

Abstract
The challenge hypothesis posits that acute increases in testosterone (T) during male–male competition enhance performance and survivability while limiting the physiological costs of consistently high T. Human challenge hypothesis research focuses on young men in industrial populations, who have higher baseline T levels than men in subsistence populations. We tested whether the Tsimane, pathogenically stressed forager-horticulturalists of the Bolivian Amazon, would express acute T increases in response to physical competition. Saliva was collected from 88 Tsimane men (aged 16–59 years) before and after a competitive soccer match. Tsimane men had significantly lower baseline levels of T (β = −0.41, p < 0.001) when compared with age-matched United States (US) males. Linear mixed-effects models were used to establish that T increased significantly immediately following competition (β = 0.23, p < 0.001), remaining high 1 h later (β = 0.09, p = 0.007); equivalent to 30.1 and 15.5 per cent increases in T, respectively. We did not find larger increases in T among winners (p = 0.412), although T increases were positively associated with self-rated performance (β = 9.07, p = 0.004). These results suggest that despite lower levels of T than US males, Tsimane males exhibit acute increases in T at the same relative magnitude reported by studies in industrialized settings, with larger increases in T for those who report better individual performance.
Keywords: challenge hypothesis, testosterone, competition, Tsimane
1. Introduction
During male–male physical competition, acute spikes in testosterone (T) are observed in a variety of species across numerous taxa [1], including humans [2] and other primates [3]. Acute increases in T have a multitude of effects on male behaviour and physiology [4], including enhanced muscular performance [5,6] and immunoredistribution of leucocytes to the periphery to prepare for potential injury [7]. The challenge hypothesis [4] postulates that the energetic and immunocompromising costs of T [8,9] have led to the evolution of a system, where T is low throughout most of the year, when males focus on parenting or survival, spiking only when necessary to mediate the trade-off between reproduction and survival in social situations, including sexual activity, resource defence, status competition and competition over mates [4].
In industrialized human societies, acute increases in male T during competition have been reported in a variety of contexts from judo to chess [2]. The magnitude of T increase in response to a challenge tends to be largest during physically competitive sports, owing to energetic mobilization during physical activity [2], which limits the interpretation of the results, confounding physical activity and psychological competition. Competitions where social status is at stake tend to have larger increases in T [10]. Evidence for a victory-induced T-response is mixed, with some studies reporting larger increases in T among winners [2,11–14], while other studies find no evidence of a T-related winner effect [10,15–18]. It has also been reported that players with larger post-game T increases tend to rate their importance in the outcome of the game as higher [19].
Industrialized populations, relatively free of energetic constraints and parasitic or pathogenic stress show elevated levels of T across all adult ages when compared with non-industrialized populations [20–22]. In advantageous energetic environments, males pay reduced marginal costs to put on a display [23,24], and thus can invest more energy in exaggerated T-based signals [9]. However, individuals in energetically limited environments face a more restrictive trade-off between investing in survival, or reproductively advantageous but immunologically and metabolically costly T [8,9]. Muscle tissue is calorically expensive to develop and maintain, particularly given that T increases fat catabolism [25]. In healthy industrialized males, reduced energetic availability from increased energy expenditure [26], or fasting [27] appears to reduce T through downregulation of the hypothalamic-pituitary-gonadal axis. Parasitic and pathogenic stress [8,28,29] also downregulate T.
Do the same transient increases in T occur during male–male competition in chronically energetically and pathogenically stressed populations? The Tsimane forager-horticulturalists of the Bolivian Amazon are an ideal population for testing T dynamics in energetically stressful environments. A wealth of demographic, behavioural and health-related data have documented their high-energy expenditure and a relatively high parasite and pathogen load [30,31]. More than 15 000 Tsimane live in 80 villages of 30–500 people in the lowlands, east of the Andes [30,32]. Medical exams indicate that at least two-thirds of adults present with signs of intestinal parasites, and one-third of all adults examined showed signs of respiratory illness [31]. Energetic expenditure for Tsimane men and women aged 40–49 is estimated to be 450–850 kcal d−1 greater than an aged-matched United States (US) sample, and the prevalence of obesity is eight times less common among the Tsimane [31]. The Tsimane do not have a recent history of inter-group violent aggression, but intra-village disputes arising from competition over horticultural land, sale of village lumber, sexual jealousy or accusations of theft are not uncommon. Men's ability to get their way in a group or influence others during community meetings is largely related to coalitional support, while physical size better predicts dyadic fight outcomes [33].
