

Abstract
Local interneurons are continuously regenerated in the olfactory bulb of adult rodents1-3. In this process, called adult neurogenesis, neural stem cells in the walls of the lateral ventricle give rise to neuroblasts that migrate for several millimeters along the rostral migratory stream (RMS) to reach and incorporate into the olfactory bulb. To study the different steps and the impact of adult-born neuron integration into preexisting olfactory circuits, it is necessary to selectively label and manipulate the activity of this specific population of neurons. The recent development of optogenetic technologies offers the opportunity to use light to precisely activate this specific cohort of neurons without affecting surrounding neurons4,5. Here, we present a series of procedures to virally express Channelrhodopsin2(ChR2)-YFP in a temporally restricted cohort of neuroblasts in the RMS before they reach the olfactory bulb and become adult-born neurons. In addition, we show how to implant and calibrate a miniature LED for chronic in vivo stimulation of ChR2-expressing neurons.
Keywords: Neuroscience, Issue 58, Olfactory bulb, Olfactory neurons, in vivo, viral transduction, mouse, LED
Protocol
1. Stereotaxic injections
The virus used in these experiments is a replication-deficient lentiviral vector based on the HIV virus to express Channelrhodopsine2-YFP (yellow fluorescent protein) fusion construct driven by a synapsin promoter6. The plasmid was generously provided by K. Deisseroth's research group (http://www.stanford.edu/group/dlab/optogenetics/index.html). Lentiviruses can be obtained from commercial sources or produced in the lab according to established protocols7. Average vector titers were in the order of 1010 transduction units/ml. Viral aliquots are stored at -80°C and should be thawed just before pipette loading. Multiple freeze/thaw cycles should be avoided. Never exposure the virus solution directly to dry ice.
Prepare glass pipettes using borosilicate glass capillaries on standard pipette puller (Sutter P-97 pipette puller). The pulling setting should be properly calibrated to yield a long and rigid pipette. Next, cut the tip of the pipette using a pair of scissors to obtain a tip of 30-40 μm in diameter. Store pulled pipettes in a dust free box.
Setup the Nanoject II microinjector with the manufacturer's instructions for delivery of 50 nL. Prepare the injection pipette by back-filling with mineral oil and insert it in to the microinjector plunger. Attach the microinjector onto a stereotaxic frame. Partially empty oil from the glass pipette before filling with the virus solution
With a micropipette, transfer 2 μl of virus solution onto a sterile piece of Parafilm. Gently lower the glass pipette into the drop of virus solution and fill the pipette with 1-2 μl. Make sure that there are no air bubbles in the pipette.
Anesthetize a mouse with 100 mg/kg Ketamine and 10 mg/kg Xylazine, diluted in sterile saline. Remove the hair from the scalp of the animal using rodent clippers, razor, scissors, or chemical depilitory agent. Disinfect the scalp with three applications of alternating 70% ethanol and betadine. All surgical instruments should also be autoclaved and then disinfected with 70% ethanol. Place the animal in the stereotaxic frame with ear bars and nose bar, assuring that the head is secure. Apply eye ointment to prevent drying of the corneas.
With a scalpel, cut the scalp from between the eyes to between the ears. Pull the skin aside to expose the skull and anchor the skin with a pair of clamps. Clean the surface of the skull. The stereotaxic apparatus is zeroed with the tip of the glass pipette at bregma. Then the tip is positioned at the injection site (Antero-Posterior, 3.30; Medio-Lateral, ±0.82, for both right and left hemispheres, respectively). The position of the nose bar has to be adjusted according to the calculation of the stereotaxic coordinates. In our experiment, the nose bar is adjusted until bregma and the injection site are aligned to the same height.
Identify the injection site. Using a high-speed surgical drill, carefully drill holes into the skull. With a bent syringe needle hooked at the tip, remove the remnants of thin bone to expose the dura mater. Check that the hole is centered at the correct position. Take care not to rupture blood vessels on the surface of the brain while drilling. If blood vessels are ruptured, gently press onto the surface of the brain with a small cotton bud or tissue for several seconds until blood flow stops.
