

Abstract
Decaying root systems of harvested trees can be a significant component of belowground carbon storage, especially in intensively managed forests where harvest occurs repeatedly in relatively short rotations. Based on destructive sampling of root systems of harvested loblolly pine trees, we estimated that root systems contained about 32% (17.2 Mg ha−1) at the time of harvest, and about 13% (6.1 Mg ha−1) of the soil organic carbon 10 years later. Based on the published roundwood output data, we estimated belowground biomass at the time of harvest for loblolly-shortleaf pine forests harvested between 1995 and 2005 in South Carolina. We then calculated C that remained in the decomposing root systems in 2005 using the decay function developed for loblolly pine. Our calculations indicate that the amount of C stored in decaying roots of loblolly-shortleaf pine forests harvested between 1995 and 2005 in South Carolina was 7.1 Tg. Using a simple extrapolation method, we estimated 331.8 Tg C stored in the decomposing roots due to timber harvest from 1995 to 2005 in the conterminous USA. To fully account for the C stored in the decomposing roots of the US forests, future studies need (1) to quantify decay rates of coarse roots for major tree species in different regions, and (2) to develop a methodology that can determine C stock in decomposing roots resulting from natural mortality.
Keywords: Carbon cycle, Carbon pool, Loblolly pine, Root biomass, Piedmont, South Carolina
Introduction
Forest ecosystem carbon storage is commonly inventoried by several separate pools: live biomass (tree and understory vegetation), standing dead trees, down dead wood, forest floor, and soil organic carbon or forest soils. Until recently (Petersson and Melin 2010), these estimates have not explicitly accounted for C stored in decaying root systems, although down dead wood has included stumps and roots of harvested trees in some studies (e.g., Woodbury et al. 2007; Smith and Heath 2008). Because of the laborious nature of root sampling (e.g., Kapeluck and Van Lear 1995), the quantity and fate of C stored in decaying coarse roots are not well studied. Decaying roots of harvested trees, before they are fully decomposed and incorporated into the soil, are part of the belowground detritus carbon pool and should be explicitly accounted for. Belowground C estimates that ignore decaying root systems may grossly underestimate the size of the belowground C pool because C concentrations in decaying root systems can be many times greater than those of the soil matrix (e.g., Van Lear et al. 2001).
In the southern United States, pine plantations have increased from ~0.7 million ha in 1952 to ~11 million ha in 1999 and are projected to rise to ~18 million ha by 2040 (Wear and Greis 2002). Most pine plantations in the South are harvested on rotations of <50 years, and rotation lengths are commonly <25 years for the 3 million ha of intensively managed plantations. In South Carolina alone, 115 million cubic meters of loblolly-shortleaf pine timber was harvested between 1995 and 2005. Thus, there is a huge inventory of root systems of loblolly-shortleaf pine trees in various stages of decay in forest soils as a direct result of timber harvest. The same is likely true for forests in other regions that are subject to similar forest management. Until this transitional C storage, i.e., between intact tree roots and soil organic matter (SOM), is quantified or estimated, global C budget models will remain inaccurate.
To demonstrate that decaying roots of harvested trees represent a significant component of belowground C at a stand scale, we estimated various components of belowground C in a recently harvested loblolly pine stand in the Clemson University Experimental Forest. To determine the size of the C pool in the decaying root systems of harvested trees at a regional scale, we analyzed timber harvesting and forest inventory data published for South Carolina between 1995 and 2005. Our objectives were (1) to show that root systems of harvested loblolly pine trees planted on abandoned agricultural land in the Piedmont of South Carolina ~50 years ago contain significant quantities of C, and (2) to quantify soil C storage in decaying roots of loblolly-shortleaf pine stands harvested between 1995 and 2005 in South Carolina.
