

Abstract
In humans declarative or explicit memory is supported by the hippocampus and related structures of the medial temporal lobe working in concert with the cerebral cortex. This paper reviews our progress in developing an animal model for studies of cortical–hippocampal interactions in memory processing. Our findings support the view that the cortex maintains various forms of memory representation and that hippocampal structures extend the persistence and mediate the organization of these codings. Specifically, the parahippocampal region, through direct and reciprocal interconnections with the cortex, is sufficient to support the convergence and extended persistence of cortical codings. The hippocampus itself is critical to the organization cortical representations in terms of relationships among items in memory and in the flexible memory expression that is the hallmark of declarative memory.
Keywords: hippocampus, cerebral cortex, memory, olfaction, rats
It is widely accepted that the hippocampus and closely related structures of the medial temporal lobe interact with the cerebral cortex in support of the persistence and organization of memories (1, 2, 3, 4, 5, 6). Here we review our own recent developments of a model system for exploring the nature of cortical and hippocampal memory representation and the specific roles played by hippocampal structures in memory formation.
This research is guided by a simple conception of cortical–hippocampal pathways in which there are three main serially and bidirectionally connected components: the cortex, the parahippocampal region, and the hippocampus itself (Fig. 1A). The beginning and end point of this system involves several tertiary or “association” cortical areas whose outputs converge on the parahippocampal region. This region, composed of the interconnected perirhinal, parahippocampal (or postrhinal), and entorhinal cortices, surround the hippocampus in both rats (7, 8) and monkeys (9, 10) and merge different sources of cortical input bound for the hippocampus. Within the hippocampus itself there are several stages of serial and parallel processing, and the outcome of this integration is sent back to the parahippocampal region, which in turn sends its main outputs to the same tertiary cortical areas that provided the source of input.
Figure 1.
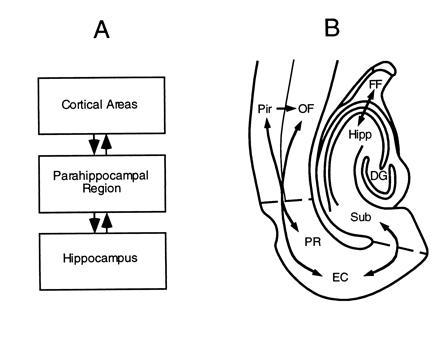
(A) Simple schematic diagram of cortical–hippocampal connections. (B) Outline of a horizontal rat brain section illustrating the locations and flow of information between components of the hippocampus, parahippocampal region, and adjacent cortical areas. DG, dentate gyrus; EC, entorhinal cortex; FF, fimbria–fornix; Hipp, hippocampus proper; OF, orbitofrontal cortex; Pir, perirhinal cortex; PR, perirhinal cortex; Sub, subiculum.
Our working hypothesis about this system is that memory representations are established and maintained in the cortex, and that the parahippocampal region and the hippocampus contribute to memory processing by modifying the persistence and organization of those cortical representations. Not all modifications of cortical representations depend on the hippocampal system. In particular hippocampal processing is not viewed as required for the “tuning” or “biasing” of cortical encodings, that is, an enhanced or diminished responsiveness to particular stimuli resulting from repeated exposure or association with reward or punishment. However, hippocampal structures receive all this information and are in position to compare and combine distinct cortical representations, and thus are viewed to play a critical role in extending the persistence of cortical memory representations and in creating and updating their organization. More specifically, we will argue that the two main components of the hippocampal region have distinct functions. First, the parahippocampal region, through its direct and reciprocal cortical connections, may be sufficient to extend the persistence of cortical memory representations. Second, the hippocampus itself plays a special role in mediating the systematic organization of cortical codings.
Olfactory Memory in Rodents as a Model System for Studies of Cortical–Hippocampal Processing
Our explorations focus on the set of interconnected olfactory–cortical and hippocampal structures that are fully evolved in rodents (Fig. 1B). Odor information initially processed by the olfactory bulb is sent directly to the piriform cortex and closely interconnected orbital prefrontal cortex (11, 12). Both of these cortical areas, as well as the olfactory bulb, project heavily to the perirhinal and entorhinal components of the parahippocampal region, which then provides the primary source of olfactory sensory information to the hippocampus itself. In the return pathway, the outputs of hippocampal processing involve direct projections from the parahippocampal region to both the piriform and orbital prefrontal cortices.
Physiological findings complement the anatomical data indicating that information processing in the olfactory and limbic systems are closely integrated during odor-guided learning and memory. During olfactory learning rats typically investigate an odor cue with about of 3–6 sniffs synchronized to the ongoing prominent theta rhythm recorded in the hippocampus (13). The reliability of this relationship is maximal just before the animal reaches accurate performance in the discrimination and just after a reversal of odor valences, two training stages when the rat is maximally attentive. In addition, hippocampal cellular activity includes firing bursts synchronized with the sniff and theta cycles, a pattern of neural activation that is consistent with optimal conditions for the induction of long-term potentiation during odor sampling (14).
In sum, olfactory–hippocampal pathways provide a straightforward and relatively simple instantiation of the general model of cortical–hippocampal interconnections. The olfactory and hippocampal systems are closely and reciprocally interconnected, and the neural activity patterns in the hippocampus reflect strong temporal integration with olfactory processing. Given these close connections, it is not surprising that rats have superb olfactory learning and memory capacities, as will be described below. Our efforts in exploring cortical–hippocampal interactions involve the development of behavioral tasks that assess different aspects of olfactory learning and memory ability, and identify specific functional contributions at each level of this system. This work involves a combination of neuropsychological studies on the effects of selective ablation of these areas, and electrophysiological studies that characterize the coding properties of single neurons in each area during memory performance.
