

Abstract
Temozolomide (TMZ) has become a key therapeutic agent in patients with malignant gliomas; however, its survival benefit remains unsatisfactory. Valproic acid (VPA) has emerged as an anticancer drug via inhibition of histone deacetylases (HDACs), but the therapeutic advantages of a combination with VPA and TMZ remain poorly understood. The main aim of the present study was to determine whether an antitumor effect could be potentiated by a combination of VPA and TMZ, especially in TMZ-resistant cell lines. A combination of VPA and TMZ had a significantly enhanced antitumor effect in TMZ-resistant malignant glioma cells (T98 and U138). This enhanced antitumor effect correlated with VPA-mediated reduced O6-methylguanine-DNA methyltransferase (MGMT) expression, which plays an important role in cellular resistance to alkylating agents. In vitro, the combination of these drugs enhanced the apoptotic and autophagic cell death, as well as suppressed the migratory activities in TMZ-resistant cell lines. Furthermore, in vivo efficacy experiment showed that treatment of combination of VPA and TMZ significantly inhibited tumor growth compared with the monotherapy groups of mice. These results suggest that the clinical efficacy of TMZ chemotherapy in TMZ-resistant malignant glioma may be improved by combination with VPA.
1. Introduction
Malignant gliomas are the most common primary tumors of the central nervous system. Although multimodality treatments exist, including extensive tumor resection, radiation therapy, and chemotherapy, their prognosis is poor. Recently, the alkylating agent temozolomide (3,4-dihydro-3-methyl-4-oxoimidazo-[5,1-d]-1,2,3,5-tetrazine-8-carboxamide, TMZ) has received much attention as a treatment for malignant gliomas [1]. A trial of concomitant and adjuvant TMZ in addition to radiotherapy for new glioblastomas demonstrated an increase in median survival from 12.1 to 14.6 months and an increase in the 2-year survival rate from 10 to 26% compared with radiotherapy alone [2]. However, recent studies have indicated that the resistance to TMZ observed in malignant gliomas is related to the DNA repair enzyme, O6-methylguanine-DNA methyltransferase (MGMT), leading to the replication of DNA and the growth of tumors [3–6].
Valproic acid (VPA) is an approved drug for the treatment of epileptic seizures, bipolar disorders, and migraine and acts via inhibition of the transamination of gamma-aminobutyric acid. VPA is a short-chain fatty acid that inhibits histone deacetylases (HDACs) [7–9]. HDACs play an important role in chromatin remodeling and gene expression via posttranslational modification of chromatin-associated histones. HDAC inhibition induces tumor cell differentiation, apoptosis, and growth arrest [10, 11]. VPA has been examined as an HDAC inhibitor (HDACI) in numerous preclinical and clinical trials for solid tumors and leukemias [12, 13].
Recent investigation shows that VPA enhanced the apoptotic cell death by TMZ in human glioma cell lines [14]. However, antitumor effects of VPA in TMZ-resistant glioma cells remain poorly documented. In this study, we investigated the ability of VPA to increase the sensitivity of four human malignant glioma cell lines (U87, U138, T98, and U251) to the cytotoxic effects of TMZ in vitro. We also examined whether VPA regulates the expression of MGMT and enhances cytotoxicity in TMZ-resistant glioma cells.
2. Materials and Methods
2.1. Cell Culture and Reagents
U87, U138, T98, and U251 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). U87 and T98 cells were grown in Eagle's Minimum Essential Medium (Invitrogen, Grand Island, NY), and U138 and U251 cells were grown in Dulbecco's modified Eagle's medium (Invitrogen). Both media were supplemented with 2 mmol/L L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 10% fetal bovine serum (FBS), purchased from Invitrogen (Carlsbad, CA, USA). Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2. Temozolomide (3,4-dihydro-3-methyl-4-oxoimidazo-[5,1-d]-1,2,3,5-tetrazine-8-carboxamide, TMZ), valproic acid (VPA), and autophagy inhibitor 3-methyladenine (3-MA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The caspase inhibitor Z-VAD-FMK was acquired from R&D Systems (Minneapolis, MN, USA).
2.2. Growth Inhibitory Effect of VPA or TMZ in Glioma Cells
The inhibition of the growth of the four glioma cell lines by VPA or TMZ was evaluated from the concentration required for 50% growth inhibition (IC50) compared with untreated controls. Cells were seeded into wells of a 96-well plate at a density of 1 × 103 cells/well in medium containing 10% FBS and were incubated for 72 h. Subsequently, cells were washed twice with the medium and incubated with fresh medium (control), medium containing 0.5–16 mM VPA, or medium containing 0.5–1,000 μM TMZ. After exposure to the various concentrations of VPA or TMZ for 48 h, cells were detached by trypsinization and the viable cell population was determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), as described previously [15].
