

Abstract
The bisdichloroacetyldiamine WIN 18,446 reversibly inhibits spermatogenesis in many species, including humans; however, the mechanism by which WIN 18,446 functions is unknown. As retinoic acid is essential for spermatogenesis, we hypothesized that WIN 18,446 might inhibit retinoic acid biosynthesis from retinol (vitamin A) within the testes by inhibiting the enzyme aldehyde dehydrogenase 1a2 (ALDH1a2). We studied the effect of WIN 18,446 on ALDH1a2 enzyme activity in vitro, and on spermatogenesis and fertility in vivo, in mature male rabbits for 16 weeks. WIN 18,446 markedly inhibited ALDH1a2 enzyme activity in vitro with an IC50 of 0.3 μM. In vivo, the oral administration of 200 mg/kg WIN 18,446 to male rabbits for 16 weeks significantly reduced intratesticular concentrations of retinoic acid, severely impaired spermatogenesis, and caused infertility. Reduced concentrations of intratesticular retinoic acid were apparent after only 4 weeks of treatment and preceded the decrease in sperm counts and the loss of mature germ cells in tissue samples. Sperm counts and fertility recovered after treatment was discontinued. These findings demonstrate that bisdichloroacetyldiamines such as WIN 18,446 reversibly suppress spermatogenesis via inhibition of testicular retinoic acid biosynthesis by ALDH1a2. These findings suggest that ALDH1a2 is a promising target for the development of a reversible, nonhormonal male contraceptive.
Keywords: Retinol, vitamin A, male contraception, sperm concentration
Almost 50 years ago, the oral administration of the bisdichloroacetyldiamine WIN 18,446 was shown in man to safely and reversibly inhibit spermatogenesis (Heller et al, 1961), raising hope that it might be useful as a male contraceptive. This compound was initially being developed as an amebicide (Berberian et al, 1961), but was noted to cause marked impairment of spermatogenesis in rodents, without significant effects on other tissues (Coulston et al, 1960). Follow-up investigations in dogs and rhesus monkeys demonstrated that oral administration of WIN 18,446 led to a complete arrest of spermatogenesis in testicular biopsies (Beyler et al, 1961; Heller et al, 1963). Notably, WIN 18,446 had no effect on serum testosterone concentrations, implying that the mechanism by which WIN 18,446 suppressed spermatogenesis was not hormonal in nature. More recently, WIN 18,446 was found to be a safe, effective, and reversible oral contraceptive in male wolves (Asa et al, 1996), cats (Munson et al, 2004), and shrews (Singh and Dominic, 1980). Unfortunately, the utility of WIN 18,446 as a contraceptive in man is hampered by the fact that men taking WIN 18,446 experience a “disulfiram reaction” when they drink alcohol. Because of this, further development of WIN 18,446 as a male contraceptive was abandoned without an understanding of its mechanism of action.
Disulfiram (Antabuse) is a medication used in the management of alcoholism (Suh et al, 2006). Individuals taking disulfiram feel well as long as they do not drink alcohol. If they do imbibe, they experience a disulfiram reaction, which is characterized by flushing, diaphoresis, dysphoria, nausea, vomiting, and other symptoms. This reaction is mediated by disulfiram's inhibition of the liver enzyme aldehyde dehydrogenase-2 (ALDH2). ALDH2 catalyzes the conversion of acetaldehyde, the main metabolite of ethanol, to acetic acid (Heilig and Egli, 2006). The combination of disulfiram and alcohol therefore leads to increased serum concentrations of acetaldehyde and the symptoms of the disulfiram reaction (Deitrich et al, 2007). The presence of a disulfiram reaction when WIN 18,446 and alcohol are coadministered strongly suggests that WIN 18,446 also inhibits ALDH2.
Nineteen isozymes of aldehyde dehydrogenase have been identified (Marchitti et al, 2008). ALDH2, the isozyme involved in alcohol metabolism, is expressed in many tissues, including the liver, lung, and kidney (Steward et al, 1996). In 1996, a novel aldehyde dehydrogenase called aldehyde dehydrogenase 1a2 (ALDH1a2) was cloned from the rat and found to be expressed almost exclusively in the testes (Wang et al, 1996). ALDH1a2 localizes to developing germ cells within the seminiferous tubules and strongly binds retinal, but does not recognize acetaldehyde as a substrate, suggesting that ALDH1a2 functions in testicular retinoic acid biosynthesis (Wang et al, 1996; Vernet et al, 2006a).
It has been recognized since 1925 that vitamin A is required for spermatogenesis (Wolbach and Howe, 1925). Vitamin A is converted in tissues to its principal biologically active derivative, retinoic acid. In the testes, retinoic acid is necessary both for the initiation of spermatogenesis at puberty and for the maintenance of spermatogenesis in adults (Lufkin et al, 1993; Ghyselinck et al, 2006; Vernet et al, 2006b). Dietary beta-carotene and retinol are transported to the testes, where they are converted to retinoic acid within the developing germ cells by alcohol and aldehyde dehydrogenases (Bishop and Griswold, 1987; Napoli, 2000; Paik et al, 2004). Retinoic acid then binds one of several retinoic acid receptors that regulate gene expression (Chung et al, 2004; Bowles et al, 2006). Among other effects, retinoic acid induces the spermatogonial differentiation protein stimulated by retinoic acid-8 (STRA8) within the developing germ cells. STRA8 appears to play a central role in spermatogonial differentiation (Zhou et al, 2008).
As testicular retinoic acid biosynthesis requires ALDH1a2, and this enzyme is structurally similar to the one inhibited by WIN 18,446 in the disulfiram reaction (ALDH2), we hypothesized that WIN 18,446 suppressed spermatogenesis via inhibition of ALDH1a2. Therefore, we studied the effect of WIN 18,446 on ALDH1a2 enzyme activity in vitro and on spermatogenesis in vivo in adult male New Zealand white rabbits. In this way, we sought to ascertain if suppression of spermatogenesis by WIN 18,446 was mediated by the inhibition of testicular retinoic acid biosynthesis.
