. Furthermore, an increase in the N2 may be seen as well, indicating an increase in inhibitory control (Table 1).
Table 1.
Hypothetical results from fMRI and ERP in relation to three hypotheses of inhibition of retrieval.
| Hypothesis: Results: | Competitive Attentional Inhibition | Direct Inhibition | Reactivation Inhibition |
|---|---|---|---|
| fMRI NT>T | |||
| PFC Activation (cognitive/inhibitory control process) | Increased/Different/No Change | Increase | Increase |
| Posterior Activation (memory retrieval/sensory processing) | No Change | No Change | Decrease |
| ERP NT>T | |||
| FN400 Old/New Effect (recognition memory) | Decrease | Decrease | No Change |
| Parietal Old/New Effect (recollection memory) | Decrease | Decrease | Decrease |
| Right Frontal Old/New Effect (reconstruction memory) | Decrease | Decrease | Decrease |
| N2 Effect (cognitive/inhibitory control process) | No Change | Increase | Increase |
Reactivation Inhibition may also be expected to show increased activation in LPFC regions associated with inhibitory control. One clear difference between this hypothesis and Direct Inhibition suggests that inhibition is generally focused on the retrieval process by interrupting pattern completion. Reducing the likelihood of pattern completion would result in decreased fMRI activation in posterior cortex specifically, in regions supporting memory representation (e.g., hippocampus, visual cortex). A second difference between this and the Direct Inhibition hypothesis, is that the ERP FN400 effect would not indicate any reduced signal. The main reason for suggesting this difference is that during Direct Inhibition the single neural representation of the memory is targeted for inhibition. This hypothetically would affect the state of the entire ensemble and has been suggested to be a lasting influence over many trials that inhibit the memory. Whereas, during Reactivation Inhibition only the retrieval process is affected and thus it first requires quick recognition of a cue that feeds forward to the hippocampus and then retrieval is subsequently altered. Because Reactivation Inhibition is hypothesized to modulate the retrieval process of the hippocampus, access to information sensitive to recognition should be spared and thus, the FN400 (reflecting recognition) should be one example of this dissociation. Conversely, reductions in the parietal and right frontal old/new effects should be found, suggesting that retrieval or subsequent reconstruction processes have been reduced. Increases in the N2 may be seen as well, indicating an increase in inhibitory control (Table 1).
A major difference between the Reactivation vs. Direct Inhibition Hypotheses is whether inhibition is generally directed towards a cortical region (i.e., hippocampus) or a specific neuronal representation of a particular memory. It should be noted that using the neuroimaging literature to differentiate the two hypotheses relies on the notion that inhibitory influence over a single neural representation is largely undetectable by fMRI. Unfortunately, this remains largely an unclear question by current resolution of fMRI and the dearth of single cell/intracranial recordings in these tasks. However, standard fMRI techniques using 3mm isotropic voxels represent tens of thousands of neurons (Hassabis et al., 2009; Leopold & Wilke, 2005). Moreover, because of spatial smoothing and registration of individual brains to a standard template, the likelihood of specific single memories from different individuals overlapping and contributing to the BOLD response appears to be highly unlikely.
During Competitive Attentional Inhibition, regions of the PFC may show increased fMRI activation as well, although because this hypothesis is largely based on focusing attention on diversionary stimuli, perhaps different regions of the PFC than those predicted in the other two hypotheses will be found. Alternatively and perhaps more likely, no differences in PFC activation may be found by focusing attention on diversionary stimuli vs. a target stimulus, because both processes likely involve attention, retrieval and maintenance of information. The accessibility to memory information indicated by the FN400 should be reduced, because competing representations have an inhibitory affect on the target representation through local collateral inhibition. Similarly, parietal and right frontal old/new effects may be reduced, as diversionary stimuli may become more accessible to retrieval and reconstruction processes (Table 1).
While these predictions differentiate the three hypotheses in the inhibition of retrieval, it must be noted that using neuroimaging evidence to do so, introduces a problem of forward inference (using qualitatively different patterns of brain activity to differentiate between cognitive theories). Although significant strides have been made in formalizing appropriate inferences (i.e., reverse, forward) that can be made using neuroimaging, there still is debate concerning the most reliable way to estimate cognitive processes from neuroimaging data. In order to best deal with this problem, the former predictions attempt to use multiple neuroimaging methods (i.e., ERP, fMRI) and results to provide complimentary data. Of course, there still will be the problem of forward inference, which hopefully will lead to testing of the specific predictions of these hypotheses and the resultant model.
4.3.1 White Bear (WB) Experiments
Only two neuroimaging studies examine inhibition of retrieval as assessed by the WB task. One fMRI study examined the neural mechanisms employed under three conditions: suppression of a relevant occurring thought (suppress), clear the mind of all thoughts (clear), or think freely while fixating (free thought) (Wyland et al., 2003). Although not identical to standard WB studies, the procedure is essentially the same, except that cognitive control is focused on a current thought or all thoughts as opposed to a white bear. Results indicate increased activation (suppress and clear vs. free thought) in the anterior cingulate cortex (ACC), bilateral insula/IFG (BA 44/45/46), and right parietal cortex. The authors suggest that increased activation of the ACC represents monitoring of conflict during unsuccessful attempts at suppression/clear instructions. Furthermore, increased activation of right insula/IFG replicates studies that have identified cognitive control regions during the inhibition of behavioral responses and/or somatosensory information. Thus, increased activity in these LPFC regions associated with inhibitory control support both the Direct and Reactivation Inhibition hypotheses of inhibiting retrieval. Because no reduced activation was seen in posterior cortex, only partial support for the Reactivation Inhibition hypothesis of inhibiting retrieval is apparent.
The second fMRI study used the WB task to assess the differential role of cognitive control as a sustained process vs. transient process (Mitchell et al., 2007). Although the study is framed within cognitive control processes, the results are informative of possible mechanisms involved when inhibiting retrieval. Although the behavioral results showed no difference in thought occurrence (measured by a button press), the imaging data provide some interesting findings. fMRI data was analyzed in a sustained (block) versus transient (single trial) manner. Sustained activity (suppression vs. free thinking) was found in rMFG, rIFG, and the caudate. Transient activity (suppression vs. free thinking) was found in parietal and occipital cortex and the ACC. The authors propose that sustained activations indicate aspects of greater cognitive control during the suppression condition and that transient activation of the ACC may be involved in conflict monitoring of unsuccessful attempts during the suppression condition. Increased activation of rMFG and rIFG associated with inhibitory control supports the Direct and Reactivation Inhibition hypotheses of inhibition of retrieval. Again, because no reduced activation is seen in posterior cortex, only partial support for the Reactivation Inhibition hypothesis of inhibiting retrieval is apparent.
4.3.2 Directed Forgetting (DF) Experiments
Few neuroimaging studies have examined inhibition of retrieval using the DF task, but those that do use the item method. Because the item method frequently indicates increased recognition of TBR vs. TBF material, this method of the DF task is usually associated with encoding rather than retrieval processes. Nonetheless, the studies included here are framed under selective rehearsal vs. inhibitory mechanisms (see behavioral DF section), and thus they may provide insight into the neural mechanisms that underlie reductions in memory information during retrieval.
An fMRI study by Wylie and colleagues (2007) conducted with the item method DF task using words showed behavioral recognition DF effects, while fMRI during the study phase revealed increased activity (TBF>TBR) in medial frontal and temporal lobe regions (Wylie et al., 2007). When the authors examined which trials were later remembered vs. forgotten, increased activation was seen in inferior parietal and hippocampal regions. The converse contrast (forgotten vs. remembered) showed increased activation in the rIFG. The rIFG also exhibited the greatest increase in activity for TBF trials that were subsequently forgotten, perhaps indicating increased inhibitory control. This study raises the possibility that attempts to control memory in the item method DF task are not only caused by selective rehearsal, but perhaps by inhibitory mechanisms as well.
Recently, Nowicka and colleagues (2010) examined differences between emotional and neutral pictures using fMRI during the item method DF task. While both types of stimuli showed behavioral recognition DF effects, negative as compared to neutral images showed reduced DF. fMRI results during the study phase showed increased activation (TBF>TBR) of rMFG, right temporal, occipital and parietal lobes. Examining only correctly forgotten trials revealed similar activations. During the test phase forgotten vs. new stimuli elicited no differences in activation, indicating a lack of retrieval for both items. The authors suggested that these results indicate that successful forgetting in the item method DF task reflects inhibitory control by right hemispheric regions. Furthermore, this control may function to inhibit encoding during the study phase, which subsequently reduces the amount of information accessible at retrieval during the test phase.
Two ERP studies using the item method DF task with words also assessed selective rehearsal vs. inhibitory mechanisms (Ullsperger et al., 2000; Van Hoof et al., 2009). Behavioral recognition results from both experiments indicated a DF effect. The FN400 showed decreased signal for TBF vs. TBR items, while the parietal old/new effect appeared absent for TBF items (Ullsperger et al., 2000). The authors suggest that the decreased FN400 ERP signal is consistent with the idea that TBF items are encoded differentially, but that the lack of parietal old/new ERP signal is indicative of retrieval inhibition (Ullsperger et al., 2000). Examining ERP effects of forgotten vs. remembered items revealed that remembered items yielded increased FN400 and parietal old/new effects, while only TBF-forgotten items showed decreased parietal old/new effects and an increased N2, as compared to New items (Van Hoof et al., 2009). These results indicate increased rehearsal, which is apparent in recognition testing and increases in the FN400, although the absence or decrease in the parietal old/new and increased N2 effect for TBF-forgotten items may be indicative of inhibition at retrieval stages.
