

Abstract
Studies in a multitude of models including cell culture, animal and clinical studies demonstrate that citrus-derived flavonoids have therapeutic potential to attenuate dyslipidemia, correct hyperinsulinemia and hyperglycemia, and reduce atherosclerosis. Emerging evidence suggests the metabolic regulators, PPARα and PGC1α, are targets of the citrus flavonoids, and their activation may be at least partially responsible for mediating their metabolic effects. Molecular studies will add significantly to the concept of these flavonoids as viable and promising therapeutic agents to treat the dysregulation of lipid homeostasis, metabolic disease, and its cardiovascular complications.
1. Introduction
Metabolic syndrome is a clustering of risk factors including dyslipidemia, hypertension, insulin resistance, and visceral obesity for the development of type 2 diabetes mellitus and premature atherosclerosis [1–3]. While pharmacological agents have been successfully used to target major risk factors including hypertension, plasma cholesterol, and blood sugar, data collected from 1994–2005 suggest that the prevalence of hypertension, diabetes and obesity and the number of people diagnosed with CVD are increasing in all groups of Canadians [4]. The increased risk was most dramatic in the youngest group studied (aged 12–34 years old). In light of the increasing prevalence of metabolic dysregulation, new therapies are required. In the search for new targets and interventions to improve underlying risk factors and prevent the complications of atherosclerosis, attention has turned to nontraditional therapies including a group of naturally-occurring compounds known as flavonoids.
2. Flavonoids and Metabolic Disease
Flavonoids are abundant in the human diet as several thousands have been identified in a large variety of beverages and foods [5]. They have been classified based on their structural variation and degree of oxidation [5]. Flavonoid intake is inversely associated with coronary heart disease as men in the highest tertile of flavonoid intake experience the lowest relative risk of mortality from coronary artery disease [6]. An inverse relationship between flavonoid consumption and many risk factors for heart disease [7] including lower blood pressure and improved flow mediated dilatation [8], improved weight management [9], and improved dyslipidemia [8, 10] has also been established, suggesting that flavonoids have multiple targets. In human studies, wide variability has been observed in the effects of flavonoid-rich foods on these markers of cardiovascular disease risk [8]. Understanding the specific targets responsible for risk reduction is difficult to delineate in such a complex system. Therefore, recent attention has turned to purified flavonoids and the examination of their pharmacological properties.
Many studies have estimated flavonoid intake, and while the family of flavan-3-ols including catechins found in tea and chocolate often make up the majority [11], the flavanones also contribute significantly to total daily intake [12]. The citrus flavanones hesperitin and naringenin are members of this family (reviewed in [13]). Naringin, the glycoside form of naringenin found in citrus fruits, is hydrolyzed by intestinal microflora to the flavanone naringenin. Making up a smaller percentage of total intake is the class of flavones which contains the polymethoxylated polyphenols nobiletin and tangeretin, two flavonoids found abundantly in oranges and tangerines [12]. Administration of grapefruit or orange juice to hypercholesterolemic casein-fed rabbits reduced low density lipoprotein cholesterol (LDL-C) and hepatic lipid accumulation suggesting components of citrus may have lipid lowering properties [10]. In rats with streptozotocin- (STZ-) induced diabetes, i.p. injection of naringenin decreased blood glucose and improved dyslipidemia [14]. These early data suggested that citrus-derived flavonoids may improve metabolic health in patients with insulin resistance and prevent diabetes and CVD.
While a direct relationship has not been clearly established, a common thread linking many of the features of the metabolic syndrome includes the aberrant tissue deposition of lipid including cholesterol, ceramides, and triglycerides [15]. The dyslipidemia of metabolic syndrome, which is characterized by an overproduction of hepatic very low density lipoproteins (VLDL-) and low high density lipoproteins (HDL-) cholesterol [16], is thought to be a significant contributor to enhanced tissue lipid deposition. Triglyceride (TG) formation is the primary method of metabolic fuel storage for most species [17] and the formation of TG can protect cells from fatty acid (FA) overload and subsequent lipotoxicity [15]. Ectopic lipid accumulation has been described in many tissues including the heart, arteries, liver, muscle, adipose tissue, and pancreas and has been linked to the dysfunction of these tissues in models of insulin resistance and metabolic syndrome [15, 18]. Therefore, one mechanism by which flavonoids have been described to prevent metabolic dysregulation is to limit ectopic lipid accumulation and stimulate fatty acid and glucose utilization.
3. PPARα-Mediated Regulation of Metabolism
Peroxisome proliferator-activated receptor alpha (PPARα) is a member of the family of nuclear hormone receptors which function as ligand activated transcription factors, with a signature type II zinc finger DNA binding motif, to control the expression of specific genes involved in fatty acid utilization [19, 20] (Figure 1). Liver, muscle, kidney, and brown adipose tissue express high levels of PPARα [20, 21]. The endogenous ligands for PPARα activation include fatty acids and their metabolites [22]. PPARs control expression of genes by partnering with RXR and binding to peroxisome proliferator response elements (PPREs) in the promoter of target genes, ultimately resulting in the stimulation of FA oxidation. In mice, receptor agonists cause proliferation of peroxisomes which serve to oxidize long chain fatty acids and detoxify xenobiotic compounds [23]. PPARα is an activator of genes involved in β-oxidation including carnitine palmitoyl transferase 1 (CPT1α) and acyl-CoA oxidase (ACOX). In the liver, fatty acid oxidation is primarily regulated by the level of CPT1 [24]. CPT1α expression is controlled by a complex of transcription factors including PPARγ-coactivator 1 α (PGC1α) and PPARα [24]. CPT1 is found on the outer surface of the mitochondria and transports fatty acids into the mitochondria by the formation of an acyl-carnitine molecule. Upon entry into the inner side of the mitochondrial membrane, CPT2 removes the carnitine, reforming acyl-CoA [24].
Figure 1.
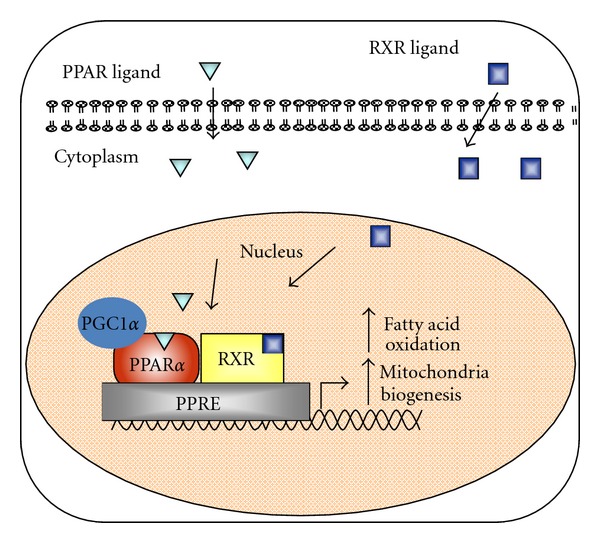
Regulation of gene expression by peroxisome proliferator-activated receptors. The nuclear hormone receptor PPARα induces transcription through formation of a heterodimer with the retinoic X receptor and binding to peroxisome proliferator response elements (most are direct repeats with one intervening nucleotide) in the promoter of genes involved in fatty acid oxidation. PGC1α is an important PPARα coactivator in tissues that undergo extensive oxidative metabolism and induce mitochondrial expansion.