Multiple Forms of Odor Memory Representation Within the Olfactory Cortex
Our starting point in describing the operations of this system involves a characterization of neural coding in the olfactory cortical areas that constitute the relevant perceptual representations for odor-guided learning. Based on our findings that rats with damage to the orbitofrontal cortex are severely impaired in olfactory learning (15, 16) and on the model described above, we expected that encodings of odor identity and the distinct significance of individual odors would be observed in olfactory cortical areas. In addition, we expected to observe firing patterns in olfactory cortical neurons that reflect both the extended persistence of odor representations and the organization of odor memories, properties that might depend on hippocampal processing. We sought neurophysiological correlates of all these aspects of memory processing in rats performing a specially designed odor discrimination task (17, 18).
Odor Coding and the Tuning and Biasing of Odor Representations.
The basic discrimination task involved a sequence of trials in which the rat was signaled by a light that it could initiate an odor presentation by poking its nose into a stimulus sampling port. When the odor was an assigned as a rewarded (S+) stimulus an additional nose poke into a separate water port resulted in water delivery, and no reward was given regardless of the response for other (S−) odors. On each trial one of eight different odor stimuli (identified here by number and assigned reward values) was presented, half of which were associated with a sweetened water reward and the other half not rewarded. We examined the coding of odor identity by comparing neural activity during odor sampling across the eight odors. In addition we assessed whether cortical odor codings could be “biased” or “tuned” in accordance with the significance of items by comparing responses across the groups of S+ and S− stimuli.
We found that many neurons in the orbitofrontal cortex and piriform cortex encode both odor identity and odor valence (Fig. 2). Although a few cells fired only to one odor, most were coarsely tuned as reflected by different levels of activation across the eight stimuli (Fig. 2A). In addition, most cells fired more strongly either to all the S+ odors or to all the S− odors. For example, in Fig. 2B the response was stronger to all S+ odors, and strongest of all to odors 1+ and 5+. An equivalent proportion of cells showed the opposite pattern, firing more strongly to all S− odors. The example shown in Fig. 2C shows larger responses to all S− odors, with the greatest response to odor 4−. This pattern of findings indicates, not surprisingly, that olfactory cortical areas encode odor identity. In addition, assigned stimulus valences are also very prominently represented in the cortex, indicating that cortical odor representations are indeed tuned or biased in association with reward contingencies.
Figure 2.
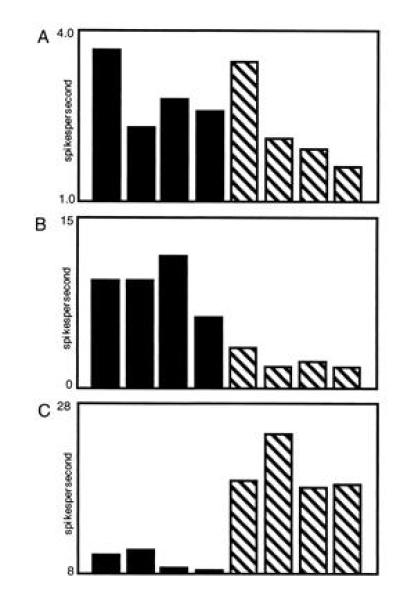
Odor responses of olfactory cortex neurons. The dark bars represent neural responses to S+ odors and striped bars represent responses to S− odors. OF, orbitofrontal.
Persistent Odor Representations in the Olfactory Cortex.
The odors were presented in a pseudorandom order (with specific exceptions described below) so that we could evaluate the extent to which particular odor responses were influenced by neural representations established on the preceding trial. In these analyses we controlled for the valence of the stimulus by separately analyzing data for preceding S+ and S− cues, and measured responses during odor sampling on subsequent trials. We found numerous cases of particular stimuli controlling subsequent odor responses. For example, in Fig. 3 the neural response during odor sampling was weak when the preceding trial involved odor 1+ and strong for other preceding S+ odors. This pattern of results indicates that a persistent representation of specific odors is carried over into later sensory processing. This capacity might subserve short-term memory required for comparisons between sequentially presented stimuli.
Figure 3.
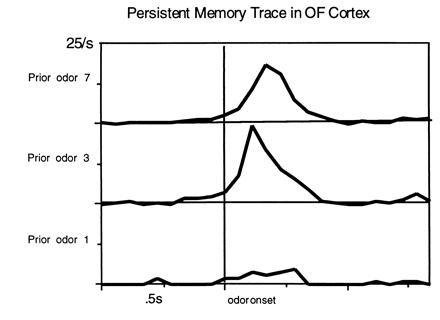
Short-term memory correlates of the activity of olfactory cortical neurons. Each panel illustrates the neural response of a cell to various current odors depending upon the stimulus on the preceding trial. OF, orbitofrontal.
Organized Odor Representations in Olfactory Cortex.
How might one detect an organization of odor representations in the cerebral cortex? Our efforts were directed at a simple kind of learned stimulus organization that might be found in sensory cortex, an acquired association between two odors such that presentation of one of them predicts the subsequent occurrence of the other (19). Assuming rats can learn a predictable relationship between odor stimuli, a neural reflection of the acquired association might be observed in the ability of presentation of the first odor to evoke or modify the neural representation of the second. We trained rats to associate sequentially presented odors by including specific exceptions to the otherwise random stimulus presentation order. For example, odors 5+ and 6− always were followed by odor 7+; as a control, odor 7+ also sometimes followed other odors unpredicted. Rats learned these incidental predictabilities in the odor sequence, as reflected in more rapid trial initiation when an S+ was predicted and slowed initiation of trials where S− odors were predicted.