Methods
Field sampling was conducted at the Clemson Experimental Forest (CEF) in Clemson, South Carolina, USA. We sampled a mature (48 years old) loblolly pine plantation (Fig. 1), a 16-year-old loblolly pine stand that naturally regenerated when a part of the mature stand was harvested, and a recently harvested pine stand. The three sampled stands had similar topography (upper- or mid-slope position, 2–15% slope, and southern exposure). Soils were formed in residuum granite, gneiss, and schist and have relatively thin sandy loam surface horizons underlain by firm red clay loam and clay. Poor agricultural practices before reforestation caused extensive erosion and formed deep gullies and ephemeral stream channels on these sites. Both the fertility and the organic matter contents are low.CEF has a humid temperate climate, with a high of 25°C in July, a low of 7°C in January, and an annual precipitation of 1,310 mm.
Fig. 1.

A mature loblolly pine stand growing in poorly eroded soils at CEF, Clemson, South Carolina, USA (Photo credit: G. Geoff Wang)
To determine soil organic carbon in the mineral soil, we collected composite mineral soil samples at 0–10-, 10–30-, and 30–50-cm depths from 18 randomly located points throughout the recently harvested stand. These sampling depths generally corresponded to the A, B, and C horizons. If a randomly located point fell on a tree stump or the core hit an intact object (e.g., root or rocks), then the sample location was offset one meter and re-cored. These composite samples, each consisting of four cores per depth per location, were analyzed for SOM using a Perkin Elmer Model 240C Elemental Analyzer. The extremely low and stable C concentrations in the 50–100 cm soil depth were extrapolated from regressions of C concentration versus soil depth using data from upper soil depths (Van Lear et al. 1995; Jobbagy and Jackson 2000).
Carbon stored in root systems and old root channels had been previously measured and reported by Van Lear et al. (1995) for the same four soil depths (0–10, 10–30, 30–50, and 50–100 cm) in the mature and 16-year-old stands. In brief, we determined root biomass using a combination of excavation, regression, and coring methods developed by Kapeluck and Van Lear (1995). Coarse-root biomass estimates were generated in four diameter classes (taproot, >5.0, 2.5–5.0, and 0.6–2.5 cm) by applying regressions to a 100% inventory of tree diameters in the mature stand. Taproot biomass distribution by depth was determined by dividing the volume of the average taproot (based on tree of average diameter) into sections corresponding to depth of soil sampling. Taproots of pine trees in this stand rarely penetrated more than 1 m into the C horizon of the Pacolet fine sandy loam. Fine root biomass (roots <0.6-cm diameter) was estimated from 60 randomly located soil cores (10-cm diameter) taken to a depth of 75 cm. Few fine roots grew below this depth, where soil strength generally exceeded 1.5 g cm−3. Root biomass and C were allocated to the same four soil depths (0–10, 10–30, 30–50, and 50–100 cm) using data on root distribution from soil profiles (Van Lear et al. 1995).To estimate the amount of C remaining in decaying root systems at different years since harvesting, we applied the following locally developed decay function (Van Lear et al. 1995):
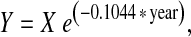 |
1 |
where Y, C remaining; X, initial quantity of C; e, base of natural logarithm 2.718; −0.1044, overall decay coefficient; and year, years since harvest.
The volume of soil to a depth of 1 m occupied by old root channels was estimated from the analyses of 60 1-m2 soil profiles randomly located within the site (McCollum 1992). Outlines of root channels and rocks and/or rock cavities were traced onto acetate sheets. In the laboratory, sketches were measured to determine their areas. We considered these soil profiles analogous to planar intersections employed to measure volume of forest fuels (Brown 1974), and we used these profile sketches to convert root channel cross-sectional areas into volumes.
We obtained loblolly-shortleaf pine roundwood output data from the biennial reports published in 1995 (Johnson et al. 1997), 1997 (Johnson and Bischoff 1999), 1999 (Johnson et al. 2002), 2001 (Johnson et al. 2004), 2003 (Johnson and Knight 2006), and 2005 (Johnson and Smith 2007). Based on the percent of total roundwood output that was considered as growing stock volume and growing stock use rate (Bentley and Johnson 2006; Bentley and Harper 2006), we converted roundwood output to harvested growing stock volume. The average of the two successive years was used for deriving the total growing stock volume for the non-reported years (i.e., 1996, 1998, 2000, 2002, and 2004). Because the area of timber harvest was not reported in the biennial reports, we assumed that growing stock volume of the harvested stands averaged 280 m3 ha−1 (Baker and Langdon 1990), based on which the area of timber harvest was estimated for each year.
