

Abstract
Hepatic fibrosis (HF) is a progressive condition with serious clinical complications arising from abnormal proliferation and amassing of tough fibrous scar tissue. This defiance of collagen fibers becomes fatal due to ultimate failure of liver functions. Participation of various cell types, interlinked cellular events, and large number of mediator molecules make the fibrotic process enormously complex and dynamic. However, with better appreciation of underlying cellular and molecular mechanisms of fibrosis, the assumption that HF cannot be cured is gradually changing. Recent findings have underlined the therapeutic potential of a number of synthetic compounds as well as plant derivatives for cessation or even the reversal of the processes that transforms the liver into fibrotic tissue. It is expected that future inputs will provide a conceptual framework to develop more specific strategies that would facilitate the assessment of risk factors, shortlist early diagnosis biomarkers, and eventually guide development of effective therapeutic alternatives.
Keywords: Antifibrotic strategies, biochemical abnormalities, cell types, gene therapy, hepatic fibrosis
Hepatic fibrosis (HF) is a pathological condition resulting in abnormal proliferation and accumulation of tough fibrous connective tissue (scar tissue) in the liver. Although the formation of scar tissue is a normal body response to injury, in fibrosis, this healing process goes erroneous. Normal process of wound healing involves collagen deposition; however, the chronic activation of this healing mechanism leads to liver pathology. Among a variety of causes/factors or stimuli, which bring about this transformation are: chronic infection by hepatitis B, C viruses and parasite Schistosoma, chronic alcoholism and/or exposure to certain drugs and toxins, infections, non-alcoholic steatohepatitis (NASH), inherited metabolic diseases like hematochromatosis, Wilson's disease, α-1 antitrypsin deficiency, autoimmune diseases such as primary biliary cirrhosis, and auto-immune hepatitis.[1] Generally, HF begins with the stimulation of inflammatory immune cells to secrete cytokines, growth factors, and other activator molecules. These chemical messengers direct hepatic stellate cells (HSCs) to activate and synthesize collagen, glycoproteins (such as fibronectin), and proteoglycans. Deposition of the abnormal products of stellate cells, along with portal myofibroblasts, bone marrow-derived cells, and epithelial mesenchymal tissues (EMT), build-up of extracellular matrix (ECM, nonfunctional connective tissue) in the liver, and the impairment by collagenolysis are simultaneous processes.[2–5] In the long run, it may either lead to cirrhosis and related complications or may couple with carcinogenesis and ultimate death due to failure of normal liver functions.[6,7] Nearly 3 billion and 180 million people have been exposed to hepatitis B and C virus, respectively.[8,9] Epidemiological studies have shown that in China, hepatitis B viral (HBV) infection is the major cause of liver fibrosis, whereas in the United States, Europe, and Japan, hepatitis C viral (HCV) infection and alcohol are the main causes.[10,11] In the sub-Saharan African region, Schistosoma mansoni infection is reported to be the major cause of HF, resulting in almost 0.3 million deaths annually.[12] This review covers recent information on the general mechanisms of HF, its impact on some biochemical parameters, and therapeutic potential of certain antifibrotic agents. The discussion outlines possible strategies and applications of the published information while designing and formulating new treatment regimen.
FUNCTIONAL CONSEQUENCES OF DIFFERENT CELL TYPES
HSCs
HSCs, also known as fat-storing cells or perisinusoidal lipocytes, represent almost 5–8% of all healthy liver cells. They are located near the hepatocyte laminated in the perisinusoidal space of Disse by means of star-like dendritic cytoplasmic processes extending along and around the hepatic endothelial cells and hypothesized of being mesenchymal in origin.[13] HSCs show two different phenotypes: quiescent in the normal liver and activated in the diseased. As a consequence of this transformation, diseased phenotype has altered functions. In the following, we summarize morphological and functional differentiation of the two phenotypes of HSCs.
Quiescent HSCs
Quiescent HSCs have a star-like shape and their cytoplasm encloses numerous lipid droplets, which contain retinoids, triglycerides, cholesterol, and free fatty acids.[14] Storage and limited release of retinoids is a major function of HSC in the healthy liver. An essential and prominent structural feature of HSCs is the presence of microfilament bundles of actin and intermediate filaments such as desmin, vimentin, and synemin.[15] HSCs also express the LIM-homeodomain protein Lhx2, a transcription factor responsible for maintaining their quiescence.[16] Quiescent HSCs express peroxisome proliferator-activated receptor-γ (PPAR-γ), a nuclear receptor considered as fundamental transcriptional regulator for adipogenesis that also displays antifibrogenic effects by inhibiting type I collagen expression at the transcriptional level.[17] Quiescent HSCs also regulate the expression of hepatocyte growth factor (HGF), TGF-β, insulin-like growth factor-I (IGF-I), and other cytokines in an auto- and paracrine manner [Figure 1].[18–20] However, the phenotype lacks expression of fatty acid synthase (FAS) receptor CD95 (a cellular surface protein with a molecular weight of 42-52 kDa that promotes apoptosis).[20] Reportedly, HSCs have a role in the expression of some other important neural proteins like glial fibrillary acidic proteins, neuronal growth factor, synaptophysin, RhoN, glutamine synthetase, and neurotrophin receptors.[21]
Figure 1.
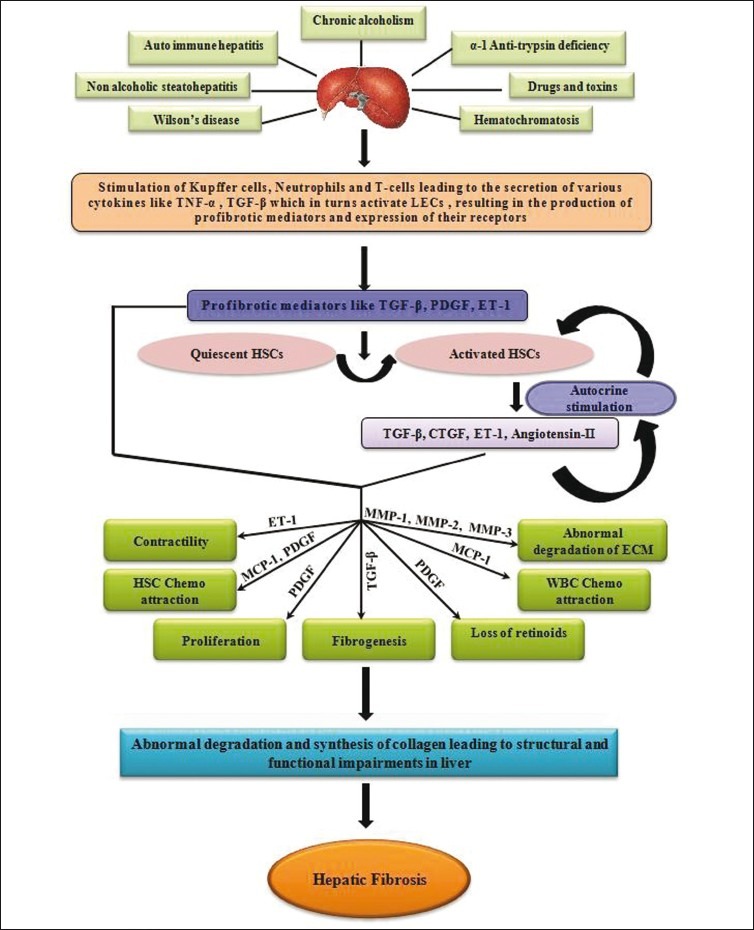
Early causes and mechanisms of matrix degradation and progression of hepatic fibrosis. Stimulation of Kupffer cells, neutrophils, and T-cells cause secretion of various cytokines and profibrotic mediator to convert quiescent to activated hepatic stellate cells (HSCs). HSCs are associated with matrix degradation due to increased production of membrane type matrix metalloproteinase-1 (MMP-1), matrix metalloproteinase-2, -3 (MMP-2, -3), and tissue inhibitors of metalloproteinases (TIMPs), leading to amassing of scar tissue
Activated HSCs
Activation of hepatic stellate cells depends on a number of factors discussed under “Introduction,” which are either directly or indirectly involved in progression of the HF.[1] Activated HSCs develop into myofibroblast-like cell types, which are differentiated by the loss of lipid droplets, lack of glial fibrillary acidic proteins, and increased cell proliferation. Consequently, excessive synthesis of ECM components occurs, causing increased expression of α-smooth muscle actin (α-SMA) and changes in the expression of L-type voltage-operated Ca2+ channels, which are known to mediate Ca2+ influx and regulate cellular contraction.[22] The activation of HSCs is controlled by the gene expression, which is itself regulated by various transcription factors briefly described below.
FoxO
Forkhead box gene group O. Its functions and its intracellular localization are regulated by growth factor (mainly PDGF) activated kinases, especially phosphoinositide 3-kinase and protein kinase-B (PKB), through phosphorylation.[23,24] Phosphorylation suppresses transactivation and promotes the translocation of FoxO proteins from the nucleus to the cytosol, reducing the expression of their target genes. The transcription factor FoxO is a key player in controlling the trans-differentiation and proliferation of HSCs that leads to liver fibrosis in vivo.[25]
ILK
Integrin-linked kinase, plays a crucial role in HSC activation, fibrogenesis, and transducing signals from the ECM or from two known growth factors, TGF-β and ET-1, to the cytoplasm.[26] ILK couples the integrins and growth factors to downstream signaling pathways, which favor the suppression of apoptosis and promote cell cycle progression. The increased expression of ILK is unregulated during HF.[26]
PPAR-γ
Peroxisome proliferator-activated receptor-γ, a nuclear receptor transcriptional factor considered to be the fundamental transcriptional regulator for adipogenesis (anabolic pathways) and also reported to have antifibrogenic activity, is expressed by quiescent HSCs. Reduced activity of PPAR-γ, results in increased HSC activation and proliferation.[17]
Kupffer Cells in HF
Kupffer cells are highly phagocytic tissue macrophages of the liver, responsible for the removal of circulating microorganisms, immune complexes, and debris from the blood stream as also detoxicating endotoxins. They constitute about 15% of the total liver cell population. Moreover, these Kupffer cells take up different substances from the circulation via receptor-mediated endocytosis. Being a part of the innate immune system, these cells play an important role in the regulation of inflammatory processes in liver by secreting cytokines such as TNF-α, IL-1, IL-6, and reactive oxygen species (ROS), which promote chemotaxis, phagocytosis, and ROS production by other inflammatory cells.[27]
Kupffer cells, when treated with gadolinium chloride, produce interstitial collagenase MMP-13, which reduces ECM deposition during experimental fibrosis.[28] In addition, activated Kupffer cells can effectively kill HSC by a caspase 9-dependent mechanism via the involvement of TNF-related apoptosis-inducing ligand (TRAIL).[29,30] Their antifibrotic effect is evident by the presence of IL-10, another important cytokine that has anti-inflammatory and antifibrotic effects, especially in the early stages of fibrosis and during acute liver injury.[31] While decreasing collagen production, IL-10 up-regulates collagenase secretion, resulting in a reduction of collagen deposition.
