

Abstract
Marijuana is a preparation of the flower, as well as the leaves and seeds, of the plant Cannabis sativa. Marijuana has been used for medicinal and recreational purposes for thousands of years due to its psychoactive effects including euphoria, sedation, and analgesia. Although it has been suspected for decades that marijuana has adverse effects on female fertility, the underlying molecular mechanism was not clear. The discovery of cannabinoid receptors and endocannabinoids has advanced studies if cannabinoid signaling. Since then, numerous studies have been published on cannabinoid signaling in female reproductive events, including preimplantation embryo development, oviductal embryo transport, embryo implantation, placentation, and parturition. This review focuses on various aspects of endocannabinoid signaling in female fertility.
Keywords: Cannabinoids, embryo, uterus, oviduct, implantation, placentation
I. Endocannabinoid System
Although marijuana has been used for thousands of years, research on cannabinoids did not begin until the discovery of the major psychoactive component of marijuana, Δ9-tetrahydrocannabinol (Δ9-THC).1 Later, cannabinoid-like compounds were identified in vivo, collectively named endocannabinoids. The two most studied endocannabinoids are N-arachidonoylethanolamine (also known as anandamide, AEA) and 2 arachidonoylglycerol (2-AG).2−4 Both AEA and 2-AG are produced on demand, although 2-AG levels are much higher than those of AEA.5 Δ9-THC and endocannabinoids exert their function by targeting two major G protein-coupled cannabinoid receptors, brain-type CNR1, encoded by Cnr1,6,7 and spleen-type CNR2, encoded by Cnr2.8 CNR1 is abundantly expressed in the central nervous system, but further studies showed that CNR1 is also present in other peripheral tissues, albeit at lower levels. In contrast, CNR2 is mostly expressed in immune cells.9−11 In addition to CNR1 and CNR2, it has also been shown that GPR5512,13 and the transient receptor potential vanilloid 1 (TRPV1)14 are capable of mediating some function of cannabinoids/endocannabinoids.
Levels of endocannabinoids in vivo are tightly regulated by enzymes for endocannabinoid synthesis and degradation. AEA is derived from the precursor N-acyl phosphatidylethanolamine (NAPE), but studies on knockout mice suggest that AEA can be generated via multiple enzymatic pathways,15,16 involving NAPE-hydrolyzing phospholipase D (Nape-pld),17,18 α/β-hydrolase 4 (Abh4),19 and a protein tyrosine phosphatase, Ptpn22.20 It is, however, widely accepted that degradation of AEA is mainly mediated by a membrane-bound fatty acid amide hydrolase (FAAH) to ethanolamine and arachidonic acid (AA)21,22 and that FAAH is critical for regulating both the magnitude and duration of AEA signaling.23 In addition, FAAH is capable of hydrolyzing other fatty acid amides, including 2-AG.
2-AG is produced by the cleavage of its precursor lipid molecule, diacylglycerol (DAG), by a membrane-bound sn1-diacylglycerol lipase (DAGL),24 which exists in vivo in two different isoforms, DAGLα and DAGLβ. 2-AG is degraded to AA and glycerol by either FAAH or a membrane-associated serine hydrolase, monoacylglycerol lipase (MAGL).25
The endocannabinoid system is not limited to the components described above, but other endogenous ligands as well as numerous synthetic agonists and antagonists can target cannabinoid receptors. These compounds can also orthosterically or allosterically interact with other non-CB receptors.26
II. Endocannabinoid Signaling in the Mouse Female Reproductive System
Fertilization, preimplantation embryo development, oviductal embryo transport, implantation, decidualization, placentation, and finally parturition are all critical events for the success of pregnancy. These events involve sperm, eggs, fertilized embryos, oviducts, and the uterus. This review discusses various aspects of the endocannabinoid system from preimplantation embryo development to parturition, in the context of the uterine environment.