We used forest inventory data to calculate fractional aboveground biomass (AGBf; except foliage) of loblolly-shortleaf pine for each county for the inventory years 1993, 2001, and 2005 based on a previously developed regression equation for loblolly-shortleaf pine (Hu and Wang 2008):
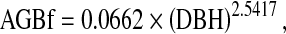 |
2 |
where AGBf, fractional aboveground biomass excluding foliage (kg); DBH, diameter at breast height (cm); and 0.0662 and 2.5417, regression parameters.
Foliage biomass was estimated using the constant foliage ratio method. Published studies (Ter-Mikaelian and Korzukhin 1997; Wagner and Ter-Mikaelian 1999) report that the proportion of foliage within the AGB averages 0.07 (with range from 0.01 to 0.14) for softwoods. Using this proportion and AGBf calculated from Eq. 2, we determined total AGB for loblolly-shortleaf pine for each county in 1993, 2001, and 2005.
Based on the timberland area and total growing stock volume of loblolly-shortleaf pine in each county from FIA data in the inventory years 1993 and 2001 and the AGB estimated above, we calculated a biomass expansion factor (BEF, defined as the ratio of AGB of all living trees with DBH ≥2.54 cm to growing stock volume for all trees with DBH ≥12.7 cm; Mg m−3) and growing stock volume per unit area (GSVD; m3 ha−1). Based on these calculations, BEF was regressed on GSVD using the inverse function:
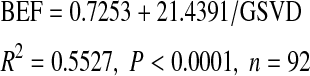 |
3 |
We validated this model using the 2003 inventory data. When the calculated BEF was used to convert growing stock volume to biomass, there was a very good agreement (R2 = 0.989, n = 46) with the biomass estimated from Eq. 2.
Using the average GSVD of 280 m3 ha−1, a BEF of 0.801868 was estimated from Eq. 3. This BEF was used to convert the total growing stock volume to the total AGB for the harvested loblolly-shortleaf pine forests from 1995 to 2005. The total AGB divided by the harvested area resulted in AGB density (AGBD), based on which belowground (coarse and fine roots) biomass density (BGBD) was calculated for harvested loblolly-shortleaf pine from 1995 to 2005 using the following regression equation developed for temperate forests by Cairns et al. (1997):
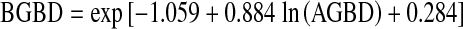 |
4 |
Belowground biomass at the time of harvest was then calculated by multiplying BGBD by the harvested area. We estimated remaining belowground biomass of harvested loblolly-shortleaf pine at any given year since harvesting using the negative exponential decay function developed for loblolly pine.
Results
Belowground C estimates were calculated for a recently harvested loblolly pine stand. Carbon content of the root system was 17.2 Mg ha−1 at time of harvest, 89% of which was in roots in the upper 50 cm of the soil profile (Table 1). Carbon sequestered in tree roots represented 32% of soil organic C at the time of harvest. An additional 5.5 Mg ha−1 C was contained in much older root channels, which should be a component of soil organic C. Based on the locally developed decay function (Eq. 1), 10.2, 6.1, and 1.3 Mg ha−1 C would remain in the decomposing root system 5, 10, and 25 years after harvest, respectively (Fig. 2). These quantities of C were equivalent to 19.0, 11.4, and 2.4% of the soil C (Table 1).
Table 1.
Belowground carbon contents of mineral soil, root systems, and old root channels
| Soil depth (cm) | Mineral soil carbon (Mg ha−1) |
Root system carbon (Mg ha−1) |
Old root channel carbon (Mg ha−1) |
Total carbon (Mg ha−1) |
|---|---|---|---|---|
| 0–10 | 13.4 | 6.7 | 2.1 | 22.2 |
| 10–30 | 14.5 | 5.8 | 1.9 | 22.2 |
| 30–50 | 9.1 | 2.8 | 0.9 | 12.8 |
| 50–100 | 11.1 | 1.9 | 0.6 | 13.6 |
| Total | 48.1 | 17.2 | 5.5 | 70.8 |
Carbon content of root systems is estimated at the time of harvest, representing the maximum amount of C in decomposing roots
Fig. 2.