Materials and Methods
In Vitro Inhibition of ALDH1a2 by WIN 18,446
To produce cells expressing high levels of ALDH1a2 activity, a cDNA for ALDH1a2 was cloned into lentiviral plasmids under the control of EF1a promoter and virally transduced into the lung cancer cell line H1229, which expresses little ALDH activity at baseline (Zaiss et al, 2002). Expression of ALDH1a2 was then verified by Western blot analysis as described previously (Moreb et al, 2002). Inhibition of ALDH1a2 by WIN 18,446 (N,N′-bis(dichloroacetyl)-1,8-octanediamine; Acros Organics, Geel, Belgium) was determined using lysate of the transduced H1229 cells. Lysate was incubated with 10 μM all-trans-retinal in the presence of varying concentrations of WIN 18,446 for 30 minutes at 37°C. The reaction was stopped by adding 100% ethanol containing 0.025 M potassium hydroxide. Retinoic acid was extracted and measured by high-performance liquid chromatography as described previously (Kane et al, 2005; Paik et al, 2005). Loss during extraction was accounted for by adjusting the recovery to that of known concentrations of the internal standard, acitretin (Sigma-Aldrich, St Louis, Missouri).
Spermatogenesis in Rabbits
All procedures in rabbits were approved in advance by the Institutional Animal Use and Care Committee at the University of Washington. Six-month-old male New Zealand white rabbits were dosed orally for 4, 8, and 16 weeks (n = 4/group) with 200 mg/kg WIN 18,446 daily, as this dose had previously been shown to be effective at suppressing spermatogenesis in a variety of other species (Coulston et al, 1960; Beyler et al, 1961; Heller et al, 1963; Singh and Dominic, 1980; Asa et al, 1996; Munson et al, 2004), and could be used to determine the effect of WIN 18,446 administration on intratesticular retinoic acid biosynthesis and function during suppression of spermatogenesis. The WIN 18,446 was compounded in a simple syrup base at a concentration of 100 mg/mL with banana crème flavoring and dosed orally using a syringe. Spermatogenesis was assessed by having the animals ejaculate sperm into an artificial vagina at baseline and weekly during treatment and recovery (Naughton et al, 2003). Sperm count, concentration, and motility were quantified manually using a hemocytometer (Seed et al, 1996). Two chambers were counted and averaged for each time point. At baseline, and after 4, 8, and 16 weeks of treatment, anesthetized rabbits underwent a unilateral orchiectomy to ascertain the impact of treatment on spermatogenesis by histology, intratesticular retinoic acid concentrations, and Stra8 expression. For histological examinations, approximately 0.1–0.3 g testis was placed in Bouin solution for fixation and paraffin embedment, sectioned at 5 μm, and stained with hematoxylin–periodic acid-Schiff stain. For testicular and epididymal sperm counts, approximately 0.3–0.6 g of the testis and 0.1–0.3 g of the cauda epididymis were individually weighed and homogenized in 0.1 M sodium phosphate buffer, pH 7.4, containing 0.1% Triton X-100, using an all-glass Kontes 15- mL homogenizer. Testes were homogenized with 8 strokes and the epididymis was homogenized with 15 strokes. Ten microliters of the homogenate was loaded into each side of a Neubauer phase contrast hemocytometer and counted. Total homogenization-resistant spermatids or epididymal sperm per gram or per organ were calculated by correcting for the number of squares counted, dilution, volume, and weight (Amann, 1986).
For the measurement of tissue retinoic acid, tissues were homogenized on ice with ice-cold 0.9% saline to make a 10% homogenate using an OMNI International homogenizer (10 strokes at 1500 rev/min). Up to 500 μL of tissue homogenate was combined with 10 mL internal standard (10 nM D5-dimethyl retinoic acid in acetonitrile) and extracted with 2 mL dichloromethane. Following the complete evaporation of dichloromethane, the organic residue was reconstituted with 60 mL of acetonitrile. Resolution of the retinoids was obtained with an extended C-18 Agilent ZORBAX column (100 mm × 2.1 mm, 3 mm) using a Waters Quattro Premier XE liquid chromatography/tandem mass spectrometry with an injection volume of 20 mL.
For the measurement of testicular Stra8 mRNA expression, total RNA was isolated from testes homogenized in Trizol (Invitrogen, Carlsbad, California) according to the manufacturer's protocol. Four testes were used for each time point. One microgram of total RNA was reverse transcribed using the iScript kit (BioRad, Hercules, California). Quantitative PCR was performed by using Fast SYBR Green Mastermix (Applied Biosystems, Foster City, California) on an ABI7500 Fast Real-Time PCR instrument (Applied Biosystems). Stra8 primers amplify a 154-bp product (primers: 5′-GATGCTGGGGA GAAGTTTCA-3′ and 5′-AATCGTCGTCATCGAAGGTC-3′); control Rps2 primers amplify a 112-bp product (primers: 5′-CTGACTCCCGACCTCTGGAAA-3′ and 5′-GAGCCTGGG TCCTCTGAACA-3′). Results were analyzed using the Δ-Δ Ct method with Rps2 as the normalization control (Zhou et al, 2008).
Fertility was assessed by mating experiments. Six-month-old females were placed with treated animals both after 16 weeks of treatment and after recovery from treatment, and copulation was observed. Fifteen days after mating, the females were sacrificed and the number of embryos counted.
Statistical Analysis
For the analysis of the testicular spermatids, Stra8 expression, and retinoic acid concentrations, as well as the sperm parameters in the ejaculates, mean values were calculated at each time point and compared with baseline by the Wilcoxon signed rank test. No corrections were made for multiple comparisons. For assessment of fertility, the number of embryos between treatment and recovery sired by a given male were compared using a 2-sample t-test with unequal variances. STATA (version 8.0; College Park, Texas) was used for all statistical analysis. For all comparisons, an α of ≤ .05 was considered significant.