2.3. Western Blot Analysis
Antibodies were obtained from commercial sources: anti-MGMT from Cell Signaling Technology (Danvers, MA, USA), anti-β-actin from Applied Biological Materials (Richmond, BC, Canada), anti-LC3B from Sigma, and anti-Bcl-2, anti-Bak, and anti-caspase-3 from New England Biolabs (Ipswich, MA, USA). For western blot analysis, cells were rinsed with phosphate-buffered saline (PBS) and lysed for 30 min on ice in RIPA-B buffer (0.5% Nonidet P-40, 20 mM Tris, pH 8.0, 50 mM NaCl, 50 mM NaF, 100 μM Na3VO4, 1 mM dithiothreitol, and 50 μg/mL of phenylmethanesulfonyl fluoride). The insoluble material was removed by centrifugation at 12,000 rpm for 20 min. The supernatant was resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the resolved proteins were transferred to a nitrocellulose blot membrane. The blots were blocked in PBS with 5% skim milk and 0.05% Tween-20, incubated with the appropriate antibodies, and subsequently incubated with secondary antibodies conjugated with horseradish peroxidase. The blots were assayed using an enhanced chemiluminescence detection system (Amersham Biosciences, Piscataway, NJ, USA).
2.4. Cell Migration Assay
The migratory ability of glioma cells (T98 and U138) was determined using Transwell plates (Corning Costar, Cambridge, MA, USA) that were 6.5 mm in diameter with 8 μm pore filters. In brief, cells were suspended in serum-free medium and seeded into the upper well, and 600 μL of 20% FBS-containing medium was placed in the lower well of a Transwell plate. Following incubation for 5 h at 37°C, cells that had not migrated from the upper side of the filter were scraped off with a cotton swab, and filters were stained with the Diff-Quik™ three-step stain set (Sysmex, Kobe, Japan). The number of cells that had migrated to the lower side of the filter was counted under a light microscope at ×200 magnification in five randomly selected fields.
2.5. Animal Experiments
Two established human glioblastoma tumor cell lines (T98 and U138) were used in this experiment. However, U138 cell line was not tumorigenic when injected into BALB/c nude mice (female, 6 weeks old). BALB/c nude mice bearing T98 tumors were randomized to four groups (n = 5 in each group) and treated when the subcutaneous tumors had reached a volume between 100 and 200 mm3. VPA (300 mg/kg) was administered i.p. 6 h before the i.p. injection of TMZ (50 mg/kg). Control (PBS) mice, or mice receiving VPA or TMZ alone, also received the corresponding vehicle. Treatments were repeated at 24 h intervals for a total of five doses. Tumor length (a) and width (b) were measured in situ at 4-day intervals using digital calipers, and tumor volumes were calculated according to the following formula: V(mm3) = a × b2/2. The significance of differences between control and treated tumors was evaluated using ANOVA.
2.6. Statistical Analysis
All data are expressed as means ± SEMs. The significance of differences between different test conditions was determined using one-way ANOVA and Student's t-test. Probability values <0.05 were considered significant.
3. Results
3.1. Antitumor Effect of VPA and TMZ on Human Malignant Glioma Cells
To determine the effects of VPA or TMZ alone in malignant glioma cells, we treated four malignant glioma cell lines with 0–16 mM VPA or 0–1,000 μM TMZ for 72 h and assessed the number of viable cells. VPA and TMZ had a growth inhibitory effect on all tumor cell lines in a dose-dependent manner. The IC50 value of VPA for U87 and U251 cells was 6 mM, whereas the IC50 value of VPA for T98 and U138 cells exceeded 8 mM. In contrast, the IC50 of TMZ for U87 and U251 was <500 μM, although T98 and U138 cells were resistant to TMZ (IC50 > 500 μM; Figure 1). Based on these results, a concentration of VPA of 4 mM and a concentration of TMZ of 50 μM, which were lower concentrations than the respective IC50 value, were chosen for the following in vitro combination experiments.
Figure 1.
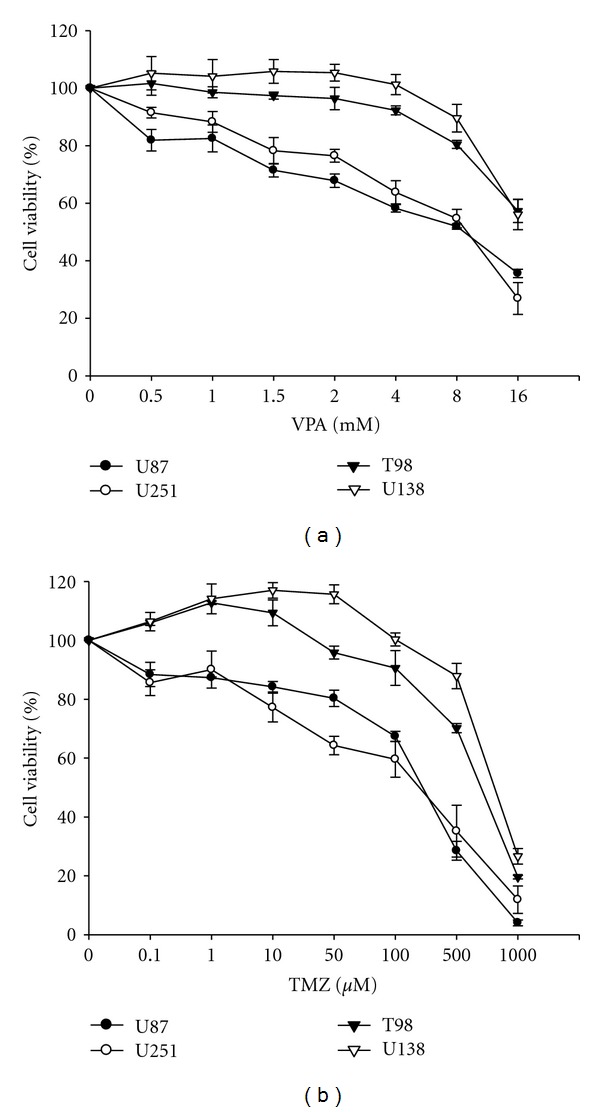
Sensitivity of four human glioma cell lines to TMZ and VPA. (a) The number of viable cells was counted at 72 h after the addition of VPA (0–16 mM) to the culture medium. (b) The viability of glioma cells was also analyzed via MTT assay 72 h after TMZ (0–1,000 μM) treatment. Points, mean; bars, SE. The results are representative of three independent experiments.