2. Predictions
Regardless of basal T or condition, males engaged in physically competitive situations would benefit from acute increases in T, which rapidly enhance muscle performance and redistributes immune resources to prepare for injury. We hypothesize that despite pathogenic and parasitic stress resulting in lower levels of baseline T, Tsimane males will express a relative increase in T during competition similar to males in industrialized populations. We also expect to find larger increases in T among winners, as increased post-game T could augment future competitive ability. Positive associations between self-rated performance and increases in T are also hypothesized, as they may represent a personal victory, independent of team success.
3. Methods
In August 2010, teams from eight Tsimane communities competed in one of two inter-village soccer tournaments held by the researchers. Soccer is a common form of recreation among the Tsimane; men in these communities reported playing an average of 3.1 (s.d. = 1.5) days per week. The majority of games occur within communities with little or no audience present. On weekends, some neighbouring communities play against one another, frequently in the presence of audiences. Our tournaments brought distant villages together, with large mixed-sex audiences. Teams from each community were given uniforms, and the winning teams received trophies, creating a competitive atmosphere. Eighty-eight male participants provided saliva specimens via passive drool 15 min before, 10 min after and 1 h after their first soccer match. All games started at 11.00 h or 12.00 h to control for diurnal variation in T [22]. Following the match, players were separated from their team-mates and asked to identify the best and worst players in their team, and then how well they played in comparison with a five-point ladder scale. Height, weight and age were collected from participants for use as control variables, with age ranging from 16 to 59 years (mean age 26.8; table 1). Anthropometric measurements were not available for one individual; his body mass index (BMI) was coded as missing. Specimens were frozen in liquid nitrogen until transferred to dry ice and flown to the University of Washington where they were stored for four months at −80°C. Previous reports indicate that salivary T is stable for at least 36 months when stored at −80°C [34].
Table 1.
Descriptive statistics for 82 Tsimane males including (a) age and BMI, (b) testosterone before and after a soccer match, and (c) per cent change in testosterone.
| mean | s.e. | min | max | 95% CI | ||
|---|---|---|---|---|---|---|
| (a) | age (years) | 26.8 | 0.99 | 16 | 59 | [24.8, 28.7] |
| BMI (kg m−2) | 23.7 | 0.29 | 16.7 | 29.7 | [23.1, 24.3] | |
| (b) | T 15 min pre-game (pg ml−1) | 182.9 | 8.49 | 31.3 | 397.0 | [166.0, 199.8] |
| T 10 min post game (pg ml−1) | 231.3 | 10.66 | 50.7 | 487.8 | [210.1, 252.5] | |
| T 60 min post game (pg ml−1) | 202.0 | 9.77 | 72.9 | 461.9 | [182.6, 221.5] | |
| (c) | % change in T 10 min post | 30.1 | 4.05 | −48.4 | 193.9 | [22.0, 38.2] |
| % change in T 60 min post | 15.5 | 6.70 | −54.0 | 212.3 | [2.2, 28.9] |
Specimens were thawed and centrifuged at 2800 r.p.m. for 20 min, and the aqueous layer aliquoted for assay. All specimens had gone through two freeze–thaw cycles when initially assayed. Saliva exhibiting blood contamination (n = 22 specimens) [34], or inadequate volume (n = 5 specimens) were excluded, leaving a total of 237 usable specimens from 82 individuals. Salivary T was measured via an in-house enzyme immunoassay [27] that cross-reacts 100 per cent with T, 57.4 per cent with 5-α-dihydrotestosterone, 0.27 per cent with androstenodione and less than 0.05 per cent with other androgens [35]. Specimens were run in duplicate, with each participant's samples run on the same plate to reduce bias owing to inter-plate variation. Individuals were randomized between plates to ensure that teams were not placed together on the same plate. The within- and between-assay coefficients of variation for this project (n = 24 plates) were 4.5 and 9.3 per cent, respectively, for the low (403.1 pg ml−1) and 5.5% and 11.4 per cent for the high (681.4 pg ml−1) in-house controls.