We found two kinds of neural correlates of the learned odor associations. First we observed cells that fired differently in response to an odor depending on whether the preceding odor predicted its occurrence. For example, in Fig. 4A the response to odor 1+ was much reduced when it was predicted than when it was unexpected. Second, we identified olfactory cortex cells that fired in anticipation of a predicted odor. In the example shown in Fig. 4B the cell fired maximally prior to as well as during the presentation of odor 1+ when it was predicted. By contrast there was almost no response when the odor was not predicted by its associate. This pattern of results indicates that learned stimulus associations are indeed established within the olfactory cortex.
Figure 4.

Neural correlates of odor–odor associations in the olfactory cortex. See text for description.
In sum, our characterization of olfactory cortical neurons indicated that the piriform and orbitofrontal cortical areas encode aspects of stimulus memories that should occur independent of hippocampal function, specifically the tuning and biasing of odor representations, as well as aspects of memory that may depend on hippocampal function, such as the persistence of odor memories and odor–odor associations.
Memory Processing Within the Hippocampal System: Which Structures Extend the Persistence of Odor Memories?
The earliest reports of amnesia in humans following damage to the hippocampal region emphasized the critical role of the hippocampus in bridging between immediate memory and permanent memory formation (20). In amnesic patients and animals with hippocampal region damage, deficits in memory persistence are characteristically observed as a dissociation between intact immediate memory versus subsequent abnormally rapid forgetting. Initially it was thought that the hippocampus itself played the primary role in extending retention beyond immediate memory, but recent data have brought this view into question (21, 22). Our studies have focused on the issue of which components of the hippocampal system are critical to the persistence of odor memory representations.
Efforts to delineate the anatomical structures involved in maintaining a memory trace have focused on the delayed nonmatch to sample task (DNMS). In most versions of this task the subject is initially presented with a sample memory cue. This is followed by a delay phase during which the memory for that cue must be maintained. Finally, in the choice phase, the subject is presented with the sample and a novel stimulus and the unfamiliar “nonmatch” cue must be selected. The load on memory can be increased by lengthening the delay phase or by presenting a “list” of sample cues prior to a series of choice recognition tests. This paradigm was first developed for monkeys by using three-dimensional junk objects that provide rich and salient cues for this species (23, 24). Damage to the hippocampal system results in a performance deficit that is dependent on the duration of the delay or the length of the sample “list,” revealing a delay-dependent deficit in new learning similar to that observed in human amnesics (25, 26).
To assess which components of the hippocampal system are critical to the maintenance of odor memory representations we developed a variant of the DNMS task that used odor cues and involved a stimulus presentation protocol that could be used to assess neural responses to single stimuli as well as behavioral responses in accordance with the nonmatch memory contingency. In this version of the task, a continuous series of single odor cues is presented with the contingency that an appropriate response to each odor is rewarded only if that odor was different from (that is, a nonmatch with) the preceding odor. Rats were trained initially with a short (3-sec) interval between odor presentations. Subsequently the interval between odor presentations was manipulated to vary the retention delay, allowing an assessment of forgetting rate as in the studies on monkeys.
In an initial experiment we compared the effects of selective ablation of the parahippocampal region versus that of damage to the fornix, a major fiber bundle that connects the hippocampus itself with subcortical structures (ref. 27; see Fig. 1B). Normal rats acquire this task within ≈150 trials and neither lesion affected the acquisition rate. In subsequent tests across memory delays, intact rats showed 90% or better retention at the shortest (3-sec) delay and their memory performance gradually declined as the delay was increased. Rats with damage to the parahippocampal region also showed good retention at the shortest delay, but their performance declined abnormally rapidly, showing a severe deficit within 1 min. By contrast rats with fornix lesions were unimpaired across delays, showing the same gradual memory decay as intact rats. These results indicate that neither hippocampal component is critical to odor perception, acquisition of the nonmatch rule, or immediate memory. However, the parahippocampal region is critical to extending the persistence of cortical representations beyond immediate memory in rats, as it is in monkeys (28, 29, 30). Furthermore, through its direct and reciprocal connections with the cortex, the parahippocampal region is sufficient to mediate this memory function independent of hippocampal processing. The results of this processing may be reflected in the persistent effects of odor representations we observed in olfactory cortex, as described above.
In complementary electrophysiological studies we have also examined the response properties of neurons in the parahippocampal region and the hippocampus in rats performing the odor-guided DNMS task (31). Cells in the perirhinal cortex, in the lateral entorhinal cortex, and in the subiculum showed selective or coarse tuning to odor stimuli, indicating that the parahippocampal region encodes specific odor representations. In addition, odor representations in each of these areas was observed to persist through the memory delay. Some cells showed striking odor-specific activity during odor sampling and throughout the delay period, such as the example shown in Fig. 5. To detect persistent memory representations across all cells, we focused on the trial period immediately preceding the initiation of trials. At this time the overt behavior of the animal is consistent (it is approaching the stimulus port), allowing us to determine if neural activity at this time varies with the identity of the odor that must be remembered during that period. A substantial number of cells throughout the parahippocampal region fired differentially associated with the preceding sample odor at this time. Some cells maintained differential activity for both short and long memory delays, whereas others lost the representation at the longer delay.
