

Abstract
Our previous research demonstrated that psychopathy-related fear deficits involve abnormalities in attention that undermine sensitivity to peripheral information. In the present study, we specified this attention-mediated abnormality in a new sample of 87 prisoners assessed with Hare's Psychopathy Checklist-Revised (Hare, 2003). We measured fear-potentiated startle (FPS) under four conditions that crossed attentional focus (threat vs. alternative) with early versus late presentation of threat cues. The psychopathic deficit in FPS was apparent only in the early-alternative-focus condition, in which threat cues were presented after the alternative goal-directed focus was established. Furthermore, psychopathy interacted with working memory capacity in the late-alternative-focus condition, which suggests that individuals high in psychopathy and working memory capacity were able to maintain a set-related alternative focus that reduced FPS. The results not only provide new evidence that attention moderates the fearlessness of psychopathic individuals, but also implicate an early attention bottleneck as a proximal mechanism for deficient response modulation in psychopathy.
Keywords: psychopathy, fear, attention, higher-order cognitive processes, multilocus selection
Attentional processes play an important role in the fear deficit of psychopathic individuals. According to the response modulation theory, psychopathic individuals appear insensitive to fear-related stimuli because they fail to reallocate attention to secondary affective information while engaged in goal-directed behavior (MacCoon, Wallace, & Newman, 2004; Patterson & Newman, 1993). This difficulty balancing the demands of goal-directed processing and secondary information processing creates a bias whereby psychopathic individuals are less responsive to affective information unless it is a central aspect of their goal-directed focus of attention.
We previously used an instructed fear-conditioning task to demonstrate that the fearlessness of psychopathic offenders is moderated by their focus of attention (Newman, Curtin, Bertsch, & Baskin-Sommers, 2010). Fear-potentiated startle (FPS) was measured as participants categorized colored letter stimuli under three conditions. One condition required participants to respond on the basis of the threat-relevant aspect of the stimuli (i.e., the color that predicted electric shocks). Two other conditions required participants to respond on the basis of an alternative, threat-irrelevant aspect of the stimuli (i.e., the case of the letter or the match or mismatch in a two-back task). In the alternative-focus conditions, the threat-relevant information was outside the primary focus of goal-directed behavior. Psychopathy scores (as measured by the Psychopathy Checklist-Revised, PCL-R; Hare, 2003) were significantly inversely related to FPS under conditions that required participants to focus on a threat-irrelevant dimension of stimuli. In contrast, psychopathy scores were unrelated to FPS in the condition that focused attention on the threat-relevant dimension. These results are consistent with the proposal that psychopathy involves abnormalities in attention that undermine sensitivity to emotion-related cues that normally modulate goal-directed behavior.
Although this research provided some of the strongest evidence to date that the fear deficit of psychopathic individuals is moderated by attention, the study did not specify the attentional mechanism underlying this effect. Goal-directed behavior requires attention to focus on relevant stimuli and ignore potential distractors. However, the mechanism for such goal-relevant control may vary depending on the demands of a specific situation, and it may involve diverse neural and cognitive systems. A useful framework for understanding the different types of attentional selection emanates from cognitive neuroscience research on the locus of attentional selection (Driver, 2001; Knudsen, 2007; Luck & Hillyard, 1999). Within this framework, attentional selection and attentional limitations may occur as a function of filtering prior to stimulus identification (early stage) or as a function of engagement of higher-order cognitive processes after stimulus identification (later stage). Depending on task demands, attentional selection and attentional limitations may also occur at multiple loci of selection (multilocus selection).
Early stages of selective attention occur as a function of a bottleneck that, once established, blocks the processing of secondary information that is not goal relevant (Driver, 2001). Although this bottleneck is most commonly associated with perceptual load (Lavie, Hirst, de Fockert, & Viding, 2004), there is also evidence that preperceptual filtering may be based on features such as spatial location or visual properties available prior to stimulus identification (Luck & Hillyard, 1999). Such early attentional influences can affect neural activity in the visual cortex (Kastner & Ungerleider, 2000), filter the processing of sensory information (Hillyard, Vogel, & Luck, 1998), and prevent the perception of secondary information (Lavie et al., 2004). Thus, for psychopathic individuals, once the distinction between primary and secondary information is established, the bottleneck filter may limit the processing of secondary information, such as salient threat stimuli.
Selective attention may also operate at a later, postperceptual stage (e.g., Luck & Hillyard, 1999). At some point between the occurrence of a stimulus and its effect on behavior, stimulus-driven and goal-relevant information compete for representation, selection, and control (Desimone & Duncan, 1995). At this later stage, selection occurs as a function of higher-order cognitive processes, such as memory and response selection, which signal the importance of specific information (Deutsch & Deutsch, 1963; Lavie et al., 2004). Thus, it is also possible that psychopathic individuals initially perceive and identify both primary and secondary information but are particularly adept at using higher-order processes to resolve the competition between goal-relevant and secondary demands on attention.