Dynamic regulation of fatty acid oxidation is provided by interaction between acetyl-CoA carboxylase (ACC), a member of the fatty acid synthesis pathway, and CPT1, through the intermediate malonyl-CoA. When malonyl-CoA levels are high, they allosterically inhibit CPT1 and de novo lipogenesis continues. When the cell experiences a drop in malonyl-CoA, ACC is inhibited and fatty acid oxidation is stimulated [25, 26]. In liver-specific Fas−/− mice significant increases in malonyl-CoA were observed to inhibit CPT1α resulting in hepatic steatosis [27]. Recently, adenovirus-mediated hepatic expression of a permanently active mutant form of Cpt1α in high-fat fed obese mice, resulted in increased hepatic FA oxidation, thereby reducing hepatic TG content and ameliorating insulin resistance [28]. These studies highlight the importance of CPT1α-regulated fatty acid oxidation in maintaining lipid balance in the cell.
Many nuclear receptor cofactors exist which provide additional control of gene expression [29]. In the liver, coactivators important for induction of PPARα-mediated gene transcription and regulation of lipid metabolism include PGC1α. PGC1α regulates a number of hepatic metabolic pathways, including gluconeogenesis, mitochondrial expansion, and FA oxidation [30]. Once bound to a transcription factor, PGC1α allows the interaction of histone modifiers, the transcriptional initiation complex and DNA [31] (Figure 1). Similar to PPARα, hepatic PGC1α is induced by fasting, through a cAMP response element [32]. Mice with hepatic-specific knockout of Pgc1α accumulate liver TG, due to impaired β-oxidation, and enhanced SREBP1c-stimulated FA and TG synthesis [33, 34]. It is known that PGC1α increases genes involved in mitochondrial expansion through induction of the nuclear respiratory factors [31]. Furthermore, PGC1α interacts with and coactivates PPARα to transcriptionally regulate the genes involved in FA oxidation and increase palmitate utilization rates [29].
PPARα is most active in response to a drop in blood glucose, whether by fasting or by exercise [20]. In addition to replenishing ATP, the process of FA oxidation also provides the reducing cofactors required for gluconeogenesis and can induce the utilization of FA for the production of ketone bodies [35]. Mice with whole body deficiency of PPARα have decreased capacity for the oxidation FA including palmitic acid [36] and elevated total- and HDL-cholesterol levels [23], after a 48 h fast. Furthermore, Pparα−/− mice exhibit hypoglycemia, significant lipid accumulation in the liver, increased circulating nonesterified fatty acids, and decreased ketogenesis [37].
Pharmacological activators of PPARα, including the fibrate family of drugs, have demonstrated efficacy for reducing plasma TG and raising HDL in dyslipidemic patients and are often used in patients with insulin resistance. A recent meta-analysis of trials involving hypertriglyceridemic patients demonstrated that fibrates reduce vascular events [38], suggesting a renewed interest in the activation of PPARα to prevent CVD in select patients with dyslipidemia.
4. Regulation of Lipid Metabolism and Correction of Dyslipidemia by Citrus Flavonoids
Like fibrates, many citrus-derived flavonoids exhibit lipid-lowering properties. The consumption of 1-2 grapefruits/day significantly reduced cholesterol and triglyceride in hypercholesterolemic patients suffering from coronary artery disease [39, 40]. In a clinical trial involving hypercholesterolemic patients, the citrus flavonoid naringin (400 mg/day) was shown to reduce LDL-cholesterol by 17% and triglycerides by 14% [41]. However, in other recent human trials, capsules of hesperitin (800 mg/day) or naringin (500 mg/day) produced no lipid lowering benefit in moderately hypercholesterolemic individuals [42].
In more mechanistic studies, it was demonstrated in cultured hepatoma cells that naringenin reduces CE availability for lipoprotein assembly and secretion through inhibition of ACAT and MTP activities [43, 44]. However, subsequent studies established that the reduction in CE accumulation within the lumen of the endoplasmic reticulum (ER) did not mediate the naringenin-induced inhibition of apolipoprotein B100 (apoB100) assembly. Instead it was the reduction in TG accumulation within the ER lumen by inhibition of MTP that primarily facilitated the naringenin-mediated decrease in apoB secretion [43, 44]. In detailed pulse-chase studies, naringenin decreased apoB100 secretion from HepG2 cells even in the presence of oleate, through enhanced apoB degradation [44]. Studies in rat liver McARH7777 cells, transfected with constructs containing human apoB of various lengths, revealed that naringenin only inhibited the secretion of apoB-containing particles that required lipidation, whereas shorter length apoB constructs, which require minimal lipidation for secretion, were unaffected [43]. These in vitro findings suggest that naringenin limits TG availability for apoB secretion.
Metabolic studies in Ldlr−/− mice demonstrated that a western type diet stimulated the production rate of VLDL-TG and apoB100 when compared with chow-fed mice [45]. Supplementation of the western diet with 3% naringenin completely prevented the overproduction of apoB100 and TG, supporting the previous in vitro data that naringenin prevents VLDL-apoB100 secretion. The pattern of hepatic lipid accumulation mirrored TG secretion, as hepatic TG was increased 2-fold in western-fed animals compared to chow-fed mice and levels were significantly reduced by naringenin. Significant increases in the hepatic mRNA expression of Srebf1c (5-fold) and increased TG synthesis (1.3-fold) were related to the significant hyperinsulinemia induced by the western diet. These parameters were all normalized by 3% naringenin. In concert with the decrease in insulin-stimulated lipogenesis, naringenin stimulated hepatic expression of Cpt1α, Acox, and fatty acid β-oxidation to prevent hepatic TG accumulation [45]. In ICR rats fed 1% naringenin for 21 days, significantly increased hepatic levels of mRNA coding for enzymes involved in peroxisomal and omega fatty acid oxidation were observed [46]. Although a pharmacological dose was used in these studies, concentrations of dietary naringenin as low as 0.6% have been reported to lower hepatic triglyceride levels in rats fed a high-fat diet; however, these effects were not observed in rats fed a high carbohydrate diet [47].