(a). United States-Tsimane salivary testosterone comparison methods
US male salivary T specimens (n = 40 specimens) were collected as a part of a separate, non-competitive experiment [27]. Twenty-three healthy weighing men (mean BMI 23.7 kg m−2) aged 19–36 years (mean age 26.7) provided saliva specimens at 11.00 h on two consecutive control days, while undergoing no experimental manipulations. These US data, analysed in the same laboratory using the same enzyme immunoassay [35], will be directly compared with Tsimane pre-competition (baseline) salivary T levels to examine differences in basal T.
(b). Statistical methods
Salivary T concentrations were normalized with a log-transformation. To control for the non-independence of repeated specimens from each participant, linear mixed-effects regressions with individuals modelled as a random effect [36] were used to determine the effect of competition on log-transformed salivary T 10 min and 1 h after a soccer match. Adipose tissue can metabolize T into oestrogen [25], so BMI was included as a control variable in all models. Study design controlled the time of day at which specimens were collected; wakeup times were reported to be invariant; thus, time of specimen collection was not included as a control variable. In the Tsimane competition data, there were no association between T and age (p = 0.805), or T change from before to immediately following the game and age (p = 0.371), thus age was not included as a control variable. The effects of age may be nonlinear; additional analyses modelling age as quadratic or dummy variables (dividing age into quartiles and decades), find no association between age and T or change in T. Analyses were conducted in STATA 12.1 (College Station, TX, USA).
4. Results
(a). Tsimane–United States salivary testosterone comparison
Tsimane pre-competition saliva specimens (n = 82) were compared with age-matched US saliva specimens (n = 40) collected as controls during a non-competition study [27] to establish differences in baseline T levels. Tsimane male salivary T (182.9 pg ml−1, s.d. = 76.0) was significantly lower (β = −0.411, p < 0.001) when compared with US males (266.8 pg ml−1, s.d. = 90.2), controlling for age and BMI. These results were not owing to a wider age range of Tsimane participants—a secondary analysis limiting the age range of Tsimane participants to match US participants also found significantly lower salivary T (β = −0.409, p < 0.001) among Tsimane men.
(b). Tsimane soccer competition
T in the Tsimane soccer players was significantly higher after the game when compared with baseline levels (β = 0.23, p < 0.001) and remained high 1 h after the game (β = 0.08, p = 0.009; figure 1). The mean per cent change in T from 15 min before the game to 10 min after the game was 30.1 per cent, and from before to 1 h after the game was 15.5 per cent (table 1). Winning was neither associated with T (p = 0.558), nor per cent change in T (p = 0.450). Men with higher baseline T had smaller relative increases in T immediately following the game (β = −32.85, p < 0.001) and 1 h after the game (β = −46.41, p < 0.001).
Figure 1.
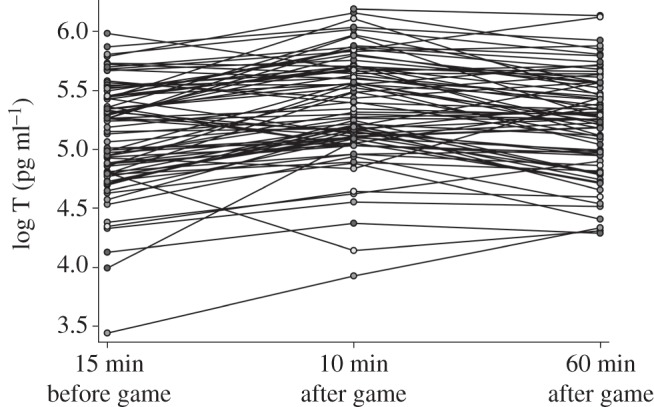
Individual log-salivary testosterone (T) 15 min before, 10 min after and 60 min after a soccer match for 82 Tsimane men.
Additional linear mixed-effects models with individuals and teams coded as random effects established that higher self-rated performance was associated with larger per cent increases in T from before the game to immediately following the game (β = 8.90, p = 0.005), but not 1 h after the game (p = 0.201; figure 2). Baseline T was not associated with self-rated performance (p = 0.493). Eleven goals were scored by six players; those who scored more goals trended towards larger increase in T 10 min after the game (β = 16.79, p = 0.113), and 1 h after the game (β = 32.74, p = 0.006).
Figure 2.

Individual change in salivary testosterone (T) 10 min post-game and self-rated performance for 82 Tsimane men. Filled circles, positive per cent change in T; open circles, negative per cent change in T.
