

Abstract
The generation of efficient candidate vaccines against bovine tuberculosis will contribute to the control of this zoonotic disease. Rationally attenuated Mycobacterium bovis strains generated by knockout of virulence genes are promising candidate vaccines. However, to be effective, these candidate vaccines should at least maintain the immunological properties of their virulent parental M. bovis strains. Therefore, the aim of this study was to obtain an M. bovis strain deleted in the mce2 genes and evaluate the effect of the mutation on the immunological profile elicited by the bacteria in cattle. We showed that the activation of CD4+ T cells in cattle inoculated with the mutant strain was equivalent to that in animals inoculated with the parental strain. Moreover, after in vitro stimulation, peripheral blood mononuclear cells from animals inoculated with the mutant produced higher levels of mRNA Th-1 cytokines than the parental strain. Therefore, these results indicate that the mce2 mutant is a promising candidate vaccine against bovine tuberculosis.
1. Introduction
Bovine tuberculosis (bTB) is not only serious animal and zoonotic disease that causes significant financial loss but also a public health hazard. While the main host of Mycobacterium bovis, the causative agent of bTB, is cattle, other animals, including humans, may also be affected. Infection in humans occurs when unpasteurized milk (or derivatives) is consumed or when people are in contact with infected cattle. bTB is a factor that undermines the development of the dairy and meat industry and international commerce. Therefore, it is essential to control and eradicate this disease and an efficient vaccination strategy would help in the control of bovine tuberculosis. Vaccination of cattle to control bTB is particularly demanding in high-prevalence zones where it is economically unfeasible to slaughter animals.
Rationally attenuated, live replicating mutants of M. bovis are potential vaccine candidates. The advantage of using attenuated M. bovis strains is that they produce a large number of protective antigens, including those absent from BCG [1]. Thus, vaccination with live attenuated M. bovis can induce a stronger and persistent immune stimulation, conferring higher levels of protection against tuberculosis than BCG.
The M. bovis genome contains three operons designated mce (mammalian cell entry). These operons, which encode membrane and exported proteins, are highly conserved in pathogenic and nonpathogenic mycobacteria [2]. In a previous study, we have demonstrated that the mce2 operon is essential for the survival of Mycobacterium tuberculosis, the causative agent of human tuberculosis, during infection in mice [3]. In that study, we found that M. tuberculosis knockout in the mce2 operon replicated less than its parental strain in mouse organs after intratracheal inoculation of animals. Our findings were consistent with previous observations supporting the involvement of mce operons in the host-pathogen interaction [2, 4–9]. In particular, in a recent study Marjanovic et al. [10] demonstrated that mice infected with an M. tuberculosis mutant in the mce2 operon survive longer than those infected with its parental and virulent strain, although the replication of both strains in organs was equivalent. The use of the M. tuberculosis Δmce2 strain as a vaccine confers better protection than BCG in both mice and guinea pigs challenged with a hypervirulent M. tuberculosis strain [11, 12]. M. bovis is closely related to M. tuberculosis, and both species are included in the M. tuberculosis complex. In addition, as both organisms can cause identical clinical disease in humans and are genetically extremely similar, it is likely that many of the virulence factors of M. bovis are the same as those of M. tuberculosis [13]. Based on these facts, we propose an M. bovis knockout strain in the mce2 operon as a candidate vaccine against bTB. In order to determine the immunogenic properties of this candidate, in this study we characterized the immunological profile elicited in cattle by the M. bovis mce2 mutant strain towards M. bovis antigens and compared it with that elicited by its parental and virulent strain NCTC 10772 [14]. To this end, we measured the cytokine mRNA expression in peripheral blood mononuclear cells (PBMCs) and determined the lymphocyte subsets involved in recalling activation of PBMCs from cattle inoculated with the candidate vaccine in response to M. bovis antigens. We found that inoculation of cattle with the candidate vaccine stimulates both CD4+ and CD8+ T-cell responses to produce Th1-associated cytokines.
2. Results
2.1. Construction of mce2 Mutant of M. bovis
By using the gene knock-out system described by Parish and Stoker [15], we created a mutant strain of M. bovis carrying a unmarked-chromosomal deletion of the region spanning the mce2A-mce2B genes (Figure 1(a)). The deletion was confirmed by PCR using primers that hybridize outside the deleted region. As shown in Figure 1(b), an amplicon of 1,831 bp corresponding to the mce2A and mce2B genes and adjacent regions was obtained in the wild type strain, while in the mutant, the amplified DNA fragment was of 177 bp and corresponded to the Δmce2A-mce2B locus. Primers that hybridize in sigA amplified the expected fragment in both strains, indicating the integrity of the DNA samples used in the PCR reactions. The mutant strain was designated MbΔmce2.