In high-fat fed Ldlr−/− mice, addition of nobiletin (0.1% and 0.3% w/w) resulted in a dramatic reduction in both hepatic and intestinal TG accumulation, attenuation of VLDL-TG secretion, normalization of insulin sensitivity and conferred an almost complete resistance to obesity, without effect on caloric consumption or fat absorption [48]. These studies indicate that prevention of the hepatic lipid load by nobiletin limits the availability lipid for hepatic storage, lipoprotein secretion and deposition in peripheral tissues. Others have demonstrated in hamsters fed a semipurified diet that supplementation with mixtures of 1% polymethoxylated flavonoids, containing tangeretin and nobiletin, lowered plasma concentrations of both TG and cholesterol and reduced hepatic TG [49].
Similar to naringenin, addition of nobiletin to a western diet increased hepatic β-oxidation, contributing to reduced hepatic TG, decreased VLDL secretion and the correction of insulin resistance. Both naringenin and nobiletin increased hepatic Pgc1αmRNA, which coincided with increased Cpt1αmRNA, mitochondrial DNA and hepatic FA oxidation [45]. However, nobiletin did not increase mRNA of Acox [48]. Studies in male ICR rats fed a diet containing 1% naringenin for 21 days demonstrated increased mRNA levels of enzymes involved in peroxisomal fatty acid oxidation which resulted in a significant increase (+56%) in the rate of peroxisomal fatty acid oxidation compared with controls [50].
The molecular signals for the induction of FA oxidation in liver by naringenin or nobiletin have not been identified. In HepG2 cells, using concentrations of naringenin and nobiletin that inhibited apoB100 secretion by 50–70% (100 μM naringenin, 10 μM nobiletin), we observed upregulation of PPARα target genes including Cpt1α, but not activation of a luciferase promoter containing three PPRE elements suggesting these flavonoids do not function as direct PPARα activators [45, 48]. Furthermore, induction of PparαmRNA expression and increased liver weight, characteristic of PPARα activators in mice (fibrates), did not occur with naringenin [45] or nobiletin [48].
In contrast, naringenin has been reported to stimulate PPRE promoter activity in U-2OS cells, a human osteosarcoma cell line [51]. Also, in the human liver cell line Huh-7, naringenin, at relatively high doses, induced PPRE activity 17% at 150 μM and activated a PPARα-GAL4 fusion protein by 24% at 240 μM. These authors also observed increased transcription of enzymes involved in fatty acid utilization including Pgc1α, Acox, and Ucp1 [52]. In a cell free system, naringenin did not increase the binding of PGC1α to PPARα but did significantly increase Pgc1α mRNA by 14-fold in the Huh7 hepatocyte cell line [52]. These data suggest that at high concentrations, these flavonoids may directly activate PPARα. However, in human studies oral administration of naringenin in capsules (135–199 mg) or ingestion of grapefruit juice (single dose, 8 mL of juice/kg) leads to much lower plasma naringenin concentrations (6.0–7.3 μmol/L) [53, 54] than concentrations used in vitro (50–240 μM), suggesting suboptimal doses for clinical efficacy. Therefore, further experimentation should focus on more closely linking in vivo observations with in vitro mechanistic studies.
Tangeretin also has been shown to increase hepatic protein concentrations of PPARα which was associated with increased fatty acid oxidation and a reduction in liver triglyceride [55]. Addition of a mixture of polymethoxylated flavonoids (1 : 1; tangeretin : nobiletin) to the diet of fructose-fed Golden Syrian hamsters reduced plasma TG and prevented lipid accumulation in liver and heart, but not in muscle [56]. These authors demonstrated a significant increase in hepatic PPARα protein expression, suggesting that an increase in fatty acid oxidation leads to the observed attenuation of hepatic steatosis and improved dyslipidemia [56].
5. Amelioration of Insulin Resistance by Citrus Flavonoids
Skeletal muscle is the major tissue responsible for insulin-mediated glucose uptake. In states where FA flux is increased, as in insulin resistance, both the oxidation of glucose and its conversion to glycogen are impaired [57]. Myocytes and hepatocytes can become overwhelmed by lipid, with storage and oxidation pathways at capacity, allowing FA intermediates such as ceramide and diacylglycerol to accumulate [58, 59]. FA metabolites activate serine kinase pathways including JNK and PLC which can lead to inhibition of the insulin receptor [60], preventing its activation and downstream signaling. Without activation of PI3K, GLUT4 does not undergo translocation and fusion with the plasma membrane resulting in impaired glucose uptake and utilization [61]. Pparα−/− mice fed a high-fat diet have increased TG in adipose and muscle, but do not develop insulin resistance, suggesting that PPARα-stimulated oxidation of ectopic lipid in peripheral tissues, including muscle, increases the cellular concentration of lipid derivatives which contribute to insulin resistance [35]. Therefore, treatments that promote glucose utilization by preventing cellular accumulation of TG- and FA-derived lipid metabolites may substantially improve peripheral insulin resistance and ameliorate impaired glucose uptake.
Interestingly, in western diet-fed Ldlr−/− mice, naringenin did not stimulate fatty acid oxidation in muscle nor were any changes observed in the expression of genes linked to fatty acid oxidation, such as Cpt1β, Ucp1, and Ucp3 [45]. Instead, naringenin inhibited SREBP1c-mediated de novo lipogenesis and promoted glucose uptake. These results suggest that naringenin does not act directly on muscle. Reduced muscle lipid accumulation and improved glucose utilization in this model is consequent to attenuated de novo lipogenesis and decreased uptake of lipoprotein-derived lipid, the latter a consequence of decreased VLDL secretion. By increasing hepatic FA oxidation, naringenin permits glucose to be utilized by peripheral tissues, including muscle, resulting in the prevention of both lipid accumulation and insulin resistance (Figure 2). The protective effect of naringenin in preventing the metabolic disturbances associated with high-fat feeding was also observed in wild-type C57BL/6J mice. In this model, naringenin significantly reduced plasma and hepatic lipids, normalized glucose tolerance and insulin sensitivity and prevented obesity when compared to western-fed mice, although the time course for the preventative effect was significantly longer than that required for Ldlr−/− mice [45].
Figure 2.