Although these early- and later-stage influences on selective attention occur at different points in the processing stream and involve activation of different brain regions (Hillyard & Anllo-Vento, 1998; Knudsen, 2007; Vogel, Woodman, & Luck, 2005), the processes underlying these influences may also overlap and combine to influence information processing (Luck & Hillyard, 1995). That is, selective attention may act at multiple stages of processing (Yantis & Johnston, 1990), with the ultimate effect on information processing being influenced by diverse situational and individual difference variables (multilocus selection; Awh, Vogel, & Oh, 2006; Lavie et al., 2004). Of particular relevance to this research is the notion that the early attention bottleneck may not always be triggered in an automatic, bottom-up way, but at times may be contingent on whether information and task attributes match a person's task-relevant set (e.g., attributes based on task instructions; Corbetta, Miezin, Dobmeye, Shulman, & Petersen, 1991; Folk, Remington, & Johnston, 1992). In addition, cognitive processes such as working memory may affect the on-line representation of set-relevant information (Awh et al., 2006; Haenny & Schiller, 1988). Therefore, a person's ability to perform this type of multilocus selection may be influenced by individual differences in working memory capacity (Fukuda & Vogel, 2009). Within a multilocus model, it is possible that the psychopathy-related attentional bias relates fundamentally to an early attention bottleneck, but that in contexts that require the application of a set (i.e., a specific goal-directed focus), working memory capacity is important for maintaining that bottleneck and precluding the processing of secondary information. Overall, the distinction between these stages of attentional selection provides a valuable framework for specifying the attention-related deficits of psychopathic individuals.
The primary aim of the present study was to specify the attentional abnormalities that undermine the processing of secondary affective stimuli in psychopathy and its associated factors. To this end, we modified the task we previously used (Newman et al., 2010) so that the threat information (color) and threat-irrelevant information (letter) were temporally separated (Mitchell, Richell, Leonard, & Blair, 2006). Specifically, we examined FPS in a new sample of incarcerated offenders under four experimental conditions that crossed threat versus alternative (i.e., threat-irrelevant) focus of attention with early versus late presentation of the threat-relevant cues. This manipulation yielded four conditions: early alternative focus, late alternative focus, late threat focus, and early threat focus. Of particular importance for testing our hypothesis is the fact that the experimental manipulation allowed us to examine FPS under two alternative-focus conditions. The first engaged the alternative focus of attention at an early stage, prior to the presentation of threat-relevant information (i.e., the early-alternative-focus condition). The second presented threat-relevant information prior to establishing the alternative focus of attention, thereby increasing demands for higher-order processes to select and maintain a set-congruent focus (i.e., the late-alternative-focus condition).
The following three hypotheses were developed to specify the attentional processes that moderate emotion processing in psychopathic individuals. First, to the extent that their deficit in response modulation involves an early attention bottleneck, psychopathic offenders should display deficient FPS, particularly when attention (i.e., goal-directed behavior) is engaged prior to presentation of the threat-relevant information (early-alternative-focus condition). Second, to the extent that their insensitivity to threat-related stimuli is inherently a reflection of a later stage of selection, psychopathic offenders' FPS deficit should be especially apparent under circumstances that necessarily call for higher-order processing (i.e., when the threat-relevant information is presented prior to the goal-relevant information; late-alternative-focus condition). Third, psychopathy-related differences in FPS may be moderated by working memory capacity, particularly in the late-alternative-focus condition, which requires the maintenance of an attentional set in the absence of set-relevant information to support the early attention bottleneck (i.e., multilocus selection). Because some researchers advocate parsing psychopathy into two components (Factor 1 and Factor 2; Patrick, 2007), we also examined the association between the two major psychopathy factors and FPS in this experimental design.
Method
Participants
Participants were inmates from a maximum-security prison in Wisconsin who did not participate in our previous study but met the same inclusion criteria (Newman et al., 2010). Because there is little evidence that psychopathy-related laboratory correlates generalize across race and gender, our laboratory disaggregates these samples and reports sample-specific psychopathy effects in separate studies (Baskin-Sommers, Newman, Sathasivam, & Curtin, 2010). In the present sample, participants were European American, male, and age 45 or younger; scored 70 or better on the Shipley Institute of Living Scale intelligence estimate (Zachary, 1986); had no record of schizophrenia, bipolar disorder, or psychosis; and were not using psychotropic medications (Cleckley, 1976; Hare, Harpur, & Hakstian, 1990; Hart & Hare, 1989). Ten participants were excluded because there was a baseline artifact or no response on more than 20% of the trials (Blumenthal et al., 2005). In addition, 2 outliers (Studentized residuals with Bonferroni-corrected p values < .05) were excluded from analyses. The final sample consisted of 87 inmates.
Psychopathy was assessed using the PCL-R (Hare, 2003). Information for the ratings was derived from a semistructured interview and a file review. In this study, for 13 participants randomly selected during the interview process for reliability ratings, the interrater reliabilities for PCL-R total scores, PCL-R Factor 1 (interpersonal and affective traits), and PCL-R Factor 2 (lifestyle and antisocial traits) were .97, .92, and .95, respectively.