In a recent paper, Ludowig and colleagues (2010) investigated the item method DF task using intracranial ERP recordings localized to the rhinal cortex and hippocampus in patients undergoing surgery for temporal lobe epilepsy. They propose that the rhinal cortex may be more highly active in inhibitory conditions, because it acts by modulating information from the neocortex to the hippocampus in a gating fashion. Behavioral results indicated that patients recognized significantly less TBF than TBR words, perhaps indicative of selective rehearsal. ERP effects during cue presentation showed that TBF vs. TBR cues produced increased signal in the rhinal cortex and reduced signal in the anterior hippocampus. These findings suggest that, although behavioral recognition results support selective rehearsal, ERP data show a possible mechanism of inhibition, whereby the rhinal cortex inhibits the flow of information being transmitted to the hippocampus.
Because these studies provide inconclusive results on retrieval processes, they cannot be used to support hypotheses of inhibition of retrieval. Nevertheless, these studies constitute a large portion of the neuroimaging literature and indicate that item method DF tasks used to examine inhibiting retrieval may involve alterations of encoding, as opposed to retrieval processes. These issues will be more fully addressed in the integration of neuroimaging studies section below.
4.3.3 Think/ No-Think (TNT) Experiments
A significant portion of neuroimaging studies examining inhibition of retrieval use the TNT task. The influential first study using word stimuli showed behavioral reductions of NT as compared to both T and baseline trials (Anderson et al., 2004). fMRI results indicated increased activation (NT>T) in MFG, IFG, ACC and parietal activity, whereas the occipital lobe and hippocampus showed decreased activity. Furthermore, correlations with success at inhibiting retrieval during IP and standard cued recall testing predicted increased activation of the MFG. Additional analyses also indicated that NT-forgotten items actually elicited increased activation of the hippocampus as compared to all other trials. The authors suggested that the increased hippocampal activation is indicative of intrusions during NT trials that elicit subsequent increased inhibitory control by the MFG. These findings signify that reductions in memory retrieval are associated with inhibitory control processes of the MFG in interaction with the hippocampus. Increased activation of the MFG associated with inhibitory control supports the Direct and Reactivation Inhibition hypotheses. Additionally, reduced activation in the hippocampus and specifically the visual cortex suggests that inhibitory modulation may be more generally affecting the retrieval process and pattern completion, further supporting the Reactivation Inhibition hypothesis of inhibition of retrieval.
Two fMRI studies by Depue and colleagues (2007; 2010) using the TNT task with emotional/pictorial stimuli produced similar results. The first study showed behavioral reductions of NT as compared to both T and baseline items. fMRI results showed increased activity (NT>T) in rMFG and rIFG. Decreased activation was seen in visual cortex, the pulvinar nuclei of the thalamus, the hippocampus and amygdala. Moreover, when examining NT vs. baseline trials, posterior regions showed reduced activity raising the possibility that these regions are actively being down-regulated. Furthermore, analyzing the data in quartiles (over the experimental phase) and correlating activity between the two PFC and four posterior regions demonstrated specific groupings of regions that were recruited at different stages. Increased activation of the rIFG inversely correlated with decreased activation of visual cortex and thalamus during the first attempts at inhibiting retrieval. In contrast, increased activation of the rMFG inversely correlated with decreased activation of the hippocampus and amygdala after additional attempts at inhibiting retrieval. This last grouping also correlated with behavioral success during NT trials such that increased activation of the rMFG and decreased activation of the hippocampus were the only two regions that predicted increased inhibition of retrieval in a whole brain fMRI analysis (Depue et al., 2007). Therefore, inhibition of retrieval appears to be a complex process that may involve multiple mechanisms employed over the process of learning inhibitory modulation. The most robust finding is the MFG’s inhibitory modulation of the hippocampus.
To further investigate the idea that LPFC interaction with the hippocampus is a critical part of inhibiting retrieval, percentage signal change of the hippocampus across quartiles was examined (Depue et al., 2007). Results indicated that NT-forgotten trials elicited the greatest increase in activation as compared to all other trials (T or NT vs. baseline, remembered or forgotten) during the first half of the experiment, while conversely the largest decrease in activation below baseline was observed during the fourth quartile. The initial increase in hippocampal activity raises the possibility that inhibition of retrieval may induce a learning mechanism that associates an inhibitory process to NT stimuli.
The second study by Depue and colleagues (2010) examined whether LPFC interaction with the hippocampus was evident in individuals with inhibitory deficits. This study examined individuals with ADHD and compared the group to control individuals investigated in the previous study (Depue et al., 2007). Behavioral results indicated that individuals with ADHD have similar recall on T and baseline trials, but show no reductions in recall during NT trials. fMRI results (NT>T) indicated that when compared to controls, some prefrontal regions exhibit similar activation (rIFG, BA10), while the rMFG, thought to be crucial in communication with the hippocampus, showed decreased-to-little activation. Supporting this former idea, brain regions outside of the PFC (i.e., hippocampus, amygdala, visual cortex) also showed increased activation as compared to controls. Furthermore, the severity of ADHD symptoms predicted a) less correlated activity between the rMFG and hippocampus, b) increased activity of the hippocampus and c) increased recall during NT trials. In an additional analysis, 5 of the 16 individuals who showed behavioral reductions in recall were compared to the 11 that did not. These 5 individuals showed greater activation (NT>baseline) in the rMFG, whereas reduced activation was exhibited in the hippocampus and amygdala during the fourth quartile, mirroring results of control subjects. These findings, once again, highlight the important interaction between inhibitory control of the rMFG and its purported modulation of the hippocampus. Taken together, the altered inhibition of retrieval findings from ADHD individuals support the explanatory power of inhibition of retrieval processes identified in nonclinical persons. LPFC activations associated with inhibitory control support both of Direct and Reactivation Inhibition hypotheses. While reduced activation in the hippocampus, pulvinar and visual cortex further supports the Reactivation Inhibition hypothesis of inhibition of retrieval.
A recent fMRI study using the TNT task examined negative and neutral word pairs (Butler & James, 2010). Behavioral differences were reported as close to ceiling, so that interpretation of recall is difficult. Nonetheless, fMRI results for neutral words show increased activation (NT>T) for rMFG, pre- and post-central gyri, and the inferior parietal cortex. Decreased activation was found in the bilateral hippocampus and precuneus. A region of interest (ROI) analysis on the hippocampus comparing both NT and T trials to baseline showed that posterior hippocampal activity was reduced below baseline for NT trials. Examination of negative words yielded increased activity (NT>T) in similar frontal regions (e.g., rMFG, rIFG, pre- and post-central gyri), the ACC and parietal cortex, although it also showed increased activation in the hippocampus, pulvinar, and visual cortex. The authors suggested that one reason that increased activation was seen in posterior cortex for negative and not neutral stimuli is that NT trials lasted half the duration of most studies (2 vs. 4 sec) and included half the repetitions (6 vs. 12 or 16) of the previous studies by Anderson and Depue and colleagues. These findings further support the interactions between the rLPFC and hippocampus during inhibition of retrieval, although in cases where stimuli are extremely salient, more repetitions may be required to achieve successful control. Increased activation of rMFG and rIFG associated with inhibitory control supports both the Direct and Reactivation Inhibition hypotheses of inhibition of retrieval. General decreases in posterior cortex more fully support the Reactivation Inhibition hypothesis.
Two ERP studies by Bergstrom and colleagues using the TNT task with word stimuli dissociated early and late components of the ERP signal (Bergstrom et al., 2007; 2009). The first study showed behavioral reductions in NT items compared to T, but not compared to baseline. ERP results were analyzed on the basis of T and NT, as well as learned vs. not learned stimuli. Parietal old/new effects were greater for Think-learned than the other three conditions, perhaps indicative of increased retrieval processes of these items. These latter results suggest that NT items regardless of learning may undergo control processes that are aimed at reducing retrieval.
The second study included an “aided” condition during NT trials in which individuals were instructed to think of a diversionary memory to distract themselves (Bergstrom et al., 2009). Behavioral results indicated that both standard NT and aided NT items were reduced below baseline. Although, an independent probe condition (IP) used during recall testing to assess memory items (NT, T) with an alternative cue (cue-independent), indicated only a behavioral reduction in NT items from both T and baseline in the standard NT condition. ERP results showed an increased N2 effect in both NT (standard, aided) as compared to T conditions, although was larger for the standard NT items. Furthermore, the N2 showed a correlation between the magnitude of this effect and individual differences in NT item IP recall only in the standard NT condition. This finding suggests that the N2 may be associated with successful cue-independent reductions of NT items. Additional frontal effects (~300-500) elicited only by the standard NT items showed the greatest increase in signal for NT-forgotten trials as compared to, intermediate levels for NT-remembered trials and the least increase in T-remembered trials. Parietal old/new effects were decreased in the standard NT but not in the aided NT condition. These results indicate that standard NT trials elicit increased N2 and later frontal effect that is subsequently followed by reductions in parietal old/new effects, perhaps further indicative of a frontal control mechanism aimed at retrieval processes.
Another ERP study examined both the TNT task and a Stop-signal task (response inhibition) to assess the similarity of ERP components that may be involved during the inhibition of memory and motor information (Mecklinger et al., 2009). Although behavioral results showed no effect of NT items in terms of reduction below baseline, NT items did show similar ERP effects to other studies (Bergstrom et al., 2007; 2009), in that a decreased parietal old/new effect was seen for NT as compared to T items. An increased N2 effect was also present in NT>T trials and also in NT-forgotten vs. NT-remembered trials. Furthermore, correlations were found for the N2 component from the TNT and Stop-signal tasks, indicating a positive relationship between inhibition of memory and motor information. Increased N2 effects support both Direct and Reactivation Inhibition hypotheses of inhibition of retrieval.