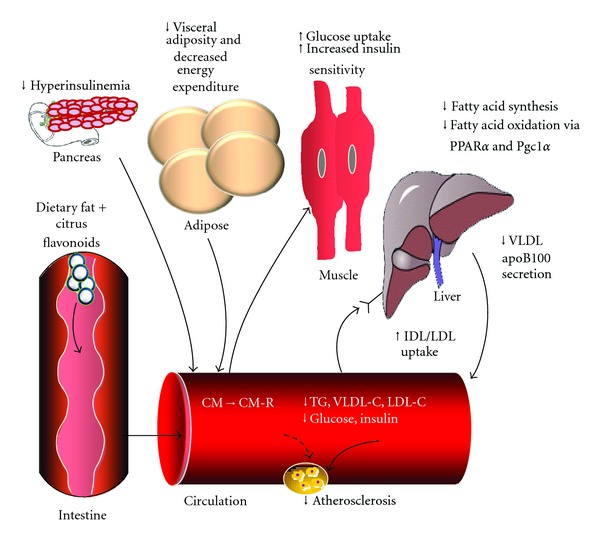
Potential mechanisms for the reduction in risk factors for atherosclerosis by flavonoids. Both in vitro and in vivo data suggest that citrus flavonoids decrease macrophage uptake of oxidized LDL and macrophage CE accumulation and improve dyslipidemia. While the complete mechanisms have not been fully defined, the nuclear hormone receptors PPARα and PGC1α represent important molecular targets. The improvement in dyslipidemia can be linked to decreased VLDL secretion as hepatic lipid availability for storage or VLDL secretion is decreased as a consequence of PPAR-stimulated fatty acid oxidation. The oxidation of fatty acids by liver prevents ectopic lipid accumulation and improves both insulin sensitivity and glucose tolerance. Arrows indicate change in response to flavonoid treatment.
6. Citrus Flavonoids as Antioxidants and Free Radical Scavengers
Flavonoids, as dictated by their structure, are reducing agents and can serve as efficient chelators of transition metals involved in cellular oxidation reactions [62, 63]. The association between flavonoids, including citrus flavonoids, and metabolic disorders and atherosclerosis has been linked to their antioxidant properties and to a reduction in oxidative stress. In vitro, naringin is an efficient free radical scavenger, but has limited antioxidant activity in mouse liver homogenates [64]. In STZ diabetic rats, naringin (40 mg/kg/day) was shown to prevent diabetic neurogenic pain through its prevention of free radical formation and antioxidant activities in the sciatic nerve [65]. In mice fed a high-fat diet, naringin (0.02%) improved indices of insulin resistance, which was attributed, in part, to improved hepatic enzymes involved in free radical scavenging and antioxidant activities [66]. Naringenin has also been demonstrated to have antioxidant properties. Naringenin (0.02%) increased hepatic glutathione peroxidase and superoxide dismutase activities and decreased hepatic and plasma TBARS [67]. In diabetic rats, naringenin (50 mg/kg/day) significantly increased enzymes with antioxidant activity in pancreas and plasma and decreased serum activities of ALT, AST, ALP, and LDH [68]. However, it is not known if naringenin or naringin has direct effects on the expression of activities of these enzymes, or if the effects are secondary to the flavonoid-induced correction of dysregulated lipid metabolism.
7. Prevention of Atherosclerosis by Citrus Flavonoids
In addition to correction of dyslipidemia and glucose intolerance, treatment with nobiletin (0.3% w/w) or naringenin (3% w/w) in Ldlr−/− mice for 6 months led to a marked reduction in the progression of atherosclerosis [48, 69]. In rabbits fed a high-cholesterol diet, supplementation of 0.1% naringin or 0.05% naringenin for 8 weeks decreased aortic fatty streak area when compared to controls [70]. In wild-type mice fed a high-fat/high-cholesterol/0.5% cholic acid in a cocoa butter diet, 0.02% naringin reduced plaque progression by 41%, but no beneficial effects of naringin were observed in ApoE−/− mice fed the same diet [71]. Naringin significantly reduced soluble E-selectin and soluble inter-cell adhesion molecule-1 in these mice [71]. Molecular studies have demonstrated that incubation of lipopolysaccharide-treated RAW-264.7 macrophages with naringenin, dose-dependently decreased the production of tumour necrosis factor (TNFα) and monocyte chemoattractant protein 1 (MCP-1) [72]. Nobiletin and its derivatives have been shown to reduce the expression and activity of macrophage scavenger receptors [73] including CD36 and SRAI/II and decrease the uptake of acetylated LDL [74]. These studies suggest that in addition to decreased exposure of the vessel wall to increased plasma lipoproteins, flavonoids directly decrease both lipoprotein uptake and the inflammatory response within macrophages of the arterial intima. PPARα activation has also been demonstrated to reduce monocyte recruitment to the vascular wall by downregulating vascular cell adhesion molecule-1, MCP-1 and interleukin-6 [75]. Ligands of PPARα have also been demonstrated to reduce foam cell formation by reducing lipid uptake and increasing lipid efflux [76]. Fenofibrate has been demonstrated to reduce foam cell formation and slow the progression of atherosclerosis in mice [77, 78], suggesting that some of the effects of citrus flavonoids may be mediated through PPARα-stimulated pathways.
8. Additional Targets of Citrus Flavonoids
While some preliminary human studies suggest metabolic benefit [39–41], an enhanced understanding of the molecular targets of natural polyphenols is required in order to identify the main mechanism of action. While PPARα has emerged as a potential target, numerous studies have also identified PPARγ as a prospective target. In diabetic male Wistar albino rats fed a high-fat diet, naringin increased PPARγ protein expression in liver. Interestingly, the authors also observed a decrease in the expression of LXR and SREBP1c and a reduction in hepatic steatosis [79]. It has also been reported that naringenin at 80 μM, activated a PPARγ-GAL4 construct up to 57% [52]. In 3T3-L1 adipocytes, both naringenin and hesperitin upregulated PPARγ at both 80 μM and 160 μM, which resulted in a significant increase in the expression of adiponectin [51]. Nobiletin (10 μM) stimulates differentiation of ST-13 preadipocytes into mature adipocytes and significantly increases the production of adiponectin protein [80].
9. Summary
Collectively, these studies add to the growing body of evidence that citrus flavonoids have marked lipid and lipoprotein lowering potential and demonstrate that naringenin and nobiletin reduce hepatic lipid accumulation and prevent lipoprotein overproduction, normalize glucose tolerance and insulin sensitivity and slow the progression of atherosclerosis (Figure 2). These beneficial metabolic effects are mediated, in part, by the metabolic regulators, PPARα, PPARγ, and PGC1α, although further studies are required to fully reveal the interaction of these flavonoids with upstream mediators in this pathway.
Conflict of Interests
The authors declare that they have no financial conflicts of interests.