PROFIBROTIC MEDIATORS
The activated HSCs produce a number of cytokines and active peptides that promote their constrictive, proliferative, and transformative properties in an autocrine manner, promoting the development of liver fibrosis.[32–35] Among fibrotic mediators, TGF-β is certainly one of the most important polypeptides along with PDGF, which is regarded as a potent stellate cell mitogen.[36] Activated HSCs are known to up-regulate and enhance the autocrine effect of receptors for IL-10, FAS (CD95), PDGF, FGF, VEGF, TGF-β1, and p75 (a member of the TNF receptor super family).[20,32,35,37,38] At the same time, these HSCs also down-regulate the expression of IGF-I receptor, three TGF-β type receptors, and signaling mediators commonly named “Smad proteins.”[20,39]
In HSCs, TGF-β participate in intracellular signaling cascade and transcriptional regulation of the genes: Ras, Raf-1, MEK, and MAPK.[40] Though the main source of TGF-β in the fibrotic liver is activated HSCs, liver endothelial cells and Kupffer cells also contribute to synthesis of this growth factor.[41]
Cholestasis (decreased excretion of bile) may result in accumulation of bile acids in liver, which encourages biliary epithelial cells to secrete ET-1, TNF-α, and PDGF. A 21-amino acid peptide, ET-1, induces HSC activation, invasiveness, and fibrogenesis by up-regulating type-I collagen gene expression and procollagen α-1 during the early phase.[42] The up-regulation of ET-1 is catalyzed by endothelin-converting enzyme-1 that continuously converts precursor ET-1 into mature ET-1 under favorable conditions.[42]
MOLECULAR BASIS OF HF
HF involves the activation of HSCs and over-expression and over-secretion of collagens, resulting in the excessive accumulation of ECM proteins. Broadly, development of the fibrosis has two main phases: inflammation and fibrogenesis. Initially, various hepatotoxic factors induce synthesis of mediators, which cause inflammatory reactions within hepatic cells. Mediators of inflammation phase bring about the strenuous phenotypic change from quiescent HSCs to activated HSCs, which have altered ECM composition. In the course of activation process, transformed HSCs become proliferative and acquire characteristic features. Morphologically, they appear myofibroblast-like, which are devoid of lipid droplets, lack glial fibrillary-acidic protein, have excess production of ECM components, and exhibit increased expression of α-smooth muscle actin (α-SMA) fibers. Stimulated by TGF-β released initially from Kupffer cells, activated HSCs also start to synthesize markedly increased amounts of ECM proteins; specifically type-I and type-III collagen.
Activated HSCs show chemotactic response and migrate towards regions of injury and start accumulating around damaged tissue. The same cytokines, which are mitogens for HSCs, play the role of chemo-attractants for these cells. Convincing evidence supports the secretion of chemo-attractants (for instance, monocyte chemotactic protein, MCP-1), which not only activate HSCs but also attract other activated HSCs. The cytokines promote the recruitment of monocytes and leukocytes.[43] Activated HSCs express the cytoskeleton protein, α-smooth muscle actin (α-SMA), equipping the cells with a contractile apparatus and various connective tissue proteins including collagen types I, III, and IV.[38,44,45] HSCs are thus capable of constricting individual sinusoids as well as the entire fibrotic liver. A balance between ET-1 and NO is assumed to regulate the contractile activities of HSC, wherein ET-1 is the key contractile stimulus of HSC while NO an antagonist of ET-1 produced by HSC, Kupffer cells, and liver endothelial cells.[42]
During fibrosis, the low-density matrix that is normal for healthy liver is degraded and replaced by an excess of non-functional collagenous ECM tissue. Calcium-dependent enzymes and matrix metalloproteinases (MMP) disrupt both collagen and non-collagenous compounds of ECM.[46] Activated HSCs also secrete MMP-1 (interstitial collagenase), MMP-2 (Gelatinase A), and MMP-3 (Stromelysin 1), which degrades the main components of ECM such as type IV collagen and laminin. In a way, the lytic activities indirectly perpetuate the deposition of collagen type I and III. Moreover, in fibrotic liver, markedly increased levels of tissue inhibitors of matrix metalloproteinases (TIMPs) have been recorded. They are capable of inhibiting the action of MMP-1, thereby causing the accumulation of collagen fibers [Figure 1].[47,48] In the sub-Saharan African region, Schistosoma mansoni infection is the major cause of HF, affecting more than 200 million people of tropical countries, and is accepted to be highly endemic, specifically in agricultural regions of Egypt and Sudan.[49] Fibrosis caused by schistosomes is characteristically periportal type and can be subdivided into four stages: fibroblasts recruitment, followed by HSC differentiation, their proliferation, and subsequent secretion and remodeling of ECM.[50,51] In schistosomiasis, there is granuloma formation followed by inflammation that in subsequent steps causes portal hypertension, collagen deposition, and gastrointestinal bleeding mediated by TH2 cytokines (i.e., IL-4, IL-5, IL-10) and TGF-β.[52,53] Literature shows that high levels of IL-5 and IL-13 are found in subjects with parasite-induced liver fibrosis.[54] Contrary to a general mechanism, some interesting work published in the recent past also showed inhibitory roles of TNF-α, IL-12, INF-γ, and NO, associated with the TH1 response in schistosomiasis.[53]
BIOCHEMICAL ABNORMALITIES AND HISTOLOGICAL CHANGES OBSERVED DURING PROGRESSION OF HF
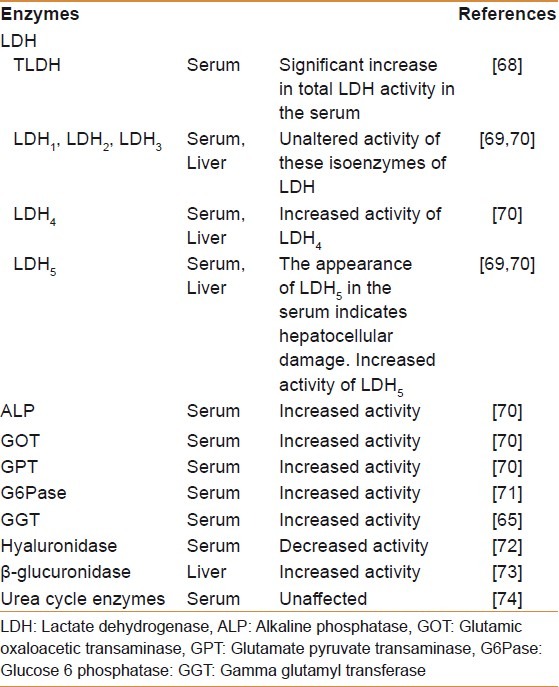
Hepatic toxicity is always accompanied by impaired hepatocyte metabolism and deposition of connective tissue components in the liver.[55,56] Update of literature reveals that biochemical abnormalities are mainly associated with disruption in the levels of intermediate metabolites or their end products in experimentally induced as well as naturally occurring HF. Moreover, enzymes like GOT, ALP, LDH, and many others have been reported to vary during the progression of HF. The summary of these abnormalities in either of the above conditions is listed in Tables 1–3.[57–79]
Table 1.
Changes in the levels of amino acid and proteins noted during the progression of hepatic fibrosis

Table 3.
Status of various biologically important molecules during liver fibrosis

Table 2.
Adjustments in the levels of different enzymes from sera and liver tissue during progression of hepatic fibrosis
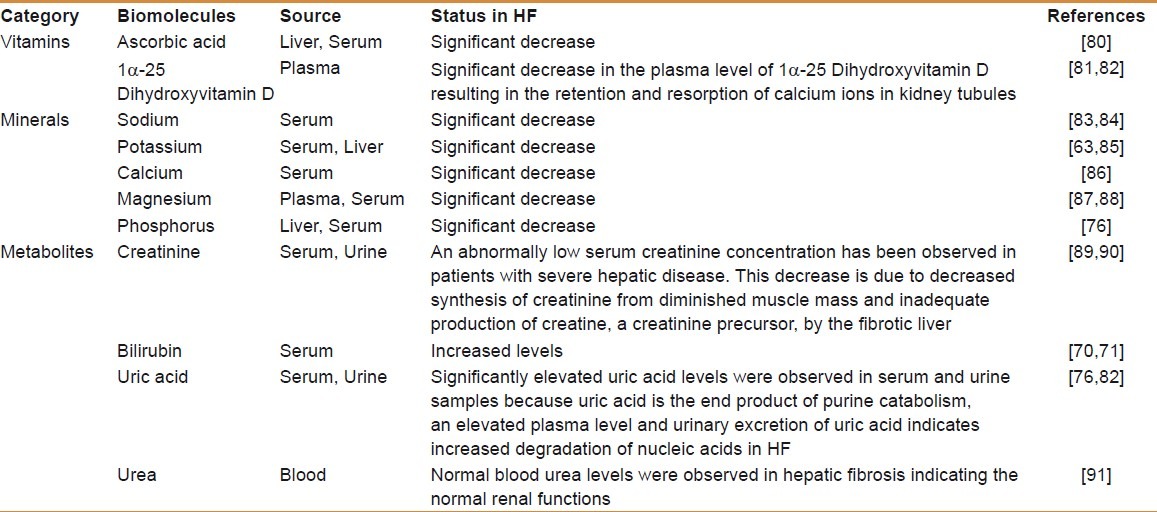
Like vitamins, minerals and metabolites also play important roles in the living systems. The minerals are components of metalloproteins and metalloenzymes, apart from acting as enzyme cofactors. As these compounds are metabolized mainly in the liver, functional impairment of hepatic tissue in fibrosis and cirrhosis alters their levels as well. A compilation of levels of important vitamins, minerals, and metabolites altered during HF is given in Table 4.[80–91]
Table 4.