Carbon storage in the decomposing root systems of a harvested loblolly pine stand at CEF, South Carolina, USA
Between 1995 and 2005, timber harvest from loblolly-shortleaf pine forests in South Carolina removed a total of 90.9 Tg AGB, with annual removal ranging from 7.5 to 10.6 Tg (Table 2). Belowground biomass (including coarse and fine roots) at the time of harvest ranged from 1.8 to 2.6 Tg between 1995 and 2005, with a total of 22.2 Tg. Belowground biomass accounted for ~20% of the total biomass.
Table 2.
Estimates of aboveground and belowground biomasses at the time of harvest and residual C in decomposing roots for the harvested loblolly-shortleaf pine forests in South Carolina
| Year | Harvested growing stock volume (m3) | Harvested area (ha) | Aboveground biomass (Tg) | Belowground Biomass (Tg) | Residual C (Tg) |
|---|---|---|---|---|---|
| 1995 | 10 304 570 | 36 802 | 8.2 | 2.0 | 0.3 |
| 1996 | 10 237 390 | 36 562 | 8.2 | 2.0 | 0.4 |
| 1997 | 10 170 200 | 36 322 | 8.1 | 2.0 | 0.4 |
| 1998 | 9 956 760 | 35 560 | 8.0 | 2.0 | 0.5 |
| 1999 | 9 743 030 | 34 806 | 7.8 | 1.9 | 0.5 |
| 2000 | 9 568 900 | 34 175 | 7.7 | 1.9 | 0.6 |
| 2001 | 9 386 710 | 33 524 | 7.5 | 1.8 | 0.6 |
| 2002 | 9 570 390 | 34 180 | 7.7 | 1.9 | 0.7 |
| 2003 | 9 754 120 | 34 846 | 7.8 | 1.9 | 0.8 |
| 2004 | 11 471 540 | 40 970 | 9.2 | 2.2 | 1.0 |
| 2005 | 13 208 600 | 47 174 | 10.6 | 2.6 | 1.3 |
| Total | 113 372 200 | 404 901 | 90.9 | 22.2 | 7.1 |
Biomass is converted to C by multiplying with a factor of 0.5
Assuming that the root systems of these harvested trees followed the same decay pattern characterized in the CEF (Eq. 1), residual C in the decomposing roots of trees harvested between 1995 and 2005 was calculated with reference to 2005 (i.e., 0 year of decomposition for trees harvested in 2005, and 10 years of decomposition for trees harvested in 1995). The decomposing roots of loblolly-shortleaf pine trees harvested between 1995 and 2005 accumulated a total belowground C of 7.1 Tg in South Carolina.
Discussion
During the growth of forests, both the above- and belowground biomasses act as “sinks” where C is sequestered for decades or even centuries (Jonsson and Wardle 2010). In southern pine forests, most of the belowground biomass is in coarse roots (Kapeluck and Van Lear 1995; Laiho and Finer 1996; Johnsen et al. 2001). Coarse-root and stump biomass has been found to increase with the age of the stand, constituting up to 90% of the total living root biomass (Laiho and Finer 1996; Ehman et al. 2002). Following harvest or natural tree mortality, root systems become a “source” of carbon as they decay in a negative exponential pattern over decades, releasing CO2 to the atmosphere. However, coarse roots persist after harvest (Johnsen et al. 2001; Ludovici et al. 2002), providing a longer term C storage mechanism than that provided by fine roots. Therefore, it is important to quantify coarse-root C in both live trees and dead, decaying root systems.