Lower the pipette and zero the Dorso-Ventral height when the tip of the pipette touches the surface of the brain. Then, lower to the correct Dorso-Ventral depth (Dorso-Ventral, 2.9 from brain surface) and wait 30 seconds for pressure equilibrium. Inject 4 times 50 nl of virus for a total of 200 nl. Wait 30 seconds between each injection.
One minute after the last injection, slowly withdraw the pipette. Repeat points 1.6-1.8 for the other hemisphere. Remove the animal from stereotaxic apparatus, clean the incision and stitch the cut skin using surgical threads. Allow the animal to recover on warming pad before returning to cage. Additionally, animals can be given a non-steroidal anti-inflammatory analgesic (e.g., Carprofen 4mg/kg) immediately after and 3 days following surgery.
2. Chronic LED implantation for in vivo optogenetic stimulation in the olfactory bulb
Animals injected with lentiviral particles bilaterally are next chronically implanted with a miniature blue LED (Osram, LED CMS 4.6W blue). Solder the LED pins to a female miniature electrical connector so that the connector is positioned on the opposite side of the LED. Check that there is no electrical short cut and identify the positive and negative pin on the connector. Test the LED by connecting it to a current controller for LED though a proper cable.
To measure and/or set the absolute light power of the LED, mount the LED on a micromanipulator and position the LED in direct contact with the photodetector of a power meter of a peak light intensity at 470 nm. Drill a pinhole of 3 mm diameter in an opaque PVC board. Affix this pinhole to the power meter and build a standard curve of optical power versus input current to calculate the power per mm2.
To measure the propagation of light through the brain, cut blocks of fresh brain tissue of 300, 500, 1000, 1500 and 2500 μm thickness in 0-4°C ACSF using a vibratome14. Place a piece of skull with a craniotomy identical to the one performed in live mice under the LED. First, measure the light power without tissue centered on LED. Then, place each block of tissue between the LED and the photodetector of the power meter, and measure the power. Build a curve of tissue thickness versus relative transmission fraction, calculated as the ratio between the power measured through tissue and without tissue.
Anesthetize a mouse with 100 mg/kg Ketamine and 10 mg/kg Xylazine, diluted in sterile saline. Clean and disinfect the LED with 70% ethanol. Remove the hair from the scalp of the animal and disinfect the scalp with three applications of alternating 70% ethanol and betadine. Then place the animal in the stereotaxic frame. Apply eye ointment to prevent drying of the corneas. Attach the connector to a stereotaxic holder. The position of the nose bar should be adjusted until the surface of the skull above the olfactory bulb is perfectly horizontal and parallel to the surface of the miniature LED.
With a scalpel, cut the scalp from 3 mm anterior to the eyes to 3 mm posterior to the ears. Pull the skin aside and anchor it with a pair of clamps. Scrape away the membranes at the surface of the skull using a razor blade. Clean and dry the skull. Apply a thin cover of cyanoacrylate adhesive to allow subsequent adhesion of dental acrylic.
Using a high-speed surgical drill, perform a rectangular craniotomy (1 mm caudal and 3 mm lateral; centered on 5.1 mm caudal, 0 mm lateral from bregma) over the olfactory bulbs. To do this, carefully thin the skull until only a thin layer of bone remains. Carefully remove the remaining bone with the help of a bent syringe needle. Special care should be taken to avoid puncturing the dura matter and rupturing the major blood vessels that lie between the two hemispheres and in the caudal border of the olfactory bulbs. Blood accumulation and coagulation at the brain surface precludes optimal light penetration into the tissue. Rinse the craniotomy and the surface of the brain with sterile saline and then dry the skull.
Attach the miniature LED to the connector. Lower the miniature LED on the top of the olfactory bulbs. The main axis of the miniature LED should be parallel to the rostral-caudal axis of the mouse, Clean with sterile saline and dry the skull. Then secure the LED to the skull with a first layer of acrylic (i.e., polycarboxylate cement), followed by a second layer of dental acrylic. The addition of two small screws may be required to fix the LED. The screws should be inserted on the skull (posterior to the LED position) and covered with both acrylics. While the acrylic is still liquid, pull back and glue the free edges of the scalp on the surface of the dental acrylic. Take care not to apply any acrylic on the connector. After the acrylic has hardened, remove the animal from stereotaxic apparatus and let it recover on a warming pad before returning to cage. Animals are left isolated in their cage to recover from the surgery for at least seven days. Additionally, animals can be given a non-steroidal anti-inflammatory analgesic (e.g., Carprofen 4mg/kg) immediately after and 3 days following surgery. LEDs should be tested before implantation and after termination the experiments.