Variations in the levels of vitamins, minerals, and other indispensable metabolites in the fibrotic liver
Substantial literature is available on the histological changes occurring during HF.[58,70] These reports invariably demonstrate significant decrease in “liver weight” and decreased “liver to body weight ratio” in N’-Nitrosodimethyl amine (NDMA)-induced HF.[59,70] Decreased protein synthesis, cell necrosis, and collapse of liver parenchyma in late stages of HF are among suggested reasons. As a reference, in this section of review, we have taken up the changes, which occur in the liver only due to NDMA administration in mammalian model [Figure 2]. Immuno-histochemical (IHC) staining using monoclonal antibodies showed the presence of α-SMA in NDMA-treated liver sections [Figure 2b]. The presence of α-SMA convincingly showed that it was synthesized by activated HSCs during the course of HF. Normal liver specimens do not stain for α-SMA [Figure 2a]. IHC staining clearly demonstrated much higher number of positive stained cells in the fibrotic area when compared with normal areas of the liver sections.[70] In a recent study, peculiar changes in the RBC rheology along with the changes in other blood-related parameters and enzymes in a mammalian fibrotic model have also been demonstrated.[79]
Figure 2.
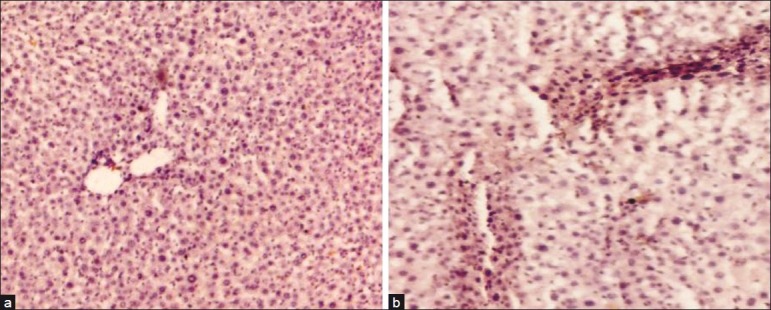
Immunohistochemical staining of α-smooth muscle actin (α-SMA) showing activated hepatic stellate cells during the progression of N’- Nitrosodimethyl amine (NDMA)-induced hepatic fibrosis in rats. (a) Control liver (×100) demonstrates absence of α-SMA staining and, (b) NDMA treatment day 21 (×100) exhibiting significant staining of α-SMA with enormous number of activated stellate cells in fibrotic zone
ANTIFIBROTIC STRATEGIES
Recent research in molecular biology has helped to discover and better characterize several elements that are perfectly and dynamically involved in making the liver fibrotic. This deeper molecular understanding of the pathogenesis of liver fibrosis has opened up opportunities for novel and exciting complementary therapeutic approaches. A remarkable achievement has been the demonstration of reversibility of advanced fibrosis and even cirrhosis after administering certain anti-fibrotic agents.[70,71,79]
Antifibrotic compounds are classified in accordance with their mechanism of action [Table 5].[92–110] For instance, compounds such as antioxidants and those with properties of reducing inflammation, promoting ECM degradation, inhibiting HSC activation and proliferation, reducing ECM production by HSC, neutralizing HSC contractile response, and stimulation of HSC apoptosis.
Table 5.
Existing antifibrotic strategies, target molecules, and their mechanism of action in obstructing or regressing hepatic fibrosis
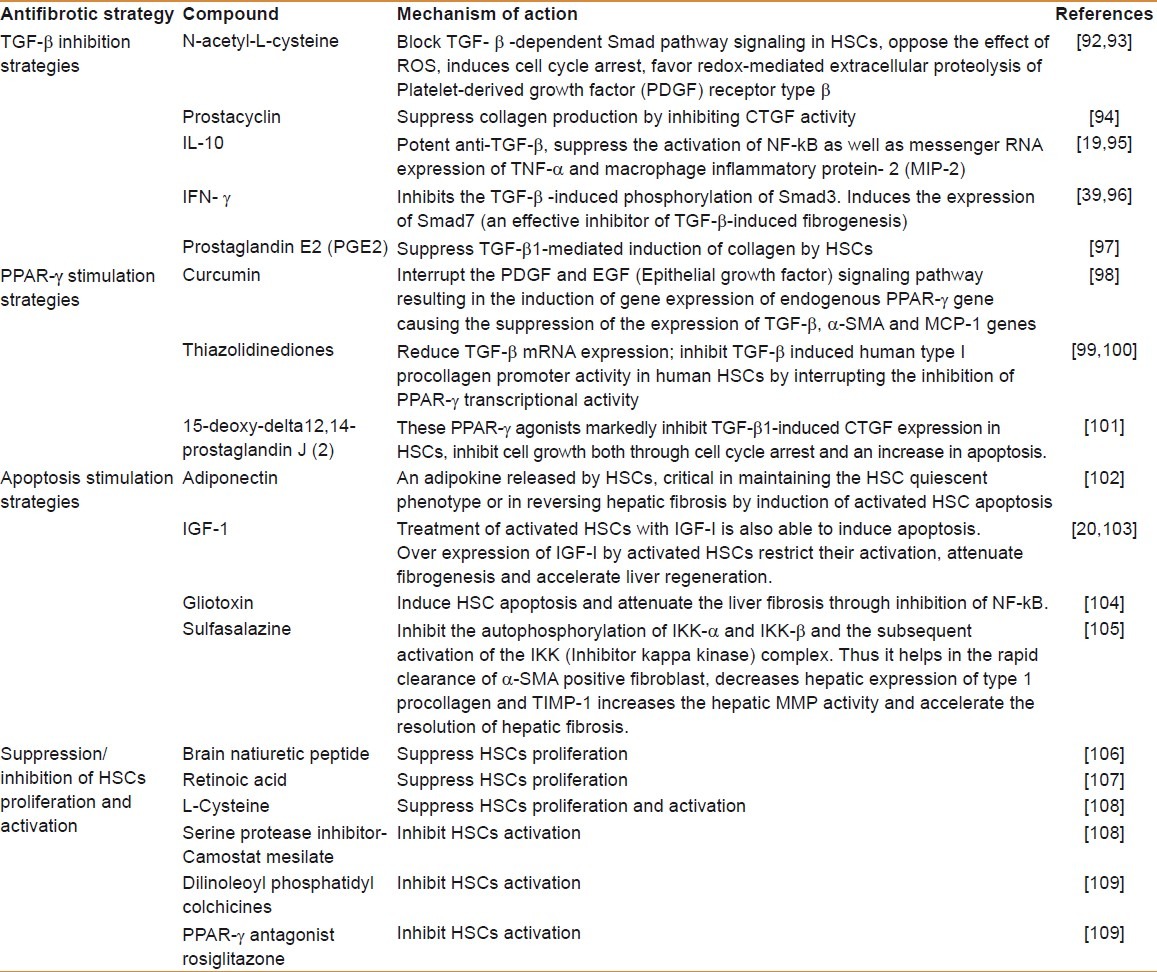
Participants like ECM proteins, some receptors mediating cell–ECM interactions, cytokines, integrins, growth factors (principally TGF-β and PDGF) and post-receptor signal regulators (i.e., Smads), and transcription factors (principally a decreased expression of PPAR-γ) can be the focus of an antifibrogenic therapeutic strategy.[39,98,99,101,109] Even though a large number of drugs have also been developed for antifibrotic effects, they have their own limitations and side effects. Interestingly, some recent studies have highlighted remarkable therapeutic effects of plant derivatives and the whole phytoextracts in reversing HF [Table 5].
Besides, few other compounds have been demonstrated to possess antifibrotic properties, as given below briefly.
Relaxin
A peptide hormone, has been shown to be antifibrotic that works by decreasing TIMP-1 and TIMP-2 expression in HSCs, which enhances matrix degradation and reduces interstitial collagen deposition.[111,112] When added to cultured TGF-β1-stimulated rat HSCs, relaxin displays anti-TGF-β effects by inhibiting collagen over-expression in vitro and reducing the level of α-SMA.[111]
Imatinib
A drug used for the treatment of chronic myelogenous leukemia and gastrointestinal stromal tumors, inhibits PDGF-β receptor kinase.[113] This is a potentially useful property for attenuating liver fibrosis. Despite proven antifibrotic effects in cultured HSCs as well as in animal models of liver fibrosis, this drug has a limitation due to its cardiotoxic side effects.[114]
Angiotensin II receptor antagonist
Circulating levels of angiotensin II (ANGII), a powerful vasoconstrictor factor, frequently increase in chronic liver diseases. In these conditions, HSCs proliferate, acquire contractile properties, and excessively synthesize endothelin, PDGF, and chemokines.[33,115] In vitro and in vivo studies on HF suggest that the angiotensin II type 1 receptor antagonist suppresses proliferation, collagen synthesis, and expression of profibrogenic cytokines (TGF-β1 and CTGF) in activated HSCs.[115–117] Two mechanisms of action have been suggested: first, the angiotensin II type 1 receptor antagonist inhibits activated HSCs by blocking angiotensin II type 1 receptors expressed on the surface of HSCs; second, it suppresses the activation of HSCs as a result of the decrease in TGF-β1. Studies on the use of botanicals or their active compounds for treating HF are summarized in Table 6.[70,71,118–124]
Table 6.
Known phytoextracts and their derivatives with probable therapeutic action used in traditional medicine to treat hepatic fibrosis

Liver fibrosis caused by S. mansoni is of great concern in tropical countries of the world, and owing to repeated anti-schistomic chemotherapy, there are reports of emergence of drug-resistant strains, which have lead researchers to search for suitable alternative medicines. Like in non-parasitic HF, botanicals or phytoextracts are given proper attention in treatment against trematode-induced liver fibrosis due to their friendly interaction and safer action. Many botanicals like Daucus carota,[125] Commiphora molmol,[126] Artemisia anmia,[127] Combretum sp.,[128] crude oil of Nigella sativa,[129–132] Zingiber officinale,[133] Solanum nigrum,[134] Allium sativum,[135,136] Curcuma longa,[137] and Camellia sinesis[138] have been reported to possess anti-schistosomal properties. Moreover, among synthetic drugs, high levels of Somatostatin and Paeoniflorin have been shown to reduce fibrosis by inhibiting HSC activation in humans and collagen synthesis in IL-13-stimulated HSC in murine S. japonicum infection, respectively.[139,140] Moreover, thiazolidinedione drug, rosiglitazone, and glucocortocoid dexamethasone, have been observed to attenuate HF by activating the PPAR-γ ligand in murine schistosomiasis.[141,142]
GENE THERAPY APPROACH FOR HF
HF involves the expression and suppression of a number of genes. Many attempts have been made to induce the cessation or the reversal of the process, but all of them did not produce desirable results, either due to the low uptake by the target cells or major side effects on other cell populations.[143] A recent report demonstrated the reversal of HF and cirrhosis in rodent model by targeting HSCs via liposomal drug delivery. The targeting complex contained siRNA against the collagen chaperon heat shock protein 47 (HSP47) bound to vitamin A.[144] These antifibrotic vitamin A–coupled liposomes decrease collagen production and promote degradation of ECM. However, thriving steps are being taken to attenuate experimental liver fibrosis with gene therapy. A brief review of novel strategies relying on gene therapy is given in Table 7.[145–155]
Table 7.