To assess working memory, we administered a computerized version of the Digits Backward subtest of the Wechsler Adult Intelligence Scale (Wechsler, 1997). This test involves presenting a sequence of digits at a rate of one digit per second. The sequence starts with two digits and then increases by one digit every two trials. Participants are instructed to repeat the digits in reverse order. The assessment stops when the participant makes two consecutive errors or achieves two correct trials during the final sequences of eight digits. One point was given for each correct trial, and points were summed to calculate the raw score.
Instructed fear-conditioning task
Presentation of all stimuli and measurement of behavioral responses was controlled by DMDX software (Forster & Forster, 2003). The instructed fear-conditioning task consisted of four conditions, with 80 trials per condition. We crossed focus of attention (threat focus or alternative focus) with the timing of this attentional focus (early onset or late onset) to yield the following conditions: early threat focus, late threat focus, early alternative focus, and late alternative focus. In the threat-focus conditions, participants had to focus on the color of a box that predicted shock administration. In the alternative-focus conditions, participants had to focus on whether a letter stimulus was capitalized (i.e., a threat-irrelevant aspect of the trial). The order of conditions was counterbalanced across participants but did not interact significantly with the effects reported.
Every trial in all conditions started with a fixation cross lasting 200 ms. Then, participants saw two stimuli presented sequentially. These stimuli consisted of a box (colored red or green) and a letter (an uppercase N or a lowercase n). Their order of appearance varied with the condition. The first stimulus appeared alone at 200 ms, and then the second stimulus appeared concurrently with the first at 400 ms. Following the offset of the stimuli, a blank screen appeared. At 1,800 ms, a descriptive word appeared on the screen, prompting participants to respond on the basis of either the first or the second stimulus, according to the condition. In the threat-focus conditions, the word “Red” or “Green” was presented, and participants pressed one of two buttons to indicate whether the word described the color of the box during that trial. In the alternative-focus conditions, the word “Upper” or “Lower” appeared, and participants pressed one of two buttons to indicate whether the word described the case of the letter during that trial. The response prompts lasted for 800 ms and were followed by an intertrial interval of 1,000 ms. The time between the onset of the first stimulus on successive trials was 3,600 ms. Figure 1 summarizes trial timing and provides an example of a trial in each condition.
Fig. 1.
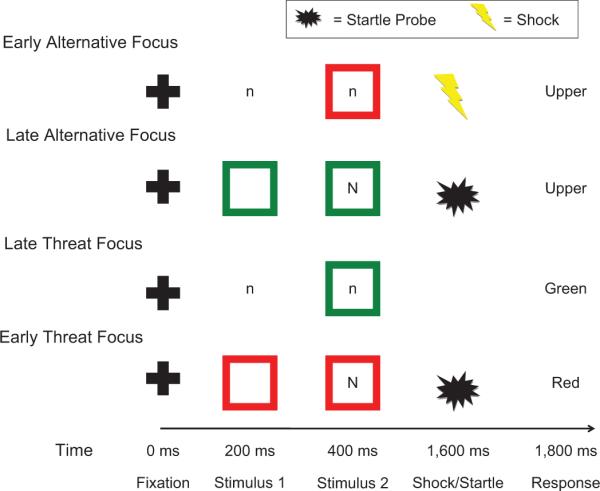
Trial structure in the four conditions used in the experiment. Every trial began with a fixation cross lasting 200 ms, after which participants saw two stimuli: a box (colored red or green) and a letter (an uppercase N or a lowercase n). The order of these two stimuli varied with condition. The first stimulus appeared alone at 200 ms, and then the second stimulus appeared concurrently with the first at 400 ms. In all four conditions, electric shocks were administered after some red boxes but never after green boxes. Following the offset of the stimuli, a blank screen appeared. White-noise startle probes were presented at 1,600 ms into the trial (i.e., during the blank screen) to measure fear-potentiated startle. At 1,800 ms, a descriptive word related to the color of the box or the case of the letter (i.e., “Green” or “Red” for threat-focus blocks; “Upper” or “Lower” for alternative-focus blocks) appeared on the screen. Participants had to indicate whether the word matched (or mismatched) the relevant feature presented during that trial.
In all conditions, participants were told that electric shocks could be administered following presentation of a red box (threat trial), but that shocks would never occur following presentation of a green box (no-threat trial). An electric shock was administered for 200 ms to two adjacent fingers on each participant's nondominant hand on 15% of threat trials in each condition. These shocks were delivered at 1,600 ms into the trial. A total of 24 shocks (6 per condition) were administered to each participant, and the intensity of shocks was calibrated to participants' subjective tolerance. The case of the letter stimulus (N or n) was unrelated to the administration of electric shocks.
For all conditions, the timing of the focus of attention (early or late) was orthogonally manipulated to examine the effects of early onset versus late onset of the attentional focus. This was accomplished by presenting the task-relevant stimulus either first or second in the trial sequence. For example, in the early-threat-focus condition, the colored box was presented first, followed by the letter stimulus.