An interesting ERP study examined how anticipatory cues affect the ERP components associated with the TNT task using face-word pairs (Hanslmayr et al., 2009). Red (NT) and green (T) crosses were presented as anticipatory cues 1 sec prior to the T and NT trials to assess anticipatory control. This study also included a separate repetition manipulation that assessed recall for NT and T trials at 5 and 10 attempts. Behavioral results showed no reduction in recall for NT items compared to baseline at 5 repetitions, but reduced recall after 10 repetitions. ERP results indicated an early and late effect that changed over the course of repetition. As repetition increased, reduced signal in the early slow-wave frontal component and later slow-wave parietal component was seen in NT trials, perhaps indicative of increased control manifest in early NT repetitions. The frontal component, furthermore suggests, control in response to the anticipatory NT cue, which consequently results in reduced retrieval as indicated by the later reduced parietal component. Supporting this view, the two ERP effects were highly correlated, such that magnitude of the early effect predicted the magnitude of the later. These results indicate an increased early slow-wave frontal component (which may be functionally related to the N2) elicited by anticipatory cues, subsequently followed by a reduced parietal component (perhaps functionally relate to parietal old/new effects) elicited during NT trials.
4.4 Integration of the Neuroimaging Studies
This section provides a brief integration of neuroimaging results and how they relate to the three previously outlined hypotheses of inhibition of retrieval. While the behavioral integration supported the framework of the three hypotheses, it lacked the ability to dissociate support for them. Because neuroimaging provides more detailed evidence of the mechanisms of inhibition of retrieval, support has emerged for one hypothesis over the others, which is illustrated in Table 2 and discussed below.
Table 2.
Neuroimaging results from individual studies and hypotheses they support (– indicates no support, x indicates partial support and X indicates full support).
| Hypothesis: Study: | Competitive Attentional Inhibition | Direct Inhibition | Reactivation Inhibition |
|---|---|---|---|
| Wyland et al., 2003 (WB, fMRI) | — | X | x |
| Mitchell et al., 2007 (WB, fMRI) | — | X | x |
| Wylie et al., 2007 (DF, fMRI) | — | — | — |
| Nowicka et al., 2010 (DF, fMRI) | — | — | — |
| Ullsperger et al., 2003 (DF, ERP) | — | — | — |
| Van Hoof et al., 2009 (DF, ERP) | — | — | — |
| Ludowig et al., 2010 (DF, EEG) | — | — | — |
| Anderson et al., 2004 (TNT, fMRI) | — | x | X |
| Depue et al., 2007 (TNT, fMRI) | — | x | X |
| Depue et al., 2010 (TNT, fMRI) | — | x | X |
| Butler & James, 2010 (TNT, fMRI) | — | x | X |
| Bergstrom et al., 2007 (TNT, ERP) | — | x | X |
| Bergstrom et al., 2009 (TNT, ERP) | — | x | X |
| Mecklinger et al., 2009 (TNT, ERP) | — | x | X |
| Hanslmayr et al., 2009 (TNT, ERP) | — | x | X |
Before discussing evidence for each hypothesis, it is necessary to briefly address the item- method DF task results, because they are not used to support any hypothesis. While the task instructions and requirements are overtly and generally the same as the list method DF, WB and TNT tasks, it is evident that the item method DF involves mechanisms that affect encoding processes that subsequently reduce recall. In contrast, the neuroimaging studies produced some commonalities that indicate shared mechanisms across the WB and TNT tasks. Results from fMRI studies indicate that increases in rLPFC (rMFG, rIFG) may support inhibitory control during retrieval, as they are exhibited in both WB and TNT tasks (Wylie et al., 2007; Nowicka et al., 2011). ERP results have illustrated decreases in the parietal old/new effect (Ullsperger et al., 2000), as well as increases in the N2 effect (Van Hoof et al., 2009), also found in both WB and TNT tasks. The joint findings of increased activity of rLPFC regions, decreased signal in the parietal old/new effect, and increased signal in the N2 effect indicate that the item method of the DF task may induce inhibitory control over encoding processes in a similar manner as retrieval processes. The degree to which these mechanisms overlap will be further discussed in the model section below.
While behavioral evidence suggested that Competitive Attentional Inhibition may cause reductions in retrieval, there is little evidence from neuroimaging that this mechanism is solely responsible for inhibition of retrieval. LPFC activations elicited by all three tasks are significant, because these brain regions have previously been strongly related to controlled inhibitory processes across several psychological domains. While the hypothesis of Competitive Attentional Inhibition involves an inhibitory influence, it does so in a relatively automatic manner, such that diversionary representations of stimuli compete for expression and may induce local collateral inhibition. It is unlikely that competitive inhibition at an individual neural representation level would be detected through fMRIs sensitivity and as such, it would not produce such strong persistent LPFC activation in regions common to controlled inhibition seen in the studies reviewed. Similarly, there is no indication that competitive inhibition would induce decreases in activation in the hippocampus, as the competition is likely to occur in PFC regions that involve WM (discussed previously). Further support from ERP evidence shows that the N2 effect is commonly found in studies across all three tasks, indicating controlled rather than automatic inhibition. Therefore, taken together, the evidence leaves little support for the Competitive Attentional Inhibition hypothesis of inhibition of retrieval.
As in the behavioral integration, it is more appropriate to address the Direct and Reactivation Inhibition hypotheses together, because the evidence used to differentiate the two is subtle, yet critical. Results from fMRI experiments using the WB and TNT tasks have clearly shown increased activation of regions of LPFC (MFG, IFG), more prominently right hemispheric, and both of these findings are putatively thought to contribute to inhibitory control (Wyland et al., 2004; Mitchell et al., 2007; Anderson et al., 2004; Depue et al., 2007; 2010; Butler & James, 2010). Multiple ERP results from the TNT Task indicate increased N2 effects, again suggesting increased inhibitory control. Indicative of this inhibitory control, reduced parietal and right frontal old/new effects are found, suggesting decreased retrieval and reconstruction processes (Bergstrom et al., 2007; 2009; Mecklinger et al., 2009; Hanslmayr et al., 2009). This evidence equally supports both hypotheses.
A key differentiation between the hypotheses occurs when examining the function of the MFG. Clear evidence supports that this region shows an interaction with the hippocampus. Multiple studies show an increase in activation of MFG and an associated decreased activation of the hippocampus for NT as compared to T and baseline trials in whole brain and ROI analyses (Anderson et al., 2004; Depue et al., 2007; 2010; Butler & James, 2010). Correlations between these two regions, as well as correlations of each of these regions independently (Anderson et al., 2004; Depue et al., 2007), and their combined correlation (Depue et al., 2010) predict behavioral success at inhibiting retrieval. These results indicate that the most evident mechanism supporting inhibition of retrieval involves the MFG’s modulation of the hippocampus. Furthermore, reduced activation in the cortices that support memory (visual cortex, fusiform gyrus, parahippocampal gyrus) is commonly found (Anderson et al., 2004; Depue et al., 2007; 2010; Butler & James, 2010). According to the Direct Inhibition hypothesis, inhibitory control targets a specific memory representation to reduce its activation and subsequently to stop its retrieval. If this were the case, it is unclear how this mechanism would account for fMRI results indicating: i) reduced activation in the hippocampus, and ii) reduced activation in posterior cortex. The most plausible explanation for these findings is that inhibition is more generally affecting the array of regions involved in the retrieval process of the hippocampus. If so, then reactivation of memory representations by pattern completion through feed-back activation of the hippocampus to the cortices supporting memory (para-hippocampal regions, visual cortex) would be reduced and may account for such results. Thus, these findings support the Reactivation Inhibition hypothesis.
It should be noted that results of deactivation of the hippocampus has received some criticism regarding the veracity of such findings. Mainly, the objection is that if one were to compare a condition of interest (i.e., NT, TBF) to a baseline condition, the results may be influenced by increased activation of the hippocampus during periods of baseline. Regardless, the logic still remains that if hippocampal activity shows the pattern of T>NT, combined with T>baseline and NT<baseline, hippocampal activity is somehow being modulated or reduced in the NT condition. Whether or not hippocampal activity is decreased in the contrast of a NT condition to a hippocampally active baseline should not matter. Indeed, if the hippocampus is normally active during periods like under-constrained mental tasks (i.e., baseline), then one would have to further argue that during NT conditions the hippocampus appears to be down-regulated from its normal active state.
While the current review focuses on inhibition of memory retrieval, it should be noted that some neuroimaging studies examining intentional retrieval and subsequent memory effects, have shown decreases in hippocampal activation for memories that were retrieved vs. not retrieved. This may be because retrieved memories could be less neurally or process demanding, which may lead to a faster response and thus reduced hippocampal activation, than ones that are attempted but unsuccessful (more difficult). In contrast, there are multiple studies highlighted in the current review indicating systematic reductions of activity or deactivation of the hippocamus correlated with decreases in n memory performance (Depue et al., 2007, 2010; Anderson et al., 2004). Therefore, at least in the context of tasks that require one to inhibit retrieval, it appears that reduced activity or deactivation of the hippocampus predicts worse memory retrieval. Similarly, cell assemblies of memory representations may go through a pruning process to limit the number of neurons in a cohesive memory representation or ensemble. Although, the consequence of these effects on resultant BOLD response related to hippocampal function remains unclear, but should be noted.