Figure 1.
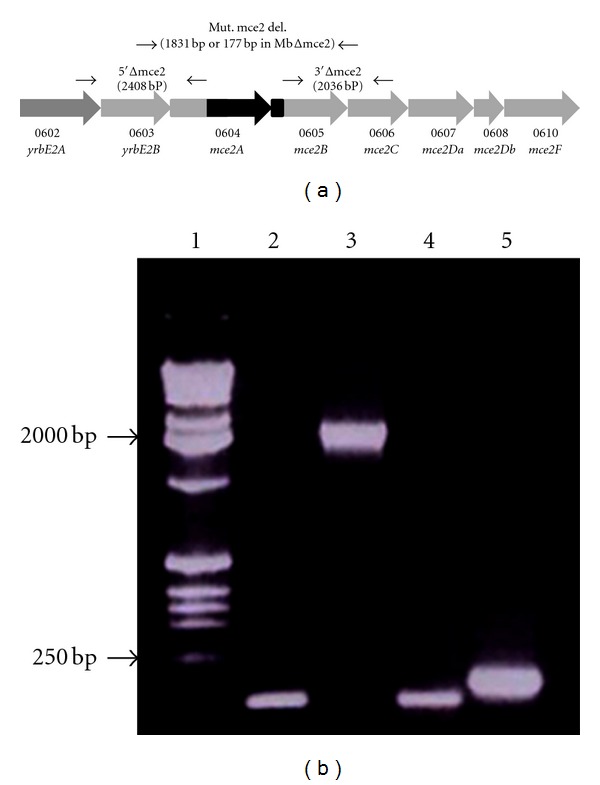
Construction of the M. bovis Δmce2 mutant strain. (a) An unmarked mutant strain of M. bovis carrying a chromosomal deletion of the region spanning the mce2A-mce2B genes was created by the gene knock-out system described by Parish and Stoker [15]. The mutant strain was designated MbΔmce2. (b) PCR analysis of MbΔmce2 and the wild type strains. PCRs were performed by using primers that hybridize outside the mce2A-mce2B region (lanes 3 and 5) or in sigA (lanes 2 and 4). Chromosomal DNA from the parental strain (lanes 2 and 3) or from the mutant strain (lanes 4 and 5) was used as template in the PCR reactions. Lane 1 is the molecular weight marker (1Kb ladder Promega).
2.2. Activation of CD4+, and CD8+ in PPDB-Stimulated PBMCs from Cattle Inoculated with the Candidate Vaccine Δmce2
To evaluate the recall response to purified protein derivative (PPDB) of lymphocyte subsets in animals inoculated with either the candidate MbΔmce2 or its wild type parental strain (NCTC 10772), we used a flow cytometry-based proliferation assay. In PBMCs isolated 15 and 90 days after infection (dpi), activation of CD4+ and CD8+ significantly increased upon stimulation with PPDB (Figures 2(a) and 2(b)). After specific stimulation, the expression of IL-2R in CD4+ cells, which indicates lymphocyte activation, increased along the time of infection (P < 0.01) with similar rates in both groups (Figure 3(a)). The expression of IL-2R in CD8+ cells also showed the same trend, but it was only significant in animals inoculated with the wild type strain (P < 0.05) (Figure 3(b)). Importantly, in PBMCs from animals before the infection, the percentages of CD4+ and CD8+ cells were not significantly altered upon PPDB stimulation, indicating that the animals used in this study were neither previously sensitized nor infected with M. bovis.
Figure 2.