Modern approaches of gene therapy used in the treatment of liver fibrosis

Out of many, stem-cell transplantation still seems to be a more promising alternative approach.[156] It is also believed that cell types including neurons, cardiac muscle cells, skeletal cells, kidney cells, liver cells, etc., have similar origin and arise from bone marrow (BM) stem cells. BM-derived stem cells have great power of regeneration and may develop into specific cellular phenotypes with different functions. These stem cells play an active role in liver repair and hepatic regeneration and may also support regeneration of cardiac/skeletal muscle and brain tissue. Recent studies suggested that hematopoietic stem cells migrate from the bone marrow to the injured liver due to the hypoxic milieu generated at the injured hepatic site and formation of chemo-attractant gradients.[157] Using markers such as aldehyde dehydrogenase, it has been demonstrated that these migrated stem cells fuse with the host hepatocytes or liver cells and help to generate fresh cell types.[158]
CONCLUSIONS
HF is a severe consequence of needless accumulation of excessive connective tissue in the liver. This amassing occurs because either ECM components are overproduced (fibrogenesis) or poorly degraded, or both contribute to it. It would be of great interest if the fibrosis pathology research proceeds to gain insights into proteomics to understand the normal and activated HSCs’ function. Only then would a comprehensive understanding of the underlying molecular mechanisms be possible. New propositions can then be put forward for the viewpoint of impairments within disease processes. The transformation from basic research to practical investigations is expected to provide novel measures for prospective therapeutics against liver diseases. Since human liver fibrosis can be caused by various pathologies (viral, alcoholic, metabolic, etc.) and each one of these might respond better to one or another drug, it is advisable to reproduce animal models to evaluate effectiveness and therapeutic potential of relatively safer and targeted drugs. Trials for safer or well-tolerated drugs with use in multiple diseases, even for schistosome-induced fibrosis,[159] are underway and will carry great importance since the real efficacy of drugs that are likely to block activation or transformation of HSCs already tested in vitro can be directly assessed. So-called modern therapeutic procedures like gene therapy, BM-derived stem cells, and approaches utilizing siRNA along with hepatocyte transplant to reconstitute normal liver function may be expected as future areas to explore possibilities of treatment against HF.
Footnotes
Source of Support: University Grants Commission (UGC) [DS Kothari-PDF], Department of Science & Technology (DST), Ministry of Science & Technology, New Delhi [No. DST/INSPIRE/2010/121]
Conflict of Interest: None declared
REFERENCES
- 1.Friedman SL. Mechanism of hepatic fibrogenesis. Gastroenterology. 2008;134:1655–69. doi: 10.1053/j.gastro.2008.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wells RG. The role of matrix stiffness in regulating cell behavior. Hepatology. 2008;47:1394–400. doi: 10.1002/hep.22193. [DOI] [PubMed] [Google Scholar]
- 3.Kalluri R, Neilson EG. Epithelial-mesenchymal transition and its implications for fibrosis. J Clin Invest. 2003;112:1776–84. doi: 10.1172/JCI20530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wells RG, Kruglov E, Dranoff JA. Autocrine release of TGF-beta by portal fibroblasts regulates cell growth. FEBS Lett. 2004;559:107–10. doi: 10.1016/S0014-5793(04)00037-7. [DOI] [PubMed] [Google Scholar]
- 5.Forbes SJ, Russo FP, Rey V, Burra P, Rugge M, Wright NA, et al. A significant proportion of myofibroblasts are of bone marrow origin in human liver fibrosis. Gastroenterology. 2004;126:955–63. doi: 10.1053/j.gastro.2004.02.025. [DOI] [PubMed] [Google Scholar]
- 6.Heidelbaugh JJ, Bruderly M. Cirrhosis and chronic liver failure: Part I.Diagnosis and evaluation. Am Fam Physician. 2006;74:756–62. [PubMed] [Google Scholar]
- 7.Perz JF, Armstrong GL, Farrington LA, Hutin YJ, Bell BP. The contributions of hepatitis B virus and hepatitis C virus infections to cirrhosis and primary liver cancer worldwide. J Hepatol. 2006;45:529–38. doi: 10.1016/j.jhep.2006.05.013. [DOI] [PubMed] [Google Scholar]
- 8.Lavanchy D. Worldwide epidemiology of HBV infection, disease burden, and vaccine prevention. J Clin Virol. 2005;34(Suppl 1):S1–3. doi: 10.1016/s1386-6532(05)00384-7. [DOI] [PubMed] [Google Scholar]
- 9.Simmonds P, Bukh J, Combet C, Deleage G, Enomoto N, Feinstone S, et al. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology. 2005;42:962–73. doi: 10.1002/hep.20819. [DOI] [PubMed] [Google Scholar]
- 10.Seeff LB, Hoofnagle JH. Appendix: The national institutes of health consensus development conference management of hepatitis C 2002. Clin Liver Dis. 2003;7:261–87. doi: 10.1016/s1089-3261(02)00078-8. [DOI] [PubMed] [Google Scholar]
- 11.Koike K. Antiviral treatment of hepatitis C: present status and future prospects. J Infect Chemother. 2006;12:227–32. doi: 10.1007/s10156-006-0460-0. [DOI] [PubMed] [Google Scholar]
- 12.van der Werf MJ, de Vlas SJ, Brooker S, Looman CW, Nagelkerke NJ, Habbema JD, et al. Quantification of clinical morbidity associated with schistosome infection in sub-Saharan Africa. Acta Trop. 2003;86:125–39. doi: 10.1016/s0001-706x(03)00029-9. [DOI] [PubMed] [Google Scholar]
- 13.Geerts A. On the origin of stellate cells: mesodermal, endodermal or neuroectodermal? J Hepatol. 2004;40:331–4. doi: 10.1016/j.jhep.2003.12.006. [DOI] [PubMed] [Google Scholar]
- 14.Vogel S, Gamble MV, Blaner WS. Biosynthesis, absorption, metabolism and transport of retinoids. In: Nau H, Blaner WS, editors. Retinoids: The biochemical and molecular basis of vitamin A and retinoid action. Berlin: Springer; 1999. pp. 31–95. [Google Scholar]
- 15.Uyama N, Zhoa L, van Rossen E, Hirrako Y, Reynaert H, Adams DH, et al. Hepatic stellate cells express synemin, a protein bridging intermediate filaments to focal adhesions. Gut. 2006;55:1276–89. doi: 10.1136/gut.2005.078865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kolterud A, Wandzioch E, Carlsson L. Lhx2 is expressed in the septum transversum mesenchyme that becomes an integral part of the liver and the formation of these cells is independent of function of Lhx2. Gene Expr Patterns. 2004;4:521–8. doi: 10.1016/j.modgep.2004.03.001. [DOI] [PubMed] [Google Scholar]
- 17.Yang L, Chan CC, Kwon OS, Liu S, McGhee J, Stimpson SA, et al. Regulation of peroxisome proliferator-activated receptor-γ in liver fibrosis. Am J Physiol Gastrointest Liver Physiol. 2006;291:G902–11. doi: 10.1152/ajpgi.00124.2006. [DOI] [PubMed] [Google Scholar]
- 18.Matsumoto K, Nakamura T. Emerging multipotent aspects of hepatocyte growth factor. J Biochem. 1996;119:591–600. doi: 10.1093/oxfordjournals.jbchem.a021283. [DOI] [PubMed] [Google Scholar]
- 19.Gressner AM, Weiskirchen R, Breitkopf K, Dooley S. Roles of TGF-β in hepatic fibrosis. Front Biosci. 2002;7:d793–807. doi: 10.2741/A812. [DOI] [PubMed] [Google Scholar]
- 20.Saile B, DiRocco P, Dudas J, El-Armouche H, Sebb H, Eisenbach C, et al. IGF-1 induces DNA synthesis and apoptosis in rat liver hepatic stellate cells (HSC) proliferation in rat liver myofibroblasts (rMF) Lab Invest. 2004;84:1037–49. doi: 10.1038/labinvest.3700116. [DOI] [PubMed] [Google Scholar]
- 21.Haussinger D, Kubitz R, Reinehr R, Bode JG, Schliess F. Molecular aspects of medicine: from experimental to clinical Hepatology. Mol Aspects Med. 2004;25:221–360. doi: 10.1016/j.mam.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 22.Gassull X, Battaler R, Gines P, Sancho-Bru P, Nicolas JM, Gorbig MN, et al. Human myofibroblastic hepatic stellate cells express Ca2+-activated K+ channels that modulate the effects of endothelin-1 and nitric oxide. J Hepatol. 2001;35:739–48. doi: 10.1016/s0168-8278(01)00198-2. [DOI] [PubMed] [Google Scholar]
- 23.Nakae J, Barr V, Accili D. Differential regulation of gene expression by insulin and IGF-1 receptors correlates with phosphorylation of a single amino acid residue in the forkhead transcription factor FKHR. EMBO J. 2000;19:989–96. doi: 10.1093/emboj/19.5.989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van der Heide LP, Hoekman MF, Smidt MP. The ins and outs of FoxO shuttling: Mechanism of FoxO translocation and transcriptional regulation. Biochem J. 2004;380:297–309. doi: 10.1042/BJ20040167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Adachi M, Osawa Y, Uchinami H, Kitamura T, Accili D, Brenner DA. The forkhead transcription factor FoxO1 regulates proliferation and transdifferentiation of hepatic stellate cells. Gastroenterology. 2007;132:1434–46. doi: 10.1053/j.gastro.2007.01.033. [DOI] [PubMed] [Google Scholar]