Acknowledgments
Research from the authors' laboratory was funded by Grants from the Heart and Stroke Foundation of Ontario (T-7007 and PG-5967) and the Canadian Institutes of Health Research (MT-8014).
Abbreviations
- ACOX:
Acyl-CoA oxidase
- ACAT:
Acyl-CoA:cholesterol acyltransferase
- apoB100:
Apolipoprotein B100
- CPT1:
Carnitine palmitoyl transferase 1
- CE:
Cholesteryl ester
- CVD:
Cardiovascular disease
- DGAT1/2:
Diacylglycerol acyl transferase 1/2
- FA:
Fatty acid
- FAS:
Fatty acid synthase
- FC:
Free cholesterol
- HDL:
High density lipoprotein
- LDL:
Low density lipoprotein
- MCP:
Monocyte chemmoattractant protein 1
- MTP:
Microsomal triglyceride transfer protein
- PI3K:
Phosphoinositide-3-kinase
- PPAR:
Peroxisome proliferator-activated receptor
- PPRE:
PPAR response element
- PGC1α:
Peroxisome proliferator-activated receptor gamma coactivator 1α
- SREBP1c:
Sterol regulatory element binding protein
- TG:
Triglyceride
- TNFα:
Tumour necrosis factor
- VLDL:
Very low density lipoprotein
- UCP-1:
Uncoupling protein 1.
References
- 1.Garg A, Haffner SM. Insulin resistance and atherosclerosis: an overview. Diabetes Care. 1996;19(3, article 274) doi: 10.2337/diacare.19.3.274. [DOI] [PubMed] [Google Scholar]
- 2.Ginsberg HN, Huang LS. The insulin resistance syndrome: impact on lipoprotein metebolism and atherothrombosis. Journal of Cardiovascular Risk. 2000;7(5):325–331. doi: 10.1177/204748730000700505. [DOI] [PubMed] [Google Scholar]
- 3.Ginsberg HN, Zhang YL, Hernandez-Ono A. Metabolic syndrome: focus on dyslipidemia. Obesity. 2006;14(supplement 1):41S–49S. doi: 10.1038/oby.2006.281. [DOI] [PubMed] [Google Scholar]
- 4.Lee DS, Chiu M, Manuel DG, et al. Trends in risk factors for cardiovascular disease in Canada: temporal, socio-demographic and geographic factors. CMAJ. 2009;181(3-4):E55–E66. doi: 10.1503/cmaj.081629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Scalbert A, Williamson G. Dietary intake and bioavailability of polyphenols. Journal of Nutrition. 2000;130(8, supplement):2073S–2085S. doi: 10.1093/jn/130.8.2073S. [DOI] [PubMed] [Google Scholar]
- 6.Hertog MGL, Feskens EJM, Hollman PCH, Katan MB, Kromhout D. Dietary antioxidant flavonoids and risk of coronary heart disease: the Zutphen Elderly Study. The Lancet. 1993;342(8878):1007–1011. doi: 10.1016/0140-6736(93)92876-u. [DOI] [PubMed] [Google Scholar]
- 7.Bazzano LA. The high cost of not consuming fruits and vegetables. Journal of the American Dietetic Association. 2006;106(9):1364–1368. doi: 10.1016/j.jada.2006.06.021. [DOI] [PubMed] [Google Scholar]
- 8.Hooper L, Kroon PA, Rimm EB, et al. Flavonoids, flavonoid-rich foods, and cardiovascular risk: a meta-analysis of randomized controlled trials. American Journal of Clinical Nutrition. 2008;88(1):38–50. doi: 10.1093/ajcn/88.1.38. [DOI] [PubMed] [Google Scholar]
- 9.Tohill BC, Seymour J, Serdula M, Kettel-Khan L, Rolls BJ. What epidemiologic studies tell us about the relationship between fruit and vegetable consumption and body weight. Nutrition Reviews. 2004;62(10):365–374. doi: 10.1111/j.1753-4887.2004.tb00007.x. [DOI] [PubMed] [Google Scholar]
- 10.Kurowska EM, Spence JD, Jordan J, et al. HDL-cholesterol-raising effect of orange juice in subjects with hypercholesterolemia. American Journal of Clinical Nutrition. 2000;72(5):1095–1100. doi: 10.1093/ajcn/72.5.1095. [DOI] [PubMed] [Google Scholar]
- 11.Ock KC, Sang JC, Song WO. Estimated dietary flavonoid intake and major food sources of U.S. adults. Journal of Nutrition. 2007;137(5):1244–1252. doi: 10.1093/jn/137.5.1244. [DOI] [PubMed] [Google Scholar]
- 12.Zamora-Ros R, Knaze V, Luján-Barroso L, et al. Estimated dietary intakes of flavonols, flavanones and flavones in the European Prospective Investigation into Cancer and Nutrition (EPIC) 24 hour dietary recall cohort. British Journal of Nutrition. 2011;106(12):1915–1925. doi: 10.1017/S000711451100239X. [DOI] [PubMed] [Google Scholar]
- 13.Mulvihill EE, Huff MW. Antiatherogenic properties of flavonoids: implications for cardiovascular health. Canadian Journal of Cardiology. 2010;26(supplement A):17A–21A. doi: 10.1016/s0828-282x(10)71056-4. [DOI] [PubMed] [Google Scholar]
- 14.Choi JS, Yokozawa T, Oura H. Improvement of hyperglycemia and hyperlipemia in streptozotocin-diabetic rats by a methanolic extract of Prunus davidiana stems and its main component, prunin. Planta Medica. 1991;57(3):208–211. doi: 10.1055/s-2006-960075. [DOI] [PubMed] [Google Scholar]
- 15.Unger RH, Scherer PE. Gluttony, sloth and the metabolic syndrome: a roadmap to lipotoxicity. Trends in Endocrinology and Metabolism. 2010;21(6):345–352. doi: 10.1016/j.tem.2010.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Grundy SM. Metabolic syndrome: a multiplex cardiovascular risk factor. The Journal of Clinical Endocrinology and Metabolism. 2007;92(2):399–404. doi: 10.1210/jc.2006-0513. [DOI] [PubMed] [Google Scholar]
- 17.Yen CLE, Stone SJ, Koliwad S, Harris C, Farese RV. DGAT enzymes and triacylglycerol biosynthesis. Journal of Lipid Research. 2008;49(11):2283–2301. doi: 10.1194/jlr.R800018-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Khan RS, Drosatos K, Goldberg IJ. Creating and curing fatty hearts. Current Opinion in Clinical Nutrition and Metabolic Care. 2010;13(2):145–149. doi: 10.1097/MCO.0b013e3283357272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lee CH, Olson P, Evans RM. Minireview: lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology. 2003;144(6):2201–2207. doi: 10.1210/en.2003-0288. [DOI] [PubMed] [Google Scholar]