3.2. Combined Effect of VPA and TMZ in Human Malignant Glioma Cells
To assess the effect of the combination of VPA and TMZ in the malignant glioma cell lines, cells were incubated in culture medium containing 4 mM VPA, 50 μM TMZ, or a combination of 4 mM VPA and 50 μM TMZ for 72 h. As shown in Figure 2(a), the combination of VPA and TMZ had a combined antitumor effect in U87 and U251 cells. In addition, a significant enhanced antitumor effect of the combined treatment of VPA and TMZ was also observed in T98 (P = 0.034, combination treatment versus single treatment) and U138 cells (P = 0.042, combination treatment versus single treatment; Figure 2(b)). These results suggest that the combination of VPA with TMZ has combined or enhanced antitumor effects in the malignant glioma cell lines. Furthermore, this finding implies that the cell growth inhibitory effect of TMZ is enhanced by VPA in TMZ-resistant cells.
Figure 2.
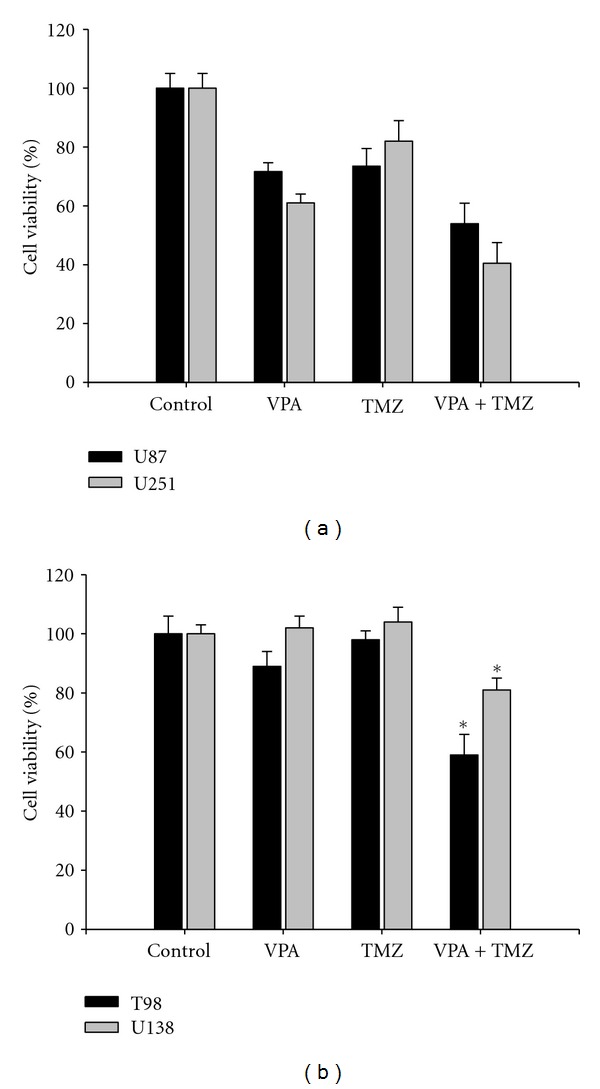
Antitumor effect of a combination of VPA with TMZ in glioma cell lines. Malignant glioma cell lines were treated with VPA (4 mM) or TMZ (50 μM), alone or in combination, for 72 h. (a) The combination of VPA with TMZ had combined antitumor effects in U87 and U251 cells. (b) Significant enhanced antitumor effects were observed in T98 and U138 cells (TMZ-resistant glioma cell lines). Columns, mean; bars, SE. *P < 0.05, ANOVA. The results are representative of three independent experiments.