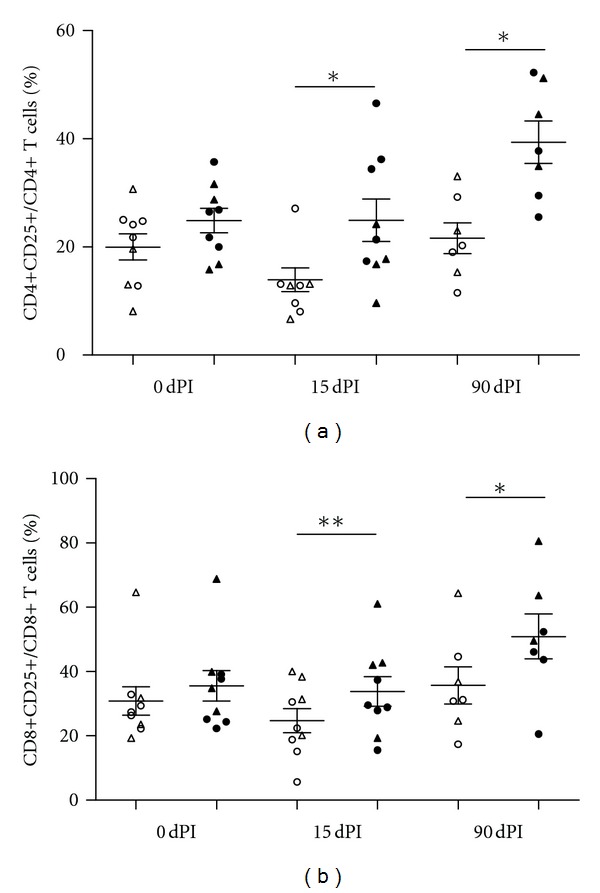
Percentages of lymphocyte cell subsets CD4+ (a) and CD8+ (b) expressing CD25 of PBMCs from animals inoculated with either MbΔmce2 (n = 4, triangle) or NCTC 10772 (n = 3, circle) stimulated with PPDB (black) or PBS (white) at different time points. Data were analyzed using the Wilcoxon matched pair test for cells with and without PPDB stimulation (*statistically significant P < 0.05 and **statistically significant P < 0.01). The means ± SEM are indicated.
Figure 3.
Percentages of the activated lymphocyte cell subsets CD4+ (a) or CD8+ (b) of PBMCs stimulated with PPDB from animals inoculated with MbΔmce2 (white square) or NCTC 10772 (black triangle). The means ± SEM are indicated. *statistically significant P < 0.05.
2.3. Expression of Cytokines in PBMCs from Cattle Inoculated with the Candidate Vaccine MbΔmce2
In order to evaluate Th1 and Th2 immune responses in cattle inoculated with the candidate vaccine strain MbΔmce2, we assessed the cytokine expression profile in PBMCs by measuring cytokine mRNAs after stimulating the cells with PPDB (Figure 4). Values for sequential samples were normalized to values before inoculation for each animal.
Figure 4.
Relative cytokine gene expression. Gene expression was measured in PBMCs from animals infected with either MbΔmce2 (n = 4, white bars) or NCTC 10772 (n = 3, gray bars) stimulated with PPDB at 15 and 90 dpi. Relative gene expression was calculated using the 2-ΔΔCt method with E correction, using pol II and gadph mRNA expression as reference genes and the preimmune condition as the calibrator. Data were analyzed using a two-tailed unpaired t-test (**P < 0.01, ***P < 0.001). The bars indicate the average ratios of infected animals/uninfected animals ± SEM.
PBMCs obtained from cattle inoculated with MbΔmce2 at 15 dpi responded to PPDB stimulation by expressing 16.2-fold more IFN-γ mRNA than cells from animals before inoculation. In contrast, a lower level of IFN-γ was detected in animals inoculated with the wild type strain (P < 0.05). However, at 90 dpi, the expression of IFN-γ increased in cattle inoculated with the wild type, while it showed only a slight reduction in those inoculated with the mutant. The expression of interleukin-2 (IL-2) in PBMCs from both animal groups was significantly different from that in PBMCs from animals before the inoculation. However, the expression level of this cytokine was equivalent in both groups and maintained during the time points assayed. Only the group inoculated with the mutant responded to PPDB stimulation with production of IL-12 and TNF-α (P < 0.0001).
IL-10 and IL-4 gene expression was evaluated to assess Th2 responses. Stimulation with PPDB induced greater IL-10 gene expression in animals inoculated with the mutant than in those inoculated with the wild type strain at both time points assayed (P < 0.005). At 15 dpi, the expression of IL-4 was similar in both groups, while at 90 dpi, the level of IL-4 was higher in the animals inoculated with the wild type strain (P < 0.005).
