

Abstract
The mesopontine rostromedial tegmental nucleus (RMTg) is a GABAergic structure in the ventral midbrain and rostral pons that, when activated, inhibits dopaminergic neurons in the ventral tegmental area and substantia nigra compacta. Additional strong outputs from the RMTg to the pedunculopontine tegmental nucleus pars dissipata, dorsal raphe nucleus, and the pontomedullary gigantocellular reticular formation were identified by anterograde tracing. RMTg neurons projecting to the ventral tegmental area express the immediate early gene Fos upon psychostimulant administration. The present study was undertaken to determine if neurons in the RMTg that project to the additional structures listed above also express Fos upon psychostimulant administration and, if so, whether single neurons in the RMTg project to more than one of these structures. We found that about 50% of RMTg neurons exhibiting retrograde labeling after injections of retrograde tracer in the dorsal raphe or pars dissipata of the pedunculopontine tegmental nucleus express Fos after acute methamphetamine exposure. Also, we observed that a significant number of RMTg neurons project both to the ventral tegmental area and one of these structures. In contrast, methamphetamine-elicited Fos expression was not observed in RMTg neurons labeled with retrograde tracer following injections into the pontomedullary reticular formation. The findings suggest that the RMTg is an integrative modulator of multiple rostrally projecting structures.
Keywords: RMTg, Psychostimulant, Fos, Addiction
Introduction
The ventral tegmental area (VTA) gives rise to the mesocorticolimbic dopaminergic system (Dahlstrom and Fuxe 1964) and is the substrate for the initiation of behavioral sensitization to psychostimulants (Kalivas and Webber 1988; Kalivas and Stewart 1991; Hooks et al. 1992; Perugini and Vezina 1994; Cador et al. 1995, 1999; Bjijou et al. 1996; Vezina 1996; Vanderschuren and Kalivas 2000). Dopamine (DA) neurons in the VTA respond with increases or decreases in their firing rates to novelty, reward, aversive stimuli, cues predicting rewards or aversive stimuli, and omissions of expected rewards or aversive stimuli (Schultz 1998, 2007; Bromberg-Martin et al. 2010). Understanding of the mechanisms by which the activity of DA neurons is linked to circumstances, however, remains incomplete. The lateral habenula (LHb), which increases firing in response to aversive stimuli and reward omission (Christoph et al. 1986; Gao et al. 1996; Ji and Shepard 2007; Matsumoto and Hikosaka 2007; Shumake et al. 2010; Hong et al. 2011), could conceivably inhibit dopamine neurons in turn, due to its direct projections to the VTA (Herkenhan and Nauta 1979; Araki et al. 1988; Omelchenko et al. 2009). However, LHb projections to the ventral tegmental area are mainly glutamatergic (Geisler et al. 2007; Brinschwitz et al. 2010), thus suggesting that LHb inhibition of DA neurons is mediated indirectly, possibly by a relay in another structure.
Evidence that such a mediator region exists was provided by Chou et al. (2004), who showed that a cluster of cells in the paramedian tegmentum behind the VTA projects to the VTA and influences fear-elicited behaviors. Subsequently, Jhou and Gallagher (2007) (same person as Chou in Chou et al. 2004) showed that these neurons are activated by aversive stimuli. In definitive papers, Jhou and colleagues (Jhou et al. 2009a, b) showed that this structure is GABAergic, receives dense projections from the LHb, and projects strongly to the VTA and substantia nigra compacta (SNC). Earlier, Scammell et al. (2000) had identified in rats a cluster of neurons that express Fos after administration of modafinil, and designated it as the “retroVTA”. Later, Perrotti et al. (2005) showed a similar cluster that expressed deltaFosB, a long-lived splice variant of FosB, after the chronic administration of amphetamine and called it the “posterior tail of the VTA”. Geisler et al. (2008) demonstrated what appears to be the same cluster of Fos-expressing neurons after cocaine self-administration, and showed that the neurons comprising it project to the VTA. Upon further investigation, Jhou et al. (2009b) and Kaufling et al. (2009, 2010a) concluded that all these workers were studying the same structure, which Jhou et al. (2009b) named the mesopontine rostromedial tegmental nucleus (RMTg). This GABAergic midbrain structure has now been shown to mediate the LHb influence on the VTA (Hong et al. 2011): glutamatergic LHb neurons project to GABAergic neurons of the RMTg, which in turn exert an inhibitory influence on dopaminergic VTA/SNC neurons. Jhou et al. (2009b) utilized the anterogradely transported tracer, PHA-L, to characterize the efferent projections of the RMTg, and showed particularly robust projections to the pedunculopontine tegmental nucleus pars dissipata (PPTg), dorsal raphe nucleus (DR), and pontomedullary paramedian gigantocellular reticular formation (RtGi), indicating that important influences of the RMTg are not necessarily limited to the VTA.
At present, the criteria available to designate a neuron as belonging to the RMTg include input from the LHb, projection to the VTA/SNC, GABAergic phenotype, and the expression of Fos following some aversive stimuli (Jhou et al. 2009a), but invariably after administration of any psychostimulant drug (Kaufling et al. 2010b). The present study was undertaken to determine if neurons in the RMTg that project to the PPTg, DR, and RtGi also express Fos upon psychostimulant administration. At least for the DR and PPTg, this was found to be the case, which led us to examine if single neurons in the RMTg might project to more than one of these structures. We herein show that single neurons of the RMTg do indeed project to multiple targets, suggesting that RMTg neurons may be involved in modulating activity coordinately in several structures, including the PPTg, DR, and VTA.
Materials and methods
Male Sprague–Dawley rats (Harlan, Indianapolis, IN, USA) weighing 225–250 g were used in accordance with the guidelines mandated in the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The rats were housed on a 12-h light–dark cycle in groups of two to four until surgeries were performed, after which all were singly housed. Access to food and water was provided ad libitum. Unless otherwise stated, chemicals were purchased from Sigma (St. Louis, MO, USA).
Tracer injections
The rats were deeply anesthetized by intraperitoneal injections of a mixture of ketamine (72 mg/kg) and xylazine (11.2 mg/kg). The anesthetic was injected as a cocktail consisting of 45% ketamine (100 mg/ml), 35% xylazine (20 mg/ml), and 20% saline at a dose of 0.16 ml/100 g of body weight. The anesthetized rats were placed into a Kopf stereotaxic instrument and an incision was made to expose the scull, in which a small burr hole was made over the injection site. The retrograde tracers, cholera toxin β subunit (CTβ; List Biological Laboratories, Campbell, CA; 1% in ddH2O) or Fluoro-Gold (FG; Fluorochrome Inc., Englewood, CO; 1% in 0.1 M cacodylate buffer, pH 7.4) were injected into targeted brain structures (VTA, DR, PPTg or RtGi) using 1.0 mm filament-containing borosilicate glass pipettes pulled to tip diameters of 15–20 μm. The tracer solution was injected ionotophoretically using positive current pulses (7 s on and 7 s off for 15 min) of 1 μA (for FG) and 3 μA (for CTβ). In order to generate more retrograde labeling in the vicinity of the RMTg, some injections of CTβ into RtGi were done with glass pipettes pulled to tip diameters of 30–40 μm from which the tracer was expelled by air pressure, resulting in an injected volume of approximately 0.1 μl. The incisions were closed with wound clips, and the rats were kept warm until they awakened.
Fixation of brains
Three days following tracer injections, the rats were given an intraperitoneal injection of methamphetamine (10 mg/kg). Two hours following methamphetamine injections, the rats were deeply anesthetized as described and perfused transaortically with 0.01 M Sorensen’s phosphate buffer (SPB; pH 7.4) containing 0.9% sodium chloride and 2.5% sucrose, followed by 0.1 M SPB (pH 7.4) containing 4% paraformaldehyde and 2.5% sucrose. The perfused brains were removed and placed in the same fixative for 4 h and then in a 30% sucrose solution overnight. Five adjacent series of 50 μm sections were collected, each representing an entire brain from the frontal pole to the caudal medulla at sampled intervals of 250 μm. Each series of sections was stored in a separate glass vial at −20°C in cryoprotectant consisting of 0.01 M SPB containing 30% sucrose and 30% ethylene glycol.
Immunohistochemistry
Some series of sections from each case were processed for immunoperoxidase and others for immunofluorescence histochemistry. Sections processed for immunoperoxidase histochemistry, which produces permanent, non-fading preparations, were used for the evaluation of injection sites, efficacy of retrograde transport of the tracer and general distribution of retrograde labeling relative to the Fos-expressing RMTg. However, double-immunoperoxidase preparations show double labeling unreliably. Accordingly, additional sections processed for immunofluorescence histochemistry were used to reliably demonstrate and quantify double labeling.