Figure 5.
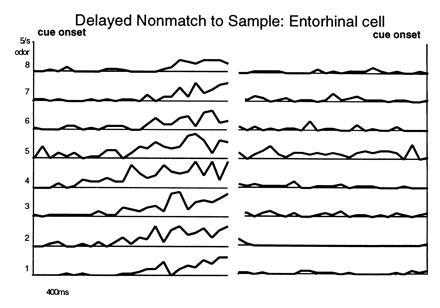
Neural activity in the parahippocampal region associated with DNMS performance. (Left) Responses of the cell during the 2 sec following the onset of different odors. (Right) Activity during the last 2 sec of the that delay period—i.e., prior to the onset of the subsequent odor cue. Note greater activity for odor 5 in both periods.
To assess odor memory representations in the hippocampus itself we recorded from putative pyramidal neurons in CA1 of rats performing the same task. A large proportion of hippocampal cells fired associated with one or more of each identifiable behavioral event in the task. A subset of these cells was selectively active during stimulus sampling, and the activity of some of these hippocampal cells reflected the “match” or “nonmatch” relationship critical to performance in this task (ref. 32; Fig. 6). However, unlike cells in the parahippocampal region, none of these cells firing associated with particular odors that composed specific comparisons. Rather cellular activity reflected all comparisons with the same outcome. We interpreted this finding as entirely consistent with the results of our lesion studies—it appears that the hippocampus itself is not involved in the encoding and storage of representations for specific items in the this task. Instead it appears the hippocampus encodes abstract relations among cues in DNMS, as in many other situations (33). In this task the hippocampus may mediate memory for match and nonmatch trial outcomes, even though this information contributes little directly to performance on this simple recognition task. Consistent with our data from the lesion study, the parahippocampal region, but not the hippocampus, maintains specific and persistent representations sufficient to support recognition performance.
Figure 6.
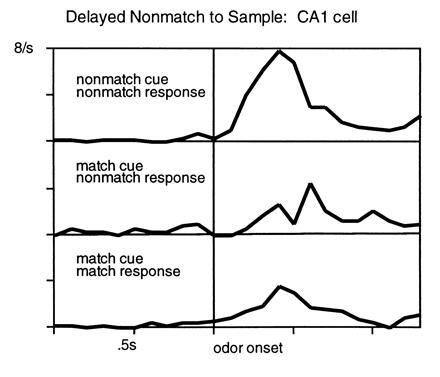
Neural activity in the hippocampus associated with DNMS performance. See text for description.
The Role of Hippocampal Structures in the Organization of Memory Representations
In addition to its role in extending the persistence of memory representations many investigators have also suggested that the hippocampus is critical for only one “kind of memory” or one form of memory representation. In humans, there is considerable agreement that the hippocampal region is critical specifically for declarative memory, the capacity for conscious and explicit recollection (34, 35, 36, 37). By contrast, the acquisition of biases or adaptations to individual items, engaged through repetition of the learning event and revealed typically by implicit measures of memory, is intact following hippocampal damage. To study the representational features of hippocampal-dependent memory in animals we have focused on two characteristic performance capacities associated with declarative memory: the ability to store and remember relationships among perceptually distinct items and the ability to express these memories flexibly in novel situations. Furthermore, as in our studies on the olfactory cortex, this work involves performance in learning and remembering relationships between odor stimuli as a prototypical example of declarative memory processing.
In attempting to understand the neural mechanisms that underlie learning-stimulus relations it is important to consider two general ways by which stimulus representations could become bound to one another. In his classic considerations of the “binding problem” in perception and memory, William James (38) suggested that stimuli may either be conceived as not distinct from one another and consequently might be bound by a conceptual fusion or, alternatively, might be discriminated as separate and then bound by association in memory. Indeed these two forms of stimulus binding can be distinguished in the performance of human amnesics. Amnesics are typically impaired in learning new associations, but with extra effort they do sometimes succeed. In these cases the associations appear to be too well bound such that amnesics find it abnormally difficult to express memory for the original elements of a successfully acquired association when the elements are subsequently separated. Such associations are characterized as “hyperspecific” in that they can be expressed only in highly constrained conditions that imitate the conditions of original learning. For example, in one experiment that involved learning baseball facts in a question-and-answer format, a densely amnesic subject could correctly recall answers only if the test procedure included precise repetitions of the original questions used during learning (35).
Hyperspecificity of associations has also been observed in animals with damage to the hippocampal system. For example, in our own previous work we found that rats with damage to the hippocampal system were abnormally inclined to bind together the representations of stimuli that were closely juxtaposed in olfactory or spatial learning (39). These rats were able to perform odor-discrimination problems when they had to choose between two discriminative cues presented in frequently experienced pairings but, unlike normal rats, they could not recognize the same stimuli in probe trials that involved novel pairings of familiar odor cues taken from different discrimination problems. Similarly, we found that rats with hippocampal system damage could learn to use distal spatial cues to locate an escape platform in the Morris water maze when they were allowed to begin trials from a consistent starting point but, unlike normal rats, they could not use these same cues to navigate to the escape locus in probe trials where they had to view those familiar stimuli from novel starting points in the maze. Our interpretation of this data is that amnesia associated with damage to the hippocampal system distinguishes between James’ two forms of binding; amnesics are abnormally inclined to fuse rather than distinguish and associate items.