When studying the forest carbon cycle, root systems of the recently harvested trees are not considered a component of soil organic C, yet fully decomposed root systems are obviously a part of the soil. Therefore, decomposing root systems (i.e., root systems at the time of harvest and before fully incorporated into the soil) represent a transitional form of belowground C storage. Our study demonstrates that decomposing roots of harvested trees in managed forests contained a large quantity of belowground C at both stand and regional scales. Unless this portion of belowground C storage is explicitly accounted for in ecosystem models, our estimations of the C budget will remain inaccurate. With projected increases in harvested areas due to the expansion of pine plantations (Wear and Greis 2002) and/or decrease in rotation age due to intensive forest management practices (e.g., tree improvement, fertilization, etc.) (Wear and Greis 2002), C sequestered in decomposing root systems will also increase. On the other hand, any management practices that harvest biomass from stump and/or coarse roots would decrease the C storage by decomposing root systems.
This study indicates that C sequestered in tree roots at the time of harvest represented 32% of the soil organic C stored in heavily eroded soils of the upper Piedmont in South Carolina on which loblolly pine plantations were planted about 50 years earlier. After 10 years, decomposing roots of the harvested trees would still represent 13% of the soil organic C. This percentage, however, may be inflated somewhat because we only measured soil organic C to a depth of 1 m. The poor soils of our study sites may also contribute to the high percentage of C sequestered in decomposing roots. Trimble (1974) estimated that an average of 23 cm of top soil, including associated organic matter, was lost in this region between 1880 and 1930 due to poor agriculture practices. As a result, our study sites were left with low quantities of soil organic C. Although decomposing roots may sequester different percentages of soil C depending on soil conditions, they are an important component of the belowground C pool and should not be ignored in future C budget models.
Our previous studies have shown that loblolly pine trees growing in these poor soils develop large tapering taproots that extend vertically to a depth of about 1 m, and lateral roots project radially from the root collar area just below the soil surface (Kapeluck and Van Lear 1995). Complete root systems represented about 20% of total tree biomass, 60% of which was within 1 m of stump centers, and 90% of which occupied the top 50 cm of soil (Kapeluck and Van Lear 1995; Miller et al. 2006). Immediately after harvest, these root systems occupied <1% of soil volume to a depth of 1 m at the stand level. Stumps of harvested trees rapidly become covered with detritus and neither random nor systematic soil sampling is likely an adequate method to capture these root systems because of the small volume of soil they occupy. Therefore, similar methodology like the one developed in this study must be used to determine C stored in decomposing root systems of harvested trees.
In this study, we determined that C associated with old root channels was also a significant component of soil organic C. A previous study indicated that these old channels are of critical importance for the development of new roots for next generation of loblolly pine stands (Van Lear et al. 2001). Because these old root channels occupy a small soil volume, C determined through randomly located soil core sampling may not accurately capture this C component in the soil unless the sample size becomes very large. This sampling error may become significant, especially on poor sites like ours, and sampling methods that minimize this error should be developed and used in future C-accounting models.
Loblolly pine is the major timber species in the southern USA. To show the significance of decomposing roots as a belowground C pool at the regional scale, we calculated the quantity of C stored in decomposing roots for loblolly-shortleaf pine forests harvested between 1995 and 2005 in South Carolina. Our results indicate that C stored in decomposing roots due to harvest of loblolly-shortleaf pine forests amounted to 7.1 Tg in South Carolina. However, SC timberland only accounts for 5.7% of the total timberland in the 14 southern States (i.e., Alabama, Arkansas, Florida, Georgia, Kentucky, Louisiana, Mississippi, North Carolina, Oklahoma, South Carolina, Texas, Virginia, and West Virginia). If we assume that the amount of harvest is proportional to the size of timberland area in each state, then the total C stored in decomposing roots of harvested loblolly-shortleaf forests would be 124.6 Tg in the southern USA. If we take into account all types of forests, the figure would be much larger. Given timber harvest data are published periodically at county, state, and national levels, the method developed in our study can be easily applied to estimate C stored in decomposing roots of harvested trees if decay rates of coarse roots are determined for different species and different regions. However, information on decay rates of coarse roots is rather limited for major timber species in various regions of the USA, and more studies are needed.