3.3. Effect of VPA on the Expression of MGMT in TMZ-Resistant Glioma Cells
To examine the involvement of MGMT in the malignant glioma cell lines, the expression of the MGMT protein was analyzed by western blot. We detected expression of MGMT in the TMZ-resistant cell lines T98 and U138. In contrast, MGMT protein expression was not detected in the TMZ-sensitive cell lines U87 and U251 (Figure 3(a)). This result indicates that the sensitivity of TMZ is correlated with the expression of the MGMT protein, which is consistent with the results of a previous study [3]. To determine the effect of VPA on the expression of MGMT in TMZ-resistant glioma cells (T98 and U138), western blot analyses were performed after the application of various concentrations of VPA (0–8 mM). The expression of MGMT in T98 and U138 cells was decreased by VPA in a dose-dependent manner (Figure 3(b)). We also evaluated the combination effect under the concentrations of VPA (0–8 mM) and TMZ (50 μM). The cell viability was decreased by VPA in a dose-dependent manner (Figure 3(c)). Moreover, cell death induced by combination can be significantly alleviated by caspase inhibitor Z-VAD-fmk (20 μM) and autophagy inhibitor 3-MA (2 mM) (P < 0.05, combination treatment versus inhibitor treatment). Taken together, these results suggest that VPA downregulates MGMT and is associated closely with TMZ sensitivity in TMZ-resistant glioma cells.
Figure 3.
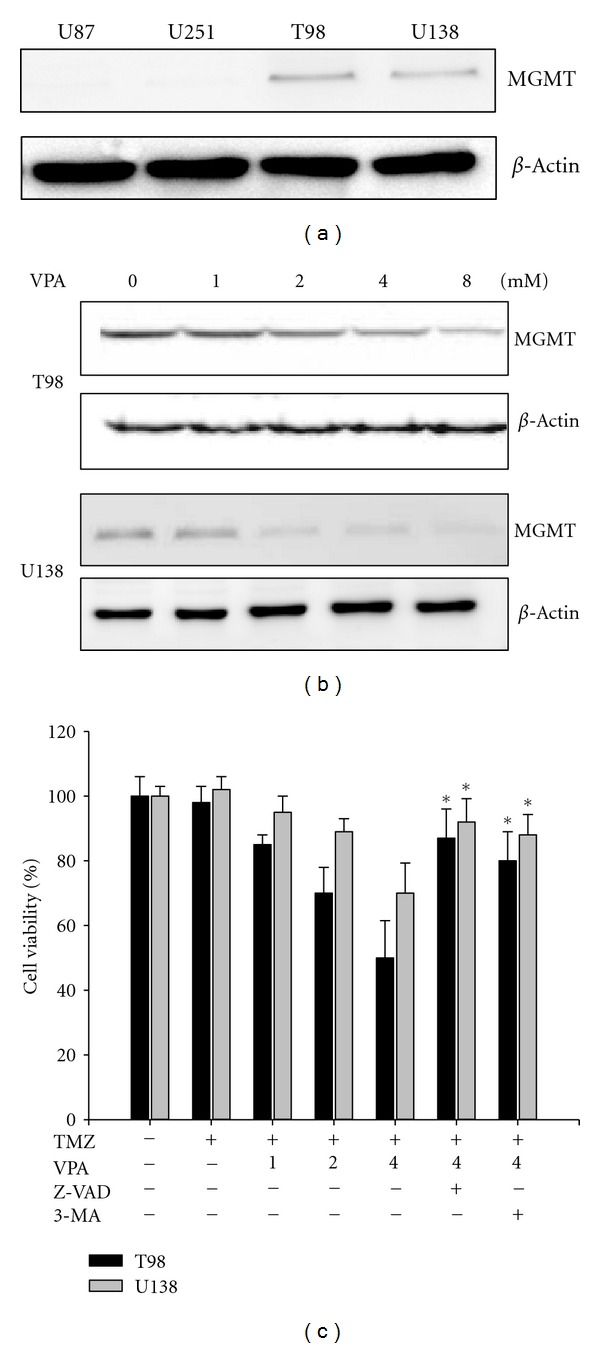
Effect of VPA on the expression of MGMT in TMZ-resistant glioma cells. (a) Cell lysates of four human glioma cell lines were subjected to western blotting using an anti-MGMT antibody. This revealed an absence of MGMT expression in the U87 and U251 cell lines. In contrast, the other two cell lines, T98 and U138, exhibited MGMT expression at the protein level. (b) MGMT expression in the T98 and U138 TMZ-resistant cells treated with varying concentrations of VPA (0–8 mM). β-Actin was used as a loading control. (c) Cells were exposed to medium containing various concentrations of VPA (0–4 mM) with 50 μM TMZ for 72 h and were measured using MTT assay. Columns, mean; bars, SE. *P < 0.05, Student's t-test. The results are representative of three independent experiments.