A light electrical cable is used to tether the animal to a current controller for LED coupled to a pulse generator (Master-8) to control the timing and intensity of the LED. The thin cable allows for complete freedom of movement. Take care to respect the polarity of the connector. Inverting the polarity may break the LED.
3. Representative Results:
ChR2-YFP is expressed in transduced neurons approximately 48 hours after injection, and remains stable for several months7. Figure 1 is a representative picture of a sagittal section of a paraformaldehyde-fixed animal 15 days post-injection. Note the dense labeling and the size of the injection site in the RMS. A large number of newborn cells have migrated along the RMS to colonize the different layers of the olfactory bulb (Fig. 1). The fusion protein is expressed in all neuron compartments, including the soma, dendrites, and spines.
Miniature LED implantation allows large scale and inexpensive optical stimulation of a broad region of interest. The animal is tethered to the LED controller with a light electrical cable that allows a complete freedom of movement. By characterizing the light diffusion in the brain tissue and the threshold of ChR2 activation, we estimate that optical activation could be triggered at a depth of ~2mm below the surface of the LED (Fig. 2A). To monitor the functional activation of ChR2-expressing neurons in vivo, the expression of c-fos, a marker of neuronal activity, can be analyzed after repetitive light stimulation (Fig. 2B). As an alternative, in vitro electrophysiological recordings of ChR2-expressing neurons coupled to focal light stimulation can be performed14. Finally, two-photon targeted loose-patch recordings have been used to study the functional activation of ChR2-expressing cortical neurons by similar miniature LED in vivo9.
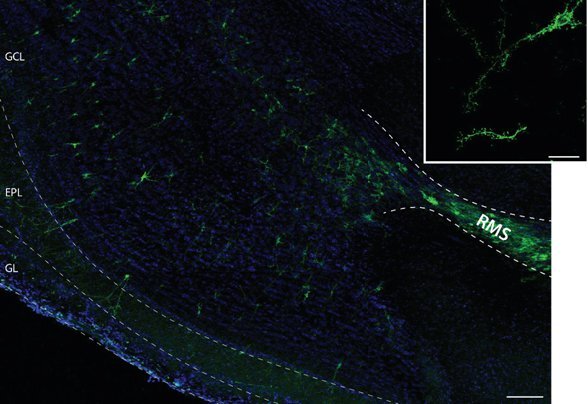 Figure 1. ChR2-YFP expression from lentiviral injection into the rostral migratory stream. Representative confocal images of a saggital sections of the olfactory bulb of a mouse 15 days after injections of a lentivirus in the rostral RMS. Note the presence of scattered YFP newborn neurons migrating from the end of the RMS to the different layers of the OB. Most of newborn cells (95%) are granule cells, which soma lie in the Granule Cell Layer (GCL) and dendrites arborize in the External Plexiform Layer (EPL). A small population of newborn neurons migrate up to the most superficial layer of the OB, in the Glomerular Layer (GL). Scale bar, 100μm. Inset, high magnification of a newborn granule cell, showing the membrane expression of ChR2-YFP in the dendrites as well as in spines. Scale bar, 20μm.
Figure 1. ChR2-YFP expression from lentiviral injection into the rostral migratory stream. Representative confocal images of a saggital sections of the olfactory bulb of a mouse 15 days after injections of a lentivirus in the rostral RMS. Note the presence of scattered YFP newborn neurons migrating from the end of the RMS to the different layers of the OB. Most of newborn cells (95%) are granule cells, which soma lie in the Granule Cell Layer (GCL) and dendrites arborize in the External Plexiform Layer (EPL). A small population of newborn neurons migrate up to the most superficial layer of the OB, in the Glomerular Layer (GL). Scale bar, 100μm. Inset, high magnification of a newborn granule cell, showing the membrane expression of ChR2-YFP in the dendrites as well as in spines. Scale bar, 20μm.