- 20.Alaynick WA. Nuclear receptors, mitochondria and lipid metabolism. Mitochondrion. 2008;8(4):329–337. doi: 10.1016/j.mito.2008.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bookout AL, Jeong Y, Downes M, Yu RT, Evans RM, Mangelsdorf DJ. Anatomical profiling of nuclear receptor expression reveals a hierarchical transcriptional network. Cell. 2006;126(4):789–799. doi: 10.1016/j.cell.2006.06.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Forman BM, Chen J, Evans RM. Hypolipidemic drugs, polyunsaturated fatty acids, and eicosanoids are ligands for peroxisome proliferator-activated receptors α and δ . Proceedings of the National Academy of Sciences of the United States of America. 1997;94(9):4312–4317. doi: 10.1073/pnas.94.9.4312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Peters JM, Cattley RC, Gonzalez FJ. Role of PPARα in the mechanism of action of the nongenotoxic carcinogen and peroxisome proliferator Wy-14,643. Carcinogenesis. 1997;18(11):2029–2033. doi: 10.1093/carcin/18.11.2029. [DOI] [PubMed] [Google Scholar]
- 24.Bonnefont JP, Djouadi F, Prip-Buus C, Gobin S, Munnich A, Bastin J. Carnitine palmitoyltransferases 1 and 2: biochemical, molecular and medical aspects. Molecular Aspects of Medicine. 2004;25(5-6):495–520. doi: 10.1016/j.mam.2004.06.004. [DOI] [PubMed] [Google Scholar]
- 25.Schreurs M, Kuipers F, Van Der Leij FR. Regulatory enzymes of mitochondrial β-oxidation as targets for treatment of the metabolic syndrome. Obesity Reviews. 2010;11(5):380–388. doi: 10.1111/j.1467-789X.2009.00642.x. [DOI] [PubMed] [Google Scholar]
- 26.Schreurs M, van Dijk TH, Gerding A, Havinga R, Reijngoud DJ, Kuipers F. Soraphen, an inhibitor of the acetyl-CoA carboxylase system, improves peripheral insulin sensitivity in mice fed a high-fat diet. Diabetes, Obesity and Metabolism. 2009;11(10):987–991. doi: 10.1111/j.1463-1326.2009.01078.x. [DOI] [PubMed] [Google Scholar]
- 27.Chakravarthy MV, Pan Z, Zhu Y, et al. New hepatic fat activates PPARα to maintain glucose, lipid, and cholesterol homeostasis. Cell Metabolism. 2005;1(5):309–322. doi: 10.1016/j.cmet.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 28.Orellana-Gavaldà JM, Herrero L, Malandrino MI, et al. Molecular therapy for obesity and diabetes based on a long-term increase in hepatic fatty-acid oxidation. Hepatology. 2011;53(3):821–832. doi: 10.1002/hep.24140. [DOI] [PubMed] [Google Scholar]
- 29.Yu S, Reddy JK. Transcription coactivators for peroxisome proliferator-activated receptors. Biochimica et Biophysica Acta. 2007;1771(8):936–951. doi: 10.1016/j.bbalip.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 30.Lin J, Handschin C, Spiegelman BM. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metabolism. 2005;1(6):361–370. doi: 10.1016/j.cmet.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 31.Handschin C, Spiegelman BM. Peroxisome proliferator-activated receptor γ coactivator 1 coactivators, energy homeostasis, and metabolism. Endocrine Reviews. 2006;27(7):728–735. doi: 10.1210/er.2006-0037. [DOI] [PubMed] [Google Scholar]
- 32.Herzig S, Long F, Jhala US, et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature. 2001;413(6852):179–183. doi: 10.1038/35093131. [DOI] [PubMed] [Google Scholar]
- 33.Leone TC, Lehman JJ, Finck BN, et al. PGC-1alpha deficiency causes multi-system energy metabolic derangements: muscle dysfunction, abnormal weight control and hepatic steatosis. PLoS Biology. 2005;3(4, article e101) doi: 10.1371/journal.pbio.0030101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Estall JL, Kahn M, Cooper MP, et al. Sensitivity of lipid metabolism and insulin signaling to genetic alterations in hepatic peroxisome proliferator-activated receptor-γ coactivator-1α expression. Diabetes. 2009;58(7):1499–1508. doi: 10.2337/db08-1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guerre-Millo M, Rouault C, Poulain P, et al. PPAR-α-null mice are protected from high-fat diet-induced insulin resistance. Diabetes. 2001;50(12):2809–2814. doi: 10.2337/diabetes.50.12.2809. [DOI] [PubMed] [Google Scholar]
- 36.Aoyama T, Peters JM, Iritani N, et al. Altered constitutive expression of fatty acid-metabolizing enzymes in mice lacking the peroxisome proliferator-activated receptor α (PPARα) The Journal of Biological Chemistry. 1998;273(10):5678–5684. doi: 10.1074/jbc.273.10.5678. [DOI] [PubMed] [Google Scholar]
- 37.Leone TC, Weinheimer CJ, Kelly DP. A critical role for the peroxisome proliferator-activated receptor α (PPARα) in the cellular fasting response: the PPARα-null mouse as a model of fatty acid oxidation disorders. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(13):7473–7478. doi: 10.1073/pnas.96.13.7473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lee M, Saver JL, Towfighi A, Chow J, Ovbiagele B. Efficacy of fibrates for cardiovascular risk reduction in persons with atherogenic dyslipidemia: a meta-analysis. Atherosclerosis. 2011;217(2):492–498. doi: 10.1016/j.atherosclerosis.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 39.Gorinstein S, Caspi A, Libman I, Katrich E, Lerner HT, Trakhtenberg S. Preventive effects of diets supplemented with sweetie fruits in hypercholesterolemic patients suffering from coronary artery disease. Preventive Medicine. 2004;38(6):841–847. doi: 10.1016/j.ypmed.2003.12.021. [DOI] [PubMed] [Google Scholar]
- 40.Gorinstein S, Caspi A, Libman I, et al. Red grapefruit positively influences serum triglyceride level in patients suffering from coronary atherosclerosis: studies in vitro and in humans. Journal of Agricultural and Food Chemistry. 2006;54(5):1887–1892. doi: 10.1021/jf058171g. [DOI] [PubMed] [Google Scholar]