No single position (forward, midfielder, defender, goalkeeper) had a statistically different T response when compared with other positions. When collapsed into defensive (defenders and goalkeepers) and offensive players (forwards and midfielders), defensive players trended towards smaller increases in T when compared with the offensive players (β = −15.92, p = 0.054), controlling for BMI. However, 11 participants reported switching between defensive and offensive positions during the game; excluding these individuals attenuated the trend (β = −14.50, p = 0.106). A logistic regression found that defensive players tended to report poorer self-rated performance than offensive players (β = −0.91, p < 0.001). Analysis examining the association between relative change in T and position was re-run with self-rated performance as a control variable, finding no association (p = 0.497).
5. Discussion
The Tsimane men in this sample had significantly lower baseline (pre-competition) levels of salivary T when compared with age-matched US males, controlling for age and BMI. These results replicate the finding that industrialized populations sheltered from many energetic and pathogenic stresses have significantly higher levels of T than men in non-industrialized settings [20,21,37–39]. Previous studies find evidence of a pre-competition rise in T [11,13,18,40], thus our baseline Tsimane T measure may be artificially inflated, suggesting that basal Tsimane T may be even lower.
Age was not associated with basal Tsimane T levels, or changes in T following competition. These results replicate those of previous studies suggesting that the typical pattern of decreasing T across the lifespan in industrialized populations [41] may not be generalizable to forager-horticulturalist populations [20,21]. Most [2], though not all [13], studies of acute changes in T during competition are conducted in young males, thus little is known about competition-related changes in T in older males. A US study reports that age did not affect the magnitude of increase in salivary T from men exposed to sexual stimuli [42], raising the possibility that acute changes in T owing to behavioural stimuli may occur independent of age.
Despite lower baseline T, Tsimane males expressed significant increases in T during physical competition. The average increase in T from before the game to immediately following the game was 30.1 per cent. A review of 12 physical competition-related studies in industrialized populations found an average T increase of 37 per cent, with a confidence interval ranging from a 13–61% increase in T [2]. The Tsimane T increase is of a comparable relative change to that observed in industrialized settings, despite lower absolute T levels. Studies of human male–male physical competition have focused on industrialized populations with high, perhaps, evolutionarily novel levels of T; this study extends these findings to non-industrialized populations. Caution must be taken when interpreting changes in T as purely owing to competition, which omits changes in T owing to physical activity; previous studies suggest that physically demanding competitions result in a larger increase in T than non-physical competitions [2].
Acute, short-term increases in T are beneficial regardless of current environmental condition, enhancing immediate muscle performance and redistributing leucocytes to peripheral tissues in preparation for injury. Previous studies find that poor environmental conditions or illness can reduce T-mediated reproductive or competitive behaviour in some avian species [43,44]. In these studies, lower T was associated with avoiding territorial behaviour that could induce a fight; thus facilitating avoidance of dangerous conflicts when an individual is in poor condition. However, these studies examined the role of baseline T on behaviour, and did not measure acute changes in T during competition. Once engaged in a physical confrontation, enhanced muscle physiology and immunoredistribution would be advantageous regardless of individual condition, in order to facilitate fighting ability or escape.
Men with higher baseline T had decreased relative change in T when compared to those with lower baseline T. This is interpreted as a function of the pre-competition rise in T [11,13,18,40]; some men may have had a pre-competition spike in T prior to their first saliva specimen, while others had a spike in T at the onset of the game.
When collapsed into defensive and offensive positions, there was a trend towards offensive players presenting a larger relative increase in T. When controlling for self-rated performance, there ceased to be an association between relative change in T and field position type, suggesting that change in T is not simply owing to increased physical activity among offensive players, and is probably dependent on personal investment, regardless of position.
There was no evidence of a winner effect in this sample. Previous reports find evidence for [2,11–14], and against [10,15–18] winner effects. Some variation in winner effects may be owing to the circumstances in which competitions are held; men playing against ingroups tend to exhibit a reduced T response to winning when compared with those competing against outgroups [45,46]. In this case, men were competing for their village, against other villages, in a high-status event; a situation normally associated with a winner effect [2,45]. However, Tsimane communities are relatively informal and fluid; migration between communities is not uncommon. The more salient ingroup could have been family members or friends who were playing against each other. This could suggest that some of the men were competing against ingroups—a situation which reduces a T-related winner effect [45,46]. We did not collect data beforehand on who was expected to win each match—anecdotal evidence suggests that the winning teams of each tournament were reliably predicted before the matches, which could have reduced individual investment in the game [15].