Immunoperoxidase histochemistry
For immunoperoxidase processing, one series from each case was immersed in 0.1 M SPB containing 0.1% Triton X-100 (SPB-t) and anti-Fos antibody (Calbiochem, San Diego, CA, USA, catalog no. PC38) made in rabbit at a dilution of 1:3,000. The following day, the sections were rinsed in 0.1 M SPB-t and immersed for 1 h in 0.1 M SPB-t containing biotinylated antibody made in donkey against rabbit IgGs at a dilution of 1:200 (Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA, catalog no. 711-065-152). Afterward, the sections were rinsed in 0.1 M SPB-t and then immersed for 1 h in 0.1 M SPB containing avidin–biotin-peroxidase complex (ABC) at a dilution of 1:200 (Vector Laboratories, Burlingame, CA, USA). After rinsing in 0.1 M SPB, the sections were reacted in 0.025 M Tris buffer (pH 8.0) containing 0.015% 3,3′-diaminobenzidine (DAB), 0.4% nickel ammonium sulfate, and 0.003% hydrogen peroxide (NiDAB), which generates an insoluble black reaction product. The sections were then rinsed in 0.1 M SPB and immersed overnight in 0.1 M SPB containing polyclonal antibodies against CTβ (anti-CTβ, List Biological Laboratories, Cambell, CA, USA, catalog no. 7032A5) made in goat and used at a dilution of 1:4,000. The following day, after rinsing in 0.1 M SPB, the sections were immersed for 1 h in 0.1 M SPB containing biotinylated antibody made in donkey against goat IgGs at a dilution of 1:200 (Jackson, catalog no. 705-065-003). Afterward, the sections were rinsed in 0.1 M SPB and then immersed for 1 h in 0.1 M SPB containing ABC at a dilution of 1:200 (Vector). After rinsing in 0.1 M SPB, the sections were reacted in 0.1 M SPB containing 0.05% 3,3′-diaminobenzidine and 0.003% hydrogen peroxide (DAB), which generates an insoluble brown reaction product. In this variation of the immunoperoxidase protocol and those to follow, the sections were then mounted onto gelatin-coated slides and coverslipped with Permount (Fisher, Pittsburgh, PA, USA).
For immunoperoxidase in cases involving injection of FG into one brain area and CTβ into a different site, one series from each case was immersed overnight in 0.1 M SPB-t containing anti-FG (Chemicon International, Temecula, CA, USA, catalog no. AB153), made in rabbit at a dilution of 1:3,000. The following day, the sections were rinsed in 0.1 M SPB-t and immersed for 1 h in 0.1 M SPB containing biotinylated antibody made in donkey against rabbit IgGs at a dilution of 1:200 (Jackson). Afterward, the sections were rinsed in 0.1 M SPB-t and then immersed for 1 h in 0.1 M SPB containing ABC at a dilution of 1:200 (Vector). After rinsing in 0.1 M SPB, the sections were reacted in NiDAB. The sections were then rinsed in SPB and immersed overnight in SPB containing polyclonal anti-CTβ used at a dilution of 1:4,000. The following day, after rinsing in 0.1 M SPB, the sections were immersed for 1 h in 0.1 M SPB containing biotinylated antibody made in donkey against goat IgGs at a dilution of 1:200 (Jackson). Afterward, the sections were rinsed in 0.1 M SPB and then immersed for 1 h in 0.1 M SPB containing ABC at a dilution of 1:200 (Vector). After more rinsing in 0.1 M SPB, the sections were reacted in NiDAB, as described above.
For injections into the DR, one series of each case was immersed overnight in 0.1 M SPB-t containing anti-5-hydroxytryptophan (5-HT, Immunostar, Hudson, WI, USA, catalog no. 2446), made in rabbit at a dilution of 1:1,000. For injections into the PPTg, one series from each case was immersed overnight in 0.1 M SPB-t containing anti-nitric oxide synthase (Nos, Sigma, catalog no. 6715) made in rabbit at a dilution of 1:3,000. The following day, the sections that had been immersed in either anti-5-HT or anti-Nos were further processed similarly. They were rinsed in 0.1 M SPB-t and immersed for 1 h in 0.1 M SPB containing biotinylated antibody made in donkey against rabbit IgGs at a dilution of 1:200 (Jackson). Afterward, the sections were rinsed in 0.1 M SPB-t and then immersed for 1 h in 0.1 M SPB containing ABC at a dilution of 1:200 (Vector). After more rinsing in 0.1 M SPB, the sections were reacted in NiDAB.
Fluorescence immunohistochemistry
For immunofluorescence processing, one series from each case was immersed overnight in 0.1 M SPB-t and polyclonal anti-CTβ used at a dilution of 1:4,000. The following day, the sections were rinsed in 0.1 M SPB-t and immersed for 1 h in 0.1 M SPB-t containing DyLight 488 conjugated to anti-sheep IGg made in donkey (Jackson, catalog no. 713-485-14), which recognizes the anti-CTβ made in goat, used at a concentration of 1:200. The sections then were rinsed in 0.1 M SPB and immersed overnight in 0.1 M SPB containing anti-Fos at a concentration of 1:3,000. The following day, the sections were rinsed in 0.1 M SPB and immersed for 1 h in 0.1 M SPB containing DyLight 594 conjugated to anti-rabbit IGg made in donkey (Jackson, catalog no. 711-515-152) used at 1:200. The sections were rinsed in 0.1 M SPB and mounted and coverslipped with ProLong Gold antifade reagent (Invitrogen).
For immunofluorescence in cases involving injection of FG into one brain site and CTβ into another, one series from each case was first rinsed in 0.1 M SPB-t and immersed overnight in 0.1 M SPB-t containing anti-CTβ at a concentration of 1:4,000. The following day, the sections were rinsed in 0.1 M SPB-t and immersed for 1 h in 0.1 M SPB-t containing DyLight 488-anti-sheep IGg conjugate (Jackson) at a concentration of 1:200. The sections were rinsed in 0.1 M SPB and immersed overnight in 0.1 M SPB containing anti-FG at a concentration of 1:5,000. The following day, the sections were rinsed in 0.1 M SPB and immersed for 1 h in 0.1 M SPB containing DyLight 594-anti-rabbit conjugate (Jackson) at a concentration of 1:200. The sections were rinsed in 0.1 M SPB and mounted onto gelatin-coated slides and coverslipped with ProLong Gold.
Analysis of labeling
Sections were viewed with an Olympus BX51 microscope in the brightfield or immunofluorescence modes, and digital micrographs were generated using a DVC 2000C-00-GE-MBF digital camera. Neurons were plotted with the aid of the Neurolucida hardware–software platform (MBF Bioscience, Williston, VT, USA, version 5.65) such that retrogradely labeled neurons, Fos-containing neurons and double-labeled neurons were designated with distinct markers. These plots were exported into Adobe Illustrator CS2 to generate maps showing three sections of the RMTg spaced at 250 μm for each case. Quantification of the markers for Fos, retrograde labeling and double labeling was performed with the aid of the Neurolucida NeuroExplorer program (MBF Bioscience, version 4.70.3). Counts were generated uniformly across all cases by delineating the RMTg with a circle of arbitrarily designated area (0.32 mm2) surrounding the spot of densest Fos labeling. The region just surrounding the RMTg, was designated by a larger (1.15 mm2) circle from which the area of the smaller circle was subtracted, leaving 0.83 mm2. The numbers of double-labeled neurons, i.e. Fos-expressing retrogradely labeled neurons within these two arbitrarily designated regions were expressed as percentages of total retrogradely labeled neurons therein. In order to determine if enrichments of double labeling were present in the RMTg, the mean percentages of double-labeled neurons within the RMTg circle were compared with those in the arbitrarily defined area just outside of the RMTg using a paired t test with a 95% confidence interval. For each injection site or combination of injection sites, 4–11 cases were used (Tables 1, 2). Statistics were done with the aid of SPSS software.
Table 1.