- 41.Jung UJ, Kim HJ, Lee JS, et al. Naringin supplementation lowers plasma lipids and enhances erythrocyte antioxidant enzyme activities in hypercholesterolemic subjects. Clinical Nutrition. 2003;22(6):561–568. doi: 10.1016/s0261-5614(03)00059-1. [DOI] [PubMed] [Google Scholar]
- 42.Demonty I, Lin Y, Zebregs YEMP, et al. The citrus flavonoids hesperidin and naringin do not affect serum cholesterol in moderately hypercholesterolemic men and women. Journal of Nutrition. 2010;140(9):1615–1620. doi: 10.3945/jn.110.124735. [DOI] [PubMed] [Google Scholar]
- 43.Borradaile NM, De Dreu LE, Barrett PHR, Behrsin CD, Huff MW. Hepatocyte ApoB-containing lipoprotein secretion is decreased by the grapefruit flavonoid, naringenin, via inhibition of MTP-mediated microsomal triglyceride accumulation. Biochemistry. 2003;42(5):1283–1291. doi: 10.1021/bi026731o. [DOI] [PubMed] [Google Scholar]
- 44.Borradaile NM, De Dreu LE, Barrett PHR, Huff MW. Inhibition of hepatocyte apoB secretion by naringenin: enhanced rapid intracellular degradation independent of reduced microsomal cholesteryl esters. Journal of Lipid Research. 2002;43(9):1544–1554. doi: 10.1194/jlr.m200115-jlr200. [DOI] [PubMed] [Google Scholar]
- 45.Mulvihill EE, Allister EM, Sutherland BG, et al. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor-null mice with diet-induced insulin resistance. Diabetes. 2009;58(10):2198–2210. doi: 10.2337/db09-0634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Huong DTT, Takahashi Y, Ide T. Activity and mRNA levels of enzymes involved in hepatic fatty acid oxidation in mice fed citrus flavonoids. Nutrition. 2006;22(5):546–552. doi: 10.1016/j.nut.2005.11.006. [DOI] [PubMed] [Google Scholar]
- 47.Wood N. Hepatolipidemic effects of naringenin in high cornstarch- versus high coconut oil-fed rats. Journal of Medicinal Food. 2004;7(3):315–319. doi: 10.1089/jmf.2004.7.315. [DOI] [PubMed] [Google Scholar]
- 48.Mulvihill EE, Assini JM, Lee JK, et al. Nobiletin attenuates VLDL overproduction, dyslipidemia, and atherosclerosis in mice with diet-induced insulin resistance. Diabetes. 2011;60(5):1446–1457. doi: 10.2337/db10-0589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kurowska EM, Manthey JA. Hypolipidemic effects and absorption of citrus polymethoxylated flavones in hamsters with diet-induced hypercholesterolemia. Journal of Agricultural and Food Chemistry. 2004;52(10):2879–2886. doi: 10.1021/jf035354z. [DOI] [PubMed] [Google Scholar]
- 50.Jung UJ, Lee MK, Park YB, Kang MA, Choi MS. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mRNA levels in type-2 diabetic mice. International Journal of Biochemistry and Cell Biology. 2006;38(7):1134–1145. doi: 10.1016/j.biocel.2005.12.002. [DOI] [PubMed] [Google Scholar]
- 51.Liu L, Shan S, Zhang K, Ning ZQ, Lu XP, Cheng YY. Naringenin and hesperetin, two flavonoids derived from Citrus aurantium up-regulate transcription of adiponectin. Phytotherapy Research. 2008;22(10):1400–1403. doi: 10.1002/ptr.2504. [DOI] [PubMed] [Google Scholar]
- 52.Goldwasser J, Cohen PY, Yang E, Balaguer P, Yarmush ML, Nahmias Y. Transcriptional regulation of human and rat hepatic lipid metabolism by the grapefruit flavonoid naringenin: role of PPARα, PPARγ and LXRα . PLoS ONE. 2010;5(8) doi: 10.1371/journal.pone.0012399. Article ID e12399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Erlund I, Meririnne E, Alfthan G, Aro A. Human nutrition and metabolism: plasma kinetics and urinary excretion of the flavanones naringenin and hesperetin in humans after ingestion of orange juice and grapefruit juice. Journal of Nutrition. 2001;131(2):235–241. doi: 10.1093/jn/131.2.235. [DOI] [PubMed] [Google Scholar]
- 54.Kanaze FI, Bounartzi MI, Georgarakis M, Niopas I. Pharmacokinetics of the citrus flavanone aglycones hesperetin and naringenin after single oral administration in human subjects. European Journal of Clinical Nutrition. 2007;61(4):472–477. doi: 10.1038/sj.ejcn.1602543. [DOI] [PubMed] [Google Scholar]
- 55.Kurowska EM, Manthey JA, Casaschi A, Theriault AG. Modulation of HepG2 cell net apolipoprotein B secretion by the citrus polymethoxyflavone tangeretin. Lipids. 2004;39(2):143–151. doi: 10.1007/s11745-004-1212-8. [DOI] [PubMed] [Google Scholar]
- 56.Li RW, Theriault AG, Au K, et al. Citrus polymethoxylated flavones improve lipid and glucose homeostasis and modulate adipocytokines in fructose-induced insulin resistant hamsters. Life Sciences. 2006;79(4):365–373. doi: 10.1016/j.lfs.2006.01.023. [DOI] [PubMed] [Google Scholar]
- 57.Kashyap SR, Belfort R, Berria R, et al. Discordant effects of a chronic physiological increase in plasma FFA on insulin signaling in healthy subjects with or without a family history of type 2 diabetes. American Journal of Physiology. 2004;287(3):E537–E546. doi: 10.1152/ajpendo.00541.2003. [DOI] [PubMed] [Google Scholar]
- 58.Adiels M, Taskinen MR, Borén J. Fatty liver, insulin resistance, and dyslipidemia. Current Diabetes Reports. 2008;8(1):60–64. doi: 10.1007/s11892-008-0011-4. [DOI] [PubMed] [Google Scholar]
- 59.Koves TR, Ussher JR, Noland RC, et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metabolism. 2008;7(1):45–56. doi: 10.1016/j.cmet.2007.10.013. [DOI] [PubMed] [Google Scholar]
- 60.Schenk S, Saberi M, Olefsky JM. Insulin sensitivity: modulation by nutrients and inflammation. The Journal of Clinical Investigation. 2008;118(9):2992–3002. doi: 10.1172/JCI34260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Klip A. The many ways to regulate glucose transporter 4. Applied Physiology, Nutrition and Metabolism. 2009;34(3):481–487. doi: 10.1139/H09-047. [DOI] [PubMed] [Google Scholar]