Endocannabinoid Signaling in Fertilization and Preimplantation Embryo Development
Fertilization occurs when a sperm fuses with an egg in the oviduct;27,28 the fertilized egg is now called an embryo. After fertilization, the embryo undergoes several rounds of successive mitotic cell divisions spanning a 2-cell through 16-cell stage, ultimately forming a ball of compacted cells termed morula. At the morula stage, the embryo starts migrating toward the uterine lumen. As the morula gradually moves through the uterotubal junction and enters into the uterus, a small cavity (blastocoel) starts appearing within the embryo, marking the beginning of the blastocyst stage. A blastocyst consists of the outer layer of trophectoderm (TE) cells and the inner cell mass (ICM).29,30
In preimplantation mouse embryos, CNR1 expression is present from late 2-cell stage to blastocyst stage. In the blastocyst, CNR1 is primarily expressed in the trophectoderm. In contrast, CNR2 is expressed in embryos from 1-cell through the blastocyst stage, but in the latter stage, CNR2 expression is primarily restricted to the ICM.31 In the oviduct and uterus, only CNR1, but not CNR2, is detected.32,33 In addition to the presence of these two receptors, FAAH and NAPE-PLD are also present in the preimplantation embryos from the 2-cell stage to the blastocyst stage. FAAH is mainly detected in the outer cell layers of morula and in the trophectoderm of blastocysts.
On-time preimplantation embryo development is critical to achieving an implantation-competent blastocyst, and it requires finely tuned endocannabinoid signaling. For example, preimplantation embryo development becomes derailed with higher levels of cannabinoid/endocannabinoid signaling. The development of 2-cell embryos to blastocysts was shown to be arrested at high levels of AEA, 2-AG, Δ9-THC, or a synthetic cannabinoid agonist WIN55212-2.31,34 Moreover, blastocysts treated with high levels of AEA had a reduced number of TE cells, and blastocyst zona-hatching became sluggish.35,36 The adverse effects of high level cannabinoid/endocannabinoid signaling was attenuated by SR141716A and AM251 (CNR1 selective antagonists), but not by SR144528 (a CNR2 specific antagonist). These results were further confirmed by the observation that a CNR2 agonist, AM663, failed to affect embryo development.34 Collectively, higher levels of cannabinoid signaling via CNR1 inhibit normal embryo development.
Complete sequestration of CNR1-mediated signaling causes developmental arrest of embryos. The generation of Cnr1 and Cnr2 null mice provided useful tools to study the effects of loss-function of cannabinoid signaling. Cnr1–/– and Cnr1–/–/Cnr2–/– females were mated with males of the same genotype, and embryos from their oviduct/uteri were collected and examined. The null embryos, recovered from oviducts on day 3 and from uteri on day 4 of pregnancy, showed retarded growth compared with embryos derived from wild-type (WT) females.37 This abnormal embryo development may be the consequence of oviductal and/or embryonic Cnr1 deficiency. Observations that heterozygous embryos recovered from Cnr1–/– females mated with WT males showed normal embryo development33 indicating that embryonic CNR1, but not maternal CNR1, directs appropriate embryo development.38 These findings suggest that either silenced or augmented CNR1 signaling leads to abnormal embryo development. These results provide evidence that tightly regulated levels of cannabinoid/endocannabinoid signaling under normal physiological conditions is conducive to normal development. This was further supported by the fact that trophoblast outgrowth in vitro was promoted at a low level (7 nM) but became retarded at higher levels (28 nM) of AEA.39
Although Cnr2–/– embryos showed some aberrant development,37 CNR2’s localization in the ICM,40 but not TE cells,41 suggests that CNR2-mediated cannabinoid signaling has a different biological function compared to that mediated by CNR1. Thus, it is possible that CNR2 plays a role in ICM development.
Collectively, normal preimplantation embryo development requires appropriate endocannabinoid signaling. The growth and differentiation of TE cells is regulated by CNR1 mediated cannabinoid/endocannabinoid signaling.
Endocannabinoid Signaling and Oviductal–Uterine Embryo Transport
In mice, embryos develop within the oviduct until day 3 of pregnancy. Thereafter, embryos at the late morula stage start migrating toward the uterine lumen, forming blastocysts therein on day 4. Coordinated oscillation of the cilia on the oviductal epithelia and muscle contractions are required for the normal passage through the oviduct into the uterine lumen. Failure of this passage through the oviductal–uterine junction results in embryo retention. Preimplantation embryos thus retained within the oviduct can transform into blastocysts and remain viable for some time. In humans, the entrapped embryos can implant in the fallopian tube, and leads to tubal pregnancy in women, which is fatal in many occasions.42,43 Therefore, normal and timely oviductal transport of the embryo is critical to on-time and normal implantation.
In the mouse oviduct, mRNA of Cnr1, but not Cnr2, is detected, and levels of AEA are higher than those in the mouse brain.44 Notably, Faah levels are lower in the oviduct isthmus compared to levels in the ampullary region, whereas Nape-pld shows the reverse pattern, being higher in the isthmus.44,45 These observations suggest that there is a gradient of AEA levels in the oviduct.