Numbers of retrogradely labeled and double-labeled neurons following brainstem tracer injections
| Injection site | Retrogradely labeled neurons | Double-labeled neurons | Percent double-labeled neurons | |||
|---|---|---|---|---|---|---|
| Case | RMTg | Outside | RMTg | Outside | RMTg | Outside |
| VTA | ||||||
| 10192 | 152 | 118 | 74 | 17 | 48% | 14% |
| 11077 | 150 | 60 | 75 | 17 | 50% | 28% |
| 11080 | 59 | 18 | 32 | 1 | 54% | 5% |
| 11083 | 115 | 115 | 46 | 7 | 40% | 6% |
| 12051 | 176 | 131 | 198 | 28 | 47% | 14% |
| Mean ± SE | 130 ± 35 | 88 ± 21 | 85 ± 29 | 14 ± 4 | 48 ± 2 | 13 ± 4 |
| p = 0.001a | ||||||
| PPTg | ||||||
| 10193 | 84 | 101 | 33 | 10 | 40% | 9% |
| 10202 | 104 | 80 | 80 | 18 | 77% | 30% |
| 10203 | 84 | 20 | 35 | 6 | 42% | 22% |
| 11058 | 77 | 52 | 50 | 14 | 65% | 26% |
| 11059 | 72 | 72 | 46 | 18 | 64% | 25% |
| 12050 | 117 | 107 | 59 | 8 | 50% | 7% |
| Mean ± SE | 89 ± 7 | 72 ± 13 | 50 ± 7 | 12 ± 2 | 54 ± 6 | 19 ± 4 |
| p = 0.001 | ||||||
| DR | ||||||
| 10200 | 63 | 80 | 22 | 4 | 35% | 5% |
| 11060 | 91 | 140 | 44 | 32 | 48% | 22% |
| 11061 | 44 | 98 | 30 | 2 | 68% | 2% |
| 11119 | 66 | 92 | 34 | 3 | 51% | 4% |
| 12053 | 50 | 65 | 23 | 7 | 46% | 11% |
| Mean ± SE | 71 ± 11 | 97 ± 10 | 35 ± 5 | 9 ± 4 | 47 ± 5 | 9 ± 4 |
| p = 0.005 | ||||||
| RtGi | ||||||
| 11180 | 37 | 83 | 5 | 9 | 14% | 11% |
| 11184 | 23 | 14 | 2 | 2 | 8% | 14% |
| 11191 | 25 | 131 | 1 | 21 | 4% | 16% |
| 11192 | 38 | 129 | 8 | 27 | 21% | 21% |
| 11214 | 17 | 43 | 1 | 4 | 6% | 9% |
| 11215 | 32 | 72 | 2 | 6 | 6% | 8% |
| 11216 | 49 | 76 | 2 | 11 | 4% | 14% |
| 11217 | 27 | 73 | 2 | 15 | 7% | 20% |
| 11223 | 26 | 125 | 5 | 23 | 19% | 18% |
| 11224 | 40 | 99 | 3 | 15 | 7% | 15% |
| 11225 | 33 | 89 | 1 | 18 | 3% | 20% |
| Mean ± SE | 31 ± 3 | 85 ± 10 | 3 ± 0.5 | 13 ± 2 | 8 ± 2 | 15 ± 1 |
| p = 0.008 | ||||||
All entries reflect data from the RMTg and region just surrounding it (see “Materials and methods”) from 3 sections/case
aPaired samples t test
Table 2.
Numbers of retrogradely labeled and double-labeled neurons for tracer (FG and CTβ) injection combinations
| Injection combination | Retrogradely labeled neurons | Double-labeled neurons | Percent double-labeled neurons | ||||
|---|---|---|---|---|---|---|---|
| Case | RMTg | Surround | RMTg | Surround | RMTg | Surround | |
| PPTg–VTA | |||||||
| 11082 | 230 (108, 122)a | 127 (64, 63) | 28 | 2 | 12% | 2% | |
| 11083 | 272 (129, 143) | 162 (67, 95) | 35 | 6 | 13% | 3% | |
| 11090 | 164 (71, 93) | 108 (65, 43) | 8 | 0 | 5% | 0% | |
| 12051 | 146 (57, 89) | 112 (62, 50) | 9 | 1 | 6% | 1% | |
| Mean ± SE | 203 ± 29 | 127 ± 12 | 20 ± 12 | 2 ± 1 | 9 ± 2 | 1 ± 0.5 | |
| p = 0.01b | |||||||
| DR–VTA | |||||||
| 11118 | 226 (115, 111) | 207 (92, 115) | 18 | 3 | 8% | 1% | |
| 11119 | 188 (74, 114) | 145 (68, 77) | 20 | 6 | 10% | 4% | |
| 11121 | 191 (78, 113) | 147 (65, 82) | 23 | 4 | 12% | 3% | |
| 12052 | 203 (81, 122) | 123 (44, 79) | 18 | 4 | 9% | 3% | |
| Mean ± SE | 202 ± 8 | 155 ± 18 | 20 ± 1 | 4 ± 0.5 | 10 ± 1 | 2 ± 0.5 | |
| p = 0.02 | |||||||
| PPTg–DR | |||||||
| 11260 | 98 (60, 38) | 65 (33, 32) | 6 | 0 | 6% | 0% | |
| 11261 | 102 (42, 60) | 36 (19, 17) | 9 | 0 | 9% | 0% | |
| 11264 | 124 (53, 71) | 172 (74, 98) | 4 | 3 | 3% | 2% | |
| 11265 | 65 (49, 16) | 46 (25, 21) | 3 | 0 | 5% | 0% | |
| 12055 | 70 (29, 41) | 54 (20, 34) | 3 | 1 | 4% | 2% | |
| Mean ± SE | 91 ± 10 | 74 ± 24 | 5 ± 1 | 0.8 ± 0.5 | 5 ± 1 | 0.5 ± 0.5 | |
| p = 0.02 | |||||||
All entries reflect data from the RMTg and region just surrounding it (see “Materials and methods”) from 3 sections/case
aInjection 1, injection 2
bPaired samples t test
Results
Single CTβ injections
VTA
Consistent with a published report in which the retrograde tracer Fluoro-Gold was used (Jhou et al. 2009b), injections of CTβ into the VTA exhibited double labeling with Fos immunoreactivity in 48 ± 2% of retrogradely labeled neurons (n = 5) within the cluster of Fos-immunoreactive neurons designated as the RMTg, as compared to 13 ± 4% double labeling of retrogradely labeled neurons in the area immediately surrounding the RMTg (Figs. 1, 3a–c, 7). A representative plot of the results for one VTA case is shown in Fig. 3, which illustrates rostral (Fig. 3a) and more caudal (Fig. 3b) levels of the RMTg. The maps in Fig. 3 and similar maps to follow (i.e., Figs. 6, 11) show localization of the retrograde tracer, Fos expression, and, if present, double-labeled neurons. In contrast, in cases in which no methamphetamine was administered, there were few Fos-immunoreactive neurons (data not shown), and in cases in which the injection sites were misplaced, there was no enrichment of retrogradely labeled or double-labeled neurons in the RMTg as compared to the surrounding region.
Fig. 1.
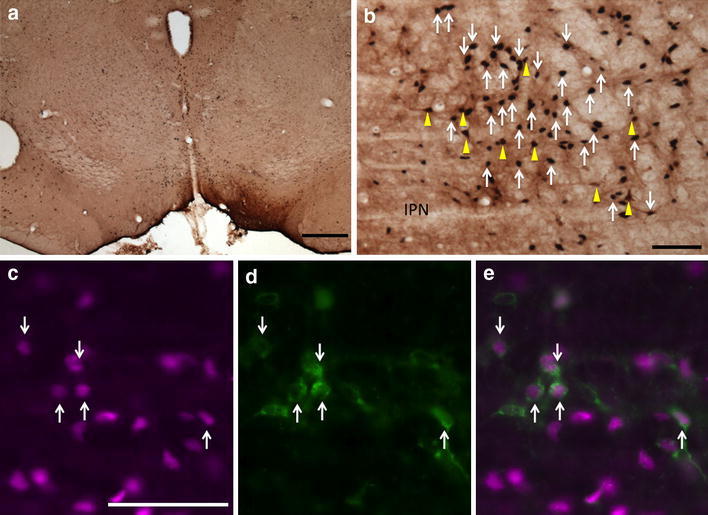
Fos-expressing retrogradely labeled neurons in the RMTg following injection of CTβ in the VTA. a CTβ injection site in the VTA. b Immunoperoxidase detection of retrogradely labeled neurons (brown, yellow arrows) and Fos expression (black, white arrows) in the RMTg. c–e Immunofluorescence images of RMTg neurons showing Fos immunoreactivity (c, magenta), retrograde labeling (d, green), and a merged image (e). White arrows in c–e indicate double-labeled neurons. The round hole in the section at the left edge of the micrograph is a tissue punch marking the right hemisphere. IPN interpeduncular nucleus. Scale bars 400 μm in a; 100 μm in b and c
Fig. 3.
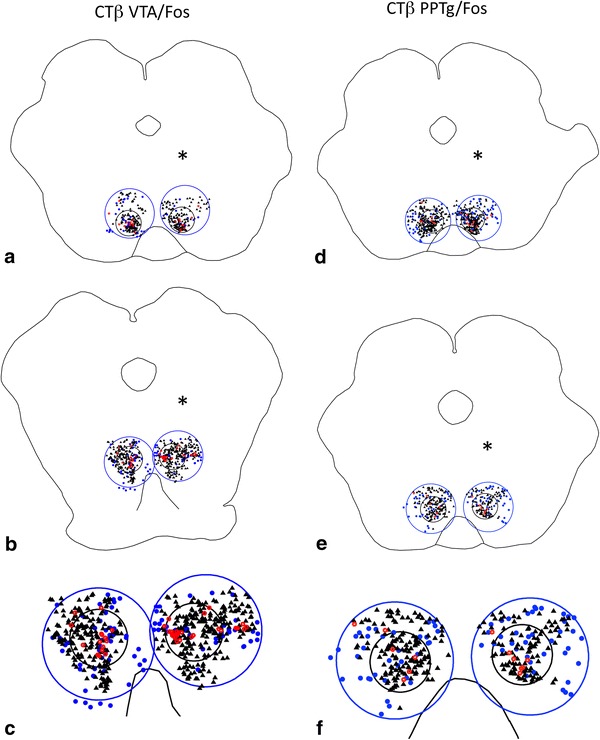
Representative plots at rostral (a, d) and caudal (b, e) levels of the RMTg (small black circles) from cases that received injections of tracer in the VTA (a–c) and PPTg (d–f) and were processed to exhibit retrogradely labeled neurons and Fos expression. Brains were injected on the sides indicated by the asterisks. Large blue circles indicate the regions bordering the RMTg. c and f Show enlargements of the caudal levels. Black triangles Fos expressing neurons, blue dots retrogradely labeled neurons, red circles double-labeled neurons
Fig. 7.