Elicitation and measurement of the startle response
In 64 of the 320 trials, a startle-eliciting noise probe was presented at 1,600 ms into the trial. The probes were equally distributed across threat and no-threat trials in all four conditions so that each participant received 16 probes per condition. Probes were also equally distributed across uppercase-letter and lowercase-letter trials. These probes lasted for 50 ms and consisted of a 102-dB white-noise burst with a near-instantaneous rise time. Probes were separated by a minimum of 13 s and never occurred on the same trial as shock administration. Startle eyeblink electromyographic activity was sampled at 2000 Hz with a bandpass filter (30–500 Hz; 24 dB/octave roll-off) from electrodes placed on the orbicularis oculi muscle under the right eye according to published guidelines (Blumenthal et al., 2005). Off-line processing included epoching (−50 ms to 250 ms surrounding the probe), rectification and smoothing (30-Hz low-pass filter following rectification), and baseline correction. Startle-blink magnitude was scored as the peak response between 20 ms and 120 ms after probe onset. Fear response to threat cues was indexed by FPS, which was calculated by subtracting blink-response magnitude to probes following green (no threat) boxes from blink-response magnitude to probes following red (threat) boxes.
Results
Psychopathy total scores
FPS was analyzed in a general linear model (GLM) with condition as a within-subjects categorical factor and psychopathy total score (mean centered and standardized) as a between-subjects quantitative factor. See Table 1 for GLM point estimates for raw startle and FPS in the four conditions, separately for low and high levels of psychopathy.
Table 1.
General Linear Model Point Estimates for Startle Responses in the Four Conditions of the Experiment
| Early alternative focus |
Late alternative focus |
Late threat focus |
Early threat focus |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Psychopathy | No threat | Threat | FPS | No threat | Threat | FPS | No threat | Threat | FPS | No threat | Threat | FPS |
| Low psychopathy | 49.27 (8.15) | 78.93 (11.89) | 29.66 (6.68) | 48.66 (8.58) | 71.06 (11.00) | 22.40 (5.17) | 41.89 (8.16) | 64.90 (10.76) | 23.01 (6.03) | 56.75 (8.47] | 72.84 (11.13) | 16.09 (5.08) |
| High psychopathy | 46.87 (8.15) | 50.80 (11.89) | 3.93 (6.68) | 39.73 (8.58) | 51.58 (11.00) | 11.85 (5.17] | 44.72 (8.16) | 59.91 (10.76) | 15.19 (6.03) | 33.82 (8.47] | 51.94 (11.13) | 18.12 (5.08) |
| Condition mean | 48.07 (4.48) | 64.87 (6.61) | 16.80 (3.78) | 44.19 (4.72) | 61.32 (6.08) | 17.13 (2.86) | 43.31 (4.84) | 62.41 (5.19) | 19.10 (3.32) | 45.29 (4.73) | 62.39 (6.16) | 17.10 (2.79) |
Note: Fear-potentiated startle (FPS) was calculated by subtracting blink-response magnitude to probes following green (no threat) boxes from blink-response magnitude to probes following red (threat) boxes in each of the four conditions. Psychopathy scores were obtained using the Psychopathy Checklist-Revised (PCL-R; Hare, 2003). Because we analyzed psychopathy continuously instead of using an extreme-groups design, the values presented are point estimates (i.e., estimated using regression analyses) for low and high psychopathy points (1.5 SD below and above the sample mean PCL-R total score, respectively) on the distribution. Numbers in parentheses are standard errors.
The effect of condition on FPS was not significant (p = .86), indicating that FPS was comparable across all four conditions. Similarly, the effect of psychopathy was not significant (p = .18), indicating that psychopathy total scores were not consistently related to FPS across conditions.
As predicted, the relationship between psychopathy and FPS was moderated by condition (i.e., Psychopathy × Condition interaction), F(3, 255) = 2.971, p = .032, ηp2 = .03. This significant interaction was decomposed using three orthogonal (Helmert) interaction contrasts designed to identify which conditions selectively revealed inverse psychopathy-FPS relationships. The first interaction contrast indicated that psychopathy's relationship with FPS in the early-alternative-focus condition differed significantly from psychopathy's relationship with FPS in all other conditions,1F(1, 85) = 7.18, p = .009, ηp2 = .082 (Fig. 2). Follow-up simple-effects tests indicated that psychopathy was significantly inversely related to FPS in the early-alternative-focus condition (b = −8.59, p = .023), but not in the remaining conditions (b = −1.82, p = .45).
Fig. 2.
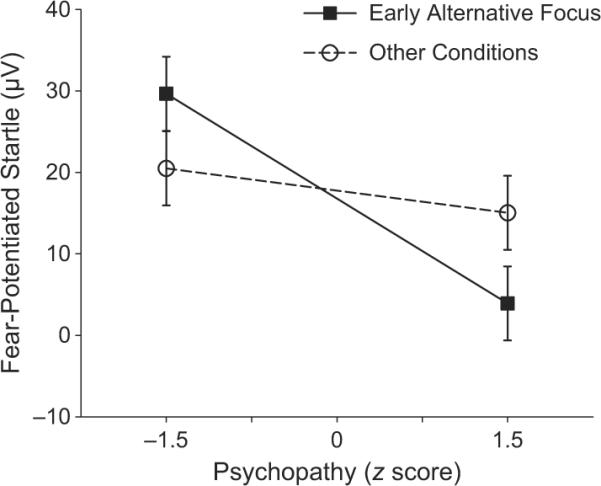
Fear-potentiated startle as a function of psychopathy (±1.5 SD from the mean) and condition (early-alternative-focus condition and the average of the three other conditions). Raw psychopathy scores, estimated from the Psychopathy Checklist-Revised (Hare, 2003), were mean-centered and standardized (z scores). Error bars represent ±1 SE.