3.4. Effects of Apoptotic and Autophagic Cell Death by VPA and TMZ Combination in TMZ-Resistant Glioma Cells
To identify the synergistic effects of VPA on TMZ-induced apoptosis, we performed western blot analyses using antibodies against Bcl-2, Bak, and caspase-3 in T98 and U138 cells treated with 4 mM VPA and/or 50 μM TMZ. The expression of the proapoptotic molecule Bak was increased by VPA alone and, to a much greater extent, by the VPA and TMZ combination. Cleaved caspase-3, which is a terminal effector of apoptosis, was also increased by VPA and also to a much greater extent by the VPA and TMZ combination. However, the expression of the antiapoptotic molecule Bcl-2 was decreased by VPA alone and by the VPA and TMZ combination. Similar patterns were observed for U138 cells (Figure 4(a)). In addition, we examined the expression of two microtubule-associated protein light-chain-3 (LC3) forms, LC3-I and LC3-II, using western blot analysis, since LC3-II correlates with the extent of autophagosome formation [16]. Expression of LC3-II increased in TMZ-resistant glioma cells treated with VPA and TMZ combination. These findings suggest that the combination of VPA and TMZ enhances TMZ-induced apoptotic and autophagic cell death in TMZ-resistant glioma cells.
Figure 4.

Induction of apoptotic and autophagic cell death in TMZ-resistant glioma cells by VPA and TMZ combination. T98 and U138 cells were untreated or treated with VPA, TMZ, or their combination. (a) The expression of MGMT, Bcl-2, Bak, and caspase-3 was evaluated using western blotting. (b) Conversion of LC3-I to LC3-II was also determined by western blotting. β-actin was used as a loading control. The results are representative of three independent experiments.
3.5. Suppression of TMZ-Resistant Glioma Cell Migration by VPA and TMZ Combination
It has been reported that TMZ modifies caveolin-1 expression in vitro and in vivo in glioma cells, a feature that directly affects glioma cell migration properties. TMZ decreases β1 integrin expression and modulates Cyr61 signaling and actin cytoskeleton polymerization in GBM cell lines [17]. We investigated the effect of VPA and TMZ on TMZ-resistant glioma cell migration using a transwell migration assay. VPA and TMZ separately did not affect cell migration, but the combination of VPA and TMZ was significantly suppressed the migration of T98 cells (P = 0.021, combination treatment versus single treatment) and U138 cells (P > 0.05) (Figure 5). This result suggests that VPA sensitizes the antimigratory activity of TMZ in TMZ-resistant glioma cells.
Figure 5.

Antimigration effect of a combination of VPA and TMZ in TMZ-resistant glioma cells. Transwell migration assay demonstrates the migration of TMZ-resistant glioma cells in response to 20% FBS. (a) Representative photomicrographs of stained filters showing migrating T98 and U138 (data not shown). Magnification, ×100. (b) T98 and U138 cells were pretreated with VPA (4 mM) or TMZ (50 μM), alone or in combination, for 72 h. Serum-induced T98 cell migration was significantly inhibited by VPA and TMZ combination. Serum-free medium (SFM) was used as a experimental control. Columns, mean; bars, SE. *P < 0.05, ANOVA. The results are representative of three independent experiments.
3.6. Reduction of TMZ-Resistant Glioma Xenograft Growth after Treatment with a Combination of VPA and TMZ
We also investigated the synergistic effects of a combination of VPA and TMZ in a human glioma xenograft-nude mouse model. TMZ (50 mg/kg) alone did not decrease the growth of the T98 tumor (P > 0.05). VPA (300 mg/kg) alone suppressed T98 tumor growth slightly (P > 0.05). In contrast, a combination of VPA and TMZ decreased T98 tumor growth to a very significant extent (P = 0.003, combination treatment versus single treatment). Therefore, the combination treatment decreased tumor growth significantly in the TMZ-resistant glioma xenografts compared with the monotherapies (Figure 6).
Figure 6.
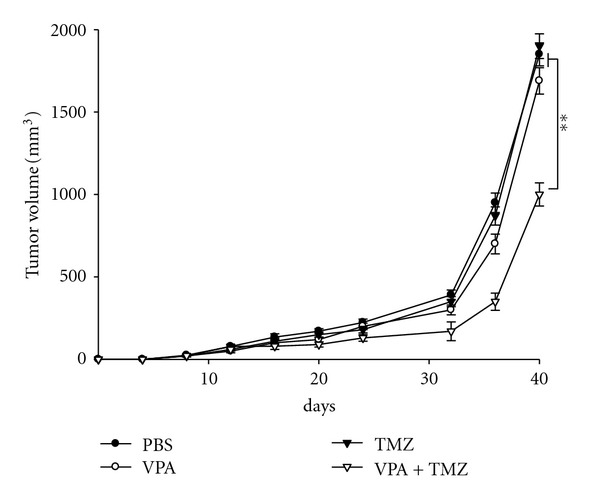
Inhibition of the growth of T98 xenografts treated with VPA and TMZ. BALB/c nude mice bearing T98 cells were distributed into four treatment groups: filled circle, PSB; open circle, VPA (300 mg/kg); filled triangle, TMZ (50 mg/kg); open triangle, VPA (300 mg/kg) + TMZ (50 mg/kg). Points, mean; bars, SE. **P < 0.01, ANOVA. PBS was used as a control. The results are representative of two independent experiments.