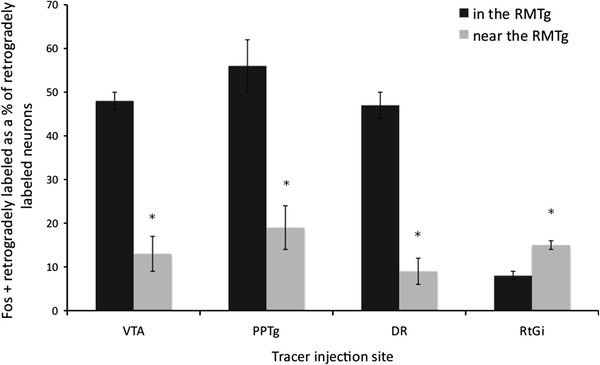
Graph showing the percentages of double-labeled neurons in the RMTg compared to the region surrounding it for four different injection sites: PPTg (n = 6), DR (n = 5), VTA (n = 5), and RtGi (n = 11). *p < 0.01 (paired t test)
Fig. 6.
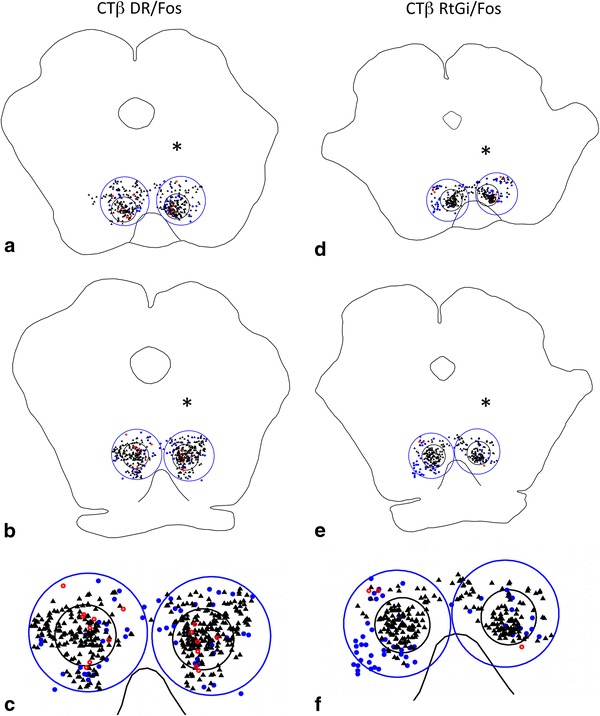
Representative plots at rostral (a, d) and caudal (b, e) levels of the RMTg (small black circles) from cases that received injections of tracer in the DR (a–c) and RtGi (d–f) and were processed to exhibit retrogradely labeled neurons and Fos expression. Brains were injected on the sides indicated by the asterisks. Large blue circles indicate the regions bordering the RMTg. c and f Show enlargements of the caudal levels. Black triangles Fos expressing neurons, blue dots retrogradely labeled neurons, red circles double-labeled neurons
Fig. 11.
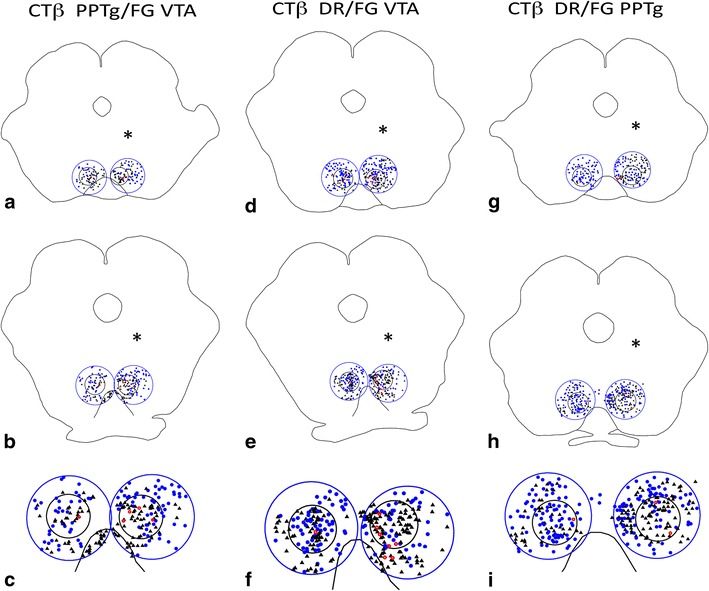
Representative plots at rostral (a, d, g) and caudal (b, e, h) levels of the RMTg (small black circles) from cases that received injections of CTβ tracer in the PPTg and FG in the VTA (a–c), cases that received injections of CTβ tracer in the DR and FG in the VTA (d–f), and cases that received injections of CTβ tracer in the DR and FG in the PPTg (g–i) and were processed to exhibit retrogradely labeled neurons. Brains were injected on the sides indicated by the asterisks. Large blue circles indicate the regions bordering the RMTg. c, f, and i Show enlargements of the caudal levels. Black triangles FG-containing neurons, blue dots CTβ-containing neurons; red circles double-labeled neurons
PPTg
Following injections of CTβ into the pars dissipata of the PPTg, 56 ± 6% of the retrogradely labeled neurons (n = 6) within the RMTg exhibited Fos-immunoreactivity compared to 19 ± 4% in the area immediately surrounding the RMTg (Figs. 2, 3d–f, 7). The locations of injection sites (Fig. 2a) within the PPTg dissipata were verified by demonstrating Nos expression (Fig. 2b), which co-localizes in cholinergic neurons that characterize the PPTg region (Armstrong et al. 1983). A representative plot of the results for one PPTg case is shown in Fig. 3, which illustrates rostral (Fig. 3d) and caudal (Fig. 3f) levels of the RMTg.
Fig. 2.
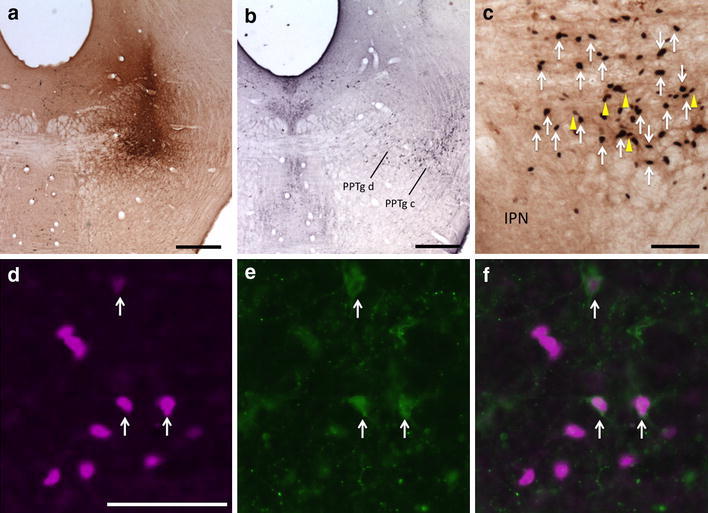
Fos-expressing retrogradely labeled neurons in the RMTg following injection of CTβ in the PPTg. a CTβ injection site in the PPTg. b Adjacent section processed for Nos immunoreactivity (black) verifies that the injection is in the PPTg, pars dissipata. c Retrogradely labeled neurons (yellow arrows) and Fos expression (white arrows) in the RMTg. d–f Immunofluorescence images of RMTg neurons showing Fos immunoreactivity (d, magenta), retrogradely labeled neurons (e, green), and a merged image (f). White arrows indicate double labeling (d–f). PPTg c pedunculopontine tegmental nucleus, pars compacta; PPTg d pedunculopontine tegmental nucleus, pars dissipata; IPN interpeduncular nucleus. Scale bars 400 μm in a and b; 100 μm in c and d
DR
47 ± 5% of retrogradely labeled neurons (n = 5) in the RMTg were double labeled following injections of CTβ in the DR compared to 9 ± 4% in the area immediately surrounding the RMTg (Figs. 4, 6a–c, 7). DR injection sites (Fig. 4a) were verified by demonstrating 5-HT immunoreactivity in adjacent sections (Fig. 4b), which identified serotoninergic neurons that characterize that structure (Aghajanian et al. 1967).
Fig. 4.