The interaction contrast comparing psychopathy-FPS relationships in the late-alterative-focus condition with psychopathy-FPS relationships in the two threat-focus conditions was not significant (p = .32),3 and the contrast comparing the early-threat-focus condition with the late-threat-focus condition was also not significant (p = .33). These findings indicate that the psychopathy-FPS relationships were roughly comparable in the late-alternative-focus and two threat-focus conditions. Overall, this pattern of results demonstrated that increasing psychopathy scores were associated with decreasing fear responses only when attention was actively engaged in an alternative (threat-irrelevant) focus at an early stage (i.e., prior to presentation of threat-relevant information).
Psychopathy total scores and working memory (Digits Backward)
As a second step in this hierarchical analysis, Digits Backward scores were added to the GLM to determine whether working memory capacity plays a role in the ability to maintain an attention bottleneck (i.e., multilocus of selection). Digits Backward scores were mean-centered and standardized.
The relationship between psychopathy, condition, and FPS was moderated by Digits Backward score, F(3, 249) = 3.18, p = .029, ηp2 = .037. This significant omnibus interaction was decomposed using three orthogonal interaction contrasts (described previously). Digits Backward score did not significantly moderate the first interaction contrast (early alternative focus compared with other conditions; p = .24). However, Digits Backward score did significantly moderate the second interaction contrast (late alternative focus compared with the two threat-focus conditions), F(1, 83) = 9.02, p = .004, ηp2 = .098 (Fig. 3). Follow-up tests indicated that psychopathy and Digits Backward score significantly interacted to predict FPS in the late-alternative-focus condition (b = −6.15, p = .04), but not in the threat-focus conditions (b = 1.58, p = .57). Finally, Digits Backward score did not significantly moderate the remaining interaction contrast comparing the two threat-focus conditions (p = .48).
Fig. 3.

Fear-potentiated startle (FPS) as a function of psychopathy (±1.5 SD from the mean) and working memory (low vs. high). Results are shown for the late-alternative-focus condition (left panel) and the two threat-focus conditions (right panel). Raw psychopathy scores, estimated from the Psychopathy Checklist-Revised (Hare, 2003), were mean-centered and standardized (z scores). Scores for low and high working memory (±1.5 SD from the mean, respectively) were measured with the Digits Backward subtest of the Wechsler Adult Intelligence Scale (Wechsler, 1997). Error bars represent ±1 SE.
PCL-R–factor scores
Secondary analyses were conducted with psychopathy-factor scores to determine whether the psychopathy effect observed in the early-alternative-focus condition and the Psychopathy × Digits Backward effect observed in the late-alternative-focus condition would hold for Factor 1 and Factor 2, in addition to PCL-R total score. We repeated all of our analyses for Factors 1 and 2 and replicated the results for both factors. In other words, both Factor 1 and Factor 2 interacted significantly with the early-alternative-focus contrast (p = .009 and p = .05, respectively). Digits Backward score also moderated both the Factor 1 and Factor 2 late-alternative-focus contrast (p = .02 and p = .005, respectively). In addition, these analyses revealed a significant main effect for Factor 1, F(1, 85) = 5.54, p = .021, ηp2 = .06; this effect reflects lower FPS when the Factor 1 score is higher across all conditions. However, as noted, this main effect of Factor 1 was qualified by the significant early-alternative-focus contrast demonstrating that this FPS deficit was primarily due to reduced FPS in the early-alternative-focus condition.
Discussion
In three key ways, the present study provides powerful support for the role of attention in moderating the fear deficits of psychopathic individuals. First, despite the use of a new sample of offenders and a different experimental paradigm, the results replicate the central findings of our previous study (Newman et al., 2010). As in that study, psychopathy-related deficits in FPS were found only in conditions in which the threat information was peripheral to the primary task. Also replicating our previous results, the current findings showed no evidence for a psychopathy-related deficit in FPS when the threat-relevant dimension of the experimental stimuli was goal relevant (i.e., primary focus of attention). Beyond the conceptual replication, the design of the present study allowed us to address potential limitations of our previous study and more precisely explore the attentional mechanism.