Normal oviductal transport of embryos is disturbed in the absence of maternal CNR1. Studies using Cnr1–/– females mated with WT males showed that fewer embryos were recovered from Cnr1–/– uteri on the morning of day 4 of pregnancy. Given the fact that Cnr1+/– embryos have normal preimplantation development in Cnr1–/– oviducts, it suggests that the failure of embryos to enter the uterine lumen is related to the absence of Cnr1.33,37 In addition, the observation that Cnr1–/–/Cnr2–/–, but not WT or Cnr2–/–, mice also have similar oviductal retention of embryos suggests that normal oviductal transport requires CNR1 but not CNR2. The fact that Cnr1+/– embryos are morphologically healthy in Cnr1–/– uteri on day 4 of pregnancy indicates that maternal Cnr1 deficiency causes embryo retention. Reciprocal embryo transfer experiments further confirmed that CNR1 is critical for appropriate transport of embryos through the oviduct; only Cnr1–/– recipients displayed oviductal retention of embryos, irrespective of embryonic genotypes.33 Additionally, normal oviductal transport in WT mice was disrupted by treatment with a CNR1 specific blocker, while CNR2 antagonists had no effect.33 Higher cannabinoid signaling also affects oviductal embryo transport. Faah–/– mice with higher oviductal AEA levels than those in WT oviducts have similar oviductal retention of embryos. Moreover, WT mice exposed to Δ9-THC or methanandamide (a stable AEA analogue) phenocopied Faah–/– mice with oviductal transport defects, which was rescued by CNR1 antagonist treatment.44
All these results lend additional support to the hypothesis that either higher or lower endocannabinoid signaling adversely affects female reproduction in mice since normal oviductal transportation is disrupted in either absence of cannabinoid signaling in Cnr1–/– mice or high cannabinoid signaling in Faah–/– mice. Previous studies have shown that it is the wave of oviductal muscle movement that sends the embryos into the uterus and that this movement is regulated by the sympathetic nervous system.46 Circular muscles relax upon activation of β2 adrenergic receptors (β2-AR), while they contract on activation of α1 adrenergic receptors (α1-AR). The alternant stimulation of these two receptors makes a wave of muscle movement, which propels embryos from the oviduct to the uterus. However, under exposure of the antagonist of either receptor, the oviduct cannot help move embryos to the uterus.46,47 The fact that CNR1 is colocalized with β2-AR/α1-AR indicates that endocannabinoid signaling may influence oviductal muscle movement through the regulation of adrenergic signaling. It was shown that oviductal adrenergic nerve terminals released more noradrenaline in Cnr1–/– than in WT females and that higher noradrenaline levels activated α1-AR to keep the smooth muscle in a contractile tone. This effect can be counterbalanced by administration of isoproterenol, a β2-AR agonist, which restored normal embryo transport in Cnr1–/– mice.33 In WT mice, treatment of methanandamide decreased the release of noradrenaline, and it was blocked by a CNR1 specific antagonist. Similarly, circular muscles maintained a relaxation phase in Faah–/– mice, where AEA levels were higher than in WT mice.44 In conclusion, either higher or silenced endocannabinoid signaling disrupts the balance between muscular contraction and relaxation, impairs normal wave movement through the oviduct, and consequently derails normal oviductal transport of embryos.
Endocannabinoid Signaling and Implantation
Normal embryo development and on time arrival in the uterus are all preparations for the next critical step in pregnancy: implantation. Implantation can occur only when the blastocyst becomes implantation-competent and the uterus achieves the receptive phase.48 A reciprocal interaction between the blastocyst and receptive uterus initiates the attachment of the blastocyst trophectoderm with the uterine luminal epithelium at night on day 4 of pregnancy.48−51 In mice, uterine receptivity can be achieved when the uterus is exposed to estrogen after 24–48 h of progesterone (P4) priming. The window of uterine receptivity is transient and lasts only for about 24 h. If implantation occurs beyond the normal window, there is an adverse ripple effect throughout the course of gestation, leading to a compromised pregnancy outcome. However, the gene network that confers uterine receptivity and on-time implantation is far from clear.