Results
In Vitro Inhibition of ALDH1a2 by WIN 18,446
WIN 18,446 potently inhibited the ability of ALDH1a2 to convert retinal (10 μM) to retinoic acid in vitro with an IC50 of 0.3 μM (Figure 1). Concentrations of WIN 18,446 greater than 1 μM suppressed over 75% of the formation from retinal under these assay conditions, demonstrating that WIN 18,446 is a strong inhibitor of retinal oxidation. WIN 18,446 also potently inhibited ALDH2 activity in a similar fashion (data not shown).
Figure 1.
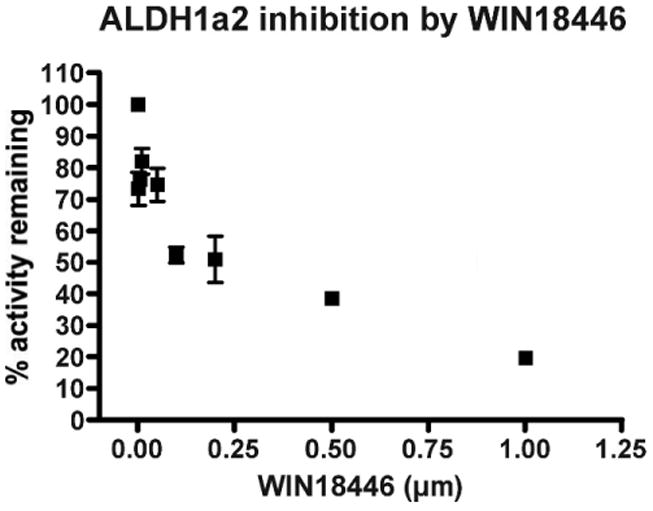
WIN 18,446 is a potent inhibitor of the testicular retinal dehydrogenase, ALsDH1a2. Lysate from H1229 cells transduced with ALDH1a2 was incubated for 30 minutes with 10 μM all-trans-retinal in the presence of various concentrations of WIN 18,446, and retinoic acid was measured by high-performance liquid chromatography. All experiments were conducted in triplicate. Values are means ± SEM (note that error bars are contained within the symbol in certain instances).
Spermatogenesis in Rabbits
Treatment of rabbits with WIN 18,446 at an oral daily dose of 200 mg/kg resulted in marked suppression of testicular weight over time (Table 1). After 16 weeks of treatment, testicular weight was reduced almost 75% compared with untreated controls. Conversely, treatment with WIN 18,446 had no apparent effect on epididymal weight compared with baseline.
Table 1.
Organ weight and testicular and epididymal spermatid counts in New Zealand white rabbits dosed orally with 200 mg/kg WIN 18,446 daily for 4, 8, or 16 weeks (n = 4/group); all values are means ±SEM
| Treatment Duration, wk | Testicular Weight, g | Epididymal Weight, g | Testicular Spermatids, millions | Cauda Epididymal Spermatids, millions |
|---|---|---|---|---|
| 0, control | 3.6 ± 0.1 | 1.4 ± 0.1 | 380 ± 49 | 506 ± 118 |
| 4 | 2.3 ± 0.2a | 1.5 ± 0.2 | 277 ± 26 | 389 ± 96 |
| 8 | 1.3 ± 0.1b | 1.4 ± 0.1 | 78 ± 62a | 55 ± 23b |
| 16 | 0.9 ± 0.1c | 1.6 ± 0.2 | 3 ± 0.6c | 5.5 ± 4.0c |
P < .05 compared with baseline.
P < .01 compared with baseline.
P < .001 compared with baseline.
Treatment of rabbits with WIN 18,446 dramatically suppressed spermatogenesis, both as measured by testicular and epididymal sperm counts from orchiectomy specimens (Table 1) as well as in ejaculated sperm from the 3 animals that were able to ejaculate into the artificial vagina (Figure 2). Interestingly, there were no significant reductions in any measure of spermatogenesis after 4 weeks of treatment. After 8 weeks of treatment, however, significant reductions in ejaculated sperm concentrations and testicular and epididymal sperm counts were evident. After 16 weeks of treatment, sperm was absent from the ejaculates in all treated animals and very few spermatids were present in either the testes or the epididymides (Table 1). Sperm motility was similarly affected, dropping from greater than 75% at baseline and after 4 weeks of treatment to 14% after 8 weeks of treatment and 0% (epididymal sperm) after 16 weeks of treatment (data not shown). Ejaculated sperm concentrations remained severely suppressed for 8–10 weeks after treatment was discontinued. However, thereafter sperm concentrations rapidly increased to the anticipated 50% of baseline values in all animals. This level takes into account the 50% reduction in spermatogenetic capacity attributable to the hemiorchiectomy performed at the end of the treatment phase.
Figure 2.

Sperm concentrations over time in New Zealand white rabbits dosed orally with 200 mg/kg WIN 18,446 daily for 16 weeks (N = 3). All animals were azoospermic between week 16 of treatment and week 8 of recovery. Animals underwent a hemiorchiectomy at the end of treatment, which accounts for the 50% reduction in sperm output at the end of recovery. *indicates P < .05 compared with baseline.
Despite observed episodes of copulation, none of the animals were able to sire a pregnancy after 16 weeks of treatment. However, after recovery of spermatogenesis and 16 weeks after stopping treatment with WIN 18,446, 3 of 4 animals successfully impregnated females (mean embryo number of 6.6 ± 3.5). The fourth animal did not impregnate a female; however, this animal exhibited no mounting behavior either at the end of treatment or during recovery, suggesting a coital disorder in this animal. As this was the same animal that was unable to use the artificial vagina, and we wished to document a return of normal spermatogenesis in all animals, this rabbit's other testis was removed and examined. The testicular and epididymal weights and testicular and epididymal sperm counts and sperm motility were very similar to those of the untreated controls (data not shown).
Histological examination of testicular tissue from rabbits treated with WIN 18,446 revealed no obvious changes after 4 weeks of treatment (Figure 3C and D), compared with controls (Figure 3A and B). By 8 weeks of treatment, however, roughly one-half of the tubules revealed marked evidence of spermatogenic arrest and hypospermatogenesis (Figure 3E and F). After 16 weeks of treatment, no active spermatogenesis was apparent (Figure 3G and H). Instead, within the seminiferous tubules only spermatogonia and Sertoli cells were identifiable, implying that treatment was blocking the ability of spermatogonia to enter into meiosis.