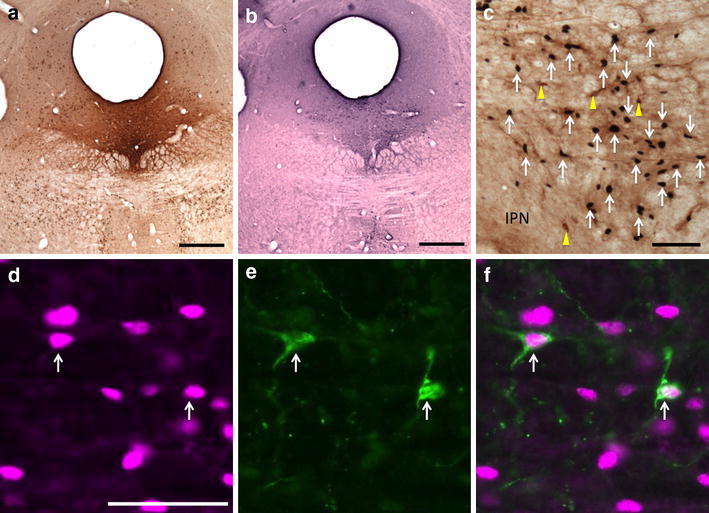
Fos-expressing retrogradely labeled neurons in the RMTg following injection of CTβ in the DR. a CTβ injection site in the DR. b Adjacent section processed for 5-HT immunoreactivity (black) verifies that the injection is in the DR. c Retrogradely labeled neurons (yellow arrows) and Fos expression (white arrows) in the RMTg. d–f Immunofluorescence images of RMTg neurons showing Fos immunoreactivity (d, magenta), retrogradely labeled neurons (e, green), and a merged image (f). White arrows indicate double labeling (d–f). IPN interpeduncular nucleus. Scale bars: 400 μm in a and b; 100 μm in c and d
RtGi
In contrast, injections of CTβ into the RtGi region (Fig. 5) resulted in few retrogradely labeled neurons in the RMTg (n = 11), of which 8 ± 2% were double labeled. Greater numbers of retrogradely labeled neurons were present in the area surrounding the RMTg following injections in the RtGi and the percentage of these that were double labeled (15 ± 1%) was comparable to that observed within the RMTg (Figs. 5, 6d–f, 7). A representative plot of the results from one RtGi injection is shown in Fig. 6, which illustrates rostral (Fig. 6d) and caudal (Fig. 6e) RMTg levels.
Fig. 5.
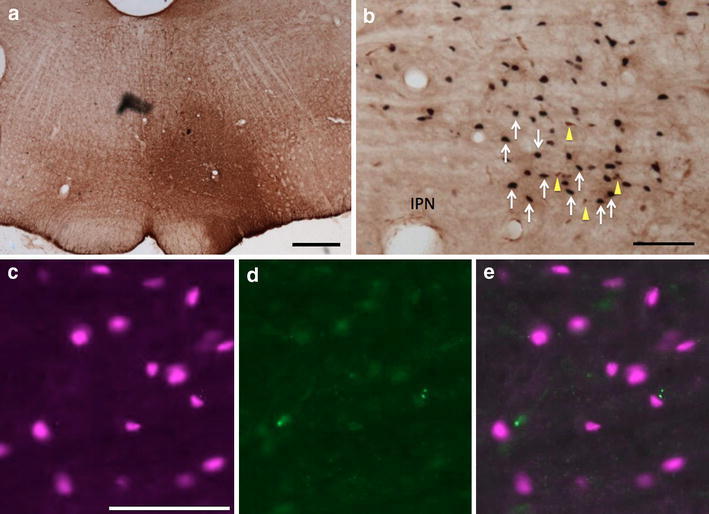
Fos-expressing retrogradely labeled neurons in the RMTg following injection of CTβ in the RtGi. a CTβ pressure injection site (see “Materials and methods” section) in the RtGi. b Immunoperoxidase detection of uncharacteristically numerous retrogradely labeled neurons (yellow arrows) and Fos expression (white arrows) in the RMTg. c–e Immunofluorescence images of RMTg neurons showing Fos immunoreactivity (c, magenta), retrograde labeling (d, green), and a merged image (e), confirming the absence of double labeling. IPN interpeduncular nucleus. Scale bars 400 μm in a; 100 μm in b and c
Double-injection tracer studies
In a series of cases in which FG was injected into the VTA and CTβ into the PPTg, 9 ± 2% of retrogradely labeled neurons within the RMTg (n = 4) exhibited both tracers, indicating RMTg neurons that project to both the VTA and PPTg, compared to 1 ± 0.5% in the area immediately surrounding the RMTg (Fig. 8). Similarly, injections of FG into the VTA and CTβ into the DR produced 10 ± 1% of retrogradely labeled neurons (n = 4) within the RMTg exhibiting both markers compared to 2 ± 0.5% in the area immediately surrounding the RMTg (Fig. 9). Injections of FG into PPTg and CTβ into DR produced 5 ± 1% of retrogradely labeled neurons (n = 5) within the RMTg exhibiting both markers compared to 0.5 ± 0.5% in the area immediately surrounding the RMTg (Fig. 10). Representative plots of these results for the PPTg/VTA, DR/VTA, and DR/PPTg injection combinations showing rostral (Fig. 11a, d, g) and more caudal (Fig. 11b, e, h) RMTg levels are shown in Fig. 11a–i
Fig. 8.
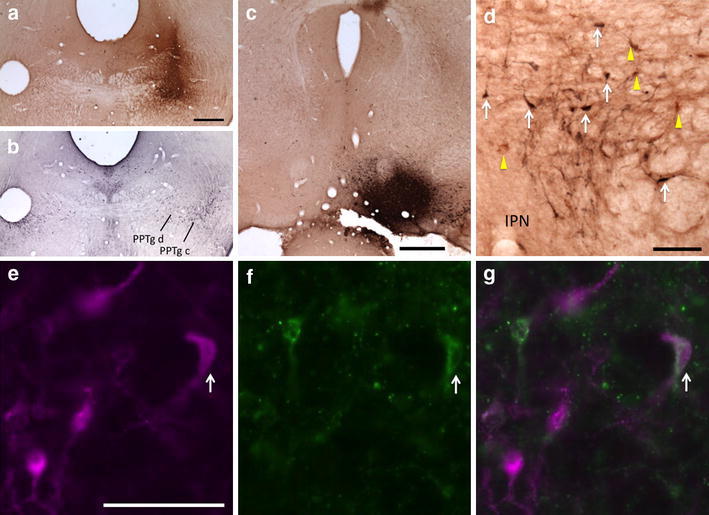
Retrogradely labeled neurons in the RMTg following injections of CTβ in the PPTg and FG in the VTA. a CTβ injection site in the PPTg. b Adjacent section processed for Nos immunoreactivity (black) verifies that the injection shown in ‘a’ is in the PPTg, pars dissipata. c FG injection site in the VTA. d CTβ-labeled neurons (yellow arrows) and FG-labeled neurons (white arrows) in the RMTg. e–g Immunofluorescence images of RMTg neurons showing FG-labeled RMTg neurons (e, magenta), CTβ-labeled RMTg neurons (f, green), and a merged image (g). White arrows indicate double labeling (e–g). PPTg c pedunculopontine tegmental nucleus, pars compacta; PPTg d pedunculopontine tegmental nucleus, pars dissipata; IPN interpeduncular nucleus. Scale bars 400 μm in a–c; 100 μm in d and e
Fig. 9.
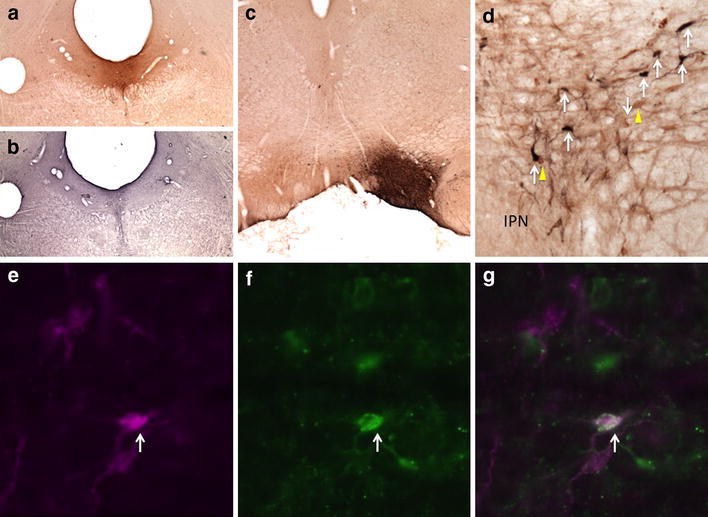
Retrogradely labeled neurons in the RMTg following injections of CTβ in the DR and FG in the VTA. a CTβ injection site in the DR. b Adjacent section processed for 5-HT immunoreactivity (black) verifies that the injection shown in a is in the DR. c FG injection site in the VTA. d CTβ-labeled neurons (yellow arrows), FG-labeled neurons (white arrows) in the RMTg. e–g Immunofluorescence images of RMTg neurons showing FG-labeled RMTg neurons (e, magenta), CTβ-labeled RMTg neurons (f, green), and a merged image (g). White arrows indicate double labeling (e–g). IPN interpeduncular nucleus. Scale bars 400 μm in a–c; 100 μm in d and e
Fig. 10.