Endocannabinoids AEA and 2-AG, as well as their putative receptor CNR1, are present in the mouse uterus. Although the levels of 2-AG are about 200-fold higher than AEA in periimplantation uteri, both showed similar patterns with higher levels at the interimplantation site and lower levels at the implantation site. The expression patterns of NAPE-PLD and FAAH explain how the gradients of AEA and 2-AG are created. Higher levels of Nape-pld mRNA and NAPE-PLD activity are found in interimplantation sites, whereas both mRNA and protein levels are lower in implantation sites.45,52 In contrast, FAAH expression and activity are higher at implantation sites. This inverse relationship is evident for MAGL and DAGLα in that DAGLα levels are higher in interimplantation sites on days 5 and 7 of pregnancy where MAGL levels are low. Conversely, DAGLα levels are lower in implantation sites with higher levels of MAGL.52 Similar phenotype was also observed in humans in that FAAH expression is highest in midsecretory stroma, while levels of NAPE-PLD are lower in secretory glands.53 These results suggest that lower levels of AEA and CNR1 are beneficial for implantation.
It was shown that uteri in the receptive phase had lower levels of AEA compared with uteri in nonreceptive phase.36 In agreement with this result, higher levels of Nape-pld mRNA and NAPE-PLD activity was found in nonreceptive uteri, whereas higher levels of FAAH were observed in the receptive uteri.45,54 AEA levels are also critical in preparing embryos for implantation. High levels of AEA or other synthetic cannabinoids inhibited blastocyst zona-hatching, while low levels of AEA accelerated trophoblast differentiation and outgrowth in culture.31,34,36 To minimize exposure to endocannabinoid signaling, the expression of CNR1 is downregulated in the blastocyst approaching implantation. For example, WT blastocysts collected from the uterus on the morning of day 4 of pregnancy had higher levels of AEA binding compared with blastocysts recovered in the evening of day 4 prior to the attachment reaction, suggesting that regulated signaling by AEA is beneficial for the implantation competency of blastocysts.37 In the same vein, immunohistochemistry experiments showed that CNR1 expression was much lower in activated blastocysts than dormant blastocysts.37,55 In addition, some evidence suggests that the implanting blastocyst influences the uterine AEA levels in a reciprocal way. The implanting blastocysts decreased Nape-pld expression and upregulated Faah levels by releasing a lipid “FAAH activator”.45,54 In humans, women with elevated peripheral AEA levels also had a higher rate of spontaneous pregnancy loss.56,57 In addition, women who had undergone IVF/ICSI embryo transfer and had successful pregnancy showed significant decreases in plasma AEA levels on the day of embryo transfer compared with that of oocyte retrieval.58 All these results point toward one concept: that appropriate and regulated endocannabinoid signaling in both the blastocyst and uterus is required for the establishment of uterine receptivity and implantation competency of blastocysts.
The mechanism by which cannabinoid/endocannabinoid signaling regulates preimplantation embryo function is still not clear. Some evidence indicates that AEA induced stimulatory and inhibitory effects are mediated by ERK and Ca2+ signaling pathways in blastocysts. AEA at a low concentration (7 nM) activates ERK signaling via CNR1, whereas AEA at a higher concentration (28 nM) fails to activate ERK but instead inhibits Ca2+ mobilization.55 These results demonstrate that endocannabinoids at different concentrations are capable of regulating various signaling pathways to exert distinct biological functions.
The intensity of endocannabinoid signaling is tightly regulated by spatiotemporal expression patterns of the key synthetic/hydrolytic enzymes and cannabinoid receptors. This orchestration is essential since endocannabinoids at only normal physiological levels are conducive to early pregnancy events, including implantation. Gene expression studies and genetically engineered mouse models have shown that endocannabinoid signaling is one of the key players in directing the process of implantation, although the exact molecular signature of this signaling in embryo–uterine interaction is not clearly understood.
Endocannabinoid Signaling and Placentation
In mice, placentation begins once the peak phase of decidualization ended in order to establish maternal–fetal circulation. The embryonic trophectoderm, after making contact with the uterine stroma, forms primary trophoblast giant cells (TGC). The polar trophectoderm, adjacent to the ICM, forms the extraembryonic ectoderm and ectoplacental cone (EPC). The EPC and extraembryonic ectoderm cells are major contributors to the placenta. The placenta is the only connection between the mother and fetus in eutherians, and its functions include the exchange of oxygen, nutrients, and waste products between the two entities. Most exchanges occur in the labyrinth layer of the placenta, consisting of mainly syncytiotrophoblasts, which is derived from EPC and the ectoderm. The spongy layer is also derived from EPC and extraembryonic ectoderm cells and provides structural support for the labyrinth layer. Most cells in the spongy layer are spongiotrophoblast cells (SPT), which are separated from maternal cells by TGC cells.59 All of the TGC, SPT, syncytiotrophoblast, and other trophoblast cells are derived from trophoblast stem (TS) cells in the trophectoderm. To execute its physiological function, the placenta requires an appropriate population and distribution of different trophoblast cells. Any aberration in trophoblast differentiation compromises normal placentation.