4. Discussion
Despite the multimodality treatments available, including surgery, radiation therapy, and chemotherapy, the prognosis of malignant gliomas remains poor. TMZ is considered as a standard chemotherapeutic regimen for the first-line treatment of malignant gliomas. However, TMZ does not exhibit uniform sensitivity to malignant gliomas. The resistance of some malignant gliomas to TMZ is related to the DNA repair enzyme MGMT, via the removal of the alkylating lesion at the O6 position of guanine. This causes the replication of DNA and the growth of malignant gliomas [3–6]. In the present study, we demonstrated that malignant glioma cell lines exhibit differential expression of the MGMT gene, which is in keeping with the results of a previous study [18, 19]. MGMT expression is suppressed at the level of transcription by CpG methylation within the MGMT promoter, and almost half of primary GBM tumor samples have evidence of promoter hypermethylation, as determined by methylation-specific PCR (MS-PCR) [20, 21]. Furthermore, tumor MGMT promoter hypermethylation was associated with prolonged survival (46% 2-year survival) in GBM patients treated with TMZ and radiation as compared to those patients without evidence of tumor hypermethylation (14% 2-year survival) [22]. In this study, the MGMT status, including promoter hypermethylation (data not shown), quantitative mRNA expression (data not shown), and protein expression in glioma cells, also confirmed that MGMT promoter methylation suppresses gene transcription and reduces MGMT protein expression. Therefore, suppression of MGMT expression mediated by promoter hypermethylation may be an important factor influencing TMZ sensitivity.
Recently, VPA has emerged as an anticancer drug because of its function as an inhibitor of HDACs. Several previous reports indicate that VPA induces cancer cell apoptosis and anticancer immunity. It also inhibits tumor-associated inflammation and angiogenesis, invasion, and proliferation of cancer cells [23, 24]. Here, VPA had an anticancer effect on human malignant glioma cell lines. However, the sensitivity to VPA varied among glioma cell lines. VPA had an anticancer effect in U87 and U251 cells at low dosages of the drug; however, T98 and U138 cells were similarly affected only at a high dosage of VPA (Figure 1). This phenomenon may reflect a difference in HDAC-induced histone hyperacetylation or posttranslational protein acetylation among the glioma cell lines. Numerous in vitro studies use concentrations of VPA in a higher dosage range, mostly from 1 up to 10 mM [25–27]. Furthermore, the sensitivity to VPA was different among glioma cell lines. Therefore, a high dosage of VPA (4 mM) was chosen for the following in vitro combination experiments. However, further work needs to be done in relation to the safety and toxicity of VPA and TMZ at optimal therapeutic concentrations.
Several preclinical and clinical studies have proved that hypomethylating drugs, such as 5-aza-2′-deoxycytidine (5-aza-CdR; decitabine), and HDACI, such as valproic acid (VPA; Depakine) and suberoylanilide hydroxamic acid (SAHA; vorinostat), have potent anticancer activity and promising therapeutic potential [13, 28–30]. Previous experimental results indicate that the anticancer effects of VPA can be increased by its combination with other drugs [31, 32]. Among them, a limited number of studies addressed the combination with VPA and other anticancer drugs for the treatment of glioma. An in vitro study of the synergistic effect of VPA and etoposide on U87, U251, and LN18 cells demonstrated that VPA sensitizes glioma cell lines to the cytotoxic effects of etoposide by inducing differentiation and upregulating the expression of p21/WAF1 and both isoforms of topoisomerase II [33]. Another study reported that the synergistic effect of the association of VPA with nitrosourea (BCNU) on human glioma cell lines in vitro was related to an increased blockage of the cell cycle with accumulation in the S-G2/M phases, rather than an increase in programmed cell death [34]. The present investigation of the effect of VPA/TMZ revealed that U87 and U251 cells were sensitive to VPA or TMZ independently. Therefore, they did not show an enhanced antitumor effect on the combination treatment. However, an enhanced antitumor effect was found in TMZ-resistant T98 and U138 cells (Figure 2). This phenomenon can be explained by a VPA-mediated sensitization of T98 and U138 cells to TMZ. The evaluation of MGMT status in T98 and U138 cells after VPA treatment demonstrated that MGMT expression was decreased by VPA in a dose-dependent manner. This result can be explained by the HDACI function of VPA, which remodels chromatin [35]. Downregulation of the MGMT protein may not remove alkylating lesions at the O6 position of guanine and increase the sensitivity to TMZ in VPA-treated TMZ-resistant cells.
5. Conclusions
This study demonstrated that the combination treatment of VPA/TMZ regulates TMZ sensitivity via the downregulation of MGMT and has anticancer effects in vitro/in vivo in TMZ-resistant glioma cells. Although further studies are needed to determine the exact mechanism underlying the inhibition of MGMT expression, these results suggest that the clinical therapeutic efficacy of TMZ in malignant glioma can be enhanced by combination with VPA.
Author's Contribution
C. H. Ryu and W. S. Yoon contributed equally to this work.
Acknowledgments
This study was supported by grants of the Korea Health technology R&D Project (A110330, A110298) and by a grant from the National R&D Program for Cancer Control (0820040), Ministry of Health & Welfare, Republic of Korea.