Our method estimated C storage in decomposing root systems based on the published equations (Eqs. 1, 2, and 4), newly developed BEF equation (Eq. 3), and the published roundwood output data, each of which could introduce errors to the estimation. A complete assessment of the uncertainty of our estimation due to all sources of error would be preferable. However, such an assessment was not possible because no error estimation was given for the roundwood output data. Because roundwood output data did not report the size of harvest area, we derived harvest areas using an average stand volume of 280 m3 ha−1 reported in the literature (Baker and Langdon 1990). We also calculated BEF using this average stand volume, and the calculated BEF was used to convert the harvested growing stock volume to biomass. Using a constant volume for all the harvested stands may therefore introduce a source of error into our estimation. However, this error may not be significant since timber harvesting occurred in mature stands, thus supporting a relatively high volume. The BEF equation developed in the study was an inverse function, indicating that BEF approaches a stable value at a high stand volume associated with mature stands. Inverse functions have been commonly used to describe biomass–volume relationships in previous studies (e.g., Fang et al. 1998).
In a recent report, Adams et al. (2006) estimated timber harvest by the US region and ownership between 1950 and 2002. For the same period from 1995 to 2002, the timber harvest of loblolly-shortleaf pine forests in South Carolina only represented 3.84 and 2.14% of the timber harvest in the southern and the conterminous USA, respectively. Assuming these percentages remained the same for the period from 1995 to 2005, a simple extrapolation results in 184.9 and 331.8 Tg of C stored in decomposing roots due to timber harvest from 1995 to 2005 in the southern and the conterminous USA, respectively. Smith and Heath (2008) reported that the size of the forest soil organic C pool in the USA in 2005 was 15 544 Tg, and the 331.8 Tg C stored in decomposing roots due to timber harvest between 1995 and 2005 represent only about 2.1% of the forest SOC pool. This low percentage is not unexpected, given that timber harvest only occurred in a very small portion of the US forests during this period. For many of the US forests that are not open to timber harvesting, decomposing roots due to natural mortality were not accounted for by the current C budget models. The C stored in these decomposing roots, although difficult to estimate based on the method proposed in our study, would likely be substantial. According to Smith and Heath (2008), dead wood and litter in the US forests stored 7193 Tg C in 2005. If we assume that dead materials generated aboveground (dead wood and litter) and belowground (root system) decompose at the same rate, the proportion of live belowground and aboveground biomass would remain the same for dead belowground and aboveground biomass (approximately 20%). The C stock in decomposing roots in the US forests in 2005 would be 1439 Tg (20% of dead wood and litter), which is 9.2% of the soil organic C pool. In a recent study, Petersson and Melin (2010) estimated the carbon pool of stump systems in Sweden based on a traditional National Forest Inventory. By matching mortality (either naturally died or harvested) data at the individual tree level within each permanent sample plots, their methods accounted for carbon storage in decaying roots due to timber harvest as well as natural mortality, and their estimates suggested that approximately five times more C was stored in stump systems than in dead wood aboveground when estimated over the period of 2003–2007. In 2005, dead wood stored 2568 Tg C in the US forests (Smith and Heath 2008), which would amount to 12 840 Tg C stored in stump systems in 2005 if we assume the same ratio of stump system C to aboveground dead wood C found by Petersson and Melin (2010). Clearly, future studies need to account for the C stock in the decomposing roots due to natural mortality in the US forests.
In summary, we have demonstrated that C stored in decomposing roots is a substantial component of SOC pool. In plantations established on highly eroded soils in the Upper Piedmont of South Carolina, C stored in decomposing roots could account for 32 and 13% of the total SOC pool at the time of harvest and 10 year after, respectively. At a regional scale, C stored in decomposing roots was estimated to be 7.09 Tg for loblolly-shortleaf pine forests harvested between 1995 and 2005 in South Carolina. Using a simple extrapolation method, we estimated 331.30 Tg C stored in the decomposing roots due to timber harvest from 1995 to 2005 in the conterminous USA. To fully account for the C stored in decomposing roots of the US forests, future studies need (1) to quantify decay rates of coarse roots for major tree species in different regions, and (2) to develop a methodology that can determine C stock in decomposing roots resulted from natural mortality.