The endocannabinoid system is present in midgestational placentas. Two endocannabinoids, AEA and 2-AG, are detected in murine placentas. It has been shown that AEA levels were elevated in Faah–/– placentas but not Cnr1–/– placentas, reinforcing the idea that FAAH is a major regulator of AEA status in mice. The placenta also has detectable levels of mRNAs for metabolic enzymes (Faah, Nape-pld, Magl, and Daglα) and cannabinoid receptors (Cnr1 and Trpv1). Immunohistochemistry results have provided evidence that FAAH and CNR1 are expressed in the EPC on day 10 of pregnancy and the SPT layer later on day 14.60 In addition, studies in various mammals, including rats and humans, indicate that the endocannabinoid system is present in those placentas.61−63 CNR1 and FAAH were identified in amniotic epithelial cells, chorionic cytotrophoblasts,61 and syncytiotrophoblasts62 in humans. To better understand the role of the endocannabinoid signaling in placentation, Cnr1 and Faah null mice were used.
Placentation was found to be compromised in Cnr1–/– females. Cnr1–/– females have a higher rate of embryo loss and lower placental weights on days 12 and 14 of pregnancy compared with those of WT mice. In Faah–/– females, the resorption rate starts to rise from day 14. These results suggest that both high and low endocannabinoid signaling compromises normal pregnancy events at midgestation, although the augmented signaling does not initiate pregnancy loss until late midgestation.60
It was found that in the absence of Cnr1, the development of the SPT layer is retarded. For example, the expression of Tpbpα, a marker gene of SPT, is much reduced in Cnr1–/– placentas on day 10 of pregnancy. However, Pl1 expression, a marker of TGC cells, is comparable in WT and Cnr1–/– placentas. Aberrant endocannabinoid signaling has adverse effects on the proliferation of trophoblast progenitor cells in the EPC, which later differentiate into SPT cells. Proliferation of Cnr1–/– trophoblast progenitor cells was much reduced, while Faah–/– trophoblast progenitor cells showed modest reduction in proliferation. These results are in agreement with results of in vitro experiments using TS cells. Proliferation of Cnr1–/–/Cnr2–/– and Faah–/– TS cells was remarkably sluggish compared to WT TS cells because of reduced activated AKT levels. Additionally, WT TS cells treated with methanandamide (7 nM) proliferated at a faster rate, but this was attenuated by a CNR1 selective antagonist, suggesting that CNR1 mediated endocannabinoid signaling regulates TS cell proliferation.60 The results show that CNR1 mediated signaling via the PI3K-AKT pathway is important for appropriate TS cell proliferation. It is interesting to note that AEA also prevented BeWo trophoblast cell proliferation in a dose-dependent manner in humans and that elevated plasma AEA levels also increased the risk factor for first trimester miscarriages.62
Interestingly, compromised development of the SPT layer is also attributed to the biased TS cell differentiation under aberrant endocannabinoid signaling. Secondary TGCs are differentiated from precursor cells present in the EPC and later in the SPT layer. Mash2 prevents trophoblast precursor cells from differentiating into secondary TGCs,64 whereas Hand1 promotes differentiation into TGCs.65Mash2 expression was shown to be much reduced in Faah–/– and Cnr1–/– placentas, indicating that Faah–/– and Cnr1–/– trophoblast precursor cells are more prone to differentiate into TGCs. These results were confirmed by the observation that the SPT marker Tpbpa was much lower in Cnr1–/–/Cnr2–/– than in WT TS cells after spontaneous differentiation of TS cells. Moreover, reciprocal embryo transfer experiments between Cnr1–/– and WT mice showed that it is the embryonic Cnr1 deficiency that causes this biased trophoblast cell differentiation.60 To further study the acute role of CNR1 during midgestation, WT females were treated with SR141716A on days 8 and 9 of pregnancy, when placentation is initiated. Further experiments showed that SR141716A treatment greatly reduced the number of SPT cells in WT placentas. In conclusion, regulated endocannabinoid signaling is required for maintaining appropriate TS cell proliferation: abnormal CNR1 signaling suppresses differentiation of TS cells into SPT cells but promotes differentiation to TGCs.
