

The sequence, structure, and function of natural biopolymers frequently appear to be optimally adapted to their metabolic and physiological tasks. So much so that “scientific creationists” frequently argue that natural biopolymers could not have possibly arisen by chance, and that the conservation of histone sequence is a sign of providential necessity rather than natural chance. Evolutionary biologists reply that random mutation and natural selection are more than capable of generating highly functional macromolecules, but direct, experimental validations of this stipulation sometimes have proven difficult to find: it is one thing to hypothesize that a complex macromolecule such as tRNA could have arisen by chance, it is quite another to prove it. Although the evolutionary basis of biology has not been in doubt for more than a hundred years, it would nonetheless be satisfying to actually witness the evolution of complex macromolecules instead of merely inferring the lengthy chain of mutation and selection that led to their fixation. Experiments in artificial or directed evolution have the potential not just to bolster evolutionary theory but to flesh out the basis of this theory—the notion that function can be acquired frequently enough by random chance to initiate and sustain life. Stephen Gould (1) is fond of saying that if we could rerun the tape of life, the biological answers that would be found would be different. Although we cannot yet rerun the full feature film, it is now possible to at least examine vignettes for molecular evolution.
Although Sol Spiegelman, Manfred Eigen, and other pioneers (2, 3) initiated the study of directed molecular evolution using Qβ bacteriophage, in recent years such experiments have become much more powerful due to the advent of facile methods for DNA synthesis and nucleic acid amplification. Now, functional nucleic acid binding species (aptamers) and catalysts can be selected from partially or completely random sequence pools (4–8). Such in vitro selection experiments can provide insights into what possibilities may have been available during the course of molecular evolution. For example, how many different sequences or structures might be functionally similar to tRNA, and thus, what is the probability that a molecule such as tRNA might have arisen by chance?
The paper by Klug et al. (9) is an excellent example of the application of in vitro selection methods to such evolutionary questions. A nonstandard amino acid, selenocysteine, is specifically incorporated into formate dehydrogenase (fdhF) of Escherichia coli during translation. An opal (UGA) codon in the genes encoding these proteins can interact with a unique aminoacylated tRNA (tRNASec). Obviously, this dual use of the opal stop codon prompts questions such as what precludes termination from occurring at selenocysteine insertion sites, and what precludes other opal codons from coding for selenocysteine? Previous work, primarily from the laboratory of August Böck (10), has shown that there is a downstream hairpin structure in the mRNA that directs this particular opal codon to be used as a selenocysteine-incorporation site. The hairpin in turn interacts with an EF-Tu variant known as SelB. Whereas the N-terminal 70% of SelB is homologous to elongation factor EF-Tu (11), the C-terminal region of the protein contains the hairpin-binding activity (12). These findings support a model in which the hairpin serves as the initial docking site for SelB and its bound selenocysteyl-tRNASec (Fig. 1). This model for selenocysteine incorporation appears to be widely applicable and holds in all prokaryotes that have been studied so far. For example, all known SelB-binding hairpins fold into a similar stem–loop structure with a consensus GGUC end loop. Studies using chemical protection (13), nuclease and iodine-cleavage protection (14), and toeprinting analysis (15) had indicated that the end loop comprised the main recognition element. This was further verified by demonstrating that mutations in this loop diminished or abolished binding (16) and a 17-nucleotide minihelix composed of the upper half of the hairpin was capable of binding SelB without loss of specificity or affinity (12).
Figure 1.
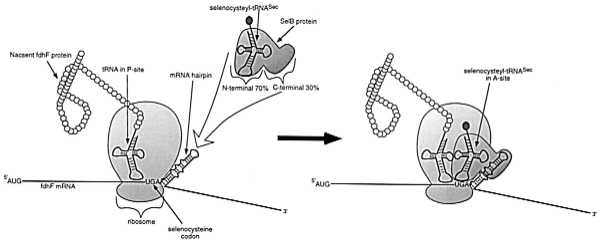
Schematic model of site-specific selenocysteine incorporation in E. coli formate dehydrogenases by alternative usage of the UGA stop codon. A SelB-GTP-selenocysteyl-tRNASec ternary complex (GTP not shown) docks to the apical region of the mRNA hairpin located immediately downstream of the opal codon. The N-terminal and central domains of SelB are homologous to EF-Tu and bind the selenocysteyl-tRNASec and GTP. The C-terminal region of SelB is responsible for recognition of the mRNA hairpin. SelB thus facilitates the incorporation of selenocysteine into the A site of the ribosome and into the nascent fDHF protein.