- 26.Shafiei MS, Rockey DC. The role of integrin-linked kinase in liver wound healing. J Biol Chem. 2006;281:24863–72. doi: 10.1074/jbc.M513544200. [DOI] [PubMed] [Google Scholar]
- 27.Luster MI, Germolec DR, Yoshida T, Kayama F, Thompson M. Endotoxin-induced cytokine gene expression and excretion in the liver. Hepatology. 1994;19:480–8. [PubMed] [Google Scholar]
- 28.Hironaka K, Sakaida I, Matsumura Y, Kaino S, Miyamoto K, Okita K. Enhanced interstitial collagenase (matrix metalloproteinase-13) production of Kupffer cell by gadolinium chloride prevents pig serum-induced rat liver fibrosis. Biochem Biophys Res Commun. 2000;267:290–5. doi: 10.1006/bbrc.1999.1910. [DOI] [PubMed] [Google Scholar]
- 29.Fischer R, Cariers A, Reinehr R, Haussinger D. Caspase 9-dependent killing of hepatic stellate cells by activated Kupffer cells. Gastroenterology. 2002;123:845–61. doi: 10.1053/gast.2002.35384. [DOI] [PubMed] [Google Scholar]
- 30.Taimr P, Higuchi H, Kocova E, Rippe RA, Friedman S, Gores GJ. Activated stellate cells express the TRAIL receptor-2/death receptor-5 and undergo TRAIL-mediated apoptosis. Hepatology. 2003;37:87–95. doi: 10.1053/jhep.2003.50002. [DOI] [PubMed] [Google Scholar]
- 31.Knolle PA, Loser P, Protzer U, Duchmann R, Schimmit E, zum Büschenfelde KH, et al. Regulation of endotoxin-induced IL-6 production in liver sinusoidal endothelial cells and Kupffer cells by IL-10. Clin Exp Immunol. 1997;107:555–61. doi: 10.1046/j.1365-2249.1997.d01-959.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Annoni G, Weiner FR, Zern M. Increased transforming growth factor β-1 gene expression in human liver disease. J Hepatol. 1992;14:259–64. doi: 10.1016/0168-8278(92)90168-o. [DOI] [PubMed] [Google Scholar]
- 33.Bataller R, Gines P, Nicolas JM, Gorbig MN, Garcia-Ramallo E, Gasull X, et al. Angiotensin II induces contraction and proliferation of human hepatic stellate cells. Gastroenterology. 2000;118:1149–56. doi: 10.1016/s0016-5085(00)70368-4. [DOI] [PubMed] [Google Scholar]
- 34.Kinnman N, Goria O, Wendum D, Gendron MC, Rey C, Poupon R, et al. Hepatic stellate cell proliferation is an early platelet-derived growth factor mediated cellular event in rat cholestatic liver injury. Lab Invest. 2001;81:1709–16. doi: 10.1038/labinvest.3780384. [DOI] [PubMed] [Google Scholar]
- 35.Pinzani M. PDGF and signal transduction in hepatic stellate cells. Front Biosci. 2002;7:d1720–6. doi: 10.2741/A875. [DOI] [PubMed] [Google Scholar]
- 36.Borkham-Kamphorst E, van Roeyen CR, Ostendorf T, Floege J, Gressner AM, Weiskirchen R. Pro-fibrogenic potential of PDGF-D in liver fibrosis. J Hepatol. 2007;46:1064–74. doi: 10.1016/j.jhep.2007.01.029. [DOI] [PubMed] [Google Scholar]
- 37.Mathurin P, Xiong S, Kharbanda KK, Veal N, Miyahara T, Motomura K, et al. IL-10 receptor and co receptor expression in quiescent and activated hepatic stellate cells. Am J Physiol Gastrointest Liver Physiol. 2002;282:981–90. doi: 10.1152/ajpgi.00293.2001. [DOI] [PubMed] [Google Scholar]
- 38.Pinzani M, Marra F. Cytokine receptors and signaling in hepatic stellate cells. Semin Liver Dis. 2001;21:397–416. doi: 10.1055/s-2001-17554. [DOI] [PubMed] [Google Scholar]
- 39.Lui C, Gaca MDA, Swenson ES, Velluci VF, Reiss M, Wells RG. Smads 2 and 3 are differentially activated by transforming growth factor β (TGF-β) in quiescent and activated hepatic stellate cells. Constitutive nuclear localization of Smads in activated cells is TGF-β-independent. J Biol Chem. 2003;278:11721–8. doi: 10.1074/jbc.M207728200. [DOI] [PubMed] [Google Scholar]
- 40.Reimann T, Hempel U, Krautwald S, Axmann A, Scheibe R, Seidel D, et al. Transforming growth factor-β1 induces activation of Ras, Raf-1, MEK and MAPK in rat hepatic stellate cells. FEBS Lett. 1997;403:57–60. doi: 10.1016/s0014-5793(97)00024-0. [DOI] [PubMed] [Google Scholar]
- 41.De Bleser PJ, Niki T, Rogiers V, Geerts A. Transforming growth factor-beta gene expression in normal and fibrotic rat liver. J Hepatol. 1997;26:886–93. doi: 10.1016/s0168-8278(97)80257-7. [DOI] [PubMed] [Google Scholar]
- 42.Rockey DC, Chung JJ. Endothelin antagonism in experimental hepatic fibrosis. Implications for endothelin in the pathogenesis of wound healing. J Clin Invest. 1996;98:1381–8. doi: 10.1172/JCI118925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pinzani M, Gesualdo L, Sabbah GM, Abboud HE. Effects of platelet-derived growth factor and other polypeptide mitogens on DNA synthesis and growth of cultured rat liver fat-storing cells. J Clin Invest. 1989;84:1786–93. doi: 10.1172/JCI114363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Maher JJ, Lozier JS, Scott MK. Rat hepatic stellate cells produce cytokine-induced neutrophil chemoattractant in culture and in vivo. Am J Physiol. 1998;275:G847–53. doi: 10.1152/ajpgi.1998.275.4.G847. [DOI] [PubMed] [Google Scholar]
- 45.Lotersztajin S, Julien B, Teixeira-Clerc F, Grenard P, Mallat A. Hepatic fibrosis: molecular mechanisms and drug targets. Annu Rev Pharmacol Toxicol. 2005;45:605–28. doi: 10.1146/annurev.pharmtox.45.120403.095906. [DOI] [PubMed] [Google Scholar]
- 46.Benyon RC, Arthur MJ. Extracellular matrix degradation and the role of hepatic stellate cells. Semin Liver Dis. 2001;21:373–84. doi: 10.1055/s-2001-17552. [DOI] [PubMed] [Google Scholar]
- 47.Iredale JP, Benyon RC, Pickering J, McCullen M, Northrop M, Pawley S, et al. Mechanisms of spontaneous resolution of rat liver fibrosis. Hepatic stellate cell apoptosis and reduced hepatic expression of metalloproteinase inhibitors. J Clin Invest. 1998;102:538–49. doi: 10.1172/JCI1018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nie QH, Duan GR, Lou XD, Xie YM, Luo H, Zhou YX, et al. Expression of TIMP-1 and TIMP-2 in rats with hepatic fibrosis. World J Gastroenterol. 2004;10:86–90. doi: 10.3748/wjg.v10.i1.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and water resources development: systematic review, meta-analysis, and estimates of people at risk. Lancet Infect Dis. 2006;6:411–25. doi: 10.1016/S1473-3099(06)70521-7. [DOI] [PubMed] [Google Scholar]
- 50.Olds GR, Griffin A, Kresina TF. Dynamics of collagen accumulation and polymorphism in murine Schistosoma japonicum. Gastroenterology. 1985;89:617–24. doi: 10.1016/0016-5085(85)90459-7. [DOI] [PubMed] [Google Scholar]
- 51.Grimaud JA. Cell-matrix interactions in schistosomal portal fibrosis: a dynamic event. Mem Inst Oswaldo Cruz. 1987;82(Suppl 4):55–65. doi: 10.1590/s0074-02761987000800010. [DOI] [PubMed] [Google Scholar]
- 52.Cheever AW, Williams ME, Wynn TA, Finkelman FD, Seder RA, Cox TM, et al. Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing TH2 cytokines while decreasing egg-induced hepatic fibrosis. J Immunol. 1994;153:753–9. [PubMed] [Google Scholar]
- 53.Wynn TA, Thompson RW, Cheever AW, Mentink-Kane MM. Immunopathogenesis of schistosomiasis. Immunol Rev. 2004;201:156–67. doi: 10.1111/j.0105-2896.2004.00176.x. [DOI] [PubMed] [Google Scholar]
- 54.Fallon PG, Richardson EJ, McKenzie GJ, McKenzie AN. Schistosome infection of transgenic mice defines distinct and contrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic agent. J Immunol. 2000;164:2585–91. doi: 10.4049/jimmunol.164.5.2585. [DOI] [PubMed] [Google Scholar]
- 55.Rojkind M, Mourelle M. Modification during regeneration and repair in collagen. In: Nimni ME, editor. The liver as bioecological system. Boca, Raton, Florida: CRC Press; 1988. pp. 137–59. [Google Scholar]
- 56.Matsui Y, Okuda Y, Nakagawa M, Kwon AH, Minoura T, Hiramatsu Y, et al. Effect of hepatocyte volume on energy status in the cirrhotic rat liver. J Gastroenterol Hepatol. 1994;9:613–9. doi: 10.1111/j.1440-1746.1994.tb01570.x. [DOI] [PubMed] [Google Scholar]
- 57.Yamada S, Hirayama C. Clinical significance of serum hydroxyproline-containing peptides with special reference to hyproprotein. Eur J Clin Invest. 1983;13:129–33. doi: 10.1111/j.1365-2362.1983.tb00077.x. [DOI] [PubMed] [Google Scholar]
- 58.George J, Chandrakasan G. Glycoprotein metabolism in dimethylnitrosamine induced hepatic fibrosis in rats. Int J Biochem Cell Biol. 1996;28:353–61. doi: 10.1016/1357-2725(95)00140-9. [DOI] [PubMed] [Google Scholar]