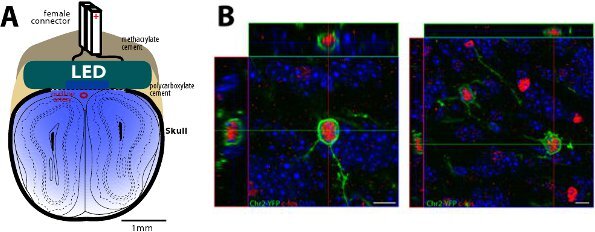 Figure 2. Diagram of implanted LED on the top of the olfactory bulb and light-evoked c-fos expression in ChR2-YFP expressing new granule cells.
A, Schematic drawing of the implanted LED above the olfactory bulbs.
B, Representative confocal images showing double-labelled ChR2-YFP+ c-fos+ granule cells one hour after light stimulation of the olfactory bulb. C-fos expression is revealed using a rabbit anti-c-fos primary antibody (Calbiochem, 1:5000) and A568-conjugated anti-rabbit secondary antibody (Invitrogen, 1:1000). Scale bar, 10μm.
Figure 2. Diagram of implanted LED on the top of the olfactory bulb and light-evoked c-fos expression in ChR2-YFP expressing new granule cells.
A, Schematic drawing of the implanted LED above the olfactory bulbs.
B, Representative confocal images showing double-labelled ChR2-YFP+ c-fos+ granule cells one hour after light stimulation of the olfactory bulb. C-fos expression is revealed using a rabbit anti-c-fos primary antibody (Calbiochem, 1:5000) and A568-conjugated anti-rabbit secondary antibody (Invitrogen, 1:1000). Scale bar, 10μm.
Discussion
Optogenetic tools have recently increased our control over selective neuronal populations, including adult born neurons in the olfactory system. Here we describe how to perform a stereotaxic injection of a lentiviral vector expressing ChR2-YFP in a specific population of adult-born neurons. By carefully monitoring the period post-injection, we are able to analyze and manipulate the activity of a cohort of adult-born neurons with a relative homogeneous age.
Because of their low diffusion spread, their high transduction efficiency and their relative good tropism for neuroblasts of the RMS, lentiviral vectors should be favored compared to others viral vectors such as retroviruses or adeno-associated viruses (AAV). Because they selectively infect dividing cells, such as stem cells or progenitors, retrovirus vectors must be injected into the subventricular zone and transduce a smaller population of newborn neurons. Compared to lentiviral vectors, AAV vectors provide a much higher spread of infection. However, AAV transduction of neuroblasts in the RMS should be avoided because of the transduction of surrounding structures such as the accessory olfactory nucleus, a region that projects axons back into the olfactory bulb. Lastly, use of implanted LEDs can be applied to other superficial structures of the brain such as cortical regions in which ChR2 expression is driven in specific populations of neurons through viral vectors or via a transgenic mouse line10.
With LED implantation, we are able to apply chronic optical stimulation to study the functional role of olfactory bulb adult neurogenesis in awake behaving animals. This simple light source provides a fast temporal control of light thanks to the intrinsically rapid on/off speed of LEDs. In addition to being inexpensive, a miniature LED light source produces sufficient power (~100mW) for ChR2 activation in a large volume of brain tissue11, even bilaterally in the case of olfactory bulbs. This is particularly important in this brain region where significant lesions may be without effect on odor quality perception due to a high degree of redundancy in the inputs from the olfactory epithelium12. Thanks to its power and its spectral bandwidth (25nm), these miniature LEDs may be used for light-evoked excitation using ChR2 as well as others versions of channelrhodopsins such as ChETA or CatCh. Miniature yellow (597nm) and red (624 or 637nm) LEDs are available to be used for red-shifted rhodopsins such as halorhodopsin and VChR113.