To illustrate a clearer picture of the fMRI findings and how they differentiate the specific Direct Inhibition from the more general Reactivation Inhibition hypotheses, meta-analytic tools (GingerALE; Turkeltaub et al., 2002) were used to combine all fMRI results across the three tasks. This analysis helps to illustrate common and overlapping PFC, hippocampus, and posterior cortical regions involved in inhibition of retrieval (Figure 2). Two different overlays were constructed without statistical inference to simply provide a visual representation of the fMRI data. The first overlay includes positive activation (NT>T) of PFC regions (anterior of post-central gyrus), while the second includes negative activation (T>NT) of posterior regions (posterior of post-central gryus, not including the cerebellum). All peaks of activity selected from the original studies were transformed into MNI if listed in Talairach space and a 10mm sphere was localized around the peak. The goal of this visual representation was to plot the findings of all studies that revealed frontal activations to illustrate common regions of PFC involved in inhibition of retrieval. Moreover, posterior cortex was also plotted to illustrate what occurs in the rest of the brain perhaps as a result of such PFC control. The analysis clearly illustrates rMFG and, to a smaller extent, rIFG as being the most commonly activated PFC regions involved across all fMRI studies. The posterior cortex clearly shows decreased activation in the ventral visual pathway moving anterior to parahippocampal and hippocampal regions.
Figure 2.
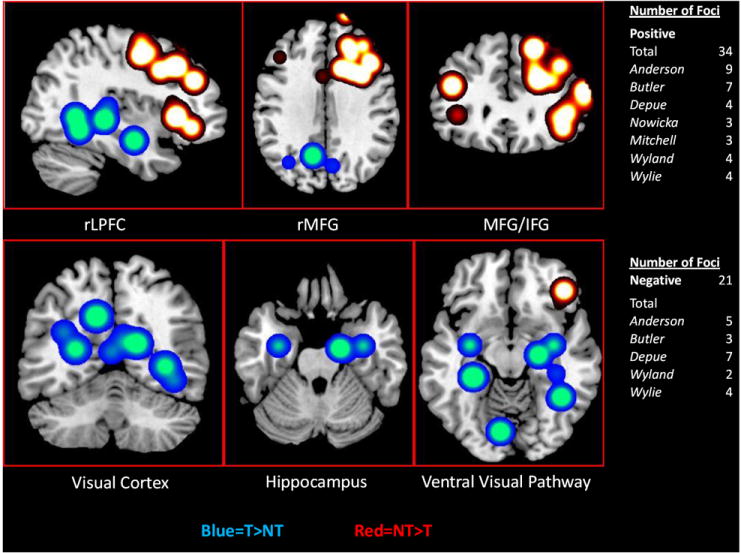
Cumulative fMRI results across all three tasks using meta-analysis techniques to illustrate common PFC (anterior to pre-central gyrus) and posterior cortical regions (posterior to pre-central gyrus) involved in inhibition of retrieval. Hot colors represent greater activation (NT>T), while cold colors represent negative activation (T>NT).
While this evidence has already been discussed individually, combining the results in the overlays gives a very strong visual representation of the confluence of wide spread deactivation in the hippocampus, parahippocampal gyrus, visual cortex and ventral visual processing stream. Thus, these results may indicate a more general inhibition of retrieval that would consequently reduce pattern completion and reactivation in posterior cortices rather than an inhibition of a specific single memory representation, once again supporting the Reactivation Inhibition hypothesis. Importantly, in order for Direct Inhibition to be as equally plausible, inhibition of a specific memory representation would have to account for such widespread reduced activation in the brain, which seems unlikely.
Further evidence providing a distinction between hypotheses comes from the FN400 ERP results. The FN400 is proposed to indicate reduced access to memory, as it is sensitive to recognition tests. This is clearly indicated in the item method of the DF task, where decreased FN400 effects are seen for TBF vs. TBR, as well as for forgotten vs. remembered information. This has been proposed to occur due to reduced encoding, resulting in a weaker representation (Rugg & Curran, 2007). Support for the Direct Inhibition hypothesis then should show decreases in the FN400 effect, because the representation of the memory has been inhibited, thus creating reduced access and recognition. Alternatively, if the retrieval process were more generally reduced, as described by Reactivation Inhibition, it would largely leave the FN400 and recognition intact. None of the reviewed studies outside of the item method DF Task (attributed to reduced encoding) produced differences in the FN400. While this represents a null finding and is worthy of further investigation, it provides continuing support for the Reactivation Inhibition hypothesis.
5. An Anatomical and Functional Model of Inhibitory Modulation of Memory Retrieval
The Reactivation Inhibition hypothesis, while supported by the behavior and neuroimaging literature, is rather incomplete unless it can be modeled with a more anatomical and functional specificity. Therefore, I attempt to illustrate a model using literature from comparative neurology/anatomy to include more fine grained neural detail. The model can then be abstracted theoretically to understand how Reactivation Inhibition serves as the primary functional mechanism of inhibition of retrieval. The goal is to provide a testable model that can guide future research examining not only inhibition of retrieval, but the broader conception of how memory information is flexibly controlled. With respect to the latter point, it will be illustrated in the model, based on the previously discussed literature, that inhibition of retrieval (as a general inhibitory concept) may include inhibitory modulation of both encoding and retrieval processes.
As a starting point, this model involves higher-order cognitive control regions; most prominently indicated by the reviewed literature is rMFG and less frequently rIFG. These brain areas are strongly implicated as being involved in cognitive, and more specifically inhibitory, control (Aron et al., 2004; Phan et al., 2005; Eisenberger et al., 2003; Ochsner et al., 2002). Detailed research examining rIFG suggests that its function is highly involved in response inhibition, and that specifically it may monitor for infrequent stimuli that require withholding of responses (Aron et al., 2004; Aron & Polldrack, 2006; Chatham et al., in press). Therefore, some inhibition of retrieval tasks have elicited increased activation of rIFG (Depue et al., 2007; Butler & James, 2010), because this region detects or monitors for stimuli (cues) associated with inhibitory responses.
The remainder of this section focuses on modeling of the rMFG, as its function in inhibitory processes has yet been defined, and it is the PFC region most commonly identified in the current review. The most frequent observation is the interaction of rMFG and the hippocampus/MTL, where increased rMFG control likely down-modulates the hippocampal retrieval process. This inhibitory effect subsequently interrupts pattern completion and the reactivation of composite memory representations, making inhibiting retrieval efficacious (Anderson et al., 2004; Depue et al., 2007; 2010; Depue & Banich, 2011; Butler & James, 2010. Moreover, a study of individuals with ADHD demonstrated a lack of behavioral inhibition of retrieval associated with a specific disruption in the rMFG-hippocampal pathway (Depue et al., 2010). Therefore, it is of critical importance to understand how interaction between the MFG and hippocampus/MTL may function to inhibit retrieval.
5.1 Three Neuroanatomical Pathways Connecting the PFC and Hippocampal Complex
In terms of modeling the mechanism of Reactivation Inhibition, comparative anatomy has strongly supported a reciprocal connection of the MFG and the hippocampus/MTL (Pandya et al., 1999; Morris et al., 1995; Goldman-Rakic et al., 1984). As illustrated in Figure 3, this literature has primarily focused on three pathways: lateral, medial, and hippocampal-prefrontal. The lateral pathway directly and reciprocally connects MFG areas 9/46 (dorsal/mid) to the entorhinal cortex via the fronto-occipital fasciculus (FOF) in the rat and non-human primate (Goldman-Rakic, et al., 1984). These connections underlie a direct route of areas 9/46 to anterior regions of the hippocampal complex, such as the entorhinal cortex/anterior parahippocampal gyrus, and more specifically input to the perforant path of the hippocampus proper (Fig. 3). The perforant path involves the neocortical connection of the entorhinal cortex to all subfields of the hippocampus proper (e.g., subiculum, CA’s, dentate gyrus). The entorhinal cortex projections to the perforant path are considered the main route in which multimodal cortical pathways reach the hippocampus proper for subsequent encoding. Illustrating this pathways function, damage to the anterior regions of the hippocampal complex/MTL results in gross encoding deficits (Petrides & Milner, 1982).
Figure 3.
The Lateral, Medial and Hippocampal-Prefrontal pathways (Hip = hippocampus proper, Retro = retrosplenial cortex, PS = presubiculum, Sub = subiculum, EC = entorhinal cortex, PHG = parahippocampal gyrus, VTA = ventral tegmental area, NAcc = nucleus accumbens core, GP = globus pallidus, VMPFC = ventromedial prefrontal cortex, MFG = middle frontal gyrus).
The medial pathway reciprocally connects the MFG, areas 9/46 (dorsal/mid) to the presubiculum via the cingulum bundle in the rat and non-human primate (Goldman-Rakic et al., 1984, Pandya et al., 1999; Morris et al., 1995). This pathway courses medially and posteriorly from areas 9/46 through the cingulum bundle, innervating the retrosplenium and presubiculum/posterior parahippocampal gyrus, thus connecting more posterior regions of the hippocampal complex/MTL (Fig. 3). Damage to more posterior (but not anterior) regions of the hippocampal complex result in WM deficits, suggesting that the posterior hippocampal complex/MTL is involved in output or retrieval processes of the hippocampus proper (Petrides & Milner, 1982).
Finally, the hippocampal-prefrontal pathway sends efferent projections from CA1/CA3 and the subiculum to the medial prefrontal cortex (mPFC) and less densely to LPFC through regions of the basal ganglia and thalamus in the rat, cat and non-human primate (Thierry et al., 2000; LaRoche et al., 2000; Swanson, 1981; Fig. 3). Because this pathway connects hippocampal regions to the ventral tegmental area and ventral striatum (e.g., nucleus accumbens, globus pallidus), it is thought to provide dopaminergic input to the medial and lateral pathways (Thierry et al., 2000).