Better individual performance, as measured by either self-rating or goals scored, was associated with a larger relative change in T following the game. These results replicate those of previous projects, which suggest that higher self-rated performance [19,45], and individual investment in the game [15] increases T response to competition. While overall changes in T confound physical activity and psychological competition, individual level performance may indicate personal investment in competition. Individual performance may be tantamount to personal victory, regardless of team success. Better performance, even if a member of a losing coalition, signals individual ability within and between coalitions, and to audience members. It should be noted that the challenge hypothesis was designed to examine changes in T in a bird model, focusing on dyadic conflicts. Human tests involving sports competitions are mediated by coalitional membership; thus in human studies, it may be important to examine individual and coalitional success.
(a). Limitations
These tournaments were high-profile events; it is not clear whether these results could be extended to normal within-community games. A major limitation of this study is that men playing in the soccer tournament were probably self-selected to be the healthiest and most competitive young Tsimane men. A one-sided t-test suggests that tournament participants who had a mean BMI of 23.7 kg m−2 were not statistically different from the average BMI of 23.5 kg m−2 for Tsimane men aged 16–59 years (n = 728 men measured since 2008). Despite self-selection, these men still presented with significantly lower levels of T when compared with age-matched US males. While this study has a larger age range than many previous studies, it offers limited insight into age-related acute T changes among older men.
It should be noted that all studies examining T dynamics during competitive sports confound changes in T owing to physical activity and psychological factors associated with competition. To disassociate these differences, additional studies (to be reported elsewhere) were conducted to examine changes in Tsimane T during non-competitive physical activity, and during competitive non-physical activity. Beyond differentiating between physical and non-physical competition, these studies also allowed us to examine changes in T: (i) in conjunction with individual measures of health status, (ii) in older men, and (iii) in the presence of different audiences.
6. Conclusion
In summary, these results suggest that the Tsimane, who have significantly lower levels of baseline T when compared with US males, exhibit an acute increase in T during physical competition. There was no evidence of increased T for winning teams, though individuals who performed better expressed larger increases in T, perhaps underscoring the importance of individual as well as coalitional success. While previous studies suggest that environmentally mediated T may decrease territorial or competitive behaviour, we propose that once engaged in physical competition, acute increases in T are physiologically beneficial regardless of individual condition.
Acknowledgements
All procedures were approved by the University of Washington Internal Review Board.
We would like to thank the participants, Emily Miner, Jeff Winking, Eleanor Brindle, Amanda Guyton, Alberto Cari, Bacillio Vie, Arnulfo Cari, Vincente Cuata, Meliton Lero, the Tsimane Health and Life History Project Staff, and two anonymous reviewers. Funded in part by NICHD R24HD042828, 5T32HD007543, NIA R01AG024119-01, R56AG024119-06 and R01AG024119-07.
References