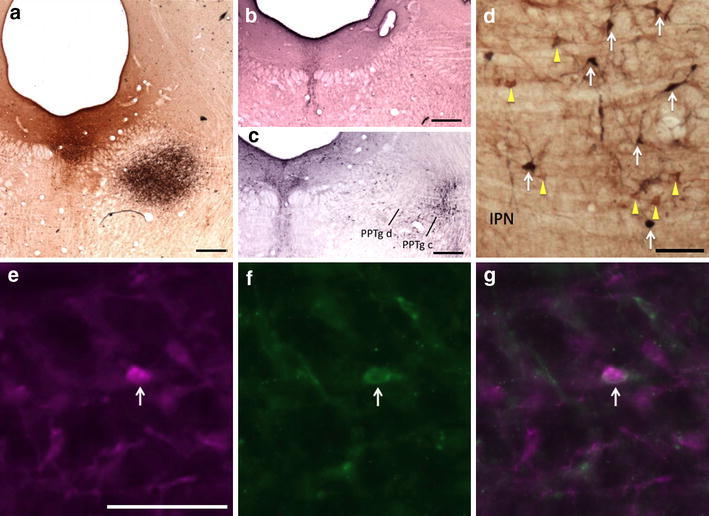
Retrogradely labeled neurons in the RMTg following injections of CTβ in the DR and FG in the PPTg. a FG injection site in the PPTg (black); CTβ injection site in the DR (brown). b Adjacent section processed for 5-HT immunoreactivity (black) verifies that the brown injection shown in a is in the DR. c Adjacent section processed for Nos immunoreactivity (black) verifies that the black injection shown in a is in the PPTg, pars dissipata. d CTβ-labeled neurons (yellow arrows) and FG-labeled neurons (white arrows) in the RMTg. e–g Immunofluorescence images of RMTg neurons showing FG-labeled RMTg neurons (e, magenta), CTβ-labeled RMTg neurons (f, green), and a merged image (g). White arrows indicate double labeling (e–g). PPTg c pedunculopontine tegmental nucleus, pars compacta; PPTg d pedunculopontine tegmental nucleus, pars dissipata; IPN interpeduncular nucleus. Scale bars 400 μm in a–c; 100 μm in d and e
Discussion
Neuroanatomical characterization of the RMTg has revealed that it is a GABAergic structure with dense afferent inputs from the LHb and SN and outputs to the VTA/SNC and other brainstem structures, including the PPTg, DR, and RtGi (Jhou et al. 2009b). The present study was undertaken to identify if RMTg neurons that project to the PPTg and DR also express Fos upon acute psychostimulant drug administration, as do RMTg neurons that project to the VTA/SNC (Geisler et al. 2008; Jhou et al. 2009b). VTA-projecting RMTg neurons that express Fos in response to psychostimulant administration are concentrated within the dense focus of Fos-expressing neurons that has come to be recognized as the RMTg per se, but such neurons were also present in the region immediately surrounding the RMTg, diminishing in number with increasing distance from the RMTg (Geisler et al. 2008). Presumably, RMTg neurons projecting to the PPTg and DR and expressing Fos after psychostimulants could be similarly distributed. Accordingly, double labeling in the RMTg was contrasted in the present study with that in the immediately surrounding region in order to emphasize the distinct character of the RMTg (Fig. 7).
Our current results for the VTA projections replicate those of our earlier study (Geisler et al. 2008) indicating that 48% of RMTg neurons that project to the VTA, as shown by retrograde labeling, express Fos following administration of methamphetamine (10 mg/kg), whereas only 13% of VTA-projecting neurons expressed Fos in the region just outside the RMTg. Also consistent with the previous study, the abundance of retrogradely labeled, Fos-expressing neurons in the rest of brainstem was negligible by comparison, although this was not objectively quantitated as was done in the previous study (Geisler et al. 2008). In turn, we found that 56% of RMTg neurons that project to the PPTg pars dissipata expressed Fos after methamphetamine, compared with 19% of PPTg-projecting neurons just outside the RMTg. Similarly, 47% of RMTg neurons that project to the DR expressed Fos, compared with 9% of DR-projecting neurons just outside the RMTg. The results for injections into the RtGi were surprising, however, with only 8% of retrogradely labeled RMTg neurons expressing Fos, compared with 15% of RtGi-projecting neurons just outside the RMTg. Overall, these results indicate that neurons in the RMTg that express Fos following methamphetamine administration project to the VTA, PPTg pars dissipata, and DR, but not substantially to the RtGi. In view of these results, we speculated that single neurons in the RMTg may project to more than one of these structures. Indeed, when the VTA and PPTg were injected in the present study with different retrograde tracers, 9% of retrogradely labeled neurons in the RMTg carried both tracers. When the VTA and DR were injected with different retrograde tracers, 10% of the retrogradely labeled neurons in the RMTg were double labeled. When the PPTg and DR were injected with different retrograde tracers, 5% of retrogradely labeled neurons in the RMTg carried both tracers. These results indicate that a significant number of RMTg neurons project to multiple targets.
RMTg projections to the PPTg and DR have received little consideration by comparison with those to the VTA/SNC. The PPTg has cholinergic (Oakman et al. 1995; Omelchenko and Sesack 2006), glutamatergic (Charara et al. 1996; Clements and Grant 1990; Geisler et al. 2007), and GABAergic (Charara et al. 1996; Jia et al. 2003; Hui-Ling and Morales 2009) projections to the VTA, and the data reported herein indicate that some or all of these are likely subject to RMTg modulation. It is well established that the VTA plays a prominent role in the initiation of behavioral sensitization (Vanderschuren and Kalivas 2000; Colussi-Mas et al. 2007) and increasing evidence implicates PPTg involvement as well (Alderson et al. 2003; Nelson et al. 2007). The present data indicate that the RMTg is also likely to influence the initiation of sensitization insofar as it appears anatomically positioned to coordinately regulate both the VTA/SNC and PPTg. As single neurons in the RMTg project to both the VTA and the PPTg, it seems reasonable to speculate that the RMTg may serve to coordinate activity in these structures. However, an alternative possibility exists that RMTg projections to the PPTg may exert less influence on VTA-projecting PPTg neurons than on PPTg neurons with other functional consequences, such as, e.g., modulation of spinal motor neuron excitability (Scarnati et al. 2011).
Neurons of the DR are primarily serotoninergic (Aghajanian et al. 1967; Baker et al. 1991; Molliver 1987), and, like the VTA/SNC (Christoph et al. 1986; Jhou et al. 2009a; Matsumoto and Hikosaka 2007) are inhibited by stimulation of LHb (Stern et al. 1979), as is release of serotonin by ascending projections from the raphe (Nishikawa and Scatton 1985). Due to the strong neuronal projections from the RMTg to the DR, it seems likely that the RMTg acts to modulate the signal from the LHb to the DR (Herkenham and Nauta 1979). Hikosaka (2010) has suggested that DR neurons may monitor the stressfulness of situations in the service of developing appropriate behavioral responses. The RMTg may convey information integrated from a variety of different descending pathways (Jhou et al. 2009b; Kaufling et al. 2009), and conduct it to the DR in order to facilitate the synthesis of appropriate responses by that structure. As was observed for RMTg projections to the PPTg, single RMTg neurons project both to the DR and VTA, suggesting again that single RMTg neurons may coordinate utilization of multiple neurotransmitters in several structures.
Despite convincing evidence from PHA-L injections that the RtGi is a target of strong RMTg projection (Jhou et al. 2009b), the present results shed doubt on this conclusion. The percentage of retrogradely labeled neurons that expressed Fos in the RMTg following CTβ injections into the RtGi was no different than in areas surrounding the RMTg and no focal accumulation of retrogradely labeled neurons in the RMTg was observed. To the contrary, retrogradely labeled neurons were relatively dense following these injections in much of the midbrain tegmentum including the retrorubral field, which is known to give rise to a robust descending projection to the vicinity of the RtGi (von Krosigk and Smith 1991; von Krosigk et al. 1992). Presumably in the earlier study (Jhou et al. 2009b), PHA-L labeled these descending projections or ones akin to them. The present result leads to the idea that the RMTg may be more involved in the modulation of structures that give rise to strong ascending projections, such as the VTA/SNC, PPTg, and DR, than in structures that have mainly descending outputs, such as the RtGi.