During placentation, some trophoblast cells invade the maternal decidual zone and reshape the maternal blood vessels to direct enough blood flow to support embryonic growth.66 Thus, trophoblast cell invasiveness is critical for successful placentation in establishing maternal–fetal circulation and exchanges. The invasion of glycogen trophoblast cells (GTC) into the decidua basalis is an indicator of trophoblast invasiveness. In WT females, an abundant number of GTC invaded into the decidual basalis; however, only a sporadic number of them with shallow invasion was found beyond the TGC border in Cnr1–/– mice.60 These results suggest that the invasive capacity of trophoblast cells is compromised in the absence of CNR1. Collectively, aberrant CNR1-mediated endocannabinoid signaling inhibits TS cell proliferation, differentiation into SPT cells, and trophoblast invasiveness, leading to defective placentation and pregnancy loss in the midgestational stage.
Endocannabinoid Signaling and Parturition
Parturition is the last encounter between the mother and fetus that leads to the birth of offspring. In humans, a pregnancy that lasts about 37 to 42 weeks is considered to be full term. A pregnancy with less than 37 weeks of gestational age is considered preterm, when a baby is not mature enough to allow normal postnatal survival. Preterm birth accounts for 10% of neonatal mortality worldwide.67 In developed countries, neonatal death decreases due to reduced infections and other causes of neonatal death, but the exact cause of preterm birth still remains unsolved. Therefore, preterm birth becomes the leading cause of neonatal mortality in developed countries and accounts for 25% of neonatal mortality in the U.S.68
It was observed that genetic loss of Cnr1, but not Cnr2, leads to preterm birth and lower birth weight. Previous reports showed that P4 withdrawal and a decreased P4 versus estradiol (E2) ratio determines the parturition timing in rodents.69 In Cnr1–/– mice, 17β-HSD, a key enzyme for estrogen biosynthesis, along with 20α-HSD, essential for metabolic conversion of P4 to an inactive metabolite, were substantially increased in Cnr1–/– ovaries, and consequently, the P4/E2 ratio decreased from day 16 of pregnancy. In fact, P4 supplement on day 18 of pregnancy restored normal parturition in Cnr1–/– mice, confirming that a decreased P4/E2 ratio is a cause of preterm labor in Cnr1–/– females.70 In summary, CNR1 deficiency induces preterm birth in mice by altering normal P4 and estrogen levels. Although the underlying causes of preterm birth remain largely unknown, endocannabinoid signaling may play a role in parturition events.
III. Closing Remarks
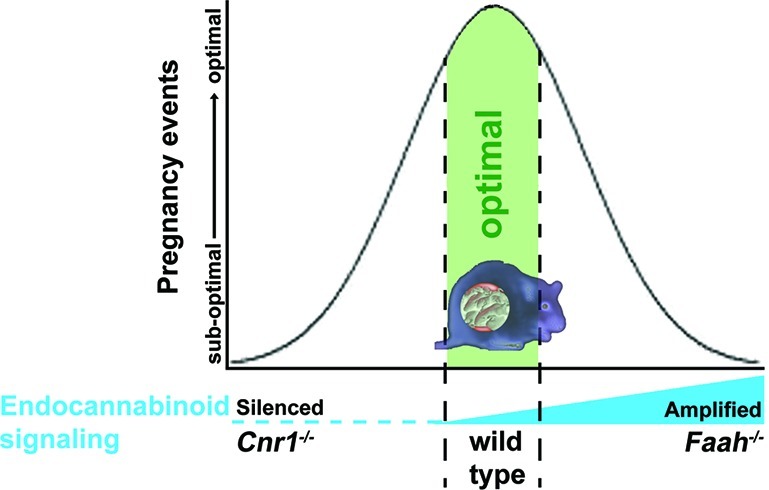
The endocannabinoid system is being discovered in many more organs, where it plays multiple physiological roles, depending on the organ. In female reproduction, the endocannabinoid system is found in different creatures from sea urchin to humans. Its conserved expression spanning different animals indicates that endocannabinoid signaling plays a key role in female reproduction. Studies in genetically manipulated mouse models showed that finely tuned endocannabinoid signaling in both the blastocyst and uterus is required for normal pregnancy events (Figure 1) since either silenced or elevated endocannabinoid signaling causes multiple defects during pregnancy (Table 1). Mouse models with the deletion of various genes within the endocannabinoid system are subfertile, suggesting that endocannabinoid signaling functions either as a regulatory rather than decisive determinant or that deficiency of this signaling is compensated for by other signaling pathways. Since genetic backgrounds and diets influence reproductive phenotypes in gene deleted mice,71 it would be interesting to see if the genetic background of mice and diet composition impact fertility phenotypes following a mutation of a gene in the endocannabinoid pathway. Nonetheless, almost all pregnancy events are regulated by endocannabinoid signaling, and in most cases, optimal outcomes can only be achieved under normal physiological endocannabinoid levels. Either silenced or enhanced endocannabinoid signaling derails these processes. This is a trait of endocannabinoid signaling that provides a route for medical manipulation in each pregnancy event. However, the in-depth mechanism by which endocannabinoid signaling exerts its role is still not clear. In addition, it would be interesting to study the mechanism behind how lower and higher cannabinoid signaling generates similar phenotypes. Further studies on endocannabinoid signaling with respect to female reproduction will generate new molecular mechanisms governing pregnancy events.