- 1.Hirschenhauser K., Oliveira R. F. 2006. Social modulation of androgens in male vertebrates: meta-analyses of the challenge hypothesis. Anim. Behav. 71, 265–277 10.1016/j.anbehav.2005.04.014 (doi:10.1016/j.anbehav.2005.04.014) [DOI] [Google Scholar]
- 2.Archer J. 2006. Testosterone and human aggression: an evaluation of the challenge hypothesis. Neurosci. Biobehav. Rev. 30, 319–345 10.1016/j.neubiorev.2004.12.007 (doi:10.1016/j.neubiorev.2004.12.007) [DOI] [PubMed] [Google Scholar]
- 3.Muller M. N., Wrangham R. W. 2004. Dominance, aggression and testosterone in wild chimpanzees: a test of the ‘challenge hypothesis’. Anim. Behav. 67, 113–123 10.1016/j.anbehav.2003.03.01 (doi:10.1016/j.anbehav.2003.03.01) [DOI] [Google Scholar]
- 4.Wingfield J. C., Hegner R. E., Dufty J., Alfred M., Ball G. F. 1990. The ‘challenge hypothesis’: theoretical implications for patterns of testosterone secretion, mating systems, and breeding strategies. Am. Nat. 136, 829–846 10.1086/285134 (doi:10.1086/285134) [DOI] [Google Scholar]
- 5.Crewther B. T., Cook C., Cardinale M., Weatherby R. P., Lowe T. 2011. Two emerging concepts for elite athletes: the short-term effects of testosterone and cortisol on the neuromuscular system and the dose-response training role of these endogenous hormones. Sports Med. 41 103–123 10.2165/11539170 (doi:10.2165/11539170) [DOI] [PubMed] [Google Scholar]
- 6.Tsai L. W., Sapolsky R. M. 1996. Rapid stimulatory effects of testosterone upon myotubule metabolism and sugar transport, as assessed by silicon microphysiometry. Aggressive Behav. 22, 357–364 10.1002/(sici)1098-2337 (doi:10.1002/(sici)1098-2337) [DOI] [Google Scholar]
- 7.Braude S., Tang-Martinez Z., Taylor G. T. 1999. Stress, testosterone, and the immunoredistribution hypothesis. Behav. Ecol. 10, 345–350 10.1093/beheco/10.3.345 (doi:10.1093/beheco/10.3.345) [DOI] [Google Scholar]
- 8.Muehlenbein M. P., Bribiescas R. G. 2005. Testosterone-mediated immune functions and male life histories. Am. J. Hum. Biol. 17, 527–558 10.1002/ajhb.20419 (doi:10.1002/ajhb.20419) [DOI] [PubMed] [Google Scholar]
- 9.Folstad I., Karter A. J. 1992. Parasites, bright males, and the immunocompetence handicap. Am. Nat. 139, 603–622 10.1086/285346 (doi:10.1086/285346) [DOI] [Google Scholar]
- 10.Mehta P. H., Josephs R. A. 2006. Testosterone change after losing predicts the decision to compete again. Horm. Behav. 50, 684–692 10.1016/j.yhbeh.2006.07.001 (doi:10.1016/j.yhbeh.2006.07.001) [DOI] [PubMed] [Google Scholar]
- 11.Booth A., Shelley G., Mazur A., Tharp G., Kittok R. 1989. Testosterone, and winning and losing in human competition. Horm. Behav. 23, 556–571 10.1016/0018-506X(89)90042-1 (doi:10.1016/0018-506X(89)90042-1) [DOI] [PubMed] [Google Scholar]
- 12.Elias M. 1981. Serum cortisol, testosterone, and testosterone-binding globulin responses to competitive fighting in human males. Aggressive Behav. 7, 215–224 10.1002/1098-2337 (doi:10.1002/1098-2337) [DOI] [Google Scholar]
- 13.Mazur A., Booth A., Dabbs J., Jr 1992. Testosterone and chess competition. Soc. Psychol. Q. 55, 70–77 10.2307/2786687 (doi:10.2307/2786687) [DOI] [Google Scholar]
- 14.Gladue B. A., Boechler M., McCaul K. D. 1989. Hormonal response to competition in human males. Aggressive Behav. 15, 409–422 10.1002/1098-2337 (doi:10.1002/1098-2337) [DOI] [Google Scholar]
- 15.Gonzalez-Bono E., Salvador A., Serrano M. A., Ricarte J. 1999. Testosterone, cortisol, and mood in a sports team competition. Horm. Behav. 35, 55–62 10.1006/hbeh.1998.1496 (doi:10.1006/hbeh.1998.1496) [DOI] [PubMed] [Google Scholar]
- 16.Wagner J. D., Flinn M. V., England B. G. 2002. Hormonal response to competition among male coalitions. Evol. Hum. Behav. 23, 437–442 10.1016/s1090-5138(02)00100-9 (doi:10.1016/s1090-5138(02)00100-9) [DOI] [Google Scholar]