Figure 3.
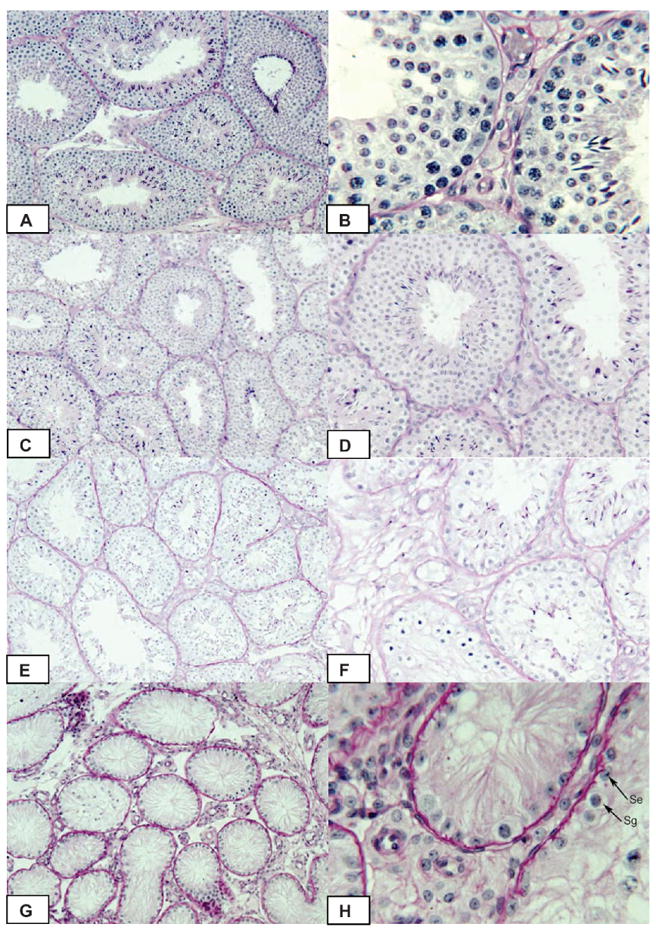
Testicular histology in New Zealand white rabbits prior to treatment (A, B) or after daily treatment with 200 mg/kg WIN 18,446 orally for 4 (C, D), 8 (E, F) or 16 weeks (G, H). Se indicates Sertoli cell; Sg, spermatogonia in Figure 3H (arrows). Original magnification ×100 (A, C, E, G); ×400 (B, D); ×200 (F, H).
Intratesticular concentrations of retinoic acid were reduced compared with baseline by approximately 60% at all time points during treatment with WIN 18,446 (Figure 4), implying a block in testicular retinoic acid biosynthesis. In contrast, testicular expression of the retinoic acid regulated gene Stra8 initially increased (Figure 5). However, after 8 and 16 weeks of treatment, Stra8 expression was significantly suppressed compared with baseline.
Figure 4.
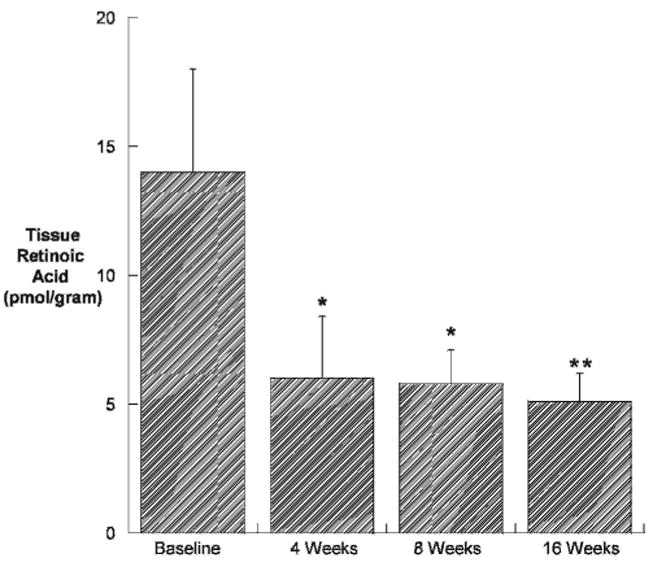
Testicular tissue retinoic acid concentration (pmol/g tissue) in New Zealand white rabbits dosed orally with 200 mg/kg WIN 18,446 daily (n = 4/group). All values are means ± SEM. *indicates P < .05 compared with placebo; **, P < .01.
Figure 5.
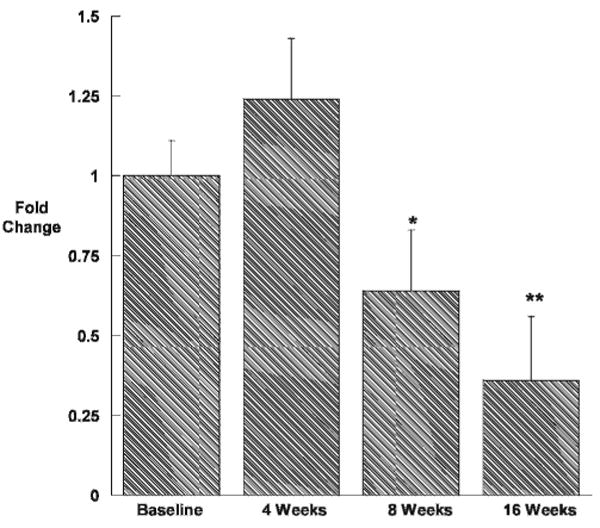
Expression of the retinoic acid–induced gene Stra8 in the testes of New Zealand white rabbits dosed orally with WIN 18,446 200 mg/kg daily (n = 4/group). All values are means ± SEM. *indicates P < .05 compared with placebo; **, P < .01.