- 62.Huxley RR, Neil HAW. The relation between dietary flavonol intake and coronary heart disease mortality: a meta-analysis of prospective cohort studies. European Journal of Clinical Nutrition. 2003;57(8):904–908. doi: 10.1038/sj.ejcn.1601624. [DOI] [PubMed] [Google Scholar]
- 63.Aherne SA, O’Brien NM. Mechanism of protection by the flavonoids, quercetin and rutin, against tert-butylhydroperoxide- and menadione-induced DNA single strand breaks in Caco-2 cells. Free Radical Biology and Medicine. 2000;29(6):507–514. doi: 10.1016/s0891-5849(00)00360-9. [DOI] [PubMed] [Google Scholar]
- 64.Yuting C, Rongliang Z, Zhongjian J, Yong J. Flavonoids as superoxide scavengers and antioxidants. Free Radical Biology and Medicine. 1990;9(1):19–21. doi: 10.1016/0891-5849(90)90045-k. [DOI] [PubMed] [Google Scholar]
- 65.Kandhare AD, Raygude KS, Ghosh P, Ghule AE, Bodhankar SL. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. doi: 10.1016/j.fitote.2012.01.010. Fitoterapia. In press. [DOI] [PubMed] [Google Scholar]
- 66.Pu P, Gao D-M, Mohamed S, et al. Naringin ameliorates metabolic syndrome by activating AMP-activated protein kinase in mice fed a high-fat diet. Archives of Biochemistry and Biophysics. 2012;518(1):61–70. doi: 10.1016/j.abb.2011.11.026. [DOI] [PubMed] [Google Scholar]
- 67.Jeon SM, Kim HK, Kim HJ, et al. Hypocholesterolemic and antioxidative effects of naringenin and its two metabolites in high-cholesterol fed rats. Translational Research. 2007;149(1):15–21. doi: 10.1016/j.trsl.2006.08.001. [DOI] [PubMed] [Google Scholar]
- 68.Annadurai T, Muralidharan AR, Joseph T, Hsu MJ, Thomas PA, Geraldine P. Antihyperglycemic and antioxidant effects of a flavanone, naringenin, in streptozotocin-nicotinamide-induced experimental diabetic rats. doi: 10.1007/s13105-011-0142-y. Journal of Physiology and Biochemistry. In press. [DOI] [PubMed] [Google Scholar]
- 69.Mulvihill EE, Assini JM, Sutherland BG, et al. Naringenin decreases progression of atherosclerosis by improving dyslipidemia in high-fat-fed low-density lipoprotein receptor-null mice. Arteriosclerosis, Thrombosis, and Vascular Biology. 2010;30(4):742–748. doi: 10.1161/ATVBAHA.109.201095. [DOI] [PubMed] [Google Scholar]
- 70.Lee CH, Jeong TS, Choi YK, et al. Anti-atherogenic effect of citrus flavonoids, naringin and naringenin, associated with hepatic ACAT and aortic VCAM-1 and MCP-1 in high cholesterol-fed rabbits. Biochemical and Biophysical Research Communications. 2001;284(3):681–688. doi: 10.1006/bbrc.2001.5001. [DOI] [PubMed] [Google Scholar]
- 71.Chanet A, Milenkovic D, Deval C, et al. Naringin, the major grapefruit flavonoid, specifically affects atherosclerosis development in diet-induced hypercholesterolemia in mice. Journal of Nutritional Biochemistry. 2012;23(5):469–477. doi: 10.1016/j.jnutbio.2011.02.001. [DOI] [PubMed] [Google Scholar]
- 72.Hirai S, Kim YI, Goto T, et al. Inhibitory effect of naringenin chalcone on inflammatory changes in the interaction between adipocytes and macrophages. Life Sciences. 2007;81(16):1272–1279. doi: 10.1016/j.lfs.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 73.Yen JH, Weng CY, Li S, et al. Citrus flavonoid 5-demethylnobiletin suppresses scavenger receptor expression in THP-1 cells and alters lipid homeostasis in HepG2 liver cells. Molecular Nutrition and Food Research. 2011;55(5):733–748. doi: 10.1002/mnfr.201000226. [DOI] [PubMed] [Google Scholar]
- 74.Whitman SC, Kurowska EM, Manthey JA, Daugherty A. Nobiletin, a citrus flavonoid isolated from tangerines, selectively inhibits class A scavenger receptor-mediated metabolism of acetylated LDL by mouse macrophages. Atherosclerosis. 2005;178(1):25–32. doi: 10.1016/j.atherosclerosis.2004.07.034. [DOI] [PubMed] [Google Scholar]
- 75.Pasceri V, Chang J, Willerson JT, Yeh ETH. Modulation of C-reactive protein-mediated monocyte chemoattractant protein-1 induction in human endothelial cells by anti-atherosclerosis drugs. Circulation. 2001;103(21):2531–2534. doi: 10.1161/01.cir.103.21.2531. [DOI] [PubMed] [Google Scholar]
- 76.Van Bilsen M, Van Nieuwenhoven FA. PPARs as therapeutic targets in cardiovascular disease. Expert Opinion on Therapeutic Targets. 2010;14(10):1029–1045. doi: 10.1517/14728222.2010.512917. [DOI] [PubMed] [Google Scholar]
- 77.Duezt H, Chao YS, Hernandez M, et al. Reduction of atherosclerosis by the peroxisome proliferator-activated receptor α agonist fenofibrate in mice. The Journal of Biological Chemistry. 2002;277(50):48051–48057. doi: 10.1074/jbc.M206966200. [DOI] [PubMed] [Google Scholar]
- 78.Hennuyer N, Tailleux A, Torpier G, et al. PPARα, but not PPARγ, activators decrease macrophage-laden atherosclerotic lesions in a nondiabetic mouse model of mixed dyslipidemia. Arteriosclerosis, Thrombosis, and Vascular Biology. 2005;25(9):1897–1902. doi: 10.1161/01.ATV.0000175756.56818.ee. [DOI] [PubMed] [Google Scholar]
- 79.Sharma AK, Bharti S, Ojha S, et al. Up-regulation of PPARγ, heat shock protein-27 and-72 by naringin attenuates insulin resistance, β-cell dysfunction, hepatic steatosis and kidney damage in a rat model of type 2 diabetes. British Journal of Nutrition. 2011;106(11):1713–1723. doi: 10.1017/S000711451100225X. [DOI] [PubMed] [Google Scholar]
- 80.Kunimasa K, Kuranuki S, Matsuura N, et al. Identification of nobiletin, a polymethoxyflavonoid, as an enhancer of adiponectin secretion. Bioorganic and Medicinal Chemistry Letters. 2009;19(7):2062–2064. doi: 10.1016/j.bmcl.2009.02.002. [DOI] [PubMed] [Google Scholar]