- 17.van der Meij L., Buunk A. P., van de Sande J. P., Salvador A. 2008. The presence of a woman increases testosterone in aggressive dominant men. Horm. Behav. 54, 640–644 10.1016/j.yhbeh.2008.07.001 (doi:10.1016/j.yhbeh.2008.07.001) [DOI] [PubMed] [Google Scholar]
- 18.Suay F., Salvador A., Gonzalez-Bono E., Sanchis C., Martinez M., Martinez-Sanchis S., Simon V. M., Montoro J. B. 1999. Effects of competition and its outcome on serum testosterone, cortisol and prolactin. Psychoneuroendocrinology 24, 551–566 10.1016/S0306-4530(99)00011-6 (doi:10.1016/S0306-4530(99)00011-6) [DOI] [PubMed] [Google Scholar]
- 19.Edwards D. A., Wetzel K., Wyner D. R. 2006. Intercollegiate soccer: saliva cortisol and testosterone are elevated during competition, and testosterone is related to status and social connectedness with team mates. Physiol. Behav. 87, 135–143 10.1016/j.physbeh.2005.09.007 (doi:10.1016/j.physbeh.2005.09.007) [DOI] [PubMed] [Google Scholar]
- 20.Ellison P. T., Bribiescas R. G., Bentley G. R., Campbell B. C., Lipson S. F., Panter-Brick C., Hill K. 2002. Population variation in age-related decline in male salivary testosterone. Hum. Reprod. 17, 3251–3253 10.1093/humrep/17.12.3251 (doi:10.1093/humrep/17.12.3251) [DOI] [PubMed] [Google Scholar]
- 21.Bribiescas R. 1996. Testosterone levels among Ache hunter–gatherer men. Hum. Nature 7, 163–188 10.1007/BF02692109 (doi:10.1007/BF02692109) [DOI] [PubMed] [Google Scholar]
- 22.Bribiescas R. G., Hill K. R. 2009. Circadian variation in salivary testosterone across age classes in Ache Amerindian males of Paraguay. Am. J. Hum. Biol. 22, 216–220 10.1002/ajhb.21012 (doi:10.1002/ajhb.21012) [DOI] [PubMed] [Google Scholar]
- 23.Bliege Bird R., Smith E. A. 2005. Signaling theory, strategic interaction, and symbolic capital. Curr. Anthropol. 46, 221–248 10.1086/427115 (doi:10.1086/427115) [DOI] [Google Scholar]
- 24.Zahavi A. 1975. Mate selection: a selection for a handicap. J. Theor. Biol. 53, 205–214 [DOI] [PubMed] [Google Scholar]
- 25.Bhasin S., et al. 2001. Testosterone dose–response relationships in healthy young men. Am. J. Physiol. Endocrinol. Metab. 281, E1172–E1181 [DOI] [PubMed] [Google Scholar]
- 26.Friedl K. E., Moore R. J., Hoyt R. W., Marchitelli L. J., Martinez-Lopez L. E., Askew E. W. 2000. Endocrine markers of semistarvation in healthy lean men in a multistressor environment. J. Appl. Physiol. 88, 1820–1830 [DOI] [PubMed] [Google Scholar]
- 27.Trumble B. C., Brindle E., Kupsik M., O'Connor K. A. 2010. Responsiveness of the reproductive axis to a single missed evening meal in young adult males. Am. J. Hum. Biol. 22, 775–781 10.1002/ajhb.21079 (doi:10.1002/ajhb.21079) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Muehlenbein M. P., Hirschtick J. L., Bonner J. Z., Swartz A. M. 2010. Toward quantifying the usage costs of human immunity: altered metabolic rates and hormone levels during acute immune activation in men. Am. J. Hum. Biol. 22, 546–556 10.1002/ajhb.21045 (doi:10.1002/ajhb.21045) [DOI] [PubMed] [Google Scholar]
- 29.Boonekamp J. J., Ros A. H., Verhulst S. 2008. Immune activation suppresses plasma testosterone level: a meta-analysis. Biol. Lett. 4, 741–744 10.1098/rsbl.2008.0347 (doi:10.1098/rsbl.2008.0347) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gurven M., Kaplan H., Supa A. Z. 2007. Mortality experience of Tsimane Amerindians of Bolivia: regional variation and temporal trends. Am. J. Hum. Biol. 19, 376–398 10.1002/ajhb.20600 (doi:10.1002/ajhb.20600) [DOI] [PubMed] [Google Scholar]
- 31.Gurven M., Kaplan H., Winking J., Rodriguez D., Vasunilashorn S., Kim J., Finch C., Crimmins E. 2009. Inflammation and infection do not promote arterial aging and cardiovascular disease risk factors among lean horticulturalists. PLoS ONE 4, e6590 (doi:10.1371/journal.pone.0006590) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Godoy R., Reyes-García V., Byron E., Leonard W., Vadez V. 2005. The effect of market economies on the well-being of indigenous peoples and on their use of renewable natural resources. Annu. Rev. Anthropol. 34, 121–138 10.1146/annurev.anthro.34.081804.120412 (doi:10.1146/annurev.anthro.34.081804.120412) [DOI] [Google Scholar]