Miniature LEDs also show limitations. To avoid heating the brain, a compromise must be found between pulse duration, pulse frequency, and light intensity. For instance, short light pulses of 5 milliseconds are recommended at high frequencies (>20Hz) and light intensity of 100 mW. Above this duration, a significant elevation of the temperature (~1-4°C) in the surrounding tissue can be measured. This parameter should be taken into account for longer light pulses (>5ms) or continuous light stimulation, notably in the case of light-evoked hyperpolarization using halorhodopsin. In addition, the current supplying the LED provides strong electrical artifact, which may preclude in vivo electrophysiological recordings when the LED is in direct contact with the tissue. Laser-coupled optic fibers constitute a possible alternative, allowing a smaller and more restricted optical stimulation. However, the rigidity of optic fibers may interfere with certain behaviors.
After characterizing of the light penetration and the functional stimulation of neurons by light, in vivo optical stimulation can be triggered in combination with behavioral experiments. Special care should be taken regarding the timing of the stimulation. Synchronization of the photostimulation with a specific behavior or task in a behavioral apparatus, notably during operant conditioning, could be decisive for establishing a causal link between cell activation and behavior.
Disclosures
No conflicts of interest declared.
Acknowledgments
This work was supported by the life insurance company "Novalis-Taitbout", Ecole des Neurosciences de Paris (ENP), the Agence Nationale de la Recherche "ANR-09-NEUR-004" in the frame of "ERA-NET NEURON" of FP7 program by the European Commission, "Foundation pour la Recherche Medicale", the Letten Foundation and the Pasteur Foundation.
References
- Lledo PM. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 2006;7:179–193. doi: 10.1038/nrn1867. [DOI] [PubMed] [Google Scholar]
- Alonso M. Turning astrocytes from the rostral migratory stream into neurons: a role for the olfactory sensory organ. J. Neurosci. 2008;28:11089–11102. doi: 10.1523/JNEUROSCI.3713-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mouret A. Learning and survival of newly generated neurons: when time matters. J. Neurosci. 2008;28:11511–11516. doi: 10.1523/JNEUROSCI.2954-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bardy C. How, when, and where new inhibitory neurons release neurotransmitters in the adult olfactory bulb. J. Neurosci. 2010;30:17023–17034. doi: 10.1523/JNEUROSCI.4543-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haubensak W. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature. 2010;468:270–276. doi: 10.1038/nature09553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang F. Multimodal fast optical interrogation of neural circuitry. Nature. 2007;446:633–639. doi: 10.1038/nature05744. [DOI] [PubMed] [Google Scholar]
- Wang X, McManus M. Lentivirus Production. J. Vis. Exp. 2009. pp. e1499–e1499. [DOI] [PMC free article] [PubMed]
- Cardin JA. Targeted optogenetic stimulation and recording of neurons in vivo using cell-type-specific expression of Channelrhodopsin-2. Nature protocols. 2010;5:247–254. doi: 10.1038/nprot.2009.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber D. Sparse optical microstimulation in barrel cortex drives learned behaviour in freely moving mice. Nature. 2010;451:61–64. doi: 10.1038/nature06445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arenkiel BR. In vivo light-induced activation of neural circuitry in transgenic mice expressing channelrhodopsin-2. Neuron. 54:205–218. doi: 10.1016/j.neuron.2007.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albeanu DF. LED arrays as cost effective and efficient light sources for widefield microscopy. PLoS ONE. 2008;3:2146–2146. doi: 10.1371/journal.pone.0002146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slotnick B, Bodyak N. Odor Discrimination and Odor Quality Perception in Rats with Disruption of Connections between the Olfactory Epithelium and Olfactory Bulbs. J. Neurosci. 2002;22:4205–4216. doi: 10.1523/JNEUROSCI.22-10-04205.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gradinaru V. Molecular and cellular approaches for diversifying and extending optogenetics. Cell. 2010;141:154–165. doi: 10.1016/j.cell.2010.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valley M, Wagner S, Gallarda BW, Lledo P. Using Affordable LED Arrays for Photo-Stimulation of Neurons. J. Vis. Exp. 2011. pp. e3379–e3379. [DOI] [PMC free article] [PubMed]