Treatment with WIN 18,446 did not have any significant effect on serum concentrations of testosterone (Table 2). In addition, treatment did not have any apparent effect on blood counts or indices of liver or kidney function. The white blood cell count was significantly decreased compared with baseline 8 weeks after treatment was discontinued; however, the white blood cell count returned to normal at the end of recovery. Average animal weight decreased by 5%–10% during treatment without a significant change in serum albumin, but weight returned to baseline after treatment was discontinued. Animals exhibited normal behavior, including sexual behavior, during treatment.
Table 2.
Blood counts, liver and kidney tests, and serum testosterone in New Zealand white rabbits dosed orally with 200 mg/kg of WIN 18,446 daily for 16 weeks (n = 4); all values are means ±SEM
| Treatment | Recovery | ||||||
|---|---|---|---|---|---|---|---|
| Baseline | 4 wk | 8 wk | 12 wk | 16 wk | 8 wk | 16 wk | |
| Hematology | |||||||
| Hematocrit, % | 37 ± 1.0 | 37 ± 0.8 | 37 ± 0.7 | 39 ± 0.9 | 37 ± 1.2 | 38 ± 0.8 | 36 ± 0.8 |
| White blood cell count, ×1000/μL | 7.5 ± 0.4 | 10 ± 0.4 | 9.2 ± 0.2 | 9.1 ± 0.5 | 9.6 ± 0.1 | 6.1 ± 0.5* | 7.1 ± 0.5 |
| Platelet count, ×1000/μL | 260 ± 41 | 207 ± 37 | 222 ± 21 | 222 ± 22 | 194 ± 32 | 214 ± 17 | 240 ± 14 |
| Metabolic | |||||||
| Creatinine, mg/dL | 1.1 ± 0.1 | 1.2 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 | 1.2 ± 0.1 | 1.2 ± 0.1 |
| Glucose, mg/dL | 150 ± 11 | 140 ± 3 | 136 ± 3 | 147 ± 13 | 160 ± 7 | 143 ± 11 | 138 ± 7 |
| Albumin, g/dL | 3.8 ± 0.1 | 3.8 ± 0.1 | 3.9 ± 0.1 | 3.8 ± 0.1 | 3.8 ± 0.1 | 3.9 ± 0.1 | 3.9 ± 0.1 |
| ALT, U/L | 35 ± 6 | 30 ± 6 | 30 ± 6 | 28 ± 6 | 30 ± 5 | 31 ± 6 | 30 ± 6 |
| Cholesterol, mg/dL | 18 ± 2 | 19 ± 2 | 16 ± 2 | 18 ± 2 | 15 ± 1 | 23 ± 3 | 19 ± 3 |
| Hormonal | |||||||
| Testosterone, ng/mL | 0.6 ± 0.3 | 1.9 ± 0.5 | 0.6 ± 0.3 | 0.6 ± 0.3 | 0.8 ± 0.3 | 0.5 ± 0.3 | 0.4 ± 0.1 |
Abbreviation: ALT = alanine aminotransferase.
P < .05 compared with baseline.
Discussion
In this work, for the first time, we elucidate the mechanism by which bisdichloroacetyldiamines such as WIN 18,446 suppress spermatogenesis. Specifically, we conclude that these compounds block spermatogenesis by inhibiting testicular retinoic acid biosynthesis. This conclusion is supported by the demonstration that WIN 18,446 potently inhibits the testes-specific enzyme ALDH1a2, which is involved in testicular retinoic acid biosynthesis. In addition, we have shown that the oral administration of WIN 18,446 to male rabbits significantly reduces intratesticular concentrations of retinoic acid and Stra8 expression and very severely suppresses spermatogenesis and fertility. It is possible that the observed reductions in retinoic acid and Stra8 expression could be due, in part, to the reduction in the number of testicular spermatids present during treatment. However, the observation that the reduction in testicular retinoic acid concentration occurred before any apparent reduction in spermatogenesis strongly supports the hypothesis that the reduction in spermatogenesis was a result of the inhibition of retinoic acid biosynthesis and not due to some other cause. Taken together, these findings suggest that WIN 18,446 reversibly suppresses spermatogenesis via inhibition of testicular retinoic acid biosynthesis by ALDH1a2, and identifies ALDH1a2 as a promising target for the development of novel, nonhormonal male contraceptives. Furthermore, this finding raises the possibility that a defect in the biosynthetic pathway of retinoic acid could underlie some causes of male infertility.
The finding that bisdichloroacetyldiamines suppress spermatogenesis by inhibition of testicular retinoic acid biosynthesis makes sense in light of emerging knowledge about the importance of retinoic acid in spermatogenesis. For example, retinoic acid appears to promote spermatogonia to enter into the meiotic pathway by up-regulating the expression of Kit in germ cells while also increasing expression of Kit ligand in Sertoli cells (Pellegrini et al, 2008). Furthermore, the absence of retinoic acid signaling is markedly deleterious to spermatogenesis. For example, retinoic-acid receptor-α knockout males are sterile because of degenerating or abnormal spermatogonia (Lufkin et al, 1993). This appears to be due in part to aberrant coupling between cells within the seminiferous tubules, which may be secondary to reduced expression of the junctional complex constituents connexin-40 and vimentin (Chung et al, 2010). Similarly, mice deficient in either RARγ or RXRβ are infertile, owing to low sperm counts and spermiation failure, respectively (Lohnes et al, 1993; Kastner et al, 1996). Moreover, retinoic acid receptor antagonists such as BMS-189453 potently inhibit spermatogenesis after oral doses as low as 5 mg/kg, with mice becoming temporarily infertile after 4 weeks of treatment (Schulze et al, 2001).
Given the importance of retinoic acid in spermatogenesis, one might wonder why WIN 18,446 takes so long to exhibit its contraceptive effect. It is tempting to speculate that tubules either have variable stores of retinoic acid (likely bound to cellular binding proteins) or only require it at a given stage of the spermatogenic cycle. This theory would explain the variability in tubular appearance after 8 weeks of treatment (Figure 3E and F). Perhaps tubules lacking access to retinoic acid exhibit hypospermatogenesis before tubules that have adequate stores of retinoic acid or had recently completed the stage in the spermatogenic cycle that required retinoic acid before treatment began.