All three pathways interact to modulate memory processes and allow for PFC influence or influence over PFC, either directly (lateral, medial pathways) or through a GABA-mediated striatal-pallidal-thalamo gating mechanism (hippocampal-prefrontal pathway) that opens and closes to allow information from different regions of posterior cortex to reach the PFC through the basal ganglia and thalamus (Alexander et al., 1990; Thierry et al., 2000). The hippocampal-prefrontal pathway’s connections with the nucleus accumbens core and globus pallidus allow for the mesocortical DA system to influence different aspects of memory (e.g., encoding, retrieval, maintenance; Thierry et al., 2000). One way in which this influence may be exerted is through interactions with both the lateral and medial pathways, because of common connections to the entorhinal cortex, presubiculum, and subiculum. Although the specific cell populations of the medial and lateral pathways’ projections are not well known, dense populations of GABA-mediated interneurons have been found in both the entorhinal cortex and presubiculum (Sewards & Sewards, 2003; Pinto et al., 2006; Canto et al., 2008; van Haeften et al., 1997; Manns et al., 2001). Cellular interactions of DA and GABA are highly complex at the level of the hippocampal complex and outside the scope of this review, although the interaction has been shown to both inhibit and facilitate responses in the perforant path via the entorhinal cortex (Otmakhova & Lisman, 1999; Caurana et al., 2006; Erickson et al., 2000). Thus, neurotransmitter influences on the lateral pathway may provide a means of modulating input to the hippocampus via entorhinal cortex to perforant path connections which may alter encoding/consolidation processes. Influence on the medial pathway may modulate output from the hippocampus proper at the level of the subiculum or presubiculum and therefore, be involved in selective retrieval of WM information (Pandya et al., 1999; Morris et al., 1995; Goldman-Rakic et al., 1984).
Further speculation on the influence of DA input from the hippocampal-prefrontal pathway and its effect on GABA interneurons in the projection zones of the medial and lateral pathways (i.e., entorhinal cortex, presubiculum) may indicate changes in neural oscillations. Although still tentative in their relation to memory inhibition, neural oscillations have been shown in cell assemblies, such that the timing of neural firing in various brain regions is coordinated to link or associate different components into a composite memory representation. Studies examining local field potentials in rats, as well as intracranial and magneto-encephalogram recordings in humans indicate that oscillations in the theta and gamma band locally (i.e., within hippocampus and entorhinal cortex) and globally (i.e., PFC-hippocampal interaction) may represent the synchronization of spike-timing-dependent plasticity (Hasselmo et al., 2002; Benchenane et al., 2011; Fell & Axmacher, 2011). This synchronization has been viewed as a prerequisite for LTM memory processes, such as separating encoding from retrieval dynamics and facilitating plasticity/LTP in the hippocampus and entorhinal cortex (for a review see Hasselmo et al., 2002). Infusion of muscimol (a GABAa agonist) into the medial septal areas in rats reduces theta activity, as well as theta-gamma co-modulation and subsequent memory performance (Shirvalkar et al., 2010; Chrobak et al., 1989; Givens & Olton, 1990). More strikingly, increases and decreases of PFC-hippocampal theta co-modulation (perhaps an underlying mechanism of WM) are suggested to depend on GABA interneurons that may turn on and off when PFC-hippocampal interaction is necessary for WM (Benchenane et al., 2011). Furthermore, DA injection to the PFC increases PFC-hippocampal theta co-modulation, perhaps as a result of DA increasing the strength and precision of GABA mediated interneurons (Benchenane et al., 2011). Hence, these studies indicate a linkage between DA, GABA interneurons and the effects of theta/gamma oscillations on memory processes. Therefore, inhibition of retrieval may involve MFG modulation of the hippocampus/MTL via the lateral and/or medial pathways (including DA influence from the hippocampal-prefrontal pathway), which may alter theta/gamma oscillations affecting the spike-timing synchronization of encoding (lateral pathway) or retrieval (medial pathway) processes.
5.2 Functional Implications of the Three PFC Hippocampal Pathways
Though the anatomical evidence appears to suggest that pathways exist between the MFG and the hippocampus/MTL, it still does not suggest specifically how this model is expressed functionally. That is, how would these pathways exert control to modulate the encoding or retrieval process? One determinant factor of the relative contribution of the lateral and medial pathways may depend on when control is applied to memory processes. When cues to forget are presented immediately or close to target presentation (DF task), attempts to control memory may affect the lateral pathway. In this case, the cue represents the concept that stimuli are irrelevant or should be avoided. This may trigger inhibitory control of the MFG to affect input to the hippocampus/MTL through the perforant path by increasing activity of GABA mediated interneurons in the entorhinal cortex. In turn, this may reduce spike-timing synchronization and oscillatory behavior important for continued encoding/consolidation. In support of this sequence of events, intracranial recordings in humans (previously reviewed) during an item-method DF task showed increased signal (TBF>TBR) of the entorhinal cortex, which was subsequently followed by decreased signal in the anterior hippocampus (Ludowig et al., 2010).
Conversely, in the case where memories have received training and thus have undergone longer encoding (TNT task), future presentation of the cue most likely initiates a default state of retrieval. To reduce the likelihood of retrieval from occurring, learning that a specific memory is irrelevant or should be avoided may be necessary. This process could involve associating the composite memory with reduced reward through DA influence. Indicative of learning, initial inhibitory attempts in the TNT task invoke increased hippocampal activity (Depue et al., 2007), and have been shown to be unsuccessful (Depue et al., 2006; Hanslmayr et al., 2009; Butler & James, 2010). These latter findings indicate that both cue and target are retrieved, and thus the composite memory (cue and target) may undergo learning and become associated with irrelevancy or avoidance. Once learning occurs, presentation of the cue may signal the MFG to reduce the output of the hippocampus/MTL, perhaps by increasing activity of GABA-mediated interneurons in the presubiculum or subiculum. Additionally, because the composite memory is now associated with reduced reward and thus DA through learning, it may exhibit decreased spike-timing synchronization and oscillatory behavior important for retrieval. fMRI evidence supporting this shows reduced activity (T>NT) in the posterior hippocampus/MTL (Butler & James, 2010), as well as reduced activation in the cortices supporting memory representation (Anderson et al., 2004; Depue et al., 2007; 2010) during attempts to inhibit retrieval. If reactivation is reduced, then the appropriate information about the content of specific memories would not be transmitted, either directly (medial pathway) or indirectly (throught the basal ganglia), to the PFC for WM processes.
In sum, using the previous anatomical details and functional account, a model of inhibition of retrieval involves alterations of encoding processes before stimuli have been well established, or alternatively learning to avoid the retrieval process when stimuli have been encoded well. This perhaps occurs by cognitive/inhibitory control, reflected as increased activation of MFG excitatory connections on GABA-mediated interneurons of the entorhinal cortex and presubiculum (i.e., lateral and medial pathways). Both pathways and their associated processes may be facilitated by the influence of DA input from the hippocampal-prefrontal pathway, which has been show to affect theta/gamma oscillatory behavior by reducing spike-timing synchronization. Reduced information or oscillations in the output of the hippocampus/MTL would likely involve reductions in (i) pattern completion, (ii) activity in posterior cortex, and (iii) information gated through the basal ganglia to be accessed or maintained by WM, all of which are likely to affect the amount of information recalled at any given moment.
6. Inhibitory Modulation of Memory Retrieval within a Broader Perspective of Controlled Inhibition
To further support a model of inhibition of retrieval and place it in a broader perspective, it is important to consider more well-established mechanisms of controlled inhibition over other domains. Literature examining both response inhibition and emotion regulation (Aron et al., 2004; Phan et al., 2005; Eisenberger et al., 2003; Ochsner et al., 2002) indicates a role of the rMFG and rIFG in inhibitory modulation. A clear neuroanatomical mechanism involved in the inhibition of motor responses has been mapped out extensively (Aron et al., 2004; Aron & Polldrack, 2006). The major circuit involves excitatory pathways from the rIFG to the subthalamic nucleus of the basal ganglia. This pathway excites GABA- mediated interneurons in the globus pallidus that induce inhibition by interrupting important signals that transmit information to the thalamus and subsequently to the motor cortex, as illustrated in Figure 4. This has been considered a global stop mechanism of motor movement, and has been demonstrated in tasks involving the stopping of prepotent motor responses (e.g., Stop-signal, Go/No-go; Aron et al., 2004; Aron & Polldrack, 2006).
Figure 4.
Inhibitory circuitry involved in response inhibition (orange), fear extinction and emotion regulation (green), and a proposed circuit for inhibition of memory retrieval (red), (AMY = amygdala, Central = central nucleus, ICMs = intercalated cell masses, HIP = hippocampus, BG = basal ganglia, STN = subthalamic nucleus, GP = globus pallidus).
A similarly structured circuit in the amygdala is known to mediate fear extinction, and may provide modulation of physiological responses during emotion regulation, re-appraisal, and feelings of social rejection (Phan et al., 2005; Eisenberger et al., 2003; Ochsner et al., 2002). Excitatory connections from ventromedial PFC (VMPFC) synapse on GABA-mediated interneurons (intercalated cell masses) located between the basal and central nuclei of the amygdala (Milad & Quirk, 2002; Pare et al., 2004). These control afferents, with possible higher level cognitive control mediated via the LPFC to VMPFC, exert inhibition over the central nucleus’s output to decrease physiological responses to fearful stimuli (Royer et al., 1999; Pare et al., 2004) via modulation of the autonomic nervous system and somato-sensory cortices (Fig. 4).