These considerations led us to examine the role of the hippocampal system in a classic form of stimulus–stimulus association, paired-associate learning. The verbal paired-associate task has been exceedingly useful in understanding cognitive aspects of associative learning in humans and is often applied in the assessment of amnesia. It seemed to us that a paired-associate task adapted for animals was the simplest paradigm that we could exploit for neurobiological studies on the learning of new associations between distinct and neutral stimulus events. The paired-associate task as typically used for humans involves presentation of a list of arbitrarily paired words followed by testing in which the subject is cued with the first item of each pair and must recall the second item. For rats, we designed an analogous task using odor stimuli and a recognition format that required subjects to distinguish appropriate odor pairs from a large number of foils (40).
Rats were trained to perform a nose poke into a sniff port when a signal light was illuminated. They sniffed two odors presented in rapid succession, separated by a period when airflow was reversed to prevent stimulus blending. Four rewarded odor sequences (paired associates) were composed out of eight different odors (A-B, C-D, E-F, G-H). When the rat smelled a rewarded pair (in either order, e.g., A-B or B-A), it could obtain a sweetened water reward from the water port. There were two kinds of unrewarded “foil” odor sequences. One kind (mispairings) was composed of the same odors used to form the paired associates but presented in different combinations—e.g., A-C. There were 48 of these mispair sequences. To distinguish a mispairing from a paired associate, the rat had to learn the arbitrarily assigned association between the odors. The other type of foil (nonrelational sequences) involved one of the odors A through H combined with one of four other odors that was never associated with reward (W through Z). There were 64 of these nonrelational sequences. To distinguish a nonrelational sequence from a paired associate the rat was required only to recognize the never-rewarded odor in the sequence. The inclusion of both types of foils allowed us to examine in the same subjects the effects of hippocampal system damage on associative and nonassociative learning.
We began our experiments on paired-associate learning by examining the performance of rats in which the parahippocampal region had been removed, effectively eliminating the contributions of both that area and the hippocampus itself. Intact rats and rats with parahippocampal area lesions learned to distinguish nonrelational pairs from paired associates at the same rate. In addition, normal rats gradually learned to distinguish paired associates from odor mispairings. By contrast, rats with parahippocampal lesions could not learn to distinguish paired associates from mispairings, even when given nearly twice as many training trials as normal rats. Similar findings of impaired stimulus–stimulus association have been made in monkeys (41). In a subsequent study (42) we evaluated the role of the hippocampus itself in paired-associate learning using the identical task and testing procedures. Selective neurotoxic lesions of the hippocampus also affected paired-associate learning and had no effect on learning nonrelational sequences. However, by contrast to the severe impairment observed after parahippocampal region lesions, hippocampal lesions resulted in a striking facilitation in distinguishing paired associates from mispairings. This combination of findings indicates that both areas normally contribute to paired-associate learning, and suggests their functions are different and perhaps antagonistic. The results led us to speculate that stimulus representations involved in a paired associate could be encoded in two fundamentally different and opposing ways, one subserved by the parahippocampal region and another mediated by the hippocampus (43).
One form of encoding could involve the fusion of the two odor representations just as James (38) described occurs when the items are not conceptually distinct. More recently and to differing ends, Schacter (36) characterized this type of representation as a “unitized structure,” and others (44, 45) have referred to such an encoding as a “configural” representation. Extending our results showing that the parahippocampal region can maintain persistent stimulus representations, we have suggested this area can combine items that occur sequentially as well as simultaneously (22, 43). In this way the parahippocampal region could mediate the encoding of the elements of paired associates as fused, unitized, or configural representations. Alternatively, stimulus elements in paired associates could be separately encoded and then have their representations associated in memory. An “association” of this type differs from a fused representation in that it maintains the compositionality of the elemental representations and organizes them according to the relevant relationships among the items. We have previously argued that such relational representations are mediated by the hippocampal system (39); based on the findings on paired-associate learning, our current view is that relational memory processing is mediated specifically by the hippocampus itself (43).
To distinguish these two types of stimulus–stimulus representation we developed two other variants of the paired-associate paradigm. To speed the rate of learning paired associates, we also adopted new testing methods that involved more “naturalistic” behaviors for memory testing. A central feature of our new tasks was that rats were required to express the memories of odor–odor associations in novel situations where the learned odor elements were separated and one of them had to be used to guide behavioral responses that differed from those involved in the initial learning. Because these demands of memory expression require a compositional representation, our expectation was that the hippocampus itself would be required for performance in such tests of paired-associate learning.
One of these experiments involved a “natural” form of paired associate learning by rats developed by Galef (46) to study the social transmission of odor selection. He has shown that rats learn from conspecifics which foods are preferable by experiencing the pairing of a distinctive (not necessarily novel) food odor with an odorous constituent of rat’s breath (carbon disulfide). Retention of learning in Galef’s task required rats to employ the learned association of the distinctive food odor to guide subsequent food selection during an explicit choice between multiple foods. We (ref. 47; also see ref. 48) have recently found that long-term memory for this form of paired-associate learning is blocked by selective neurotoxic lesions of the entire hippocampus, indicating that the memory for paired associates does depend on the hippocampus itself in a situation where the relevant stimulus relationships are set in a “natural” context and memory expression differs from repetition of the learning event (Fig. 7).
Figure 7.