Figure 1.
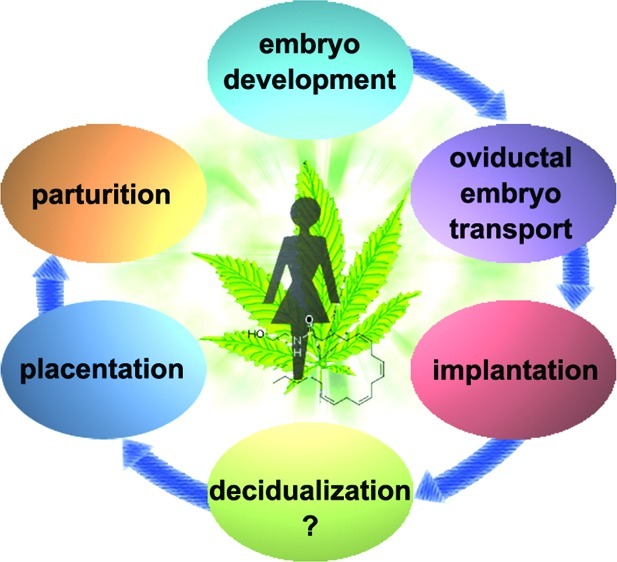
Cannabinoid/endocannabinoid signaling impacts various pregnancy events. Either silenced or augmented cannabinoid/endocannabinoid signaling disturbs pregnancy events including preimplantation embryo development, oviductal embryo transport, implantation, placentation, and parturition. The impact on decidualization is yet to be determined.
Table 1. Observed Adverse Effects of Either Silenced or Augmented Cannabinoid/Endocannabinoid Signaling during Pregnancy in Mice.
| cannabinoid
signaling |
|||
|---|---|---|---|
| silenced | optimal | augmented | |
| pregnancy events | Cnr1–/– | WT | Faah–/– |
| embryo development | retarded | normal | retarded |
| oviductal embryo transport | retention | on time | retention |
| blastocyst competency | conducive to competency | impediment to competency | |
| uterine receptivity | suboptimal | receptive | suboptimal |
| fetoplacental development | retarded/resorption | healthy | retarded/resorption |
| parturition | preterm | full term | |
Acknowledgments
We thank Serenity Curtis for editing the manuscript.