One limitation we cited in our previous research (Newman et al., 2010) was that our experimental conditions resulted in significantly different levels of FPS. This was potentially problematic because we could not rule out the hypothesis that psychopathy-related differences in FPS are found more readily in weak fear conditions (i.e., those eliciting weaker FPS; cf. Lissek, Pine, & Grillon, 2006) than in conditions involving direct or potent threats. That is, the main effect of condition in our previous study may have determined the psychopathy-related differences in FPS. In contrast to our previous study, the present study showed no main effect of condition, indicating that FPS was roughly comparable regardless of the attentional focus or temporal onset of the threat cues. Thus, the present study clarifies that psychopathy-related differences in FPS are a function of condition-related effects on attention rather than a condition-related artifact involving the overall strength of FPS.
A second focus of this study was to examine the locus of attentional selection influencing these attention-related fear deficits. Our analyses indicate that psychopathy is inherently related to an early attention bottleneck that minimizes the encoding and identification of secondary information and is not related to superior higher-order functioning; this interpretation is supported by the specific psychopathy-related deficit observed only in the early-alternative-focus condition. In the present study, psychopaths displayed a significant deficit in FPS in the early-alternative-focus condition, which was designed to establish an attention bias prior to presenting the threat-relevant information. Once established, this bias appeared to block psychopathic individuals' ability to process secondary threat information. This perspective is largely consistent with several findings showing that psychopaths are adept at focusing but have difficulty redistributing resources once attention is allocated (Jutai & Hare, 1983). Patterson and Newman (1993) posited that the allocation of attentional resources toward a goal-directed focus “seems to be relatively exaggerated in psychopaths” (p. 729). This attention bias has been found in a wide range of circumstances, including studies demonstrating perseverating approach responses in the face of passive avoidance cues (Newman & Kosson, 1986) and resistance to distraction in cued flanker tasks (Zeier, Maxwell, & Newman, 2009) and Stroop tasks (Hiatt, Schmitt, & Newman, 2004). Although the effect of psychopathy was significant only when psychopaths' alternative focus was engaged early in the information-processing stream, it is possible that attentional bias would emerge at a later stage under other circumstances (Vogel, Luck, & Shapiro, 1998).
A third focus of this study was to examine whether individual differences in working memory capacity moderate the effects of condition on FPS in psychopathy. The effects of working memory capacity on psychopathy-related differences in FPS were limited to the late-alternative-focus condition, which required participants to maintain a set-related focus in the face of emotion-related distractors (multilocus selection). Alternatively, this finding may also be consistent with an interpretation suggesting that higher-order cognitive processes resolve the competition between goal-relevant and secondary demands on attention. However, in light of the significant psychopathy-related difference in early selection and the lack of evidence for higher-order processing effects in the late-alternative-focus condition, we believe that the Psychopathy × Working Memory interaction highlights the importance of multilocus selection for maintaining the inherent attention bottleneck of psychopathic offenders. Although the bottleneck reduces or eliminates the need to use higher-order processes under certain circumstances, in other cases, the stability of the bottleneck may depend on higher-order processes to establish and maintain task-relevant, goal-directed behavior (Folk et al., 1992). There is substantial evidence that psychopathy involves a failure to process set-incongruent information (Hiatt et al., 2004; Mitchell et al., 2006). The current study suggests that such insensitivities reflect an early selection bias that may be moderated by situational demands and individual differences in higher-order cognitive processes. The fact that working memory capacity did not moderate the association between psychopathy and FPS in conditions other than the late-alternative-focus condition suggests that psychopaths do not generally use higher-order processes to suppress fear. Nevertheless, further research is needed to verify this finding and clarify the specific contexts that potentiate the importance of attention and working memory for threat processing in psychopathic individuals.
Finally, the fact that our findings for both PCL-R factors were highly similar to our findings for PCL-R total score suggests that this early selection effect is a general feature of the psychopathy construct. In addition, these findings complement our earlier findings (Newman et al., 2010) that attentional focus significantly moderated the association between FPS and each of these three dimensional constructs.
Overall, the findings reported in this article provide further support for the response modulation theory and for the importance of attentional abnormalities in psychopathy. Contemporary affective neuroscience has clearly established that attention and emotion are interdependent processes with bidirectional influences in clinical as well as nonclinical populations (Lang, Bradley, & Cuthbert, 1997; Pessoa, McKenna, Gutierrez, & Ungerleider, 2002; Phelps, 2006). In the present study, our manipulations of attentional focus and the timing of threat information demonstrated the differential importance of an attentional bottleneck for the subsequent fear processing of psychopathic individuals. Combined with the results of previous studies, these findings indicate that clarifying the anomalous cognitive-emotional interactions associated with psychopathy is a promising avenue for future research.
Acknowledgments
We thank many people at the Wisconsin Department of Corrections for making this research possible.
Funding This work was supported by Grants 5R01MH053041 and T32-MH018931 from the National Institute of Mental Health.
Footnotes
The early-alternative-focus interaction contrast and the moderation of the late-alternative-focus interaction contrast by Digits Backward score remained significant regardless of whether the 2 outliers (see the Participants section) were included in the analyses.
Because some investigators recommend using standardized transformations of the startle response prior to analysis to control for individual differences in overall reactivity and habituation, we reanalyzed our data applying a z transformation at the trial level. Although the overall results were quite similar, the first Helmert contrast (early alternative focus) was reduced to a trend, F(1, 85) = 3.39, p = .069.