- 33.von Rueden C., Gurven M., Kaplan H. 2008. The multiple dimensions of male social status in an Amazonian society. Evol. Hum. Behav. 29, 402–415 10.1016/j.evolhumbehav.2008.05.001 (doi:10.1016/j.evolhumbehav.2008.05.001) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Granger D. A., Shirtcliff E. A., Booth A., Kivlighan K. T., Schwartz E. B. 2004. The ‘trouble’ with salivary testosterone. Psychoneuroendocrinology 29, 1229–1240 10.1016/j.psyneuen.2004.02.005 (doi:10.1016/j.psyneuen.2004.02.005) [DOI] [PubMed] [Google Scholar]
- 35.Muir C., Spironello-Vella E., Pisani N., de Catanzaro D. 2001. Enzyme immunoassay of 17 beta-estradiol, estrone conjugates, and testosterone in urinary and fecal samples from male and female mice. Horm. Metab. Res. 33, 653–658 10.1055/s-2001-18692 (doi:10.1055/s-2001-18692) [DOI] [PubMed] [Google Scholar]
- 36.West B., Welch K. B., Galecki A. T. 2007. Linear mixed models: a practical guide using statistical software. Boca Raton, FL: Chapman & Hall/CRC [Google Scholar]
- 37.Campbell B., Leslie P., Campbell K. 2006. Age-related changes in testosterone and SHBG among Turkana males. Am. J. Hum. Biol. 18, 71–82 10.1002/ajhb.20468 (doi:10.1002/ajhb.20468) [DOI] [PubMed] [Google Scholar]
- 38.Ellison P. T., Lipson S. F., Meredith M. D. 1989. Salivary testosterone levels in males from the Ituri Forest of Zaïre. Am. J. Hum. Biol. 1, 21–24 10.1002/ajhb.1310010106 (doi:10.1002/ajhb.1310010106) [DOI] [PubMed] [Google Scholar]
- 39.Bentley G. R., Harrigan A. M., Campbell B., Ellison P. T. 1993. Seasonal effects on salivary testosterone levels among Lese males of the Ituri Forest, Zaire. Am. J. Hum. Biol. 5, 711–717 10.1002/ajhb.1310050614 (doi:10.1002/ajhb.1310050614) [DOI] [PubMed] [Google Scholar]
- 40.Salvador A., Suay F., Gonzalez-Bono E., Serrano M. A. 2003. Anticipatory cortisol, testosterone and psychological responses to judo competition in young men. Psychoneuroendocrinology 28, 364–375 10.1016/S0306-4530(02)00028-8 (doi:10.1016/S0306-4530(02)00028-8) [DOI] [PubMed] [Google Scholar]
- 41.Travison T. G., Araujo A. B., O'Donnell A. B., Kupelian V., McKinlay J. B. 2007. A population-level decline in serum testosterone levels in American men. J. Clin. Endocrinol. Metab. 92, 196–202 10.1210/jc.2006-1375 (doi:10.1210/jc.2006-1375) [DOI] [PubMed] [Google Scholar]
- 42.Escasa M. J., Casey J. F., Gray P. B. 2011. Salivary testosterone levels in men at a U.S. sex club. Arch. Sex Behav. 40, 921–926 10.1007/s10508-010-9711-3 (doi:10.1007/s10508-010-9711-3) [DOI] [PubMed] [Google Scholar]
- 43.Munoz N. E., Blumstein D. T., Foufopoulos J. 2010. Immune system activation affects song and territorial defense. Behav. Ecol. 21, 788–793 10.1093/beheco/arq054 (doi:10.1093/beheco/arq054) [DOI] [Google Scholar]
- 44.Neave N., Wolfson S. 2003. Testosterone, territoriality, and the ‘home advantage’. Physiol. Behav. 78, 269–275 10.1016/s0031-9384(02)00969-1 (doi:10.1016/s0031-9384(02)00969-1) [DOI] [PubMed] [Google Scholar]
- 45.Oxford J., Ponzi D., Geary D. C. 2009. Hormonal responses differ when playing violent video games against an ingroup and outgroup. Evol. Hum. Behav. 31, 201–209 10.1016/j.evolhumbehav.2009.07.002 (doi:10.1016/j.evolhumbehav.2009.07.002) [DOI] [Google Scholar]
- 46.Wagner J., Flinn M., England B. 2002. Hormonal response to competition among male coalitions. Evol. Hum. Behav. 23, 437–442 10.1016/S1090-5138(02)00100-9 (doi:10.1016/S1090-5138(02)00100-9) [DOI] [Google Scholar]