3. Discussion
The efficacy of BCG to protect against tuberculosis in humans has shown to be highly variable and depending on uncertain factors of the populations tested. Field evaluation of the protective efficacy of BCG against bovine tuberculosis has indicated that BCG is not capable of protecting 100% of the animals [16, 17]. One possible explanation of this failure is that exposure to environmental mycobacteria compromises the protection efficacy of BCG in cattle, as it happens with the use of BCG in humans [18]. To overcome the incomplete protection afforded by BCG, numerous vaccine strategies have been developed and tested in experimental vaccine assays. Among them, prime-boost strategies combining single antigens and BCG have been shown to improve the protective efficacy of BCG significantly. Inspections on the genomic sequences of M. bovis isolates and BCG strains have revealed that at least 16 regions of differences (RDs) are absent from the BCG chromosomes [19, 20]. Many of these RDs encode for powerful and protective antigens [1, 21]. Thus, an attractive alternative to the use of BCG as a veterinary vaccine is the development of attenuated M. bovis strains deleted in proteins that are nonantigenic but relevant for virulence. Here, we describe the construction of a mutant strain of M. bovis and its immunological evaluation as a candidate attenuated live vaccine against bTB. Based on previous findings indicating that inactivation or deletion of the mce2 operon attenuates M. tuberculosis [3], we obtained an attenuated nonreverting M. bovis strain by deletion of two mce2 genes. Importantly, since the experimental approach used to generate this mutant strain avoids the insertion of any selective and nonselective gene marker into the bacterial chromosome, we created this mutant in compliance with the criteria of tuberculosis vaccine development [22]. Another important aspect of this candidate vaccine strategy is that the absence of Mce2A and Mce2B proteins in the mutant strain could be used as marker to differentiate infected animals from vaccinated ones.
Given that there is no doubt about the essential protective role of CD4+ T cells to control tuberculosis infections, it is a condition that any candidate vaccine is capable of stimulating this cell population. Therefore, the most effective vaccination strategies will be those that stimulate CD4+ T-cell responses to produce Th1-associated cytokines. In accordance with these requirements, the candidate vaccine Δmce2 elicited strong T-cell responses in cattle, with activation of CD4+ following stimulation with the M. bovis-specific antigen PPDB.
It has been demonstrated that CD8+ T cells are also required for immunity against tuberculosis in a variety of animal models as well as in humans [9, 23–27]. In this regard, in the group of animals inoculated with the mutant strain, the activation of CD8+ T cells was sustained along the time of infection and increased at the last point assayed, but this increase was not statistically significant, likely due to the reduced number of animals included in the experiment. Similarly, subcutaneous inoculation of cattle with Δmce2 induced activation of CD4+ and CD8+ following specific stimulation, but only the CD4+ T cells responded at significant levels (unpublished results).
Moreover, inoculation of the mutant strain in cattle induced the production of Th1 expression of IL-12 responses with production of IL-2, IL-12, TNF-α, and IFN-γ. Surprisingly, the level of and TNF-α was higher in stimulated PBMCs from animals inoculated with the mutant than in those from animals inoculated with the wild type strain. These data indicate that, in cattle, MbΔmce2 induces a strong Th1 response that is maintained for at least 90 days. These observations are in contrast to that found by Marjanovic et al., who have shown that an M. tuberculosis mutant in the mce2 operon induces smaller amounts of proinflammatory cytokines (TNF-α, IL-6, and MCP-1) in RAW cells than the wild type H37Rv strain. However, considerable differences between the systems used in both studies could explain these disparities. In particular, these authors used an in vitro murine model to determine the cytokines produced after cell infection, while here we measured cytokines produced after antigen stimulation of total bovine PBMCs. Although it is still unclear why the mutant strain elicited stronger proinflammatory response than the wild type strain, we speculate that the lack of Mce2 lipid transporter could alter the M. bovis cell wall composition, which, in turn, could affect the host's immune response elicited by the bacteria.
Production of the anti-inflammatory cytokine IL-10 was early detected in animals inoculated with the mutant strain. In contrast, animals inoculated with the wild type strain showed low IL-10 expression at 15 and 90 dpi. At 15 dpi, IL-4 expression levels were similar in both groups, whereas at 90 dpi, the expression of this cytokine decreased in the wild type group. Together, these results suggest that anti-inflammatory responses in animals inoculated with the mutant strain were generated to attenuate the adverse effect of an exacerbated inflammation in the host's tissues driven by the highly expressed IFN-γ.
The importance of the induction of multifunctional CD4+ T cells producing different cytokines simultaneously for protection against tuberculosis has been previously addressed in a mouse model [28]. Therefore, it will be of interest to carry out further studies to establish whether multifunctional CD4+ T cells are induced in MbΔmce2-vaccinated cattle.
Given that the immune responses between bovines and humans have been considered a determinant issue in the development of diagnostic tools and vaccines for both human and bovine tuberculosis [29], the results presented here make this mutant an attractive candidate vaccine to be tested against both bovine and human tuberculosis.
4. Conclusions
Here, we demonstrated that the lack of mce2 gene expression maintains the immunogenicity properties of M. bovis, favoring its ability to protect against bovine tuberculosis. Although the results presented here make Δmce2 an attractive candidate vaccine, further studies addressing safety and innocuity concerns are necessary before testing this candidate in challenge assays.