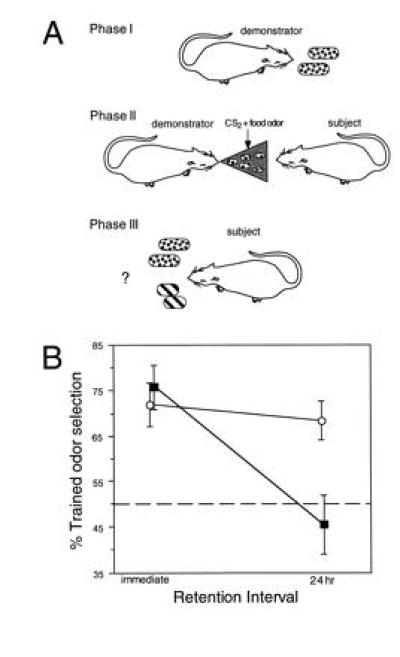
(A) Training and testing on a “naturalistic” odor–odor association. In phase I a “demonstrator” rat is given a distinctively scented food. Then, in phase II, the demonstrator is presented to an experimental subject for a brief period of social interaction. During this experience subjects associate two odors carried on the demonstrator’s breath, the distinctive food odor and carbon disulfide (CS2), a natural constituent of rats’ breath (ref. 46; phase II). In phase III, to test memory for the food odor–CS2 association, subjects are presented with the same food or another distinctively scented food, either immediately or after a 24-hr delay. After this training normal rats show a strong selection preference for the trained food odor. (B) The effects of hippocampal lesions on performance in the retention test. Retention was intact immediately after learning but severely impaired during a 24-hr retention test.
In addition, to more explicitly examine the flexibility of memory dependent on the hippocampus itself, we developed another paired-associate task that was used to assess an animal’s ability to infer relations among associated elements presented in novel configurations (49). As described above, animals with selective hippocampal damage can acquire odor–odor representations in learning responses to representations of specific odor pairings. However, we believe this learning is supported by fused stimulus representations that are “hyperspecific,” rendering the animals unable to make flexible and inferential judgments about the same items when presented in unusual ways. Exploiting rodents’ natural foraging strategies that employ olfactory cues, animals were trained with stimuli that consisted of distinctive odors added to a mixture of ground rat chow and sand through which they dug to obtain buried cereal rewards. On each paired-associate trial one of two sample odors initially presented was followed by two choice odors, each assigned as the “associate” of one of the samples and baited only when preceded by that sample (Fig. 8A). Following training on two sets of overlapping odor–odor associations, subsequent probe tests were used to characterize the extent to which learned representations supported two forms of flexible memory expression, transitivity, the ability to judge inferentially across stimulus pairs that share a common element, and symmetry, the ability to associate paired elements presented in the reverse of training order.
Figure 8.
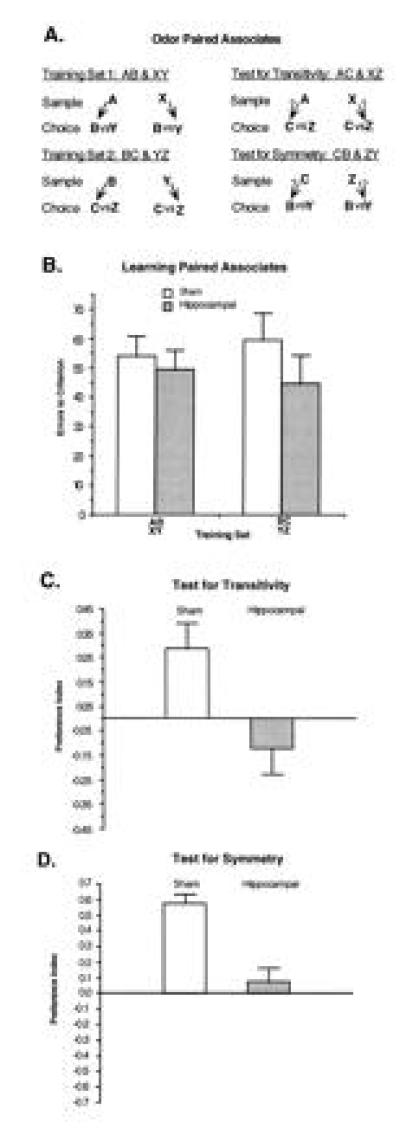
Inferential expression of odor–odor associations. (A) Schematic diagram of paired associate training and probe testing. Letters represent odor stimulus items; arrows without question marks indicate trained pairings, whereas arrows with question marks indicate expected transitive and symmetrical choices. Rats are first trained on two overlapping sets of paired associates (Left). Then (Right) they are tested for inferential expression in two ways. In the test for transitivity, they are presented with one of two sample cues from the first training set and the required to select between the choice cues from the second set, based on the shared associates of these items. In the test for symmetry or “reversibility” of the associations, they are presented with one of two choice cues from the second set and required to select the appropriate sample cue from that set. (B) Errors to criterion on acquisition of the two sets of paired associates for sham-operated and hippocampal subjects. (C) Preference indices on the test for transitive inference. For these probe trials a preference score was calculated as (X − Y)/(X + Y), where X and Y were the digging times in the transitive and alternate choices, respectively. (D) Preference indices on the test for symmetrical expression.
Intact rats learned paired associates rapidly and hippocampal damage did not affect acquisition rate on either of the two training sets, consistent with recent reports on stimulus–stimulus association learning in rats and monkeys (refs. 41 and 42; Fig. 8B). Intact rats also showed strong transitivity across the sets with a preference of ≈2:1 in favor of choice items indirectly associated with the presented sample (Fig. 8C). By contrast rats with selective hippocampal lesions were severely impaired in that they showed no evidence of transitivity. In the symmetry test, intact rats again showed the appropriate preference of ≈3:1 in the direction of the symmetrical association (Fig. 8D). By contrast, rats with hippocampal lesions again were severely impaired, showing no significant capacity for symmetry.