In addition, it is somewhat surprising that WIN 18,446 treatment reduced testicular retinoic acid concentrations only to approximately 40% of baseline values. It is possible that testicular retinoic acid is partially synthesized via non-ALDH1a2 pathways, possibly via ALDH1a1, which is also expressed in testicular tissue (Steward et al, 1996). Alternatively, retinoic acid from other tissues could be diffusing into the testes; however, less than 1% of testicular retinoic acid is derived from the circulation (Kurlandsky et al, 1995). Moreover, this finding may imply that spermatogenesis requires a certain threshold concentration of retinoic acid within the testes. Determining the minimal concentration of testicular retinoic acid required for spermatogenesis will be the subject of future research.
Importantly, treatment with WIN 18,446 did not have any toxic effects, as measured by tests of hematopoiesis and liver and kidney function; however, animals did lose 5%–10% of their body weight in the first several weeks of treatment. A slight decrease in the body weights of male cats treated with WIN 18,446 was also observed (Munson et al, 2004), and humans treated with this compound complained of upset stomachs, without changes in weight (Heller et al, 1961). Alternatively, the loss in body weight observed in our study could have been due to the stress of daily oral dosing of the medication, which has been observed in other settings with rabbits (Foote and Carney, 2000).
Interestingly, and despite a 75% reduction in testicular volume, there was no apparent change in the serum testosterone concentrations during treatment. This observation implies that the change in the volume of the testis was mostly attributable to the absence of maturing sperm in the seminiferous tubules. Reductions in testicular volume are observed in humans during treatment with male hormonal contraceptives (Wu et al, 1996). In these trials the epididymal volume is also reduced, as the epididymis is an androgen-dependent organ (Goyal et al, 1994), and hormonal contraceptives markedly reduce intratesticular concentrations of testosterone (Page et al, 2007). In contrast, epididymal weight did not change during treatment with WIN 18,446, implying that local concentrations of testosterone remained normal during treatment. The lack of change in serum testosterone and epididymal weight demonstrates that WIN 18,446 functions as a contraceptive without impacting the hypothalamic–pituitary–Leydig cell axis. Such a nonhormonal contraceptive could have significant appeal, as men could be reassured that their testosterone levels would be unaffected by treatment.
Importantly, WIN 18,446 exhibited almost no discernable toxic effects in vivo in experimental animals aside from its effect on the testes. The LD50 after oral administration of WIN 18,446 exceeds 10 000 mg/kg, and there were no deaths after the intraperitoneal injection of 750 mg/kg of WIN 18,446 in rats (Coulston et al, 1960). Although WIN 18,446 is likely safe in adult animals, WIN 18,446 and other bisdichloroacetyldiamines have teratogenic effects on rat and chicken embryos (Tasaka et al, 1991; Choy et al, 1999), similar to those observed with retinoids (Nau, 2001). Therefore, care must be taken to avoid exposure of pregnant women to these compounds.
There is a great need for new approaches to preventing unintended pregnancy. Despite currently available contraceptives, the world's population exceeds 6.9 billion and is increasing by 80 million yearly (Department of Economic and Social Affairs, 2010). Much of this population growth is unintended (Henshaw, 1998). Therefore, there is a great need for better access to existing contraceptives, improved contraceptive education, and more contraceptive options. Currently, male-directed contraceptive options are particularly limited, and despite decades of research into hormonal and immunological methods of male contraception, no regimen based on either of these approaches is near clinical approval. Bisdichloroacetyldiamines, such as WIN 18,446, safely and effectively suppress spermatogenesis in men, but cause a disulfiram reaction when coadministered with alcohol, which prevented their introduction for contraceptive purposes. We have now demonstrated that the contraceptive effect of these compounds is mediated by inhibition of testicular retinoic acid biosynthesis via the testes-specific enzyme ALDH1a2. Hopefully, this finding will allow for the development of novel, specific inhibitors of ALDH1a2 that do not cause the disulfiram reaction and can be developed into a safe, effective, and reversible form of male contraception.
Acknowledgments
Supported by the Eunice Kennedy Shriver National Institute of Child Health and Human Development, a division of the National Institute of Health, through cooperative agreement U01 HD060408 and, in part, by NIH grant R01GM081569-02S1.
We thank Ms Constance Pete and Ms Pilar Cordero for assistance with sperm analysis; William Bremner, MD, PhD, and Alvin M. Matsumoto, MD, for critical review of the manuscript; Stu Sommerville for medication compounding; and the University of Washington Veterinary Services for animal care and assistance.
References
- Amann RP. Detection of alterations in testicular and epididymal function in laboratory animals. Environ Health Perspect. 1986;70:149–158. doi: 10.1289/ehp.8670149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asa C, Zaneveld LJD, Munson L, Callahan M, Byers AP. Efficacy, safety and reversibility of a bisdiamine male-directed oral contraceptive in grey wolves (Canis lupus) J Zoo Wildl Med. 1996;27:501–506. [Google Scholar]
- Berberian DA, Slighter RG, Surrey AR. In vitro and in vivo amebicidal activity of N, N′Bis(dichloroacetyl)diamines. Antibiot Chemother. 1961;11:245–255. [Google Scholar]
- Beyler AL, Potts GO, Coulston F, Surrey AR. The selective testicular effects of certain bis-(dichloroacetyl) diamines. Endocrinology. 1961;69:819–833. [Google Scholar]
- Bishop PD, Griswold MD. Uptake and metabolism of retinol in cultured Sertoli cells: evidence for a kinetic model. Biochemistry. 1987;26:7511–7518. doi: 10.1021/bi00397a046. [DOI] [PubMed] [Google Scholar]
- Bowles J, Knight D, Smith C, Wilhelm D, Richman J, Mamiya S, Yashiro K, Chawengsaksophak K, Wilson MJ, Rossant J, Hamada H, Koopman P. Retinoic signaling determines cell fate in mice. Science. 2006;312:596–600. doi: 10.1126/science.1125691. [DOI] [PubMed] [Google Scholar]
- Choy M, Oltjen SL, Moon AJ, Armstrong MT, Armstrong PB. Bisdiamine inhibits extracellular matrix formation and cell proliferation of atrioventricular mesenchyme from developing chick heart valves. Teratology. 1999;59:148–155. doi: 10.1002/(SICI)1096-9926(199903)59:3<148::AID-TERA6>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- Chung SS, Choi C, Wang X, Hallock L, Wolgemuth DJ. Aberrant distribution of junctional complex components in retinoic acid receptor alpha-deficient mice. Micro Res Tech. 2010;73:583–596. doi: 10.1002/jemt.20797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung SS, Sung W, Wang X, Wolgemuth DJ. Retinoic acid receptor alpha is required for synchronization of spermatogenic cycles and its absence results in progressive breakdown of the spermatogenic process. Dev Dyn. 2004;230:754–766. doi: 10.1002/dvdy.20083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coulston F, Beyler AL, Drobeck HP. The biologic actions of a new series of bis(dichloroacetyl)diamines. Toxicol Appl Pharmacol. 1960;2:715–721. doi: 10.1016/0041-008x(60)90088-0. [DOI] [PubMed] [Google Scholar]
- Deitrich RA, Petersen D, Vasiliou V. Removal of acetaldehyde from the body. Novartis Found Symp. 2007;285:23–40. doi: 10.1002/9780470511848.ch3. [DOI] [PubMed] [Google Scholar]
- Department of Economic and Social Affairs of the United Nations Secretariat. World population prospects: the 2008 revision. [Accessed February 24];2010 http://esa.un.org/unpp.