Recent studies of the RMTg demonstrate that it is activated and exhibits Fos expression in response to aversive stimuli or reward omission and inhibited by rewards and reward-predictive stimuli (Jhou et al. 2009a; Hong et al. 2011). RMTg neurons also express Fos in response to acute and repeated exposure to psychostimulants (Colussi-Mas et al. 2007; Geisler et al. 2008; Jhou et al. 2009a, b; Kaufling et al. 2010a, b; Perrotti et al. 2005; Scammell et al. 2000), but this does not necessarily indicate that psychostimulants activate RMTg neurons (Zahm et al. 2010). To the contrary, Lecca et al. (2011) have shown the inhibition of RMTg neurons in response to psychostimulant administration in which case the Fos expression can be better explained as accompanying transcriptional regulation associated with neuronal inhibition. Alternatively, systemically administered psychostimulants may produce an initial depression of RMTg neuronal activity (associated with disinhibition of the DA neurons, DA release in the Acb, and euphoria) followed by a rebound excitation, during which Fos is expressed, that would depress DA release and be aversive (TC Jhou, personal communication). This idea is consistent with the observation that both appetitive and aversive sensations commonly accompany administration of psychostimulant drugs of abuse (Hutchison and Riley 2008; Lecca et al. 2011; Simpson and Riley 2005; Wise et al. 1976). Our evidence for single RMTg neurons projecting to multiple structures involved in the regulation of VTA/SNC neuronal activity and thus DA release supports this idea. This system of brainstem connections warrants further consideration when one thinks about how appetitively and aversively affective stimuli enlist opponent processes that underlie adaptive responses to environmental stimuli and maladaptive ones following repeated exposure to drugs of abuse (Daw et al. 2002; Fields 2007; Koob and Le Moal 2008; Hikosaka 2010; Jhou et al. 2009a, b; Matsumoto and Hikosaka 2007).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
References
- Aghajanian GK, Rosecrans JA, Sheard MH. Serotonin: release in the forebrain by stimulation of midbrain raphe. Science. 1967;3773:402–403. doi: 10.1126/science.156.3773.402. [DOI] [PubMed] [Google Scholar]
- Alderson HL, Faulconbridge LF, Gregory LP, Latimer MP, Winn P. Behavioural sensitisation to repeated d-amphetamine: effects of excitotoxic lesions of the pedunculopontine tegmental nucleus. Neuroscience. 2003;118:311–315. doi: 10.1016/S0306-4522(03)00152-0. [DOI] [PubMed] [Google Scholar]
- Araki M, McGeer PL, Kimura H. The efferent projections of the rat lateral habenular nucleus revealed by the PHA-L anterograde tracing method. Brain Res. 1988;441:319–330. doi: 10.1016/0006-8993(88)91410-2. [DOI] [PubMed] [Google Scholar]
- Armstrong DM, Saper CB, Levey AI, Wainer BH, Terry RD. Distribution of cholinergic neurons in rat brain: demonstrated by the immunocytochemical localization of choline acetyltransferase. J Comp Neurol. 1983;216(1):53–68. doi: 10.1002/cne.902160106. [DOI] [PubMed] [Google Scholar]
- Baker KG, Halliday GM, Hornung JP, Geffen LB, Cotton RG, Törk I. Distribution, morphology and number of monoamine-synthesizing and substance P-containing neurons in the human dorsal raphe nucleus. Neuroscience. 1991;42(3):757–775. doi: 10.1016/0306-4522(91)90043-N. [DOI] [PubMed] [Google Scholar]
- Bjijou Y, Stinus L, Le Moal M, Cador M. Evidence for selective involvement of dopamine D1 receptors of the ventral tegmental area in the behavioral sensitization induced by intra-ventral tegmental area injections of d-amphetamine. J Pharmacol Exp Ther. 1996;277(2):1177–1187. [PubMed] [Google Scholar]
- Brinschwitz K, Dittgen A, Madai VI, Lommel R, Geisler S, Veh W. Glutamatergic axons from the lateral habenula mainly terminate on GABAergic neurons of the ventral midbrain. Neuroscience. 2010;168:463–476. doi: 10.1016/j.neuroscience.2010.03.050. [DOI] [PubMed] [Google Scholar]
- Bromberg-Martin ES, Matsumoto M, Hikosaka O. Dopamine in motivational control: rewarding, aversive, and alerting. Neuron. 2010;68:815–834. doi: 10.1016/j.neuron.2010.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cador M, Bjijou Y, Caihol S, Stinus L. d-Amphetamine-induced behavioral sensitization: implication of a glutamatergic medial prefrontal cortex-ventral tegmental area innervation. Neuroscience. 1995;94(3):705–721. doi: 10.1016/S0306-4522(99)00361-9. [DOI] [PubMed] [Google Scholar]
- Cador M, Bjijou Y, Stinus L. Evidence of a complete independence of the neurobiological substrates for the induction and expression of behavioral sensitization to amphetamine. Neuroscience. 1999;65(2):385–395. doi: 10.1016/0306-4522(94)00524-9. [DOI] [PubMed] [Google Scholar]
- Charara A, Smith Y, Parent A. Glutamatergic inputs from the pedunculopontine nucleus to midbrain dopaminergic neurons in primates: phaseolus vulgaris-leucoagglutinin anterograde labeling combined with postembedding glutamate and GABA immunohistochemistry. J Comp Neurol. 1996;364:254–266. doi: 10.1002/(SICI)1096-9861(19960108)364:2<254::AID-CNE5>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- Chou TC, Baxter MG, Saper CB (2004) A novel afferent to dopaminergic neurons regulates fear-induced freezing. Soc Neurosci Abstr 30:783.13
- Christoph GR, Leonzio RJ, Wilcox KS. Stimulation of the lateral habenula inhibits dopamine-containing neurons in the substantia nigra and ventral tegmental area of the rat. J Neurosci. 1986;6:613–619. doi: 10.1523/JNEUROSCI.06-03-00613.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements JR, Grant S. Glutamate-like immunoreactivity in neurons of the laterodorsal tegmental and pedunculopontine nuclei in the rat. Neurosci Lett. 1990;120:70–73. doi: 10.1016/0304-3940(90)90170-E. [DOI] [PubMed] [Google Scholar]
- Colussi-Mas J, Geisler S, Zimmer L, Zahm DS, Bérod A. Activation of afferents to the ventral tegmental area in response to acute amphetamine: a double-labeling study. Eur J Neurosci. 2007;26:1011–1025. doi: 10.1111/j.1460-9568.2007.05738.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahlstrom A, Fuxe K. Localization of monoamines in the lower brain stem. Experientia. 1964;20:398–399. doi: 10.1007/BF02147990. [DOI] [PubMed] [Google Scholar]
- Daw ND, Kakade S, Dayan P. Opponent interactions between serotonin and dopamine. Neural Netw. 2002;15:603–616. doi: 10.1016/S0893-6080(02)00052-7. [DOI] [PubMed] [Google Scholar]
- Fields HL. Understanding how opioids contribute to reward and analgesia. Reg Anesth Pain Med. 2007;32:242–246. doi: 10.1016/j.rapm.2007.01.001. [DOI] [PubMed] [Google Scholar]
- Gao DM, Huffman D, Benabid AL. Simultaneous recording of spontaneous activities and nociceptive responses from neurons in the pars compacta of substantia nigra and in the lateral habenula. Eur J Neurosci. 1996;8(7):1474–1478. doi: 10.1111/j.1460-9568.1996.tb01609.x. [DOI] [PubMed] [Google Scholar]
- Geisler S, Derst C, Veh RW, Zahm DS. Glutamatergic afferents of the ventral tegmental area in the rat. J Neurosci. 2007;27:5730–5743. doi: 10.1523/JNEUROSCI.0012-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geisler S, Marinelli M, Degarmo B, Becker ML, Freiman AJ, Beales M, Meredith GE, Zahm DS. Prominent activation of brainstem and pallidal afferents of the ventral tegmental area by cocaine. Neuropsychopharmacology. 2008;33:2688–2700. doi: 10.1038/sj.npp.1301650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Nauta WJ. Efferent connections of the habenular nuclei in the rat. J Comp Neurol. 1979;187:19–47. doi: 10.1002/cne.901870103. [DOI] [PubMed] [Google Scholar]
- Hikosaka O. The habenula: from stress evasion to value-based decision-making. Nat Rev Neurosci. 2010;11:503–513. doi: 10.1038/nrn2866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong S, Jhou TC, Smith MK, Saleem KS, Hikosaka O. Negative reward signals from the lateral habenula to dopamine neurons are mediated by rostromedial tegmental nucleus in primates. J Neurosci. 2011;31:11457–11471. doi: 10.1523/JNEUROSCI.1384-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hooks MS, Jones HG, Liem BJ, Justice JB., Jr Sensitization and individual differences to IP amphetamine, cocaine, or caffeine following repeated intracranial amphetamine infusions. Pharmacol Biochem Behav. 1992;43(3):815–823. doi: 10.1016/0091-3057(92)90413-A. [DOI] [PubMed] [Google Scholar]
- Hui-Ling W, Morales M. Pedunculopontine and laterodorsal tegmental nuclei contain distinct populations of cholinergic, glutamatergic, and GABAergic neurons in the rat. Eur J Neurosci. 2009;29(2):340–358. doi: 10.1111/j.1460-9568.2008.06576.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchison MA, Riley AL. Adolescent exposure to nicotine alters the aversive effects of cocaine in adult rats. Neurotoxicol Teratol. 2008;30:404–411. doi: 10.1016/j.ntt.2008.04.004. [DOI] [PubMed] [Google Scholar]