Acknowledgments
This paper is Technical Contribution No. 5928 of the Clemson University Experimental Station. We thank Ben Knapp for his editorial suggestions.
Biographies
G. Geoff Wang
is a Professor in the School of Agricultural, Forest and Environmental Science at Clemson University.
David H. Van Lear
is a Professor Emeritus in the School of Agricultural, Forest and Environmental Science at Clemson University.
Huifeng Hu
is a Ph.D. candidate in the School of Agricultural, Forest and Environmental Science at Clemson University.
Peter R. Kapeluck
was a research specialist (now retired) in the School of Agricultural, Forest and Environmental Science at Clemson University.
References
- Adams, D.M., R.W. Haynes, and A.J. Daigneault. 2006. Estimated timber harvest by U.S. region and ownership, 1950–2002. USDA Forest Service, General Technical Report. PNW-GTR-659, 64 pp.
- Baker, J.B., and O.G. Langdon. 1990. Pinus taeda L., loblolly pine. In Silvics of North America, vol. 1, conifers, ed. R.M. Burns and B.H. Honkala, 497–512. Agriculture Handbook 654. Washington, DC: USDA Forest Service.
- Bentley, J.W., and R.A. Harper. 2006. Georgia harvest and utilization study, 2004. USDA Forest Service, RB-SRS-117, 25 pp.
- Bentley, J.W., and T.G. Johnson. 2006. North Carolina harvest and utilization study, 2002. USDA Forest Service, RB-SRS-109, 23 pp.
- Brown, J.K. 1974. Handbook for inventorying downed woody material. USDA Forest Service, General Technical Report, GTR-INT-16, 34 pp.
- Cairns MA, Brown S, Helmer EH, Baumgardner GA. Root biomass allocation in the world’s upland forests. Oecologia. 1997;111:1–11. doi: 10.1007/s004420050201. [DOI] [PubMed] [Google Scholar]
- Ehman JL, Schmid HP, Grimmond CSB, Randolph JC, Hanson PJ, Wayson CA, Cropley FD. An initial intercomparison of micrometeorological and ecological inventory estimates of carbon exchange in a mid-latitude deciduous forest. Global Change Biology. 2002;8:575–589. doi: 10.1046/j.1365-2486.2002.00492.x. [DOI] [Google Scholar]
- Fang JY, Wang GG, Liu GH, Xu SL. Forest biomass of China: An estimate based on the biomass–volume relationship. Ecological Applications. 1998;8:1084–1091. [Google Scholar]
- Hu HF, Wang GG. Changes in forest biomass carbon storage in the South Carolina Piedmont between 1936 and 2005. Forest and Ecology Management. 2008;255:1400–1408. doi: 10.1016/j.foreco.2007.10.064. [DOI] [Google Scholar]
- Jobbagy EG, Jackson RB. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications. 2000;10:423–436. doi: 10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2. [DOI] [Google Scholar]
- Johnsen KH, Wear JD, Oren R, Teskey RO, Sanchez F, Will R, Butnor J, Markewitz D, Richter D, Rials T, Allen HL, Seiler J, Ellsworth D, Maier C, Katul G, Dougherty PM. Meeting global policy commitments: Carbon sequestration and southern pine forests. Journal of Forestry. 2001;99:14–21. [Google Scholar]
- Johnson, T.G., and P.S. Bischoff. 1999. South Carolina’s timber industry—an assessment of timber product output and use, 1997. USDA Forest Service, RB-SRS-46, 32 pp.
- Johnson, T.G., R.A. Harper, and M.J. Bozzo. 2002. South Carolina’s timber industry—an assessment of timber product output and use, 1999. USDA Forest Service, RB-SRS-70, 42 pp.
- Johnson, T.G., R.A. Harper, and M.J. Bozzo. 2004. South Carolina’s timber industry—an assessment of timber product output and use, 2001. USDA Forest Service, RB-SRS-89, 33 pp.
- Johnson, T.G., A. Jenkins, D.P. Stratton, and P.S. Bischoff. 1997. South Carolina’s timber industry—an assessment of timber product output and use, 1995. USDA Forest Service, RB-SRS-16, 32 pp.