5. Material and Methods
5.1. Bacterial Strains and Culture Media
All cloning steps were performed in Escherichia coli Top10 (F-mcrAΔ(mrr-hsdRMS-mcrBC) φ80lacZΔM15 ΔlacX74 nupG recA1 araD139 Δ(ara-leu)7697 galE15 galK16 rpsL(StrR) endA1 λ-). E. coli were grown either in Luria-Bertani (LB) broth or on LB agar. When necessary, 50 μg/mL kanamycin was added to the LB media. M. bovis strains were grown in Middlebrook 7H9 medium supplemented with 0.05% Tween 80 or Middlebrook 7H10. Middlebrook media were supplemented with albumin 0.5%, dextrose 0.4%, and 0.4% pyruvate (ADP). To grow the knock-out M. bovis strain, Middlebrook 7H10-ADP was supplemented with either 20 μg/mL kanamycin plus 50 μg/mL X-Gal or 2% sucrose.
5.2. General DNA Methodology
PCR amplifications from genomic DNA templates were performed as previously described [30]. Each primer contained base mismatches that introduced a restriction site suitable for directional cloning. Chromosomal DNA samples were obtained as described by [31]. Purification of plasmids and DNA fragments was performed using the GFX Micro Plasmid Prep Kit (GE Healthcare) and DNA and Gel Band Purification Kit (GE Healthcare), respectively, according to the manufacturer's instructions.
5.3. Construction of the M. bovis Δmce2 Mutant Strain
Genomic regions of about 2 kb either upstream of mce2A or downstream of mce2B were obtained by PCR from M. bovis total DNA by using the following pair of primers: 3′Δmce2 and 5′Δmce2 (Table 1). The amplified fragments were cloned into the p2NIL plasmid [15]. The final delivery vector was generated by incorporation of the PacI cassette from pGOAL 17 into this last p2NIL recombinant vector. The vector generated was pretreated with UV light (100 mJ × cm−2) to induce depurination and promote recombination [22]. This UV-treated plasmid was used to electroporate the collection strain M. bovis NCTC 10772. The unmarked mutant was obtained using a two-step strategy described previously [15], and mutant clones were identified by colony PCR, using the primers described in Table 1. The mutant strain resulting from allelic exchange was designated MbΔmce2.
Table 1.
Primers used in this study.
| Primers | Sequences | Tm (°C) | Application |
|---|---|---|---|
| 3′ Δmce2 | F: GGATCCAAACGTTGGACCCAGACAAG | MbΔmce2 | |
| R: GCGGCCGCGGTGCTCATTGGTTGCTACC | |||
| 5′ Δmce2 | F: AAGCTTTGGCGCTGCCGGTGACT | MbΔmce2 | |
| R: GGATCCCGCGGCTACACCGTGAGATT | |||
| Mut. mce2 del | F: ACCCGAACTTCAATCTCACG | MbΔmce2 | |
| R: ACCTCACCTATCGCATGGTC | |||
| Pol II | F: GGACCCGTGTGGACAAGAAT | 76 | qPCR |
| R: ACAAGCCCCAGGTAATCATCC | |||
| GAPDH | F: ATCTCTGCACCTTCTGCCGA | 79 | qPCR |
| R: GCAGGAGGCATTGCTGACA | |||
| IL-2 | F: CGTGCCCAAGGTTAACGCTA | 76 | qPCR |
| R: CCATTGAATCCTTGATCTCTCTGG | |||
| IL-4 | F: TGCCCCAAAGAACACAACTG | 78 | qPCR |
| R: GCACGTGTGGCTCCTGTAGAT | |||
| IL-10 | F: GGAAGAGGTGATGCCACAGG | 84 | qPCR |
| R: AGGGCAGAAAGCGATGACAG | |||
| IL-12p35 | F: TAGCCACGAATGAGAGTTGCC | 78 | qPCR |
| R: TTTCCAGAAGCCAGACAATGC | |||
| TNF-α | F: CCCCCAGAGGGAAGAGTCC | 84 | qPCR |
| R: GGGCTACCGGCTTGTTACTTG | |||
| IFN-γ | F: AGCTGATTCAAATTCCGGTGG | 78 | qPCR |
| R: GATTTTGGCGACAGGTCATTC |
5.4. Cattle Infections
Holstein-Fresian calves (six months old) were inoculated intratracheally as described previously [32] with 106-107 colony forming units (CFU) of the mutant MbΔmce2 (four animals) or its parental virulent strain M. bovis NCTC 10772 (three animals). All experiments conformed to local and national guidelines on the use of experimental animals and category III infectious organisms. All the animals used in this study were negative for IFN-γ by ELISA assay (Bovigam) and tuberculin skin test at the beginning of the experiments but positive by both assays at the end of the experiments (data not shown). Blood samples were taken both at the beginning of the experiment for evaluation of preimmune status and at 15 and 90 dpi.