Although this interaction contrast is not significant, the effect is in the same direction as the effect in the early-alternative-focus condition. In a supplemental analysis, we collapsed across focus, comparing the two threat-focus conditions with the two alternative-focus conditions. As in our previous study (Newman et al., 2010), attentional focus significantly moderated the association between psychopathy and FPS, F(1, 85) = 6.03, p = .016. Thus, focus is important, but it appears to be particularly important when goal-directed behavior is established prior to the presentation of threat cues.
Declaration of Conflicting Interests The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
References
- Awh E, Vogel EK, Oh S-H. Interactions between attention and working memory. Neuroscience. 2006;139:201–208. doi: 10.1016/j.neuroscience.2005.08.023. [DOI] [PubMed] [Google Scholar]
- Baskin-Sommers AR, Newman JP, Sathasivam N, Curtin JJ. Evaluating the generalizability of a fear deficit in psychopathic African American offenders. Journal of Abnormal Psychology. 2010 doi: 10.1037/a0021225. Advance online publication. doi: 10.1037/a0021225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenthal TD, Cuthbert BN, Filion DL, Hackley S, Lipp OV, Van Boxtel A. Committee report: Guidelines for human startle eyeblink electromyographic studies. Psychophysiology. 2005;42:1–15. doi: 10.1111/j.1469-8986.2005.00271.x. [DOI] [PubMed] [Google Scholar]
- Cleckley H. The mask of sanity. 5th ed. Mosby; St. Louis, MO: 1976. [Google Scholar]
- Corbetta M, Miezin FM, Dobmeye S, Shulman GL, Petersen SE. Selective and divided attention during visual discriminations of shape, color, and speed: Functional anatomy by positron emission tomography. The Journal of Neuroscience. 1991;11:2383–2402. doi: 10.1523/JNEUROSCI.11-08-02383.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desimone R, Duncan J. Neural mechanisms of selective visual attention. Annual Review of Neuroscience. 1995;18:193–222. doi: 10.1146/annurev.ne.18.030195.001205. [DOI] [PubMed] [Google Scholar]
- Deutsch JA, Deutsch D. Attention: Some theoretical considerations. Psychological Review. 1963;87:272–300. doi: 10.1037/h0039515. [DOI] [PubMed] [Google Scholar]
- Driver J. A selective review of selective attention research from the past century. British Journal of Psychology. 2001;92:53–78. [PubMed] [Google Scholar]
- Folk CL, Remington RW, Johnston JC. Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance. 1992;18:1030–1044. [PubMed] [Google Scholar]
- Forster KI, Forster JC. DMDX: A Windows display program with millisecond accuracy. Behavioral Research Methods. 2003;35:116–124. doi: 10.3758/bf03195503. [DOI] [PubMed] [Google Scholar]
- Fukuda K, Vogel EK. Human variation in overriding attentional capture. The Journal of Neuroscience. 2009;29:8726–8733. doi: 10.1523/JNEUROSCI.2145-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haenny PE, Schiller PH. State dependent activity in monkey visual cortex: I. Single cell activity in Vl and V4 on visual tasks. Experimental Brain Research. 1988;69:225–244. doi: 10.1007/BF00247569. [DOI] [PubMed] [Google Scholar]
- Hare RD. Manual for the Hare Psychopathy Checklist-Revised. 2nd ed. Multi-Health Systems; Toronto, Ontario, Canada: 2003. [Google Scholar]
- Hare RD, Harpur TJ, Hakstian AR. The revised Psychopathy Checklist: Reliability and factor structure. Psychological Assessment. 1990;2:338–341. [Google Scholar]
- Hart SD, Hare RD. Discriminant validity of the Psychopathy Checklist in a forensic psychiatric population. Psychological Assessment: A Journal of Consulting and Clinical Psychology. 1989;1:211–218. [Google Scholar]
- Hiatt KD, Schmitt WA, Newman JP. Stroop tasks reveal abnormal selective attention among psychopathic offenders. Neuropsychology. 2004;18:50–59. doi: 10.1037/0894-4105.18.1.50. [DOI] [PubMed] [Google Scholar]
- Hillyard SA, Anllo-Vento L. Event-related brain potentials in the study of visual selective attention. Proceedings of the National Academy of Sciences, USA. 1998;95:781–787. doi: 10.1073/pnas.95.3.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillyard SA, Vogel EK, Luck SJ. Sensory gain control (amplification) as a mechanism of selective attention: Electrophysiological and neuroimaging evidence. Philosophical Transactions of the Royal Society B: Biological Sciences. 1998;393:1257–1270. doi: 10.1098/rstb.1998.0281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jutai JW, Hare RD. Psychopathy and selective attention during performance of a complex perceptual-motor task. Psycho-physiology. 1983;20:146–151. doi: 10.1111/j.1469-8986.1983.tb03280.x. [DOI] [PubMed] [Google Scholar]