These findings provide compelling evidence that some form of stimulus–stimulus representations can be acquired independent of the hippocampus itself (see refs. 50 and 51), although this form of representation is hyperspecific. Only a hippocampally mediated representation can support the flexible expression of associations among items within a larger organization. Collectively, the findings from our studies on paired-associate learning in animals provide an extension of classic views on human memory, such as William James’ (38) description of “memory” as involving an elaborated network of associations that can be applied across a broad range of situations, as distinct from “habits” that depend on rigid associative sequences. These findings are also entirely consistent with present day characterizations of human declarative memory, such as Cohen’s (33) description of declarative memory as “promiscuous” in its accessibility by novel routes of expression. Our experiments using a rodent model of declarative memory show this capacity is dependent on the circuitry within the hippocampus itself.
Conclusions
In everyday life surely the distinct aspects of cortical and hippocampal memory processing are intertwined. The prominence of bidirectional connections between cortical and hippocampal structures would make it difficult to have parallel coexisting short-lived and persistent, or fused and associated representations at distinct levels of the system. Rather, as indicated in our characterization of neuronal firing properties in the olfactory cortex, interactions among these structures likely results in a unified persistence and form of representation throughout the system in intact animals. How these interactions unfold among components of the cortical–hippocampal system should become a main target of interest in studies on the operation of this system.
Acknowledgments
This work was supported by grants from the National Institute of Mental Health, the National Institute on Aging, the Office of Naval Research, and the Human Frontiers Science Program.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviation: DNMS, delayed nonmatch to sample.
References
- 1.Gray J A. The Neuropsychology of Anxiety: An Enquiry into the Functions of the Septo-Hippocampal System. New York: Oxford Univ. Press; 1982. [Google Scholar]
- 2.Halgren E. In: Neuropsychology of Memory. Butters N, Squire L R, editors. New York: Guilford; 1984. pp. 165–182. [Google Scholar]
- 3.Rolls E. In: Neural Models of Plasticity: Theoretical and Empirical Approaches. Byrne J H, Berry W O, editors. New York: Academic; 1989. pp. 240–265. [Google Scholar]
- 4.Squire L R, Cohen N J, Nadel L. In: Memory Consolidation. Weingartner H, Parker E, editors. Hillsdale, NJ: Erlbaum; 1984. pp. 185–210. [Google Scholar]
- 5.Teyler T J, DiScenna P. Behav Neurosci. 1986;100:147–154. doi: 10.1037//0735-7044.100.2.147. [DOI] [PubMed] [Google Scholar]
- 6.Wickelgren W A. Psychol Rev. 1979;86:44–60. [PubMed] [Google Scholar]
- 7.Burwell R D, Witter M P, Amaral D G. Hippocampus. 1995;5:390–408. doi: 10.1002/hipo.450050503. [DOI] [PubMed] [Google Scholar]
- 8.Deacon T W, Eichenbaum H, Rosenberg P, Eckmann K W. J Comp Neurol. 1983;220:168–190. doi: 10.1002/cne.902200205. [DOI] [PubMed] [Google Scholar]
- 9.Suzuki W A, Amaral D G. J Neurosci. 1994;14:1856–1877. doi: 10.1523/JNEUROSCI.14-03-01856.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Witter M P, Groenewegen H J, Lopes da Silva F H, Lohman A H M. Progr Neurobiol. 1989;33:161–254. doi: 10.1016/0301-0082(89)90009-9. [DOI] [PubMed] [Google Scholar]
- 11.Kosel K C, Van Hoesen G W, West J R. J Comp Neurol. 1981;198:467–482. doi: 10.1002/cne.901980307. [DOI] [PubMed] [Google Scholar]
- 12.Price J L, Carmichael S T, Carnes K, Clugnet M-C, Kuroda M, Ray J P. In: Olfaction: A Model System for Computational Neuroscience. Davis J L, Eichenbaum H, editors. Cambridge, MA: MIT Press; 1991. pp. 101–120. [Google Scholar]
- 13.Macrides F, Eichenbaum H, Forbes W B. J Neurosci. 1982;2:1705–1717. doi: 10.1523/JNEUROSCI.02-12-01705.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Otto T, Eichenbaum H, Wiener S I, Wible C G. Hippocampus. 1991;1:181–192. doi: 10.1002/hipo.450010206. [DOI] [PubMed] [Google Scholar]
- 15.Eichenbaum H, Eckmann K W, Shedlack K J. Brain Behav Evol. 1980;17:255–275. doi: 10.1159/000121803. [DOI] [PubMed] [Google Scholar]