- 59.Ala-Kokko L, Pihlajaniemi T, Myers JC, Kivirikko KI, Savolainen ER. Gene expression of types, I, III, and IV collagens in hepatic fibrosis induced by dimethylnitrosamine in the rat. Biochem J. 1987;244:75–9. doi: 10.1042/bj2440075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.George J, Chandrakasan G. Molecular characteristics of dimethylnitrosamine induced fibrotic liver collagen. Biochim Biophys Acta. 1996;1292:215–22. doi: 10.1016/0167-4838(95)00202-2. [DOI] [PubMed] [Google Scholar]
- 61.Garcia–Trevijano ER, Iraburu MJ, Fontana L, Dominguez-Rosales JA, Auster A, Covarrubias-Pinedo A, et al. Transforming growth factor β1 induces the expression of alpha1(I) type(I) procollagen mRNA by a hydrogen peroxide-C/EBP beta-dependent mechanism in rat hepatic stellate cells. Hepatology. 1999;29:960–70. doi: 10.1002/hep.510290346. [DOI] [PubMed] [Google Scholar]
- 62.Panduro A, Shalaby F, Biempica L, Shafritz DA. Changes in albumin, a-fetoprotein and collagen gene transcription in CCl4-induced hepatic fibrosis. Hepatology. 1990;8:259–66. doi: 10.1002/hep.1840080212. [DOI] [PubMed] [Google Scholar]
- 63.George J. Mineral metabolism in dimethylnitrosamine-induced hepatic fibrosis. Clin Biochem. 2006;39:984–91. doi: 10.1016/j.clinbiochem.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 64.Kanzler S, Lohse AW, Keil A, Henninger J, Dienes HP, Schirmacher P, et al. TGF-β1 in liver fibrosis: an inducible transgenic mouse model to study liver fibrogenesis. Am J Physiol Liv Pathol. 1999;276:G1059–68. doi: 10.1152/ajpgi.1999.276.4.G1059. [DOI] [PubMed] [Google Scholar]
- 65.Imbert-Bismut F, Ratziu V, Pieroni L, Charlotte F, Benhamou Y, Poynard T. Biochemical markers of liver fibrosis in patients with hepatitis C virus infection: A prospective study. Lancet. 2001;357:1069–75. doi: 10.1016/S0140-6736(00)04258-6. [DOI] [PubMed] [Google Scholar]
- 66.Kawser CA, Iredale JP, Winwood PJ, Arthur MJ. Rat hepatic stellate cell expression of α 2-macroglobulin is a feature of cellular activation: implications for matrix remodeling in hepatic fibrosis. Clin Sci (Lond) 1998;95:179–86. [PubMed] [Google Scholar]
- 67.Poynard T, Bedossa P, Mathurin P, Ratziu V, Paradis V. Apolipoprotein A1 and hepatic fibrosis. J Hepatol. 1995;22(2 Suppl):107–10. [PubMed] [Google Scholar]
- 68.Hashimoto N, Ishikawa Y, Utsunomiya J. Effects of portacaval shunt, transposition and dimethylnitrosamine-induced chronic liver injury on pancreatic hormones and amino acid in dogs. J Surg Res. 1989;46:35–40. doi: 10.1016/0022-4804(89)90179-0. [DOI] [PubMed] [Google Scholar]
- 69.George J, Chandrakasan G. Lactate dehydrogenase isoenzymes in dimethylnitrosamine induced hepatic fibrosis in rats. J Clin Biochem Nutr. 1997;22:51–62. [Google Scholar]
- 70.Ahmad R, Ahmad S, Khan NU, Hasnain A. Operculina turpethum attenuates NDMA-induced toxic liver injury and clastogenicity in rats. Chem Biol Interact. 2009;181:145–53. doi: 10.1016/j.cbi.2009.06.021. [DOI] [PubMed] [Google Scholar]
- 71.George J, Suguna L, Jayalakshmi R, Chandrakasan G. Efficacy of silymarin and curcumin on dimethylnitrosamine induced liver fibrosis in rats. Biomedicine. 2006;26:18–26. [Google Scholar]
- 72.Attallah AM, Toson el-SA, El-Waseef AM, Abo-Seif MA, Omran MM, Shiha GE. Discriminant function based on hyaluronic acid and its degrading enzymes and degradation products for differentiating cirrhotic from non-cirrhotic liver disease patients in chronic HCV infection. Clin Chim Acta. 2006;369:66–72. doi: 10.1016/j.cca.2006.01.004. [DOI] [PubMed] [Google Scholar]
- 73.George J. Elevated serum β-glucuronidase reflects hepatic lysosomal fragility following toxic liver injury in rats. Biochem Cell Biol. 2008;86:235–43. doi: 10.1139/o08-038. [DOI] [PubMed] [Google Scholar]
- 74.Snodgrass PJ. Urea cycle enzyme activities are normal and inducible by a high-protein diet in CCl4 cirrhosis of rats. Hepatology. 1989;9:373–9. doi: 10.1002/hep.1840090306. [DOI] [PubMed] [Google Scholar]
- 75.Vendemiale G, Grattagliano I, Caruso ML, Servidelio G, Valintini AM, Pirrellli M, et al. Increased oxidative stress in dimethylnitrosoamine-induced liver fibrosis in rats. Effect of N-acetylcysteine and interferon-alpha. Toxicol Appl Pharmacol. 2001;175:130–9. doi: 10.1006/taap.2001.9234. [DOI] [PubMed] [Google Scholar]
- 76.George J, Chandrakasan G. Biochemical abnormalities during the progression of hepatic fibrosis induced by dimethylnitrosoamine. Clin Biochem. 2000;33:563–70. doi: 10.1016/s0009-9120(00)00170-3. [DOI] [PubMed] [Google Scholar]
- 77.Kobayashi H, Horikoshi K, Yamataka A, Yamataka T, Okazaki T, Lane GJ, et al. Hyaluronic acid: A specific prognostic indicator of hepatic damage in biliary atresia. J Pediatr Surg. 1999;34:1791–4. doi: 10.1016/s0022-3468(99)90314-7. [DOI] [PubMed] [Google Scholar]
- 78.Morrison WL, Bouchier IAD, Gibson JN, Rennie MJ. Skeletal muscle and whole body protein turnover in cirrhosis. Clin Sci (Lond) 1990;78:613–9. doi: 10.1042/cs0780613. [DOI] [PubMed] [Google Scholar]
- 79.Ahmad A, Fatima R, Maheshwari V, Ahmad R. Effect of N’-Nitrosodimethylamine on red blood cell rheology and proteomic profiles of brain in male albino rats. Interdiscip Toxicol. 2011;4:125–31. doi: 10.2478/v10102-011-0020-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.George J. Ascorbic acid concentrations in dimethylnitrosamine-induced hepatic fibrosis in rats. Clin Chim Acta. 2003;335:39–47. doi: 10.1016/s0009-8981(03)00285-7. [DOI] [PubMed] [Google Scholar]
- 81.Bengoa JM, Sitrin MD, Meredith S, Kelly SE, Shah N, Baker AL, et al. Intestinal calcium absorption and vitamin D status in chronic cholestatic liver disease. Hepatology. 1984;4:261–5. doi: 10.1002/hep.1840040215. [DOI] [PubMed] [Google Scholar]
- 82.Masuda S, Okanao T, Osawa K, Shinjo M, Suematsu T, Kobayashi T. Concentrations of vitamin D-binding protein and vitamin D metabolites in plasma of patients with liver cirrhosis. J Nutr Sci Vitaminol (Tokyo) 1989;35:225–34. doi: 10.3177/jnsv.35.225. [DOI] [PubMed] [Google Scholar]
- 83.Wong F, Blendis L. Pathophysiology of sodium retention and ascites formation in cirrhosis: Role of atrial natriuretic factor. Semin Liver Dis. 1994;14:59–70. doi: 10.1055/s-2007-1007298. [DOI] [PubMed] [Google Scholar]
- 84.Castello L, Pirisi M, Sainaghi PP, Bartoli E. Hyponatremia in liver cirrhosis: pathophysiological principles of management. Dig Liver Dis. 2005;37:73–81. doi: 10.1016/j.dld.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 85.Weiner ID, Wingo CS. Hypokalemia-consequences, causes and correction. J Am Soc Nephrol. 1997;8:1179–88. doi: 10.1681/ASN.V871179. [DOI] [PubMed] [Google Scholar]
- 86.Castro A, Ros J, Jimenez W, Claria J, Llibre J, Leivas A, et al. Intracellular calcium concentration in vascular smooth muscle cells of rats with cirrhosis. J Hepatol. 1994;21:521–6. doi: 10.1016/s0168-8278(94)80096-0. [DOI] [PubMed] [Google Scholar]
- 87.Rocchi E, Borella P, Borghi A, Paolillo F, Pradelli M, Farina F, et al. Zinc and magnesium in liver cirrhosis. Eur J Clin Invest. 1994;24:149–55. doi: 10.1111/j.1365-2362.1994.tb00980.x. [DOI] [PubMed] [Google Scholar]
- 88.Suzuki K, Oyama R, Hayashi E, Arakawa Y. Liver diseases and essential trace elements. Nippon Rinsho. 1996;54:85–92. [PubMed] [Google Scholar]
- 89.Cocchetto DM, Tschanz C, Bjornsson TD. Decreased rate of creatinine production in patients with hepatic disease: implications for estimation of creatinine clearance. Ther Drug Monit. 1983;5:161–8. doi: 10.1097/00007691-198306000-00002. [DOI] [PubMed] [Google Scholar]
- 90.Takabatake T, Ohta H, Ishida Y, Hara H, Ushiogi Y, Hattori N. Low serum creatinine levels in severe hepatic disease. Arch Intern Med. 1988;148:1313–5. [PubMed] [Google Scholar]
- 91.Ariosto F, Riggio O, Cantafora A, Colussi S, Gaudio E, Mechelli C, et al. Carbon tetrachloride induced experimental cirrhosis in the rat: A reappraisal of the model. Eur Surg Res. 1989;21:280–6. doi: 10.1159/000129038. [DOI] [PubMed] [Google Scholar]
- 92.Kim KY, Rhim T, Choi I, Kim SS. N-Acetylcysteine induces cell cycle arrest in hepatic stellate cells through its reducing activity. J Biol Chem. 2001;276:40591–8. doi: 10.1074/jbc.M100975200. [DOI] [PubMed] [Google Scholar]
- 93.Meurer SK, Lahme B, Tihaa L, Weiskirchen R, Gressner AM. N-acetyl-l-cysteine suppresses TGF-β signaling at distinct molecular steps: the biochemical and biological efficacy of a multifunctional, antifibrotic drug. Biochem Pharmacol. 2005;70:1026–34. doi: 10.1016/j.bcp.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 94.Zardi EM, Dobrina A, Amoroso A, Afeltra A. Prostacyclin in liver disease: a potential therapeutic option. Expert Opin Biol Ther. 2007;7:785–90. doi: 10.1517/14712598.7.6.785. [DOI] [PubMed] [Google Scholar]