Klug et al. (9) have attempted to discern how likely or optimal the evolution of the SelB-binding hairpin would have been. A pool of fdhF hairpin variants was synthesized in which each position contained 70% wild-type residues and 10% of each non-wild-type residue; in addition, positions corresponding to the selenocysteine-encoding opal codon were totally randomized. The RNA pool was selected for its ability to bind to SelB in vitro. The inclusion of wild-type hairpin as a specific competitor helped to ensure that the best binding species were extracted from the population. The resultant anti-SelB aptamers fell into two groups. The first was composed of minor variations of the wild-type sequence. The 16 nucleotides at the top of the hairpin were completely conserved, with the exception of two adjacent base pair interactions close to the loop that covaried between several Watson–Crick alternatives. None of the wild-type-like aptamers had a UGA codon adjacent to the stem, suggesting that the VGA codon does not directly interact with SelB. The second group of aptamers contained widely divergent sequences that nonetheless still could be folded to form stem–loop structures. Most of the alternative stem–loops bound at the same site on SelB as the original hairpin, but two of the novel structures also appeared to interact with additional epitopes on SelB. Protection studies indicated that the apical regions of both classes of aptamer hairpins were important for protein recognition.
The finding that alternative sequences and structures can bind protein targets is far from unique. For example, in one of the first applications of in vitro selection methods to a protein target, a translational operator that bound T4 DNA polymerase was randomized, binding sequences were selected, and both the wild-type and an alternative stem–loop were recovered (8). The unnatural variant bound as well as the wild-type, indicating that nature apparently had found one of two equivalent “best answers.” Nonetheless, the three-dimensional structures of the two different RNAs were virtually identical (17, 18). Similarly, a region of 16S ribosomal RNA bound by protein S8 was completely randomized and binding variants were selected (19). Three families of related sequences were found: one family was similar to the natural consensus sequence whereas the other two families deviated significantly from this sequence. However, comparison of these families suggested that they could all form a base triplex with a looped-out A residue. As a final example, Olke Uhlenbeck and coworkers (20) partially randomized tRNAPhe and selected variants that could fulfill a number of independent, functional requirements, including the ability to (i) bind to phenylalanyl-tRNA synthetase, (ii) be aminoacylated, and (iii) bind in aminoacylated form to EF-Tu. Again, the selected variants showed much greater sequence diversity than had previously been observed in nature.
Although unnatural RNA sequences may bind as well as natural sequences, it is unclear whether they can serve as functional replacements for the natural sequences. It is possible that natural evolution never arrived at these alternative solutions because they were incompatible with other functions of the RNA. For example, although variants of tRNA might be aminoacylated and carried by EF-Tu, there is no guarantee that they would fit into the A or P sites of the ribosome. In this respect, the paper by Klug et al. (9) is particularly instructive. Several of the anti-SelB aptamers were cloned into a portion of the fdhF mRNA in place of the wild-type hairpin structure; care was taken to retain spacing requirements that previously had been observed to be important for hairpin function. The chimeras were then assayed for their ability to readthrough the opal codon by facilitating the insertion of selenocysteine. An aptamer from their first group, in which the apical region was identical to the wild type except for one base pair, was found to substitute for the wild-type stem–loop with 87% efficiency. However, none of the aptamers from the second group were able to efficiently substitute despite the fact that they formed hairpin structures of similar size and contained at least some residues that were similar to those of the wild type. One of the aptamers was able to partially substitute for the wild-type hairpin (17% of wild-type activity) when only its apical region was switched. The authors concluded that some aspect of the wild-type sequence and structure beyond SelB binding was essential for selenocysteine insertion. These results can be directly compared with a study in which anti-Rev aptamers were inserted into the Rev-responsive element in place of the wild-type Rev-binding element and assayed for the ability to facilitate Rev-dependent mRNA transport (21). As was the case with the studies performed by Klug et al. (9), one family of aptamers closely resembled the wild-type sequence, whereas the other diverged significantly. Tellingly, all could be modeled to resemble the wild-type, three-dimensional shape (22). However, in contrast to the results obtained by Klug et al. (9), both families of aptamers could efficiently replace the function of the wild-type element. Moreover, when the aptamers were expressed in trans they could inhibit interaction of the Rev protein with the wild-type element and concomitantly inhibit viral replication (23). Thus, it would appear that in some instances wild-type sequences are functionally optimal, and in other cases wild-type sequences can be readily replaced by alternative sequences. The structural corollary to this conclusion may be that some RNA-binding sequences, such as the Rev-binding element, are like DNA enhancers and can function in an architecture-independent manner, whereas other RNA-binding sequences, such as the hairpin that binds SelB, function in an architecture-dependent manner. Given the model shown in Fig. 1, it is not surprising that the latter system may require precise placement of the SelB:aminoacyl tRNA complex relative to the opal codon, or that the act of binding the hairpin may create a conformational shift in SelB, causing it to more effectively place the selenocysteyl-tRNASec in the ribosome.