- 16.Eichenbaum H, Clegg R A, Feeley A. Exp Neurol. 1983;79:434–451. doi: 10.1016/0014-4886(83)90224-8. [DOI] [PubMed] [Google Scholar]
- 17.Schoenbaum G, Eichenbaum H. J Neurophysiol. 1995;74:733–750. doi: 10.1152/jn.1995.74.2.733. [DOI] [PubMed] [Google Scholar]
- 18.Schoenbaum G, Eichenbaum H. J Neurophysiol. 1995;74:751–762. doi: 10.1152/jn.1995.74.2.751. [DOI] [PubMed] [Google Scholar]
- 19.Sakai K, Miyashita Y. Nature (London) 1991;354:152–155. doi: 10.1038/354152a0. [DOI] [PubMed] [Google Scholar]
- 20.Scoville W B, Milner B. J Neurol Neurosurg Psychiatry. 1957;20:11–21. doi: 10.1136/jnnp.20.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Brown M W, Wilson F A W, Riches I P. Brain Res. 1987;409:158–162. doi: 10.1016/0006-8993(87)90753-0. [DOI] [PubMed] [Google Scholar]
- 22.Eichenbaum H, Otto T, Cohen N J. Brain Behav Sci. 1994;17:449–518. [Google Scholar]
- 23.Gaffan D. J Comp Physiol Psychol. 1974;86:1100–1109. doi: 10.1037/h0037649. [DOI] [PubMed] [Google Scholar]
- 24.Mishkin M, Delacour J. J Exp Psychol Ann Behav Proc. 1975;1:326–334. doi: 10.1037//0097-7403.1.4.326. [DOI] [PubMed] [Google Scholar]
- 25.Mishkin M. Nature (London) 1978;273:297–298. doi: 10.1038/273297a0. [DOI] [PubMed] [Google Scholar]
- 26.Zola-Morgan S, Squire L R. Behav Neurosci. 1985;99:22–34. doi: 10.1037//0735-7044.99.1.22. [DOI] [PubMed] [Google Scholar]
- 27.Otto T, Eichenbaum H. Behav Neurosci. 1992;106:1763–1776. doi: 10.1037//0735-7044.106.5.762. [DOI] [PubMed] [Google Scholar]
- 28.Meunier M, Bachevalier J, Mishkin M, Murray E A. J Neurosci. 1993;13:5418–5432. doi: 10.1523/JNEUROSCI.13-12-05418.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Murray E A, Mishkin M. J Neurosci. 1986;6:1991–2003. doi: 10.1523/JNEUROSCI.06-07-01991.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zola-Morgan S, Squire L R, Amaral D G, Suzuki W A. J Neurosci. 1989;9:4355–4370. doi: 10.1523/JNEUROSCI.09-12-04355.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Young B J, Otto T, Fox G D, Eichenbaum H. Soc Neurosci Abstr. 1995;21:943. [Google Scholar]
- 32.Otto T, Eichenbaum H. Hippocampus. 1992;2:323–334. doi: 10.1002/hipo.450020310. [DOI] [PubMed] [Google Scholar]
- 33.Eichenbaum H. Curr Opin Neurobiol. 1996;6:187–195. doi: 10.1016/s0959-4388(96)80072-9. [DOI] [PubMed] [Google Scholar]
- 34.Cohen N J, Eichenbaum H. Memory, Amnesia, and the Hippocampal System. Cambridge, MA: MIT Press; 1993. [Google Scholar]
- 35.Cohen N J, Squire L R. Science. 1980;210:207–210. doi: 10.1126/science.7414331. [DOI] [PubMed] [Google Scholar]
- 36.Schacter D L. In: Memory Systems of the Brain. Weinberger N M, McGaugh J L, Lynch G, editors. New York: Gilford; 1985. pp. 351–380. [Google Scholar]
- 37.Squire L R. Psychol Rev. 1992;99:195–231. doi: 10.1037/0033-295x.99.2.195. [DOI] [PubMed] [Google Scholar]
- 38.James W. The Principles of Psychology. New York: Holt; 1890. 1918 Ed. [Google Scholar]
- 39.Eichenbaum H, Otto T, Cohen N J. Behav Neural Biol. 1992;57:2–36. doi: 10.1016/0163-1047(92)90724-i. [DOI] [PubMed] [Google Scholar]
- 40.Bunsey M, Eichenbaum H. Behav Neurosci. 1993;107:740–747. doi: 10.1037//0735-7044.107.5.740. [DOI] [PubMed] [Google Scholar]
- 41.Murray E A, Gaffan D, Mishkin M. J Neurosci. 1993;13:4549–4561. doi: 10.1523/JNEUROSCI.13-10-04549.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bunsey M, Eichenbaum H. Soc Neurosci Abstr. 1993;19:358. [Google Scholar]
- 43.Eichenbaum H, Bunsey M. Curr Dir Psychol Sci. 1995;4:19–23. [Google Scholar]
- 44.Rescorla R A. J Comp Physiol Psychol. 1972;79:307–317. doi: 10.1037/h0032553. [DOI] [PubMed] [Google Scholar]
- 45.Sutherland R J, Rudy J W. Psychobiology. 1989;17:129–144. [Google Scholar]
- 46.Galef B G. In: Contemporary Issues in Comparative Psychology. Dewsbury D A, editor. Sunderland, MA: Sinauer; 1990. pp. 55–79. [Google Scholar]
- 47.Bunsey M, Eichenbaum H. Hippocampus. 1995;5:546–556. doi: 10.1002/hipo.450050606. [DOI] [PubMed] [Google Scholar]
- 48.Winocur G. Behav Brain Res. 1990;38:145–154. doi: 10.1016/0166-4328(90)90012-4. [DOI] [PubMed] [Google Scholar]
- 49.Bunsey M, Eichenbaum H. Nature (London) 1996;379:255–257. doi: 10.1038/379255a0. [DOI] [PubMed] [Google Scholar]
- 50.Moscovitch M. In: Memory Systems 1994. Schacter D L, Tulving E, editors. Cambridge, MA: MIT Press; 1994. pp. 269–310. [Google Scholar]
- 51.Musen G, Squire L R. Neuropsychology. 1993;7:119–135. [Google Scholar]