- 95.Yoshidome H, Kato A, Edwards MJ, Lentsch AB. Interleukin-10 suppresses hepatic ischemia/reperfusion injury in mice: implications of a central role for nuclear factor kappaB. Hepatology. 1999;30:203–8. doi: 10.1002/hep.510300120. [DOI] [PubMed] [Google Scholar]
- 96.Ulloa L, Doody J, Massague J. Inhibition of transforming growth factor-beta/SMAD signaling by the interferon-gamma/STAT pathway. Nature. 1999;397:710–13. doi: 10.1038/17826. [DOI] [PubMed] [Google Scholar]
- 97.Hui AY, Cheng AS, Chan HL, Go MY, Chan FK, Sakata R, et al. Effect of prostaglandin E2 and prostaglandin I 2 on PDGF-induced proliferation of LI90, a human hepatic stellate cell line. Prostaglandins Leukot Essent Fatty Acids. 2004;71:329–33. doi: 10.1016/j.plefa.2004.04.007. [DOI] [PubMed] [Google Scholar]
- 98.Zhou Y, Zheng S, Lin J, Zhang QJ, Chen A. The interruption of PDGF and EGF signaling pathways by curcumin stimulates gene expression of PPAR-gamma in rat activated hepatic stellate cell in vitro. Lab Invest. 2007;87:488–98. doi: 10.1038/labinvest.3700532. [DOI] [PubMed] [Google Scholar]
- 99.Han J, Hajjar DP, Tauras JM, Feng J, Gotto AM, Jr, Nicholson AC. Transforming growth factor-β1 (TGF- β1) and TGF- β2 decrease expression of CD36, the type B scavenger receptor, through mitogen-activated protein kinase phosphorylation of peroxisome proliferator-activated receptor-g. J Biol Chem. 2000;275:1241–6. doi: 10.1074/jbc.275.2.1241. [DOI] [PubMed] [Google Scholar]
- 100.Galli A, Crabb DW, Ceni E, Salzano R, Mello T, Svegliati-Baroni G, et al. Antidiabetic thiazolidinediones inhibit collagen synthesis and hepatic stellate cell activation in vivo and in vitro. Gastroenterology. 2002;122:1924–40. doi: 10.1053/gast.2002.33666. [DOI] [PubMed] [Google Scholar]
- 101.Sun K, Wang Q, Huang XH. PPAR-gamma inhibits growth of rat hepatic stellate cells and TGF beta-induced connective tissue growth factor expression. Acta Pharmacol Sin. 2006;27:715–23. doi: 10.1111/j.1745-7254.2006.00299.x. [DOI] [PubMed] [Google Scholar]
- 102.Ding X, Saxena NK, Lin S, Xu A, Srinivasan S, Anania FA. The role of leptin and adiponectin: a novel paradigm in adipocytokine regulation of liver fibrosis and stellate cell biology. Am J Pathol. 2005;166:1655–69. doi: 10.1016/S0002-9440(10)62476-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Sanz S, Pucilowska JB, Liu S, Rodríguez-Ortigosa CM, Lund PK, Brenner DA, et al. Expression of insulin-like growth factor I by activated hepatic stellate cells reduces fibrogenesis and enhances regeneration after liver injury. Gut. 2005;54:134–41. doi: 10.1136/gut.2003.024505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Elsharkawy AM, Wright MC, Hay RT, Arthur MJ, Hughes T, Bahr MJ, et al. Persistent activation of nuclear factor-kappa B in cultured rat hepatic stellate cells involves the induction of potentially novel Rel-like factors and prolonged changes in the expression of IkappaB family proteins. Hepatology. 1999;30:761–9. doi: 10.1002/hep.510300327. [DOI] [PubMed] [Google Scholar]
- 105.Oakley F, Meso M, Iredale JP, Green K, Marek CJ, Zhou X, et al. Inhibition of inhibitor of kB kinases stimulates hepatic stellate cell apoptosis and accelerated recovery from rat liver fibrosis. Gastroenterology. 2005;128:108–20. doi: 10.1053/j.gastro.2004.10.003. [DOI] [PubMed] [Google Scholar]
- 106.Sonoyama T, Tamura N, Miyashita K, Park K, Oyamada M, Taura D, et al. Inhibition of hepatic damage and liver fibrosis by brain natiuretic peptide. FEBS Lett. 2009;583:2067–70. doi: 10.1016/j.febslet.2009.05.025. [DOI] [PubMed] [Google Scholar]
- 107.Wang L, Potter JJ, Rennie-Tankersley L, Novitskiy G, Sipes J, Mezey E. Effect of retinoic acid on the development of liver fibrosis produced by carbon tetrachloride in mice. Biochim Biophys Acta. 2007;1772:66–71. doi: 10.1016/j.bbadis.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 108.Horie T, Sakaida I, Yokoya F, Nakajo M, Sonaka I, Okitab K. L-cysteine administration prevents liver fibrosis by suppressing hepatic stellate cell proliferation and activation. Biochem Biophys Res Commun. 2003;305:94–100. doi: 10.1016/s0006-291x(03)00691-0. [DOI] [PubMed] [Google Scholar]
- 109.Galli A, Crabb D, Price D, Ceni E, Salzano F, Surrenti C, et al. Peroxisome proliferator-activated receptor γ transcriptional regulation is involved in platelet-derived growth factor-induced proliferation of human hepatic stellate cells. Hepatology. 2000;31:101–8. doi: 10.1002/hep.510310117. [DOI] [PubMed] [Google Scholar]
- 110.Okuno M, Akita K, Moriwaki H, Kawada N, Ikeda K, Kaneda K, et al. Prevention of rat hepatic fibrosis by the protease inhibitor, camostat mesilate, via reduced generation of active TGF-beta. Gastroenterology. 2001;120:1784–800. doi: 10.1053/gast.2001.24832. [DOI] [PubMed] [Google Scholar]
- 111.Williams EJ, Benyon RC, Trim N, Hadwin R, Grove BH, Arthur MJ, et al. Relaxin inhibits effective collagen deposition by cultured hepatic stellate cells and decreases rat liver fibrosis in vivo. Gut. 2001;49:577–83. doi: 10.1136/gut.49.4.577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Bennett RG, Mahan KJ, Gentry-Nielsen MJ, Tuma DJ. Relaxin receptor expression in hepatic stellate cells and in cirrhotic rat liver tissue. Ann N Y Acad Sci. 2005;1041:185–9. doi: 10.1196/annals.1282.027. [DOI] [PubMed] [Google Scholar]
- 113.von Mehren M. Targeted therapy with imatinib: hits and misses? J Clin Oncol. 2005;23:8–10. doi: 10.1200/JCO.2005.09.961. [DOI] [PubMed] [Google Scholar]
- 114.Kerkela R, Grazette L, Yacobi R, Iliescu C, Patten R, Beahm C, et al. Cardio toxicity of the cancer therapeutic agent imatinib mesilate. Nat Med. 2006;12:908–16. doi: 10.1038/nm1446. [DOI] [PubMed] [Google Scholar]
- 115.Friedman SL. The answer: Angiotensin II. The question: what do inflammation, oxidant stress and fibrogenesis have in common? J Hepatol. 2004;40:1050–2. doi: 10.1016/j.jhep.2004.04.001. [DOI] [PubMed] [Google Scholar]
- 116.Wei HS, Lu HM, Li DG, Zhan YT, Wang ZR, Huang X, et al. The regulatory role of AT1 receptor on activated HSCs in hepatic fibrogenesis: effects of RAS inhibitors on hepatic fibrosis induced by CCl4. World J Gastroenterol. 2000;6:824–8. doi: 10.3748/wjg.v6.i6.824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kurikawa N, Suga M, Kuroda S, Yamada K, Ishikawa H. An angiotensin II type 1 receptor antagonist, olmesartan medoxomil, improves experimental liver fibrosis by suppression of proliferation and collagen synthesis in activated hepatic stellate cells. Br J Pharmacol. 2003;139:1085–94. doi: 10.1038/sj.bjp.0705339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zhang JP, Zhang M, Zhou JP, Liu FT, Zhou B, Xie WF, et al. Antifibrotic effects of matrine on in vitro and in vivo models of liver fibrosis in rats. Acta Pharmacol Sin. 2001;22:183–6. [PubMed] [Google Scholar]
- 119.Miyazaki T, Karube M, Matsuzaki Y, Ikegami T, Doy M, Tanaka N, et al. Taurine inhibits oxidative damage and prevents fibrosis in carbon tetrachloride-induced hepatic fibrosis. J Hepatol. 2005;43:117–25. doi: 10.1016/j.jhep.2005.01.033. [DOI] [PubMed] [Google Scholar]
- 120.Zhan Y, Li D, Wei H, Wang Z, Huang X, Xu Q, et al. Emodin on hepatic fibrosis in rats. Chin Med J (Engl) 2000;113:599–601. [PubMed] [Google Scholar]
- 121.Tan LX, Li XS, Liu ZQ, Liu LY. Effects of combination therapy of rehin and tetramethylpyrazine on experimental hepatic fibrosis induced by carbon tetrachloride. Zhonghua Gan Zang Bing Za Zhi. 2004;12:692–3. [PubMed] [Google Scholar]
- 122.Liu SQ, Yu JP, Chen HL, Luo HS, Chen SM, Yu HG. Therapeutic effects and molecular mechanisms of Ginkgo biloba extract on liver fibrosis in rats. Am J Chin Med. 2006;34:99–114. doi: 10.1142/S0192415X06003679. [DOI] [PubMed] [Google Scholar]
- 123.Chen MH, Chen SH, Wang QF, Chen JC, Chang DC, Hsu SL, et al. The molecular mechanism of gypenosides-induced G1 growth arrest of rat hepatic stellate cells. J Ethnopharmacol. 2008;117:309–17. doi: 10.1016/j.jep.2008.02.009. [DOI] [PubMed] [Google Scholar]
- 124.Wasser S, Ho JMS, Ang HK, Tan CEL. Salvia miltiorrhiza reduces experimentally-induced hepatic fibrosis in rats. J Hepatol. 1998;29:760–71. doi: 10.1016/s0168-8278(98)80257-2. [DOI] [PubMed] [Google Scholar]