However, there is an important caveat to this conclusion. In both experiments, the observation and quantitation of function may be highly dependent on assay conditions. Although the anti-Rev aptamers bound from 3- to 10-fold better than Rev in vitro, they initially appeared to facilitate mRNA transport no better than the wild-type Rev-binding element in vivo. It was hypothesized that this apparent discrepancy might be due to saturation of both wild-type and aptamer sequences with Rev. Accordingly, when the same constructs were expressed in the presence of subsaturating concentrations of Rev, the aptamers showed improved mRNA transport activity. Similarly, although anti-SelB aptamers that diverged from the wild-type hairpin sequence and structure do not appear to support selenocysteine incorporation, it should be noted that the effects of aptamer sequence and structure on overall translational efficiency were never measured. However, this is probably not a major consideration, as all the aptamers form stems with roughly equivalent stability, and it has been shown that lengthening the bottom stem by three base pairs has minimal effect on translational readthrough (much of it probably due to the greater distance between the opal codon and the hairpin apex) (16). Thus, depending on assay conditions and interpretations, unnatural sequences may function worse than, as well as, or better than wild type. True comparisons must await evolutionary competitions between organisms that contain unnatural and natural binding sequences.
References
- 1.Gould S J. Wonderful Life: The Burgess Shale and the Nature of History. New York: W. W. Norton; 1989. [Google Scholar]
- 2.Mills D R, Peterson R L, Spiegelman S. Proc Natl Acad Sci USA. 1967;58:217–224. doi: 10.1073/pnas.58.1.217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Biebricher C K, Eigen M, Gardiner W C., Jr Biochemistry. 1983;22:2544–2559. doi: 10.1021/bi00279a036. [DOI] [PubMed] [Google Scholar]
- 4.Conrad R C, Giver L, Tian Y, Ellington A D. Methods Enzymol. 1996;267:336–367. doi: 10.1016/s0076-6879(96)67022-0. [DOI] [PubMed] [Google Scholar]
- 5.Gold L. J Biol Chem. 1995;270:13581–13584. doi: 10.1074/jbc.270.23.13581. [DOI] [PubMed] [Google Scholar]
- 6.Robertson D L, Joyce G F. Nature (London) 1990;344:467–468. doi: 10.1038/344467a0. [DOI] [PubMed] [Google Scholar]
- 7.Ellington A D, Szostak J. Nature (London) 1990;346:818–822. doi: 10.1038/346818a0. [DOI] [PubMed] [Google Scholar]
- 8.Tuerk C, Gold L. Science. 1990;249:505–510. doi: 10.1126/science.2200121. [DOI] [PubMed] [Google Scholar]
- 9.Klug S J, Hüttenhofer A, Kromayer M, Famulok M. Proc Natl Acad Sci USA. 1997;94:6676–6681. doi: 10.1073/pnas.94.13.6676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zinoni F, Heider J, Bock A. Proc Natl Acad Sci USA. 1990;87:4660–4664. doi: 10.1073/pnas.87.12.4660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Forchhammer K, Leinfelder W, Bock A. Nature (London) 1989;342:453–456. doi: 10.1038/342453a0. [DOI] [PubMed] [Google Scholar]
- 12.Kromayer M, Wilting R, Tormay P, Bock A. J Mol Biol. 1996;262:413–420. doi: 10.1006/jmbi.1996.0525. [DOI] [PubMed] [Google Scholar]
- 13.Hüttenhofer A, Westhof E, Bock A. RNA. 1996;2:354–366. [PMC free article] [PubMed] [Google Scholar]
- 14.Baron C, Heider J, Bock A. Proc Natl Acad Sci USA. 1993;90:4181–4185. doi: 10.1073/pnas.90.9.4181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ringquist S, Schneider D, Gibson T, Baron C, Bock A, Gold L. Genes Dev. 1994;8:376–385. doi: 10.1101/gad.8.3.376. [DOI] [PubMed] [Google Scholar]
- 16.Heider J, Baron C, Bock A. EMBO J. 1992;11:3759–3766. doi: 10.1002/j.1460-2075.1992.tb05461.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mirmira S R, Tinoco I. Biochemistry. 1996;35:7664–7674. doi: 10.1021/bi960414y. [DOI] [PubMed] [Google Scholar]
- 18.Mirmira S R, Tinoco I. Biochemistry. 1996;35:7675–7683. doi: 10.1021/bi960415q. [DOI] [PubMed] [Google Scholar]
- 19.Moine H, Cachia C, Westhof E, Ehresmann B, Ehresmann C. RNA. 1997;3:255–268. [PMC free article] [PubMed] [Google Scholar]
- 20.Peterson E T, Blank J, Sprinzl M, Uhlenbeck O C. EMBO J. 1993;12:2959–2967. doi: 10.1002/j.1460-2075.1993.tb05958.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Symensma T L, Giver L, Zapp M, Takle G B, Ellington A D. J Virol. 1996;70:179–187. doi: 10.1128/jvi.70.1.179-187.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Leclerc F, Cedergren R, Ellington A D. Nat Struct Biol. 1994;1:293–300. doi: 10.1038/nsb0594-293. [DOI] [PubMed] [Google Scholar]
- 23.Good P D, Krikos A J, Li S X, Bertrand E, Lee N S, Giver L, Ellington A, Zaia J A, Rossi J J, Engelke D R. Gene Ther. 1997;4:45–54. doi: 10.1038/sj.gt.3300354. [DOI] [PubMed] [Google Scholar]