- Foote RH, Carney EW. The rabbit as a model for reproductive and developmental toxicity studies. Reprod Toxicol. 2000;14:477–493. doi: 10.1016/s0890-6238(00)00101-5. [DOI] [PubMed] [Google Scholar]
- Ghyselinck NB, Vernet N, Dennefeld C, Giese N, Nau H, Chambon P, Viville S, Mark M. Retinoids and spermatogenesis: lessons from mutant mice lacking the plasma retinol binding protein. Dev Dyn. 2006;235:1608–1622. doi: 10.1002/dvdy.20795. [DOI] [PubMed] [Google Scholar]
- Goyal HO, Hutto V, Maloney MA. Effects of androgen deprivation in the goat epididymis. Acta Anat. 1994;150:127–135. doi: 10.1159/000147611. [DOI] [PubMed] [Google Scholar]
- Heilig M, Egli M. Pharmacological treatment of alcohol dependence: target symptoms and target mechanisms. Pharmacol Ther. 2006;111:855–876. doi: 10.1016/j.pharmthera.2006.02.001. [DOI] [PubMed] [Google Scholar]
- Heller CG, Flageolle BY, Matson LJ. Histopathology of the human testes as affected by Bis (dichloroacetyl) diamines. Exp Mol Pathol. 1963;2:107–114. [PubMed] [Google Scholar]
- Heller CG, Moore DJ, Paulsen CA. Suppression of spermatogenesis and chronic toxicity in men by a new series of bis (dichloroacetyl) diamines. Toxicol Appl Pharmacol. 1961;3:1–11. doi: 10.1016/0041-008x(61)90002-3. [DOI] [PubMed] [Google Scholar]
- Henshaw SK. Unintended pregnancy in the US. Fam Plann Perspect. 1998;30:24–29. [PubMed] [Google Scholar]
- Kane MA, Chen N, Sparks S, Napoli JL. Quantification of endogenous retinoic acid in limited biological samples by LC/MS/MS. Biochem J. 2005;388:363–369. doi: 10.1042/BJ20041867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kastner P, Mark M, Leid M, Gansmuller A, Chin W, Grondona JM, Décimo D, Krezel W, Dierich A, Chambon P. Abnormal spermatogenesis in RXR beta mutant mice. Genes Dev. 1996;10:80–92. doi: 10.1101/gad.10.1.80. [DOI] [PubMed] [Google Scholar]
- Kurlandsky SB, Gamble MV, Ramakirshnan R, Blaner WS. Plasma delivery of retinoic acid to tissues in the rat. J Biol Chem. 1995;270:17850–17857. doi: 10.1074/jbc.270.30.17850. [DOI] [PubMed] [Google Scholar]
- Lohnes D, Kastner P, Dierich A, Mark M, LeMeur M, Chambon P. Function of retinoic acid receptor gamma in the mouse. Cell. 1993;73:643–658. doi: 10.1016/0092-8674(93)90246-m. [DOI] [PubMed] [Google Scholar]
- Lufkin T, Lohnes D, Mark M, Dierich A, Gorry P, Gaub MP, LeMeur M, Chambon P. High postnatal lethality and testis degeneration in retinoic acid receptor alpha mutant mice. Proc Natl Acad Sci U S A. 1993;90:7225–7229. doi: 10.1073/pnas.90.15.7225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchitti SA, Brocker C, Stagos D, Vasiliou V. Non-P450 aldehyde oxidizing enzymes: the aldehyde dehydrogenase superfamily. Expert Opin Drug Metab Toxicol. 2008;4:697–720. doi: 10.1517/17425250802102627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreb JS, Zucali JR, Ostmark B, Benson NA. Heterogeneity of aldehyde dehydrogenase expression in lung cancer cell lines is revealed by Aldefluor flow cytometry-based assay. Cytometry B. 2002;72:281–289. doi: 10.1002/cyto.b.20161. [DOI] [PubMed] [Google Scholar]
- Munson L, Chassy LM, Asa C. Efficacy, safety and reversibility of bisdiamine as a male contraceptive in cats. Theriogenology. 2004;62:81–92. doi: 10.1016/j.theriogenology.2003.07.026. [DOI] [PubMed] [Google Scholar]
- Napoli J. Retinoic acid: its biosynthesis and metabolism. In: Moldave K, editor. Progress in Nucleic Acid Research and Molecular Biology. San Diego, CA: Academic Press; 2000. pp. 139–188. [DOI] [PubMed] [Google Scholar]
- Nau H. Teratogenicity of isotretinoin revisited: species variation and the role of all-trans-retinoic acid. J Am Acad Dermatol. 2001;45:S183–S187. doi: 10.1067/mjd.2001.113720. [DOI] [PubMed] [Google Scholar]
- Naughton CK, Nelson DR, Thomas AJ. Development of an inexpensive artificial vagina for semen collection from rabbits. J Androl. 2003;24:712–715. doi: 10.1002/j.1939-4640.2003.tb02731.x. [DOI] [PubMed] [Google Scholar]
- Page ST, Kalhorn TF, Bremner WJ, Anawalt BD, Matsumoto AM, Amory JK. Intratesticular androgens and spermatogenesis during severe gonadotropin suppression induced by male hormonal contraceptive treatment. J Androl. 2007;28:734–741. doi: 10.2164/jandrol.107.002790. [DOI] [PubMed] [Google Scholar]