- 125.Shalaby NM, Maghraby AS, El-Hagrassy AM. Effect of Daucus carota var. boissieri extracts on immune response of Schistosoma mansoni infected mice. Folia Microbiol. 1999;44:441–8. doi: 10.1007/BF02903720. [DOI] [PubMed] [Google Scholar]
- 126.Massoud A. Myrrh; a schistosomicide for human Schistosomiasis mansoni. Ain Shams Med J. 1999;50:1401–17. [Google Scholar]
- 127.Abdel Aziz SS, El-Badawy NM. Experimental trials of an artemisinin derivative in treatment of Schistosoma mansoni infected mice. J Egypt Soc Parasitol. 2000;30:295–303. [PubMed] [Google Scholar]
- 128.McGaw LJ, Rabe T, Sparg SG, Jager AK, Eloff JN, van Staden J. An investigation on the biological activity of Combretum species. J Ethnopharmacol. 2001;75:45–50. doi: 10.1016/s0378-8741(00)00405-0. [DOI] [PubMed] [Google Scholar]
- 129.El-Qadri AA, Emara MM. Effect of Nigella sativa crude oil on skin penetration and viability of Schistosoma mansoni cercariae. New Egypt J Med. 1994;11:431–6. [Google Scholar]
- 130.Mostafa OM. Experimental use of black-seed oil against Schistosoma mansoni in albino mice: I. Some parasitological and biochemical parameters. Egypt J Med Lab Sci. 2001;10:99–113. [Google Scholar]
- 131.Mahmoud MR, El-Abhar HS, Saleh S. The effect of Nigella sativa oil against the liver damage induced by Schistosoma mansoni infection in mice. J Ethnopharmacol. 2002;70:1–11. doi: 10.1016/s0378-8741(01)00310-5. [DOI] [PubMed] [Google Scholar]
- 132.Mohamed AM, Metwally NM, Mahmoud SS. Sativa seeds against Schistosoma mansoni different stages. Mem Inst Oswaldo Cruz. 2005;100:205–11. doi: 10.1590/s0074-02762005000200016. [DOI] [PubMed] [Google Scholar]
- 133.Sanderson L, Bartlett A, Whitefield PJ. In vitro and in vivo studies on the bioactivity of ginger (Zingiber officinale) extract towards adult schistosomes and their egg production. J Helminthol. 2002;76:241–7. doi: 10.1079/JOH2002116. [DOI] [PubMed] [Google Scholar]
- 134.Ahmed AH, Rifaat MM. Effects of Solanum nigrum leaves water extract on the penetration and infectivity of Schistosoma mansoni cercariae. J Egypt Soc Parasitol. 2005;35:33–40. [PubMed] [Google Scholar]
- 135.Riad NHA, Fares NH, Mostafa OMS, Mahmoud YI. The effect of garlic on some parasitological and on hepatic tissue reactions in experimental Schistosomiasis mansoni. J Appl Sci Res. 2007;3:949–60. [Google Scholar]
- 136.Riad NHA, Fares NH, Mostafa OMS, Mahmoud YI. The effect of garlic on murine schistosomiasis mansoni: A histological and ultrastructural study on the ileum. Res J Med Med Sci. 2008;3:188–201. [Google Scholar]
- 137.Allam G. Immunomodulatory effects of curcumin treatment on murine schistosomiasis mansoni. Immunobiology. 2009;214:712–27. doi: 10.1016/j.imbio.2008.11.017. [DOI] [PubMed] [Google Scholar]
- 138.Dajem SMB, Shati AA, Adly MA, Ahmed OM, Ibrahim EH, Mostafa OMS. Green tea (Camellia sinensis) ameliorates female Schistosoma mansoni-induced changes in the liver of Balb/C mice. Saudi J Biol Sci. 2011;18:361–8. doi: 10.1016/j.sjbs.2011.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Chatterjee S, Mbaye A, Alfidja AT, Weyler J, Scott JT, Van Damme P, et al. Circulating levels of the neuropeptide hormone somatostatin may determine hepatic fibrosis in Schistosoma mansoni infections. Acta Trop. 2004;90:191–203. doi: 10.1016/j.actatropica.2003.12.002. [DOI] [PubMed] [Google Scholar]
- 140.Chu D, Luo Q, Li C, Gao Y, Yu L, Wei W, et al. Paeoniflorin inhibits TGF-beta1-mediated collagen production by Schistosoma japonicum soluble egg antigen in vitro. Parasitology. 2007;134:1611–21. doi: 10.1017/S0031182007002946. [DOI] [PubMed] [Google Scholar]
- 141.Weiner FR, Czaja MJ, Giambrone MA, Takahashi S, Biempica L, Zern MA. Transcriptional and posttranscriptional effects of dexamethasone on albumin and procollagen messenger RNAs in murine schistosomiasis. Biochemistry. 1987;26:1557–62. doi: 10.1021/bi00380a010. [DOI] [PubMed] [Google Scholar]
- 142.Chen H, He YW, Liu WQ, Zhang JH. Rosiglitazone prevents murine hepatic fibrosis induced by Schistosoma japonicum. World J Gastroenterol. 2008;14:2905–11. doi: 10.3748/wjg.14.2905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Beljaars L, Meijer DK, Poelstra K. Targeting hepatic stellate cells for cell specific treatment of liver fibrosis. Front Biosci. 2002;7:e214–22. doi: 10.2741/A917. [DOI] [PubMed] [Google Scholar]
- 144.Sato Y, Murase K, Kato J, Kobune M, Sato T, Kawano Y, et al. Resolution of liver cirrhosis using vitamin A–coupled liposomes to deliver siRNA against a collagen-specific chaperone. Nat Biotechnol. 2008;26:431–42. doi: 10.1038/nbt1396. [DOI] [PubMed] [Google Scholar]
- 145.Suzuki K, Aoki K, Ohnami S, Yoshida K, Kazui T, Kato N, et al. Adenovirus-mediated gene transfer of interferon alpha improves dimethylnitrosamine-induced liver cirrhosis in rat model. Gene Ther. 2003;10:765–73. doi: 10.1038/sj.gt.3301949. [DOI] [PubMed] [Google Scholar]
- 146.Iimuro Y, Nishio T, Morimoto T, Nitta T, Stefanovic B, Choi SK, et al. Delivery of matrix metalloproteinase-1 attenuates established liver fibrosis in the rat. Gastroenterology. 2003;124:445–58. doi: 10.1053/gast.2003.50063. [DOI] [PubMed] [Google Scholar]
- 147.Arias M, Sauer-Lehnen S, Treptau J, Janoschek N, Theuerkauf I, Buettner R, et al. Adenoviral expression of a transforming growth factor-beta1 antisense mRNA is effective in preventing liver fibrosis in bile-duct ligated rats. BMC Gastroenterol. 2003;3:29–40. doi: 10.1186/1471-230X-3-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Hung KS, Lee TH, Chou WY, Wu CL, Cho CL, Lu CN, et al. Interleukin-10 gene therapy reverses thioacetamide-induced liver fibrosis in mice. Biochem Biophys Res Commun. 2005;336:324–31. doi: 10.1016/j.bbrc.2005.08.085. [DOI] [PubMed] [Google Scholar]
- 149.Wang CH, Lee TH, Lu CN, Chou WY, Hung KS, Concejero AM, et al. Electroporative alpha-MSH gene transfer attenuates thioacetamide-induced murine hepatic fibrosis by MMP and TIMP modulation. Gene Ther. 2006;13:1000–9. doi: 10.1038/sj.gt.3302744. [DOI] [PubMed] [Google Scholar]
- 150.Dooley S, Hamzavi J, Breitkopf K, Wiercinska E, Said HM, Lorenzen J, et al. Smad7 prevents activation of hepatic stellate cells and liver fibrosis in rats. Gastroenterology. 2003;125:178–91. doi: 10.1016/s0016-5085(03)00666-8. [DOI] [PubMed] [Google Scholar]
- 151.Zhong Z, Froh M, Wheeler MD, Smutney O, Lehmann TG, Thurman RG. Viral gene delivery of superoxide dismutase attenuates experimental cholestasis-induced liver fibrosis in the rat. Gene Ther. 2002;9:183–91. doi: 10.1038/sj.gt.3301638. [DOI] [PubMed] [Google Scholar]
- 152.Tsui TY, Lau CK, Ma J, Wu X, Wang YQ, Farkas S, et al. rAAV-mediated stable expression of heme oxygenase-1 in stellate cells: a new approach to attenuate liver fibrosis in rats. Hepatology. 2005;42:335–42. doi: 10.1002/hep.20803. [DOI] [PubMed] [Google Scholar]
- 153.Siller-Lopez F, Sandoval A, Salgado S, Salazar A, Bueno M, Garcia J, et al. Treatment with human metalloproteinase-8 gene delivery ameliorates experimental rat liver cirrhosis. Gastroenterology. 2004;126:1122–33. doi: 10.1053/j.gastro.2003.12.045. [DOI] [PubMed] [Google Scholar]
- 154.Borkham-Kamphorst E, Stoll D, Gressner AM, Weiskirchen R. Antisense strategy against PDGF B-chain proves effective in preventing experimental liver fibrogenesis. Biochem Biophys Res Commun. 2004;321:413–23. doi: 10.1016/j.bbrc.2004.06.153. [DOI] [PubMed] [Google Scholar]
- 155.Matsuno Y, Iwata H, Umeda Y, Takagi H, Mori Y, Kosugi A, et al. Hepatocyte growth factor gene transfer into the liver via the portal vein using electroporation attenuates rat liver cirrhosis. Gene Ther. 2003;10:1559–66. doi: 10.1038/sj.gt.3302052. [DOI] [PubMed] [Google Scholar]
- 156.Kakinuma S, Nakauchi H, Watanabe M. Hepatic stem/progenitor cells and stem-cell transplantation for the treatment of liver disease. J Gastroenterol. 2009;44:167–72. doi: 10.1007/s00535-008-2297-z. [DOI] [PubMed] [Google Scholar]
- 157.Zhou P, Wirthlin L, McGee J, Annett G, Nolta J. Contribution of human hematopoietic stem cells to liver repair. Semin Immunopathol. 2009;31:411–19. doi: 10.1007/s00281-009-0166-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Matsumoto T, Terai S, Oishi T, Kuwashiro F, Fujisawa S, Yamamoto N, et al. Medaka as a model for human nonalcoholic Steatohepatitis. Dis Model Mech. 2010;3:431–40. doi: 10.1242/dmm.002311. [DOI] [PubMed] [Google Scholar]
- 159.Said E, Said SA, Elkashef WF, Gameil NM, Ammar EM. Tranilast ameliorates impaired hepatic functions in Schistosoma mansoni-infected mice. Inflammopharmacology. 2012;20:77–87. doi: 10.1007/s10787-011-0117-1. [DOI] [PubMed] [Google Scholar]