I propose that, akin to inhibitory modulation of motor responses and emotional reactivity, inhibition of retrieval is a cognitive process that arises through cortically-driven control over sub-cortical and older (non-neo) cortical regions (e.g., basal ganglia, amygdala, hippocampus). These mechanisms may be viewed as accompanying more recent phylogenetic cortical regions that express control over older phylogentic brain circuitries. Specifically, response inhibition involves neocortical control over motor behavior (LPFC -> basal ganglia) to alter complex approach and withdrawal locomotion. Emotion regulation involves the cortical control of the fear circuit that is expressed in fear conditioning or extinction of the fear response (LPFC/OFC-> amygdala) to alter physiological arousal. And similarly, inhibiting retrieval appears to involve neocortical control over memory (LPFC->hippocampal complex) to alter encoding and retrieval processes associated with relevant and irrelevant memories. All of these interactions appear to exemplify the relative cognitive flexibility that humans have evolved to modulate the functions of lower-order brain circuitries.
7. Conclusion
In sum, a model of inhibition of retrieval incorporating Reactivation Inhibition as the main mechanism is supported by the current behavioral, neuroimaging, and comparative neurology/anatomy literature. The neural mechanisms likely involve MFG – hippocampal/MTL interaction through the lateral and medial pathways with influence of DA input by the hippocampal-prefrontal pathway to modulate encoding and retrieval processes. This is evidenced most clearly in fMRI findings, with corroborating results from ERP studies, all of which indicate reduced hippocampal/MTL activation, reduced activation of the cortices supporting memory representation, reduced parietal and right frontal old/new effects and increased N2 effects.
While constant modulation of retrieval processes in order to select appropriate behavior for ongoing goal activity most likely occurs, it largely takes little effort to perform. However, this process is much more apparent and effortful when the information is highly salient or disturbing. Therefore, understanding these processes may be informative of dysfunction in clinical populations who exhibit problems with memory and thought (i.e., PTSD, OCD, GAD, ASD, depression). More broadly, inhibition of retrieval may function similarly to response inhibition and emotion regulation, as cognitive processes that modulate lower-order brain circuitries to adjust the flow of information through general inhibitory gating, rather than specific inhibitory influence over individual representations. Although still speculative, by fully outlining the proposed mechanisms by which inhibition of retrieval occurs, numerous opportunities to test the model are apparent. Finally, understanding these mechanisms not only informs the nature of inhibiting retrieval, but the control of memory in general, as well as inhibitory processes in the brain that contribute to flexible behavior.
Highlights.
Behavioral and neuroimaging studies of inhibition of memory retrieval were reviewed. > Results were used to differentiate three current hypotheses in control over memory. > Interaction between lateral prefrontal cortex and hippocampus were consistently seen. > A neuroanatomical model is developed to illustrate the possible mechanisms involved. > Inhibition over memory is discussed in relation to inhibition across other domains.
Acknowledgments
Appreciation is given to the anonymous reviewer for very thoughtful insights and additions to the review. This works was supported by NIMH grant # P50 – 079485.
Footnotes
The current concern in this paper is with cognitively controlled attempts to inhibit retrieval. As such, it places other non-cognitive processes that may affect memory outside the focus of this discussion. Specifically, because the tasks used to examine inhibition of retrieval are sometimes termed memory or thought suppression, occasionally a proposed relationship between the tasks and the controversial unconscious defense mechanism of Freudian repression that renders an individual unaware of a traumatic memory that may subsequently be “recovered” has been suggested (Anderson & Green 2001; Anderson et al. 2004; Erdelyi 2006). Although it has been debated that conscious inhibition of retrieval may be the counterpart to an unconscious process of repression, and that the mechanisms of how each are employed may share some common aspects (Erdelyi, 2006), data are not currently available to answer these questions convincingly, and so they lie outside the scope of this discussion.
Although the direct inhibition hypothesis has been characterized as specific inhibition of a single memory representation, which is referenced by multiple articles by Anderson and colleagues, it is noteworthy that their recent discussions and future work may alternatively view inhibition of retrieval as a more general inhibitory mechanism (Anderson, personal communication, 2011).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Alexander GE, Crutcher MD, DeLong MR. Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, ‘prefrontal’ and ‘limbic’ functions. Progressive Brain Research. 1990;85:119–146. [PubMed] [Google Scholar]
- Anderson MC. Rethinking interference theory: executive control and the mechanisms of forgetting. Journal of Memory and Language. 2003;49:415–445. [Google Scholar]
- Anderson MC. Active forgetting: Evidence for functional inhibition as a source of memory failure. Journal of Aggression, Maltreatment, & Trauma. 2001;4:185–210. [Google Scholar]
- Anderson MC, Bjork RA, Bjork EL. Remembering can cause forgetting: retrieval dynamics in long-term memory. Journal of Experimental Psychology: Learning Memory and Cognition. 1994;20:1063–87. doi: 10.1037//0278-7393.20.5.1063. [DOI] [PubMed] [Google Scholar]
- Anderson MC, Green C. Suppressing unwanted memories by executive control. Nature. 2001;410:366–369. doi: 10.1038/35066572. [DOI] [PubMed] [Google Scholar]
- Anderson MC, Ochsner KN, Kuhl B. Neural systems underlying the suppression of unwanted memories. Science. 2004;303:232–235. doi: 10.1126/science.1089504. [DOI] [PubMed] [Google Scholar]
- Aron AR, Poldrack RA. Cortical and subcortical contributions to stop signal response inhibition: Role of the subthalamic nucleus. Journal of Neuroscience. 2006;26:2424–2433. doi: 10.1523/JNEUROSCI.4682-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aron AR, Robbins TW, Poldrack RA. Inhibition and the right inferior frontal cortex. Trends in Cognitive Sciences. 2004;8:170–177. doi: 10.1016/j.tics.2004.02.010. [DOI] [PubMed] [Google Scholar]
- Benchenane K, Tiesinga PH, Battaglia FP. Oscillations in the prefrontal cortex: A gateway to memory and attention. Current Opinion in Neurobiology. 2011;21:1–11. doi: 10.1016/j.conb.2011.01.004. [DOI] [PubMed] [Google Scholar]
- Bergstrom Z, de Fockert J, Richardson-Klavehn A. ERP and behavioral evidence for direct suppression of unwanted memories. Neuroimage. 2009;48:726–737. doi: 10.1016/j.neuroimage.2009.06.051. [DOI] [PubMed] [Google Scholar]
- Bergstrom Z, Velmans M, de Fockert J, Richardson-Klavehn A. ERP evidence for successful voluntary avoidance of conscious recollection. Brain Research. 2007;1151:119–133. doi: 10.1016/j.brainres.2007.03.014. [DOI] [PubMed] [Google Scholar]
- Bjork RA. Positive forgetting: the noninterference of items intentionally forgotten. Journal of Verbal Learning and Verbal Behavior. 1970;9:255–268. [Google Scholar]
- Bjork RA. Retrieval inhibition as an adaptive mechanism in human memory. In: Roediger HL III, Craik FIM, editors. Varieties of memory and consciousness: Essays in honour of Endel Tulving. Hillsdale, NJ: Erlbaum; 1989. [Google Scholar]
- Bjork EL, Bjork RA. Intentional forgetting can increase, not decrease, residual influences of to-be-forgotten information. Journal of Experimental Psychology. 2003;29:524–531. doi: 10.1037/0278-7393.29.4.524. [DOI] [PubMed] [Google Scholar]
- Bjork EL, Bjork RA, Anderson MC. Varieties of goal-directed forgetting. In: Golding JM, MacLeod CM, editors. Intentional forgetting: Interdisciplinary approaches. Mahwah, NJ: Erlbaum; 1998. [Google Scholar]
- Bjork RA, Geiselman RE. Constituent processes in the differentiation of items in memory. Journal of Experimental Psychology: Human, Learning, & Memory. 1978;4:347–361. [Google Scholar]
- Bjork RA, Woodward AE. Directed forgetting of individuals words in free recall. Journal of Experimental Psychology. 1973;99:22–27. [Google Scholar]
- Block RA. Effects of instructions to forget in short- term memory. Journal of Experimental Psychology. 1971;89:1–9. [Google Scholar]
- Bruce D, Papay JP. Primacy effects in single trial free recall. Journal of Verbal Learning and Verbal Behavior. 1970;9:473–486. [Google Scholar]
- Bulevich JB, Roediger HL, III, Balota DA, Butler AC. Failures to find suppression of episodic memories in the think/no-think paradigm. Memory and Cognition. 2006;34:1569–1577. doi: 10.3758/bf03195920. [DOI] [PubMed] [Google Scholar]