Heparinized blood (10 mL) from each animal was used for PBMC isolation by gradient centrifugation over Histopaque 1077 (Sigma Aldrich) following the manufacturer's protocol. PBMCs were incubated at 37°C in RPMI complete medium supplemented with 10% of bovine fetal serum (Internegocios) and 20 μg/mL final concentration of PPDB (Biocor) on 12-well tissue culture plates for 16 h for RNA extraction and 48 h for flow cytometry determinations.
After three months of infection, the calves were euthanized and then thin slices of lungs and lymph nodes of the head and pulmonary region were analyzed looking for granuloma formations. Only one of the animals inoculated with the wild type NCTC 10772 strain developed macroscopic lesions compatible with tuberculosis (data not shown). These lesions were located in retropharyngeal lymph nodes. None of the animals inoculated with the mutant MbΔmce2 showed lesions.
5.5. Flow Cytometry
For flow cytometry determinations, 2 × 106 cells were incubated either with or without PPDB. To evaluate the expression of CD4 (MCA 1653A647, IgG2a), CD8 (MCA837PE, IgG2a), and CD25 (MCA2430F and MCA2430PE) surface markers, cells were stained with fluorescent-conjugated monoclonal antibodies (AdDSerotec, Oxford, UK).
Stained cells were analyzed in a FACScalibur cytometer (BD, Franklin Lakes, NJ, USA) using Cell Quest software. Analysis gates were set on lymphocytes according to forward and side scatter. Expression of IL-2R was analyzed in CD4+ and CD8+ populations. Percentages of IL-2R-expressing cells were calculated as the ratio of CD4+ or CD8+ cells expressing CD25 and total CD4+ or CD8+ cells.
5.6. Statistical Analysis
The data were organized in two sets, one for CD4+ cells and the other for CD8+ cells. Each data set recorded CDC25 expression in cells infected with two different M. bovis strains over time. Both sets were analyzed independently as repeated measure designs over time. The dependent variable was the expression of CDC25 in PPDB stimulated cells, the bacterial strain was considered a fixed factor with two levels (MbΔmce2 and NCTC 10772), the level of CDC25 in nonstimulated cells was included in the models as a covariate, and time of sampling was treated as a random factor. The analyses were performed using the nlme library of R [33, 34].
5.7. Cytokine mRNA Analysis
Total RNA was extracted using a commercial kit (QIAGEN). The quality and quantity of RNA and the synthesis of cDNA were assayed as described previously [35].
The mRNAof cytokines (IL-2, IL-4, IL-10, IL-12p35, TNF-α, and IFN-γ) was quantified by qPCR by using specific primers (Table 1). qPCR results were analyzed using REST software as described previously [10]. For each animal, the preimmune condition was used as the calibrator, and pol II and gadph were used as reference genes. Data were analyzed using a two-tailed unpaired t-test.
Acknowledgments
The present study was supported by SECyT Grant PICT2324, MinCyT UE Mercosur Grant BiotechSur, and INTA Grant AEBIO243512 and partially supported by NIH 1R01AI083084-01. F. Bigi and A. A. Cataldi are CONICET fellows. The authors thank Valeria Rocha, Javier Leiva, and Diego Soreira for technical assistance and Luis Fernandez for the bibliography provided.