- Paik J, Blaner WS, Swisshelm K. Cis-retinol dehydrogenase: 9-cisretinol metabolism and its effect on proliferation of human MCF7 breast cancer cells. Exp Cell Res. 2005;303:183–196. doi: 10.1016/j.yexcr.2004.09.019. [DOI] [PubMed] [Google Scholar]
- Paik J, Vogel S, Quadro L, Piantedosi R, Gottesman M, Lai K, Hamberger L, Vieria MM, Blaner WS. Vitamin A: overlapping delivery pathways to tissues from the circulation. J Nutr. 2004;134:S276–S280. doi: 10.1093/jn/134.1.276S. [DOI] [PubMed] [Google Scholar]
- Pellegrini M, Filippani D, Gori M, Barrios F, Lolicato F, Grimaldi P, Rossi P, Jannini EA, Geremia R, Dolci S. ATRA and KL promote differentiation toward the meiotic program of male germ cells. Cell Cycle. 2008;7:3878–3888. doi: 10.4161/cc.7.24.7262. [DOI] [PubMed] [Google Scholar]
- Schulze GE, Clay RJ, Mezza LE, Bregman CL, Buroker RA, Frantz JD. BMS-189453, a novel retinoid receptor antagonist, is a potent testicular toxin. Toxicol Sci. 2001;59:297–308. doi: 10.1093/toxsci/59.2.297. [DOI] [PubMed] [Google Scholar]
- Seed J, Chapin RE, Clegg ED, Dostal LA, Foote RH, Hurtt ME, Klinefelter GR, Makris SL, Perreault SD, Schrader S, Seyler D, Sprando R, Treinen KA, Veeramachaneni DN, Wise LD. Methods for assessing sperm motility, morphology, and counts in the rat, rabbit and dog: A consensus report. Reprod Toxicol. 1996;10:237–244. doi: 10.1016/0890-6238(96)00028-7. [DOI] [PubMed] [Google Scholar]
- Singh SK, Dominic CJ. Effect of N′N′-bis(dichloroacetyl)-1,8-octamethylenediamine (WIN 18,446) on the testis and epididymis of the musk shrew (Suncus murinus) Indian J Exp Biol. 1980;18:1217–1220. [PubMed] [Google Scholar]
- Steward MJ, Malek K, Crabb DW. Distribution of messenger RNAs for aldehyde dehydrogenase 1, aldehyde dehydrogenase 2, and aldehyde dehydrogenase 5 in human tissues. J Investig Med. 1996;44:42–46. [PubMed] [Google Scholar]
- Suh JJ, Pettinati HM, Kampman KM, O'Brien CP. The status of disulfiram: a half a century later. J Clin Psychopharmacol. 2006;26:290–302. doi: 10.1097/01.jcp.0000222512.25649.08. [DOI] [PubMed] [Google Scholar]
- Tasaka H, Takenaka H, Okamoto N, Onitsuka T, Koga Y, Hamada M. Abnormal development of cardiovascular systems in rat embryos treated with bisdiamine. Teratology. 1991;43:191–200. doi: 10.1002/tera.1420430303. [DOI] [PubMed] [Google Scholar]
- Vernet N, Dennefeld C, Guillou F, Chambon P, Ghyselinck NB, Mark M. Prepuberal testis development relies on retinoic acid but not rexinoid receptors in Sertoli cells. EMBO J. 2006a;25:5816–5825. doi: 10.1038/sj.emboj.7601447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vernet N, Dennefeld C, Rochett-Egly C, Oulad-Abdelghani M, Chambon P, Ghyselinck NB, Mark M. Retinoic acid metabolism and signaling pathways in the adult and developing mouse testis. Endocrinology. 2006b;147:96–110. doi: 10.1210/en.2005-0953. [DOI] [PubMed] [Google Scholar]
- Wang X, Penzes P, Napoli J. Cloning of a cDNA encoding an aldehyde dehydrogenase and its expression in Escherichia coli. J Biol Chem. 1996;27:16288–16293. doi: 10.1074/jbc.271.27.16288. [DOI] [PubMed] [Google Scholar]
- Wolbach SB, Howe PR. Tissue changes following deprivation of fat soluble A vitamin. J Exp Med. 1925;42:753–777. doi: 10.1084/jem.42.6.753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu FC, Farley TM, Peregoudov A, Waites GM. Effects of testosterone enanthate in normal men: experience from a multicenter contraceptive efficacy study. World Health Organization Task Force on Methods for the Regulation of Male Fertility. Fertil Steril. 1996;65:626–636. [PubMed] [Google Scholar]
- Zaiss AK, Son S, Chang LJ. RNA 3′ readthrough of oncoretrovirus and lentivirus: implications for vector safety and efficacy. J Virol. 2002;76:7209–7219. doi: 10.1128/JVI.76.14.7209-7219.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Q, Li Y, Nie R, Friel P, Metchell D, Evanoff RM, Pouchnik D, Banasik B, McCarrey JR, Small C, Griswold MD. Expression of stimulated by retinoic acid gene 8 (Stra8) and maturation of murine gonocytes and spermatogonia induced by retinoic acid in vitro. Biol Reprod. 2008;78:537–545. doi: 10.1095/biolreprod.107.064337. [DOI] [PMC free article] [PubMed] [Google Scholar]