- Johnson, T.G., and M. Knight. 2006. South Carolina’s timber industry—an assessment of timber product output and use, 2003. USDA Forest Service, RB-SRS-106, 47 pp.
- Johnson, T.G., and N. Smith. 2007. South Carolina’s timber industry—an assessment of timber product output and use, 2005. USDA Forest Service, RB-SRS-121, 28 pp.
- Jonsson M, Wardle DA. Structural equation modelling reveals plant-community drivers of carbon storage in boreal forest ecosystems. Biology Letters. 2010;6:116–119. doi: 10.1098/rsbl.2009.0613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapeluck PR, Lear DH. A technique for estimating below-stump biomass of mature loblolly pine plantations. Canadian Journal of Forest Research. 1995;25:361–365. doi: 10.1139/x95-039. [DOI] [Google Scholar]
- Laiho R, Finer L. Changes in root biomass after water-level drawdown on pine mires in southern Finland. Scandinavian Journal of Forest Research. 1996;11:251–260. doi: 10.1080/02827589609382934. [DOI] [Google Scholar]
- Ludovici KH, Zarnoch SJ, Richter DD. Modeling in situ pine root decomposition using data from a 60-year chronosequence. Canadian Journal of Forest Research. 2002;32:1675–1684. doi: 10.1139/x02-073. [DOI] [Google Scholar]
- McCollum, M.M. 1992. Density and distribution of loblolly pine (Pinus taeda L.) roots along an environmental gradient. MS Thesis, Clemson University, Clemson, 62 pp.
- Miller AT, Allen HL, Maier CA. Quantifying the coarse-root biomass of intensively managed loblolly pine plantations. Canadian Journal of Forest Research. 2006;36:12–22. doi: 10.1139/x05-229. [DOI] [Google Scholar]
- Petersson H, Melin Y. Estimating the biomass and carbon pool of stump systems at a national scale. Forest Ecology and Management. 2010;260:466–471. doi: 10.1016/j.foreco.2010.05.002. [DOI] [Google Scholar]
- Smith, J.E., and L.S. Heath. 2008. Carbon stocks and stock changes in U.S. forests. In U.S. Department of Agriculture. U.S. Agriculture and Forestry Greenhouse Gas Inventory: 1990–2005. Office of the Chief Economist. Technical Bulletin No. 1921, 65–80, C-1-C-7.
- Ter-Mikaelian MT, Korzukhin MD. Biomass equations for sixty-five North American tree species. Forest Ecology and Management. 1997;97:1–24. doi: 10.1016/S0378-1127(97)00019-4. [DOI] [Google Scholar]
- Trimble, S.W. 1974. Man-induced soil erosion on the Southern Piedmont, 1700–1970. Ankeny, IA: Soil Conservation Society of America, 80 pp.
- Van Lear, D.H., P.R. Kapeluck, and M.M. Parker. 1995. Distribution of carbon in a Piedmont soil as affected by loblolly pine management. In Carbon forms and functions in forest soils, ed. W.W. McFee and J.M. Kelly, 489–502. Madison, WI: Soil Science Society of America.
- Van Lear, D.H., P.R. Kapeluck, and W.D. Carroll. 2001. Productivity of loblolly pine as affected by decomposing root systems. In Forest soils and ecosystem sustainability, ed. J.R. Boyle and R.F. Powers, 435–444. New York, NY: Elsevier.
- Wagner RG, Ter-Mikaelian MT. Comparison of biomass component equations for four species of northern coniferous tree seedlings. Annals of Forest Science. 1999;56:193–199. doi: 10.1051/forest:19990301. [DOI] [Google Scholar]
- Wear, D.N., and J.G. Greis. 2002. Southern forest resource assessment—summary report. USDA Forest Service, GTR-SRS-53, 103 pp.
- Woodbury PB, Smith JE, Heath LS. Carbon sequestration in U.S. forest sector from 1990 to 2010. Forest Ecology and Management. 2007;241:14–27. doi: 10.1016/j.foreco.2006.12.008. [DOI] [Google Scholar]