References
- 1.Kurzen H, Schmitt S, Näher H, Möhler T. Inhibition of angiogenesis by non-toxic doses of temozolomide. Anti-Cancer Drugs. 2003;14(7):515–522. doi: 10.1097/00001813-200308000-00003. [DOI] [PubMed] [Google Scholar]
- 2.Stupp R, Mason WP, van den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. The New England Journal of Medicine. 2005;352(10):987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 3.Friedman HS, Kerby T, Calvert H. Temozolomide and treatment of malignant glioma. Clinical Cancer Research. 2000;6(7):2585–2597. [PubMed] [Google Scholar]
- 4.Silber JR, Blank A, Bobola MS, Ghatan S, Kolstoe DD, Berger MS. O6-Methylguanine-DNA methyltransferase-deficient phenotype in human gliomas: frequency and time to tumor progression after alkylating agent-based chemotherapy. Clinical Cancer Research. 1999;5(4):807–814. [PubMed] [Google Scholar]
- 5.Pegg AE. Mammalian O6-alkylguanine-DNA alkyltransferase: regulation and importance in response to alkylating carcinogenic and therapeutic agents. Cancer Research. 1990;50(19):6119–6129. [PubMed] [Google Scholar]
- 6.Bobola MS, Blank A, Berger MS, Silber JR. Contribution of O6-methylguanine-DNA methyltransferase to monofunctional alkylating-agent resistance in human brain tumor-derived cell lines. Molecular Carcinogenesis. 1995;13(2):70–80. doi: 10.1002/mc.2940130203. [DOI] [PubMed] [Google Scholar]
- 7.Göttlicher M, Minucci S, Zhu P, et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. The EMBO Journal. 2002;20(24):6969–6978. doi: 10.1093/emboj/20.24.6969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Boffa IC, Vidali G, Mann RS, Allfrey VG. Suppression of histone deacetylation in vivo and in vitro by sodium butyrate. Journal of Biological Chemistry. 1978;253(10):3364–3366. [PubMed] [Google Scholar]
- 9.Candido EPM, Reeves R, Davie JR. Sodium butyrate inhibits histone deacetylation in cultured cells. Cell. 1978;14(1):105–113. doi: 10.1016/0092-8674(78)90305-7. [DOI] [PubMed] [Google Scholar]
- 10.Knüpfer MM, Pulzer F, Schindler I, Hernáiz Driever P, Knüpfer H, Keller E. Different effects of valproic acid on proliferation and migration of malignant glioma cells in vitro. Anticancer Research. 2001;21(1A):347–352. [PubMed] [Google Scholar]
- 11.Shao Y, Gao Z, Marks PA, Jiang X. Apoptotic and autophagic cell death induced by histone deacetylase inhibitors. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(52):18030–18035. doi: 10.1073/pnas.0408345102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kuendgen A, Gattermann N. Valproic acid for the treatment of myeloid malignancies. Cancer. 2007;110(5):943–954. doi: 10.1002/cncr.22891. [DOI] [PubMed] [Google Scholar]
- 13.Duenas-Gonzalez A, Candelaria M, Perez-Plascencia C, Perez-Cardenas E, de la Cruz-Hernandez E, Herrera LA. Valproic acid as epigenetic cancer drug: preclinical, clinical and transcriptional effects on solid tumors. Cancer Treatment Reviews. 2008;34(3):206–222. doi: 10.1016/j.ctrv.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 14.Chen CH, Chang YJ, Ku MSB, Chung KT, Yang JT. Enhancement of temozolomide-induced apoptosis by valproic acid in human glioma cell lines through redox regulation. Journal of Molecular Medicine. 2011;89(3):303–315. doi: 10.1007/s00109-010-0707-1. [DOI] [PubMed] [Google Scholar]
- 15.Carmichael J, DeGraff WG, Gazdar AF. Evaluation of a tetrazolium-based semiautomated colorimetric assay: assessment of chemosensitivity testing. Cancer Research. 1987;47(4):936–942. [PubMed] [Google Scholar]
- 16.Kabeya Y, Mizushima N, Ueno T, et al. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. The EMBO Journal. 2000;19(21):5720–5728. doi: 10.1093/emboj/19.21.5720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bruyère C, Abeloos L, Lamoral-Theys D, et al. Temozolomide modifies caveolin-1 expression in experimental malignant gliomas in vitro and in vivo. Translational Oncology. 2011;4(2):92–100. doi: 10.1593/tlo.10205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yoshino A, Ogino A, Yachi K, et al. Effect of IFN-β on human glioma cell lines with temozolomide resistance. International Journal of Oncology. 2009;35(1):139–148. doi: 10.3892/ijo_00000322. [DOI] [PubMed] [Google Scholar]