References
- 1.Al-Attiyah R, Mustafa AS. Characterization of human cellular immune responses to novel Mycobacterium tuberculosis antigens encoded by genomic regions absent in Mycobacterium bovis BCG. Infection and Immunity. 2008;76(9):4190–4198. doi: 10.1128/IAI.00199-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Casali N, Riley LW. A phylogenomic analysis of the Actinomycetales mce operons. BMC Genomics. 2007;8, article 60 doi: 10.1186/1471-2164-8-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gioffré A, Infante E, Aguilar D, et al. Mutation in mce operons attenuates Mycobacterium tuberculosis virulence. Microbes and Infection. 2005;7(3):325–334. doi: 10.1016/j.micinf.2004.11.007. [DOI] [PubMed] [Google Scholar]
- 4.Arruda S, Bomfim G, Knights R, Huima-Byron T, Riley LW. Cloning of an M. tuberculosis DNA fragment associated with entry and survival inside cells. Science. 1993;261(5127):1454–1457. doi: 10.1126/science.8367727. [DOI] [PubMed] [Google Scholar]
- 5.Chitale S, Ehrt S, Kawamura I, et al. Recombinant Mycobacterium tuberculosis protein associated with mammalian cell entry. Cellular Microbiology. 2001;3(4):247–254. doi: 10.1046/j.1462-5822.2001.00110.x. [DOI] [PubMed] [Google Scholar]
- 6.Flesselles B, Anand NN, Remani J, Loosmore SM, Klein MH. Disruption of the mycobacterial cell entry gene of Mycobacterium bovis BCG results in a mutant that exhibits a reduced invasiveness for epithelial cells. FEMS Microbiology Letters. 1999;177(2):237–242. doi: 10.1111/j.1574-6968.1999.tb13738.x. [DOI] [PubMed] [Google Scholar]
- 7.Sassetti CM, Boyd DH, Rubin EJ. Genes required for mycobacterial growth defined by high density mutagenesis. Molecular Microbiology. 2003;48(1):77–84. doi: 10.1046/j.1365-2958.2003.03425.x. [DOI] [PubMed] [Google Scholar]
- 8.Shimono N, Morici L, Casali N, et al. Hypervirulent mutant of Mycobacterium tuberculosis resulting from disruption of the mce1 operon. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(26):15918–15923. doi: 10.1073/pnas.2433882100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tehilian EZ, Desel C, Forbes EK, et al. Immunogenicity and protective efficacy of prime-boost regimens with recombinant ΔureC hly+Mycobacterium bovis BCG and modified vaccinia virus ankara expressing M. tuberculosis antigen 85A against murine tuberculosis. Infection and Immunity. 2009;77(2):622–631. doi: 10.1128/IAI.00685-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Marjanovic O, Miyata T, Goodridge A, Kendall LV, Riley LW. Mce2 operon mutant strain of Mycobacterium tuberculosis is attenuated in C57BL/6 mice. Tuberculosis. 2010;90(1):50–56. doi: 10.1016/j.tube.2009.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Aguilar LD, Infante E, Bianco MV, Cataldi A, Bigi F, Pando RH. Immunogenicity and protection induced by Mycobacterium tuberculosis mce-2 and mce-3 mutants in a Balb/c mouse model of progressive pulmonary tuberculosis. Vaccine. 2006;24(13):2333–2342. doi: 10.1016/j.vaccine.2005.11.051. [DOI] [PubMed] [Google Scholar]
- 12.Obregón-Henao A, Shanley C, Bianco MV, et al. Vaccination of guinea pigs using mce operon mutants of Mycobacterium tuberculosis. Vaccine. 2011;29(26):4302–4307. doi: 10.1016/j.vaccine.2011.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hewinson RG, Vordermeier HM, Buddle BM. Use of the bovine model of tuberculosis for the development of improved vaccines and diagnostics. Tuberculosis. 2003;83(1–3):119–130. doi: 10.1016/s1472-9792(02)00062-8. [DOI] [PubMed] [Google Scholar]
- 14.Bigi F, Gioffré A, Klepp L, et al. Mutation in the P36 gene of Mycobacterium bovis provokes attenuation of the bacillus in a mouse model. Tuberculosis. 2005;85(4):221–226. doi: 10.1016/j.tube.2004.11.002. [DOI] [PubMed] [Google Scholar]
- 15.Parish T, Stoker NG. Use of flexible cassette method to generate a double unmarked Mycobacterium tuberculosis tlyA plcABC mutant by gene replacement. Microbiology. 2000;146(8):1969–1975. doi: 10.1099/00221287-146-8-1969. [DOI] [PubMed] [Google Scholar]
- 16.Buddle BM. Tuberculosis vaccines for cattle: the way forward. Expert Review of Vaccines. 2010;9(10):1121–1124. doi: 10.1586/erv.10.112. [DOI] [PubMed] [Google Scholar]
- 17.Lopez-Valencia G, Renteria-Evangelista T, Williams JDJ, Licea-Navarro A, Mora-Valle ADL, Medina-Basulto G. Field evaluation of the protective efficacy of Mycobacterium bovis BCG vaccine against bovine tuberculosis. Research in Veterinary Science. 2010;88(1):44–49. doi: 10.1016/j.rvsc.2009.05.022. [DOI] [PubMed] [Google Scholar]