- Jhou TC and Gallagher M (2007) Paramedian raphe neurons that project to midbrain dopamine neurons are activated by aversive stimuli. Soc Neurosci Abstr 33:425.5
- Jhou TC, Fields HL, Baxter MG, Saper CB, Holland PC. The rostromedial tegmental nucleus (RMTg), a GABAergic afferent to midbrain dopamine neurons, encodes aversive stimuli and inhibits motor responses. Neuron. 2009;61:786–800. doi: 10.1016/j.neuron.2009.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jhou TC, Geisler S, Marinelli M, Degarmo BA, Zahm DS. The mesopontine rostromedial tegmental nucleus: a structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta. J Comp Neurol. 2009;513:566–596. doi: 10.1002/cne.21891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji H, Shepard PD. Lateral habenula stimulation inhibits rat midbrain dopamine neurons through a GABA(A) receptor-mediated mechanism. J Neurosci. 2007;27:6923–6930. doi: 10.1523/JNEUROSCI.0958-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia H-G, Yamuy J, Sampogna S, Morales FR, Chase MH. Colocalization of γ-aminobutyric acid and acetylcholine in neurons in the laterodorsal and pedunculopontine tegmental nuclei in the cat: a light and electron microscopic study. Brain Res. 2003;992(2):205–219. doi: 10.1016/j.brainres.2003.08.062. [DOI] [PubMed] [Google Scholar]
- Kalivas PW, Stewart J. Dopamine transmission in the initiation and expression of drug- and stress-induced sensitization of motor activity. Brain Res Rev. 1991;16(3):223–244. doi: 10.1016/0165-0173(91)90007-U. [DOI] [PubMed] [Google Scholar]
- Kalivas PW, Webber B. Amphetamine injection into the ventral mesencephalon sensitizes rats to peripheral amphetamine and cocaine. J Pharmacol Exp Ther. 1988;245:1095–1102. [PubMed] [Google Scholar]
- Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M. Afferents to the GABAergic tail of the ventral tegmental area in the rat. J Comp Neurol. 2009;513:597–621. doi: 10.1002/cne.21983. [DOI] [PubMed] [Google Scholar]
- Kaufling J, Veinante P, Pawlowski SA, Freund-Mercier MJ, Barrot M. γ-Aminobyturic acid cells with cocaine-induced ΔFosB in the ventral tegmental area innervate mesolimbic neurons. Biol Psych. 2010;67(1):88–92. doi: 10.1016/j.biopsych.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Kaufling J, Waltisperger E, Bourdy R, Valera A, Veinante P, et al. Pharmacological recruitment of the GABAergic tail of the ventral tegmental area by acute drug exposure. Br J Pharmacol. 2010;161:1677–1691. doi: 10.1111/j.1476-5381.2010.00984.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF, Le Moal M. Addiction and the brain antireward system. Annu Rev Psychol. 2008;59:29–53. doi: 10.1146/annurev.psych.59.103006.093548. [DOI] [PubMed] [Google Scholar]
- Lecca S, Melis M, Luchicchi A, Ennas MG, Castelli MP, Muntoni AL, Pistis M. Effects of drugs of abuse on putative rostromedial tegmental neurons, inhibitory afferents to midbrain dopamine cells. Neuropsychopharmacology. 2011;36:589–602. doi: 10.1038/npp.2010.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto M, Hikosaka O. Lateral habenula as a source of negative reward signals in dopamine neurons. Nature. 2007;447:1111–1115. doi: 10.1038/nature05860. [DOI] [PubMed] [Google Scholar]
- Molliver M. Serotonergic neuronal systems: what their anatomic organization tells us about function. J Clin Psychopharmacol. 1987;7:3S–23S. doi: 10.1097/00004714-198712001-00002. [DOI] [PubMed] [Google Scholar]
- Nelson CL, Wetter JB, Milovanovic M, Wolf ME. The laterodorsal tegmentum contributes to behavioral sensitization to amphetamine. Neuroscience. 2007;146:41–49. doi: 10.1016/j.neuroscience.2007.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishikawa T, Scatton B. Inhibitory influence of GABA on central serotonergic transmission. Involvement of the habenulo-raphe pathways in the GABAergic inhibition of ascending cerebral serotonergic neurons. Brain Res. 1985;331:81–90. doi: 10.1016/0006-8993(85)90717-6. [DOI] [PubMed] [Google Scholar]
- Oakman SA, Faris PL, Kerr PE, Cozzari C, Hartman BK. Distribution of pontomesencephalic cholinergic neurons projecting to substantia nigra differs significantly from those projecting to ventral tegmental area. J Neurosci. 1995;15:5859–5869. doi: 10.1523/JNEUROSCI.15-09-05859.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omelchenko N, Sesack SR. Cholinergic axons in the rat ventral tegmental area synapse preferentially onto mesoaccumbens dopamine neurons. J Comp Neurol. 2006;494:863–875. doi: 10.1002/cne.20852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omelchenko N, Bell R, Sesack SR. Lateral habenula projections to dopamine and GABA neurons in the rat ventral tegmental area. Eur J Neurosci. 2009;30:1239–1250. doi: 10.1111/j.1460-9568.2009.06924.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrotti LI, Bolanos CA, Choi KH, Russo SJ, Edwards S, Ulery PG, et al. DeltaFosB accumulates in a GABAergic cell population in the posterior tail of the ventral tegmental area after psychostimulant treatment. Eur J Neurosci. 2005;21:2817–2824. doi: 10.1111/j.1460-9568.2005.04110.x. [DOI] [PubMed] [Google Scholar]
- Perugini M, Vezina P. Amphetamine administered to the ventral tegmental area sensitizes rats to the locomotor effects of nucleus accumbens amphetamine. J Pharmacol Exp Ther. 1994;270(2):690–696. [PubMed] [Google Scholar]
- Scammell TE, Estabrooke IV, McCarthy MT, Chemelli RM, Yanagi-sawa M, Miller MS, Saper CB. Hypothalamic arousal regions are activated during modafinil-induced wakefulness. J Neurosci. 2000;20:8620–8628. doi: 10.1523/JNEUROSCI.20-22-08620.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scarnati E, Florio T, Capozzo A, Confalone G, Mazzone P (2011) The pedunculopontine tegmental nucleus: implications for a role in modulating spinal cord motoneuron excitability. J Neural Transm 118:1409–1421 [DOI] [PubMed]
- Schultz W. Predictive reward signal of dopamine neurons. J Neurophysiol. 1998;80:1–27. doi: 10.1152/jn.1998.80.1.1. [DOI] [PubMed] [Google Scholar]
- Schultz W. Multiple dopamine functions at different time courses. Annu Rev Neurosci. 2007;30:259–288. doi: 10.1146/annurev.neuro.28.061604.135722. [DOI] [PubMed] [Google Scholar]
- Shumake J, Ilango A, Scheich H, Wetzel W, Ohl FW. Differential neuromodulation of acquisition and retrieval of avoidance learning by the lateral habenula and ventral tegmental area. J Neurosci. 2010;30:5876–5883. doi: 10.1523/JNEUROSCI.3604-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson GR, Riley AL. Morphine preexposure facilitates morphine place preference and attenuates morphine taste aversion. Pharmacol Biochem Behav. 2005;80:471–479. doi: 10.1016/j.pbb.2005.01.003. [DOI] [PubMed] [Google Scholar]
- Stern WC, Johnson A, Bronzino JD, Morgane PJ. Effects of electrical stimulation of the lateral habenula on single-unit activity of raphe neurons. Exp Neurol. 1979;65:326–342. doi: 10.1016/0014-4886(79)90102-X. [DOI] [PubMed] [Google Scholar]
- Vanderschuren LJ, Kalivas PW. Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology. 2000;151:99–120. doi: 10.1007/s002130000493. [DOI] [PubMed] [Google Scholar]
- Vezina P. D1 dopamine receptor activation is necessary for the induction of sensitization by amphetamine in the ventral tegmental area. J Neurosci. 1996;16(7):2411–2420. doi: 10.1523/JNEUROSCI.16-07-02411.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Krosigk M, Smith AD. Descending projections from the substantia nigra and retrorubral field to the medullary and pontomedullary reticular formation. Eur J Neurosci. 1991;3:260–273. doi: 10.1111/j.1460-9568.1991.tb00088.x. [DOI] [PubMed] [Google Scholar]
- von Krosigk M, Smith Y, Bolam JP, Smith AD. Synaptic organization of GABAergic inputs from the striatum and the globus pallidus onto neurons in the substantia nigra and retrorubral field which project to the medullary reticular formation. Neuroscience. 1992;50:531–549. doi: 10.1016/0306-4522(92)90445-8. [DOI] [PubMed] [Google Scholar]
- Wise RA, Yokel RA, DeWit H. Both positive reinforcement and conditioned aversion from amphetamine and from apomorphine in rats. Science. 1976;191:1273–1275. doi: 10.1126/science.1257748. [DOI] [PubMed] [Google Scholar]
- Zahm DS, Becker ML, Freiman AJ, Strauch S, Degarmo B, Geisler S, Meredith GE, Marinelli M. Fos after single and repeated self-administration of cocaine and saline in the rat: emphasis on the basal forebrain and recalibration of expression. Neuropsychopharmacology. 2010;35(2):445–463. doi: 10.1038/npp.2009.149. [DOI] [PMC free article] [PubMed] [Google Scholar]