- 19.Yoshino A, Ogino A, Yachi K, et al. Gene expression profiling predicts response to temozolomide in malignant gliomas. International Journal of Oncology. 2010;36(6):1367–1377. doi: 10.3892/ijo_00000621. [DOI] [PubMed] [Google Scholar]
- 20.Paz MF, Yaya-Tur R, Rojas-Marcos I, et al. CpG island hypermethylation of the DNA repair enzyme methyltransferase predicts response to temozolomide in primary gliomas. Clinical Cancer Research. 2004;10(15):4933–4938. doi: 10.1158/1078-0432.CCR-04-0392. [DOI] [PubMed] [Google Scholar]
- 21.Esteller M, Hamilton SR, Burger PC, Baylin SB, Herman JG. Inactivation of the DNA repair gene O(6)-methylguanine-DNA methyltransferase by promoter hypermethylation is a common event in primary human neoplasia. Cancer Research. 1999;59(4):793–797. [PubMed] [Google Scholar]
- 22.Hegi ME, Diserens AC, Gorlia T, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. The New England Journal of Medicine. 2005;352(10):997–1003. doi: 10.1056/NEJMoa043331. [DOI] [PubMed] [Google Scholar]
- 23.Blaheta RA, Michaelis M, Driever PH, Cinatl J., Jr. Evolving anticancer drug valproic acid: insights into the mechanism and clinical studies. Medicinal Research Reviews. 2005;25(4):383–397. doi: 10.1002/med.20027. [DOI] [PubMed] [Google Scholar]
- 24.Michaelis M, Doerr HW, Cinatl J., Jr. Valproic acid as anti-cancer drug. Current Pharmaceutical Design. 2007;13(33):3378–3393. [PubMed] [Google Scholar]
- 25.Greenblatt DY, Vaccaro AM, Jaskula-Sztul R, et al. Valproic acid activates Notch-1 signaling and regulates the neuroendocrine phenotype in carcinoid cancer cells. Oncologist. 2007;12(8):942–951. doi: 10.1634/theoncologist.12-8-942. [DOI] [PubMed] [Google Scholar]
- 26.Rodriguez-Menendez V, Gilardini A, Bossi M, et al. Valproate protective effects on cisplatin-induced peripheral neuropathy: an in vitro and in vivo study. Anticancer Research. 2008;28(1A):335–342. [PubMed] [Google Scholar]
- 27.van Nifterik KA, van den Berg J, Slotman BJ, Lafleur MV, Sminia P, Stalpers LJ. Valproic acid sensitizes human glioma cells for temozolomide and γ-radiation. Journal of Neuro-Oncology. 2012;107(1):61–67. doi: 10.1007/s11060-011-0725-z. [DOI] [PubMed] [Google Scholar]
- 28.Garcia-Manero G, Yang H, Bueso-Ramos C, et al. Phase 1 study of the histone deacetylase inhibitor vorinostat (suberoylanilide hydroxamic acid [SAHA]) in patients with advanced leukemias and myelodysplastic syndromes. Blood. 2008;111(3):1060–1066. doi: 10.1182/blood-2007-06-098061. [DOI] [PubMed] [Google Scholar]
- 29.Eyüpoglu IY, Hahnen E, Buslei R, et al. Suberoylanilide hydroxamic acid (SAHA) has potent anti-glioma properties in vitro, ex vivo and in vivo. Journal of Neurochemistry. 2005;93(4):992–999. doi: 10.1111/j.1471-4159.2005.03098.x. [DOI] [PubMed] [Google Scholar]
- 30.Marks PA, Rifkind RA, Richon VM, Breslow R, Miller T, Kelly WK. Histone deacetylases and cancer: causes and therapies. Nature Reviews Cancer. 2001;1(3):194–202. doi: 10.1038/35106079. [DOI] [PubMed] [Google Scholar]
- 31.Blaheta RA, Nau H, Michaelis M, Cinatl J., Jr. Valproate and valproate-analogues: potent tools to fight against cancer. Current Medicinal Chemistry. 2002;9(15):1417–1433. doi: 10.2174/0929867023369763. [DOI] [PubMed] [Google Scholar]
- 32.Cinatl J, Jr., Kotchetkov R, Blaheta R, Driever PH, Vogel JU, Cinatl J. Induction of differentiation and suppression of malignant phenotype of human neuroblastoma BE(2)-C cells by valproic acid: enhancement by combination with interferon-alpha. International Journal of Oncology. 2002;20(1):97–106. [PubMed] [Google Scholar]
- 33.Das CM, Aguilera D, Vasquez H, et al. Valproic acid induces p21 and topoisomerase-II (α/β) expression and synergistically enhances etoposide cytotoxicity in human glioblastoma cell lines. Journal of Neuro-Oncology. 2007;85(2):159–170. doi: 10.1007/s11060-007-9402-7. [DOI] [PubMed] [Google Scholar]
- 34.Ciusani E, Balzarotti M, Calatozzolo C, et al. Valproic acid increases the in vitro effects of nitrosureas on human glioma cell lines. Oncology Research. 2007;16(10):453–463. doi: 10.3727/096504007783338340. [DOI] [PubMed] [Google Scholar]
- 35.Mueller WC, von Deimling A. Gene regulation by methylation. Recent Results in Cancer Research. 2009;171:217–239. doi: 10.1007/978-3-540-31206-2_13. [DOI] [PubMed] [Google Scholar]