- 18.Flaherty DK, Vesosky B, Beamer GL, Stromberg P, Turner J. Exposure to Mycobacterium avium can modulate established immunity against Mycobacterium tuberculosis infection generated by Mycobacterium bovis BCG vaccination. Journal of Leukocyte Biology. 2006;80(6):1262–1271. doi: 10.1189/jlb.0606407. [DOI] [PubMed] [Google Scholar]
- 19.Behr MA, Wilson MA, Gill WP, et al. Comparative genomics of BCG vaccines by whole-genome DNA microarray. Science. 1999;284(5419):1520–1523. doi: 10.1126/science.284.5419.1520. [DOI] [PubMed] [Google Scholar]
- 20.Brosch R, Gordon SV, Marmiesse M, et al. A new evolutionary scenario for the Mycobacterium tuberculosis complex. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(6):3684–3689. doi: 10.1073/pnas.052548299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kalra M, Khuller GK, Grover A, Behera D, Wanchu A, Verma I. Utility of a combination of RD1 and RD2 antigens as a diagnostic marker for tuberculosis. Diagnostic Microbiology and Infectious Disease. 2010;66(2):153–161. doi: 10.1016/j.diagmicrobio.2009.09.005. [DOI] [PubMed] [Google Scholar]
- 22.Kamath AT, Fruth U, Brennan MJ, et al. New live mycobacterial vaccines: the Geneva consensus on essential steps towards clinical development. Vaccine. 2005;23(29):3753–3761. doi: 10.1016/j.vaccine.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 23.Caccamo N, Meraviglia S, La Mendola C, Guggino G, Dieli F, Salerno A. Phenotypical and functional analysis of memory and effector human CD8 T cells specific for mycobacterial antigens. Journal of Immunology. 2006;177(3):1780–1785. doi: 10.4049/jimmunol.177.3.1780. [DOI] [PubMed] [Google Scholar]
- 24.Chen CY, Huang D, Wang RC, et al. A critical role for CD8 T cells in a nonhuman primate model of tuberculosis. PLoS Pathogens. 2009;5(4) doi: 10.1371/journal.ppat.1000392. Article ID e1000392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liébana E, Girvin RM, Welsh M, Neill SD, Pollock JM. Generation of CD8+ T-cell responses to Mycobacterium bovis and mycobacterial antigen in experimental bovine tuberculosis. Infection and Immunity. 1999;67(3):1034–1044. doi: 10.1128/iai.67.3.1034-1044.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.McShane H, Pathan AA, Sander CR, et al. Recombinant modified vaccinia virus Ankara expressing antigen 85A boosts BCG-primed and naturally acquired antimycobacterial immunity in humans. Nature Medicine. 2004;10(12):1240–1244. doi: 10.1038/nm1128. [DOI] [PubMed] [Google Scholar]
- 27.Ronan EO, Lee LN, Beverley PCL, Tchilian EZ. Immunization of mice with a recombinant adenovirus vaccine inhibits the early growth of Mycobacterium tuberculosis after infection. Plos ONE. 2009;4(12) doi: 10.1371/journal.pone.0008235. Article ID e8235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lindenstrøm T, Agger EM, Korsholm KS, et al. Tuberculosis subunit vaccination provides long-term protective immunity characterized by multifunctional CD4 memory T cells. Journal of Immunology. 2009;182(12):8047–8055. doi: 10.4049/jimmunol.0801592. [DOI] [PubMed] [Google Scholar]
- 29.Waters WR, Palmer MV, Thacker TC, et al. Tuberculosis immunity: opportunities from studies with cattle. Clinical and Developmental Immunology. 2011;2011:11 pages. doi: 10.1155/2011/768542. Article ID 768542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Santangelo MP, Goldstein J, Alito A, et al. Negative transcriptional regulation of the mce3 operon in Mycobacterium tuberculosis. Microbiology. 2002;148(10):2997–3006. doi: 10.1099/00221287-148-10-2997. [DOI] [PubMed] [Google Scholar]
- 31.van Soolingen D, Hermans PW, de Haas PE, Soll DR, van Embden JD. Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains: evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis. Journal of Clinical Microbiology. 1991;29(11):2578–2586. doi: 10.1128/jcm.29.11.2578-2586.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Buddle BM, De Lisle GW, Pfeffer A, Aldwell FE. Immunological responses and protection against Mycobacterium bovis in calves vaccinated with a low dose of BCG. Vaccine. 1995;13(12):1123–1130. doi: 10.1016/0264-410x(94)00055-r. [DOI] [PubMed] [Google Scholar]
- 33. R Foundation for Statistical Computing R Development Core Team: R: A language and environment for statistical computing, Austria, 2011.
- 34.Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM. Mixed Effects Models and Extensions in Ecology with R. Springer Science Business Media, LLC; 2009. [Google Scholar]
- 35.Blanco FC, Schierloh P, Bianco MV, et al. Study of the immunological profile towards Mycobacterium bovis antigens in naturally infected cattle. Microbiology and Immunology. 2009;53(8):460–467. doi: 10.1111/j.1348-0421.2009.00141.x. [DOI] [PubMed] [Google Scholar]