- Kastner S, Ungerleider LG. Mechanisms of visual attention in the human cortex. Annual Review of Neuroscience. 2000;23:315–341. doi: 10.1146/annurev.neuro.23.1.315. [DOI] [PubMed] [Google Scholar]
- Knudsen EI. Fundamental components of attention. Annual Review of Neuroscience. 2007;30:57–78. doi: 10.1146/annurev.neuro.30.051606.094256. [DOI] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM, Cuthbert MM. Motivated attention: Affect, activation and action. In: Lang PJ, Simons RF, Balaban MT, editors. Attention and orienting: Sensory and motivational processes. Erlbaum; Hillsdale, NJ: 1997. pp. 97–135. [Google Scholar]
- Lavie N, Hirst A, de Fockert JW, Viding E. Load theory of selective attention and cognitive control. Journal of Experimental Psychology: General. 2004;133:339–354. doi: 10.1037/0096-3445.133.3.339. [DOI] [PubMed] [Google Scholar]
- Lissek S, Pine DS, Grillon C. The strong situation: A potential impediment to studying the psychobiology and pharmacology of anxiety disorders. Biological Psychology. 2006;72:265–270. doi: 10.1016/j.biopsycho.2005.11.004. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA. The role of attention in feature detection and conjunction discrimination: An electrophysiological analysis. International Journal of Neuroscience. 1995;80:281–297. doi: 10.3109/00207459508986105. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA. The operation of selective attention at multiple stages of processing: Evidence from human and monkey electrophysiology. In: Gazzaniga MS, editor. The new cognitive neurosciences. 2nd ed. MIT Press; Cambridge, MA: 1999. pp. 687–700. [Google Scholar]
- MacCoon DG, Wallace JF, Newman JP. Self-regulation: The context-appropriate allocation of attentional capacity to dominant and non-dominant cues. In: Baumeister RF, Vohs KD, editors. Handbook of self-regulation: Research, theory, and applications. Guilford; New York, NY: 2004. pp. 422–446. [Google Scholar]
- Mitchell DVG, Richell RA, Leonard A, Blair RJR. Emotion at the expense of cognition: Psychopathic individuals outperform controls on an operant response task. Journal of Abnormal Psychology. 2006;115:559–566. doi: 10.1037/0021-843X.115.3.559. [DOI] [PubMed] [Google Scholar]
- Newman JP, Curtin JJ, Bertsch JD, Baskin-Sommers AR. Attention moderates the fearlessness of psychopathic offenders. Biological Psychiatry. 2010;67:66–70. doi: 10.1016/j.biopsych.2009.07.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newman JP, Kosson DS. Passive avoidance learning in psychopathic and nonpsychopathic offenders. Journal of Abnormal Psychology. 1986;95:257–263. [PubMed] [Google Scholar]
- Patrick CJ. Getting to the heart of psychopathy. In: Herve H, Yuille JC, editors. The psychopath: Theory, research, and social implications. Erlbaum; Hillsdale, NJ: 2007. pp. 207–252. [Google Scholar]
- Patterson CM, Newman JP. Reflectivity and learning from aversive events: Toward a psychological mechanism for the syndromes of disinhibition. Psychological Review. 1993;100:716–736. doi: 10.1037/0033-295x.100.4.716. [DOI] [PubMed] [Google Scholar]
- Pessoa L, McKenna M, Gutierrez E, Ungerleider LG. Neural processing of emotional faces requires attention. Proceedings of the National Academy of Sciences, USA. 2002;98:683–687. doi: 10.1073/pnas.172403899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phelps EA. Emotion and cognition: Insights from studies of the human amygdala. Annual Review of Psychology. 2006;24:27–53. doi: 10.1146/annurev.psych.56.091103.070234. [DOI] [PubMed] [Google Scholar]
- Vogel EK, Luck SJ, Shapiro KL. Electrophysiological evidence for a postperceptual locus of suppression during the attentional blink. Journal of Experimental Psychology: Human Perception and Performance. 1998;24:1656–1674. doi: 10.1037//0096-1523.24.6.1656. [DOI] [PubMed] [Google Scholar]
- Vogel EK, Woodman GF, Luck SJ. Pushing around the locus of selection: Evidence for the flexible-selection hypothesis. Journal of Cognitive Neuroscience. 2005;17:1907–1922. doi: 10.1162/089892905775008599. [DOI] [PubMed] [Google Scholar]
- Wechsler D. WAIS-III administration and scoring manual. Psychological Corp; San Antonio, TX: 1997. [Google Scholar]
- Yantis S, Johnston JC. On the locus of visual selection: Evidence from focused attention tasks. Journal of Experimental Psychology. 1990;16:135–149. doi: 10.1037//0096-1523.16.1.135. [DOI] [PubMed] [Google Scholar]
- Zachary RA. Shipley Institute of Living Scale: Revised manual. Western Psychological Service; Los Angeles, CA: 1986. [Google Scholar]
- Zeier JD, Maxwell JS, Newman JP. Attention moderates the processing of inhibitory information in primary psychopathy. Journal of Abnormal Psychology. 2009;118:554–563. doi: 10.1037/a0016480. [DOI] [PMC free article] [PubMed] [Google Scholar]