- Butler AJ, James KH. The neural correlates of attempting to suppress negative versus neutral memories. Cognitive, Affective, & Behavioral Neuroscience. 2010;10:182–194. doi: 10.3758/CABN.10.2.182. [DOI] [PubMed] [Google Scholar]
- Canto CB, Wouterlood FG, Witter MP. What Does the Anatomical Organization of the Entorhinal Cortex Tell Us? Neural Plasticity. 2008 doi: 10.1155/2008/381243. 381243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caruana DA, Sorge RE, Stewart J, Chapman CA. Dopamine Has Bidirectional Effects on Synaptic Responses to Cortical Inputs in Layer II of the Lateral Entorhinal Cortex. Journal of Physiol. 2006;96:3006–3015. doi: 10.1152/jn.00572.2006. [DOI] [PubMed] [Google Scholar]
- Chrobak J, Spates MJ, Stackman RW, Walsh TJ. Hemicholinium3 prevents the working memory impairments and the cholinergic hypofunction induced by ethylcholine aziridinium ion (AF64A) Brain Research. 1989;504:269–275. doi: 10.1016/0006-8993(89)91367-x. [DOI] [PubMed] [Google Scholar]
- Depue BE, Banich MT. Increased inhibition and enhancement of memory retrieval are associated with reduced hippocampal volume. Hippocampus. 2011 doi: 10.1002/hipo.20952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depue BE, Burgess GC, Willcutt EG, Ruzic L, Banich MT. Inhibitory control of memory retrieval and motor processing associated with the right lateral prefrontal cortex: Evidence from deficits in individuals with ADHD. Neuropsychologia. 2010;48:3909–3917. doi: 10.1016/j.neuropsychologia.2010.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depue BE, Banich MT, Curran T. Prefrontal regions orchestrates the suppression of memory via a two phase process. Science. 2007;317:215–219. doi: 10.1126/science.1139560. [DOI] [PubMed] [Google Scholar]
- Depue BE, Banich MT, Curran T. Suppression of emotional and non-emotional content in memory: Effects of repetition on cognitive control. Psychological Science. 2006;17:441–447. doi: 10.1111/j.1467-9280.2006.01725.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberger NI, Lieberman MD, Williams KD. Does rejection hurt? An fMRI study of social exclusion. Science. 2003;302:290–292. doi: 10.1126/science.1089134. [DOI] [PubMed] [Google Scholar]
- Erdelyi MH. The unified theory of repression. Behavioral and Brain Sciences. 2006;29:499–551. doi: 10.1017/S0140525X06009113. [DOI] [PubMed] [Google Scholar]
- Erickson K, Drevets W, Schulkin J. Glucocorticoid regulation of diverse cognitive functions in normal and pathological emotional states. Neuroscience and Biobehavioral Reviews. 2003;27:233–246. doi: 10.1016/s0149-7634(03)00033-2. [DOI] [PubMed] [Google Scholar]
- Fell J, Axmacher N. The role of phase synchronization in memory processes. Nature. 2011;12:105–118. doi: 10.1038/nrn2979. [DOI] [PubMed] [Google Scholar]
- Frank MJ, Loughry B, O’Reilly RC. Interactions between the frontal cortex and basal ganglia in working memory: A computational model. Cognitive, Affective, and Behavioral Neuroscience. 2001;1:137–160. doi: 10.3758/cabn.1.2.137. [DOI] [PubMed] [Google Scholar]
- Geiselman RE, Bagheri B. Repetition effects in directed forgetting: Evidence for retrieval inhibition. Memory and Cognition. 1985;13:57–62. doi: 10.3758/bf03198444. [DOI] [PubMed] [Google Scholar]
- Geiselman RE, Bjork RA, Fishman DL. Disrupted retrieval in directed forgetting: A link with posthypnotic amnesia. Journal of Experimental Psychology: General. 1983;112:58–72. doi: 10.1037//0096-3445.112.1.58. [DOI] [PubMed] [Google Scholar]
- Givens BS, Olton DS. Cholinergic and GABAergic modulation of medial septal area: Effect on working memory. Behavioral Neuroscience. 1990;104:849–855. doi: 10.1037//0735-7044.104.6.849. [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS, Selemon LD, Schwartz ML. Dual pathways connecting the dorsolateral prefrontal cortex with the hippocampal formation and parahippocampal cortex in the rhesus monkey. Neuroscience. 1984;12:719–743. doi: 10.1016/0306-4522(84)90166-0. [DOI] [PubMed] [Google Scholar]
- Hanslmayr S, Leipold P, Bäuml KH. Anticipation boosts forgetting of voluntarily suppressed memories. Memory. 2010;18:252–257. doi: 10.1080/09658210903476548. [DOI] [PubMed] [Google Scholar]
- Hassabis D, Chu C, Rees G, Weiskopf N, Molyneux PD, Maguire EA. Decoding Neuronal Ensembles in the Human Hippocampus. Current Biology. 2009;19:546–554. doi: 10.1016/j.cub.2009.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasselmo ME, Bodelón C, Wyble BP. A proposed function for hippocampal theta rhythm, separate phases of encoding and retrieval enhance reversal of prior learning. Neural Comput. 2002;14:793–817. doi: 10.1162/089976602317318965. [DOI] [PubMed] [Google Scholar]
- Hazy TE, Frank MJ, O’Reilly RC. Banishing the homunculus: Making working memory work. Neuroscience. 2006;139:105–118. doi: 10.1016/j.neuroscience.2005.04.067. [DOI] [PubMed] [Google Scholar]
- Hertel PT, Calcaterra G. Intentional forgetting benefits from thought substitution. Psychonomic Bulletin & Review. 2005;12:484–489. doi: 10.3758/bf03193792. [DOI] [PubMed] [Google Scholar]
- Jonides J, Lewis RL, Nee DE, Lustig CA, Berman MG, Moore KS. The mind and brain of short-term memory. Annu Rev Psychol. 2008;59:193–224. doi: 10.1146/annurev.psych.59.103006.093615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konishi S, Watanabe T, Jimura K, Chikazoe J, Hirose S, Kimura HM, Miyashita M. Role for presupplementary motor area in inhibition of cognitive set interference. Journal of Cognitive Neuroscience. 2001;23:737–745. doi: 10.1162/jocn.2010.21480. [DOI] [PubMed] [Google Scholar]
- Levy BJ, Anderson MC. Inhibitory processes and the control of memory retrieval. Trends in Cognitive Neuroscience. 2002;6:299–305. doi: 10.1016/s1364-6613(02)01923-x. [DOI] [PubMed] [Google Scholar]
- Lambert AJ, Good KS, Kirk IJ. Testing the repression hypothesis: Effects of emotional valence on memory suppression in the Think – No think task. Consciousness and Cognition. 2010;19:281–293. doi: 10.1016/j.concog.2009.09.004. [DOI] [PubMed] [Google Scholar]
- Laroche S, Davis S, Jay TM. Plasticity at hippocampal to prefrontal cortex synapses: Dual roles in working memory and consolidation. Hippocampus. 2000;10:438–446. doi: 10.1002/1098-1063(2000)10:4<438::AID-HIPO10>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Lehman EB, Bovasso M. Development of intentional forgetting in children. In: Howe ML, Pasnak R, editors. Emerging themes in cognitive development Vol 1: Foundations. New York: Springer-Verlag; 1993. [Google Scholar]
- Leopold DA, Wilke M. Neuroimaging: Seeing the trees for the forest. Current Biology. 2005;15:R766–8. doi: 10.1016/j.cub.2005.08.055. [DOI] [PubMed] [Google Scholar]
- Ludowig E, Möller J, Bien CG, Münte TF, Elger CE, Rosburg T. Active suppression in the mediotemporal lobe during directed forgetting. Neurobiology of Learning and Memory. 2010;93:352–361. doi: 10.1016/j.nlm.2009.12.001. [DOI] [PubMed] [Google Scholar]
- MacLeod CM. The item and list methods in directed forgetting: Test differences and the role of demand characteristics. Psych Bull and Rev. 1999;6:123–129. doi: 10.3758/bf03210819. [DOI] [PubMed] [Google Scholar]
- Malkinson R. Cognitive-behavioral therapy of grief: A review and application. Research on Social Work Practice. 2001;11:671–698. [Google Scholar]
- Manns ID, Mainville L, Jones BE. Evidence for glutamate, in addition to acetylcholine and GABA, neurotransmitter synthesis in basal forebrain neurons projecting to the entorhinal cortex. Neuroscience. 2001;107:249–263. doi: 10.1016/s0306-4522(01)00302-5. [DOI] [PubMed] [Google Scholar]
- McNab F, Klingberg T. Prefrontal cortex and basal ganglia control access to working memory. Nature Neuroscience. 2008;11:103–107. doi: 10.1038/nn2024. [DOI] [PubMed] [Google Scholar]
- Mecklinger A, Parra M, Waldhauser GT. ERP correlates of intentional forgetting. Brain Research. 2009;1255:132–147. doi: 10.1016/j.brainres.2008.11.073. [DOI] [PubMed] [Google Scholar]
- Milad MR, Quirk GJ. Neurons in medial prefrontal cortex signal memory for fear extinction. Nature. 2002;240:70–74. doi: 10.1038/nature01138. [DOI] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annual Rev Neurosci. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- Mitchell JP, Heatherton TF, Kelley WM, Wyland CL, Wegner DM, Macrae CM. Separating sustained from transient aspects of cognitive control during thought suppression. Psychological Science. 2007;18:292–297. doi: 10.1111/j.1467-9280.2007.01891.x. [DOI] [PubMed] [Google Scholar]
- Morris RE, Pandya DN, Petrides M. Fiber system linking the mid-dorsolateral frontal cortex with the retrosplenial/presubicular region in the rhesus monkey. The Journal of Comparative Neurology. 1999;407:183–192. doi: 10.1002/(sici)1096-9861(19990503)407:2<183::aid-cne3>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- Morris R, Petrides M, Pandya DN. Hodological fronto-hippocampal interaction: re-discovering the cingulum bundle. Soc Neurosci Abstr. 1995;21:2. [Google Scholar]
- Munakata Y, Herd S, Chattam C, Depue BE, Banich MT, O’Reilly RC. A unified theory of inhibition. Trends in Cognitive Science. doi: 10.1016/j.tics.2011.07.011. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muther WS. Erasure or partitioning in short-term memory. Psychonomic Science. 1965;3:429–430. [Google Scholar]
- Nørby S, Lange M, Larsen A. Forgetting to forget: On the duration of voluntary suppression of neutral and emotional memories. Acta Psychologica. 2010;133:73–80. doi: 10.1016/j.actpsy.2009.10.002. [DOI] [PubMed] [Google Scholar]
- Nowicka A, Marchewka A, Jednoróg K, Tacikowski P, Brechmann A. Forgetting of emotional information is hard: An fMRI study of directed forgetting. Cerebral Cortex. 2011;21:539–549. doi: 10.1093/cercor/bhq117. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Bunge SA, Gross JJ, Gabrieli JDE. Rethinking feelings: An fMRI study of the cognitive regulation of emotion. Journal of Cognitive Neuroscience. 2002;14:1215–1299. doi: 10.1162/089892902760807212. [DOI] [PubMed] [Google Scholar]
- O’Reilly RC, Frank MJ. Making working memory work: A computational model of learning in the frontal cortex and basal ganglia. Neural Computation. 2006;18:283–328. doi: 10.1162/089976606775093909. [DOI] [PubMed] [Google Scholar]
- O’Reilly RC, Norman KA. Hippocampal and neocortical contributions to memory: Advances in the complementary learning systems framework. Trends in Cognitive Sciences. 2002;6:505–510. doi: 10.1016/s1364-6613(02)02005-3. [DOI] [PubMed] [Google Scholar]
- O’Reilly RC, McClelland JL. Hippocampal conjunctive encoding, storage, and recall: avoiding a tradeoff. Hippocampus. 1994;4:661–682. doi: 10.1002/hipo.450040605. [DOI] [PubMed] [Google Scholar]
- Otmakhova NA, Lisman J. Dopamine selectively inhibits the direct cortical pathway to the CA1 hippocampal region. The Journal of Neuroscience. 1999;19 doi: 10.1523/JNEUROSCI.19-04-01437.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paré D, Quirk GJ, LeDoux JE. New vistas on amygdala networks in conditioned fear. Journal of Neurophysiol. 2004;92:1–9. doi: 10.1152/jn.00153.2004. [DOI] [PubMed] [Google Scholar]
- Paz-Alonso PM, Ghetti S, Matlen BJ, Anderson MC, Bunge SA. Memory suppression is an active process that improves over childhood. Frontiers in Human Neuroscience. 2010;3:24. doi: 10.3389/neuro.09.024.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrides M, Milner B. Deficits on subject-ordered tasks after frontal and temporallobe lesions in man. Neuropsychologia. 1982;20:249–262. doi: 10.1016/0028-3932(82)90100-2. [DOI] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Moore GJ, Uhde TW, Tancer ME. Neural substrates for voluntary suppression of negative affect: A functional magnetic resonance imaging study. Biological Psychiatry. 2005;57:210–219. doi: 10.1016/j.biopsych.2004.10.030. [DOI] [PubMed] [Google Scholar]
- Pinto A, Fuentes C, Pare D. Feedforward inhibition regulates perirhinal transmission of neocortical inputs to the entorhinal cortex: Ultrastructural study in guinea pigs. The Journal of Comparative Neurology. 2006;495:722–734. doi: 10.1002/cne.20905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolls ET. Functions of neuronal networks in the hippocampus and neocortex in memory. In: Byrne J, et al., editors. Neural Models of Plasticity: Experimental and Theoretical Approaches. Academic Press; 1989. [Google Scholar]
- Royer S, Martina M, Paré D. An inhibitory interface gates impulse traffic between the input and output stations of the amygdala. Journal of Neurosci. 1999;19:10575–10583. doi: 10.1523/JNEUROSCI.19-23-10575.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rugg MD, Curran T. Event-related potentials and recognition memory. Trends in Cognitive Science. 2007;6:251–257. doi: 10.1016/j.tics.2007.04.004. [DOI] [PubMed] [Google Scholar]
- Salame P, Danion JM. Inhibition of inappropriate responses is preserved in the think-no-think and impaired in the random number generation tasks in schizophrenia. Journal of the International Neuropsychological Society. 2007;13:277–287. doi: 10.1017/S1355617707070300. [DOI] [PubMed] [Google Scholar]
- Sewards TV, Sewards MA. Input and output stations of the entorhinal cortex: superficial vs. deep layers or lateral vs. medial divisions? Brain Research Reviews. 2003;42:243–251. doi: 10.1016/s0165-0173(03)00175-9. [DOI] [PubMed] [Google Scholar]
- Shirvalkar PR, Rapp PR, Shapiro ML. Bidirectional changes to hippocampal theta–gamma comodulation predict memory for recent spatial episodes. PNAS. 2010;107:7054–7059. doi: 10.1073/pnas.0911184107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swanson LW. A direct projection from Ammon’s horn to prefrontal cortex in the rat. Brain Research. 1981;217:150–154. doi: 10.1016/0006-8993(81)90192-x. [DOI] [PubMed] [Google Scholar]
- Thierry A-M, Gioanni Y, Dégénétais E, Glowinski J. Hippocampo-Prefrontal Cortex Pathway: Anatomical and Electrophysiological Characteristics. Hippocampus. 2000;10:411–419. doi: 10.1002/1098-1063(2000)10:4<411::AID-HIPO7>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Tomlinson TD, Huber DE, Rieth CA, Davelaar EJ. An interference account of cue-independent forgetting in the no-think paradigm. PNAS. 2009;106:15588–15593. doi: 10.1073/pnas.0813370106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA. Meta-Analysis of the Functional Neuroanatomy of Single-Word Reading: Method and Validation. NeuroImage. 2002;16:765–780. doi: 10.1006/nimg.2002.1131. [DOI] [PubMed] [Google Scholar]
- Ullsperger M, Mecklinger A, Müller U. An electrophysiological test of directed forgetting: the role of retrieval inhibition. J Cogn Neurosci. 2000;12:924–940. doi: 10.1162/08989290051137477. [DOI] [PubMed] [Google Scholar]
- Van Hooff JC, Whitaker TA, Ford RM. Directed forgetting in direct and indirect tests of memory: Seeking evidence of retrieval inhibition using electrophysiological measures. Brain and Cognition. 2009;71:153–164. doi: 10.1016/j.bandc.2009.05.001. [DOI] [PubMed] [Google Scholar]
- Van Haeften T, Wouterlood TG, Jorritsma-Byham B, Witter MP. GABAergic presubicular projections to the medial entorhinal cortex of the rat. The Journal of Neuroscience. 1997;17:862–874. doi: 10.1523/JNEUROSCI.17-02-00862.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wegner DM. Ironic processes of mental control. Psychological Review. 1994;101:34–52. doi: 10.1037/0033-295x.101.1.34. [DOI] [PubMed] [Google Scholar]
- Wegner DM, Schneider DJ, Carter S, White T. Paradoxical effects of thought suppression. Journal of Personality and Social Psychology. 1987;53:5–13. doi: 10.1037//0022-3514.53.1.5. [DOI] [PubMed] [Google Scholar]
- Wegner DM, Erber R. The hyperaccessibility of suppressed thoughts. Journal of Personality and Social Psychology. 1992;63:903–912. [Google Scholar]
- Wegner DM. You can’t always think what you want: Problems in the suppression of unwanted thoughts. In: Zanna M, editor. Advances in experimental social psychology. San Diego, CA: Academic Press; 1992. [Google Scholar]
- Wegner DM. When the antidote is the poison: Ironic mental control processes. Psychological Science. 1997;8:148–150. [Google Scholar]
- Wegner DM, Wenzlaff RM. Mental control. In: Higgins ET, Kruglanski A, editors. Social psychology: Handbook of basic mechanisms and processes. Academic Press; 1996. [Google Scholar]
- Weiner B, Reed H. Effects of the instructional sets to remember and to forget on short-term retention: Studies of rehearsal control and retrieval inhibition (repression) Journal of Experimental Psychology. 1969;79:226–232. [Google Scholar]
- Wenzlaff RM, Wegner DM, Klein SB. The role of thought suppression in the bonding of thought and mood. Journal of Personality and Social Psychology. 1991;60:500–508. [Google Scholar]
- Wenzlaff RM, Wegner DM, Roper D. Depression and mental control: The resurgence of unwanted negative thoughts. Journal of Personality and Social Psychology. 1988;55:882–892. doi: 10.1037//0022-3514.55.6.882. [DOI] [PubMed] [Google Scholar]
- Wessel I, Wetzels S, Jelicic M, Merckelbach H. Dissociation and memory suppression: A comparison of high and low dissociative individuals performance on the Think–No Think task. Personality and Individual Differences. 2005;39:1461–1470. [Google Scholar]
- Wilson SP, Kipp S. The development of efficient inhibition: evidence from directed-forgetting tasks. Developmental Review. 1998;18:86–123. [Google Scholar]
- Wimber M, Bauml KH, Bergstrom Z, Markopoulos G, Heinze HJ, Richardson-Klavehn A. Neural markers of inhibition in human memory retrieval. The Journal of Neuroscience. 2008;10:3419–13427. doi: 10.1523/JNEUROSCI.1916-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wixted JT. The psychology and neuroscience of forgetting. Ann Rev Psychol. 2004;55:235–69. doi: 10.1146/annurev.psych.55.090902.141555. [DOI] [PubMed] [Google Scholar]
- Wyland CL, Kelley WM, Macrae CN, Gordon HL, Heatherton TF. Neural correlates of thought suppression. Neuropsychologia. 2003;41:1863–1867. doi: 10.1016/j.neuropsychologia.2003.08.001. [DOI] [PubMed] [Google Scholar]
- Wylie GR, Foxe JJ, Taylor TL. Forgetting as an active process: An fMRI investigation of item-method-directed forgetting. Cerebral Cortex. 2007;18:670–82. doi: 10.1093/cercor/bhm101. [DOI] [PubMed] [Google Scholar]