Work described in this review was supported by grants from NIH (DA006668). X.S. is supported by a Lalor Foundation postdoctoral fellowship.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
References
- Gaoni Y.; Mechoulam R. (1964) Isolation, structure, and partial synthesis of an active constituent of hashish. J. Am. Chem. Soc. 86, 1646–1647. [Google Scholar]
- Devane W. A.; Hanus L.; Breuer A.; Pertwee R. G.; Stevenson L. A.; Griffin G.; Gibson D.; Mandelbaum A.; Etinger A.; Mechoulam R. (1992) Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258, 1946–1949. [DOI] [PubMed] [Google Scholar]
- Mechoulam R.; Ben-Shabat S.; Hanus L.; Ligumsky M.; Kaminski N. E.; Schatz A. R.; Gopher A.; Almog S.; Martin B. R.; Compton D. R.; et al. (1995) Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 50, 83–90. [DOI] [PubMed] [Google Scholar]
- Sugiura T.; Kudo N.; Ojima T.; Mabuchi-Itoh K.; Yamashita A.; Waku K. (1995) Coenzyme A-dependent cleavage of membrane phospholipids in several rat tissues: ATP-independent acyl-CoA synthesis and the generation of lysophospholipids. Biochim. Biophys. Acta 1255, 167–176. [DOI] [PubMed] [Google Scholar]
- Sugiura T.; Kishimoto S.; Oka S.; Gokoh M. (2006) Biochemistry, pharmacology and physiology of 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand. Prog. Lipid Res. 45, 405–446. [DOI] [PubMed] [Google Scholar]
- Devane W. A.; Dysarz F. A. 3rd; Johnson M. R.; Melvin L. S.; Howlett A. C. (1988) Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 34, 605–613. [PubMed] [Google Scholar]
- Matsuda L. A.; Lolait S. J.; Brownstein M. J.; Young A. C.; Bonner T. I. (1990) Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 346, 561–564. [DOI] [PubMed] [Google Scholar]
- Munro S.; Thomas K. L.; Abu-Shaar M. (1993) Molecular characterization of a peripheral receptor for cannabinoids. Nature 365, 61–65. [DOI] [PubMed] [Google Scholar]
- Howlett A. C.; Barth F.; Bonner T. I.; Cabral G.; Casellas P.; Devane W. A.; Felder C. C.; Herkenham M.; Mackie K.; Martin B. R.; Mechoulam R.; Pertwee R. G. (2002) International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 54, 161–202. [DOI] [PubMed] [Google Scholar]
- Pertwee R. G.; Ross R. A. (2002) Cannabinoid receptors and their ligands. Prostaglandins, Leukotrienes, Essent. Fatty Acids 66, 101–121. [DOI] [PubMed] [Google Scholar]
- McAllister S. D.; Glass M. (2002) CB(1) and CB(2) receptor-mediated signalling: a focus on endocannabinoids. Prostaglandins, Leukotrienes, Essent. Fatty Acids 66, 161–171. [DOI] [PubMed] [Google Scholar]
- Sawzdargo M.; Nguyen T.; Lee D. K.; Lynch K. R.; Cheng R.; Heng H. H.; George S. R.; O’Dowd B. F. (1999) Identification and cloning of three novel human G protein-coupled receptor genes GPR52, PsiGPR53 and GPR55: GPR55 is extensively expressed in human brain. Brain Res. Mol. Brain Res. 64, 193–198. [DOI] [PubMed] [Google Scholar]
- McPartland J. M.; Matias I.; Di Marzo V.; Glass M. (2006) Evolutionary origins of the endocannabinoid system. Gene 370, 64–74. [DOI] [PubMed] [Google Scholar]
- Van Der Stelt M.; Di Marzo V. (2004) Endovanilloids. Putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur. J. Biochem. 271, 1827–1834. [DOI] [PubMed] [Google Scholar]
- Leung D.; Saghatelian A.; Simon G. M.; Cravatt B. F. (2006) Inactivation of N-acyl phosphatidylethanolamine phospholipase D reveals multiple mechanisms for the biosynthesis of endocannabinoids. Biochemistry 45, 4720–4726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon G. M.; Cravatt B. F. (2010) Characterization of mice lacking candidate N-acyl ethanolamine biosynthetic enzymes provides evidence for multiple pathways that contribute to endocannabinoid production in vivo. Mol. Biosyst. 6, 1411–1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natarajan V.; Reddy P. V.; Schmid P. C.; Schmid H. H. (1982) N-Acylation of ethanolamine phospholipids in canine myocardium. Biochim. Biophys. Acta 712, 342–355. [DOI] [PubMed] [Google Scholar]
- Natarajan V.; Schmid P. C.; Reddy P. V.; Schmid H. H. (1984) Catabolism of N-acylethanolamine phospholipids by dog brain preparations. J. Neurochem. 42, 1613–1619. [DOI] [PubMed] [Google Scholar]
- Simon G. M.; Cravatt B. F. (2006) Endocannabinoid biosynthesis proceeding through glycerophospho-N-acyl ethanolamine and a role for alpha/beta-hydrolase 4 in this pathway. J. Biol. Chem. 281, 26465–26472. [DOI] [PubMed] [Google Scholar]
- Liu J.; Wang L.; Harvey-White J.; Osei-Hyiaman D.; Razdan R.; Gong Q.; Chan A. C.; Zhou Z.; Huang B. X.; Kim H. Y.; Kunos G. (2006) A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. U.S.A. 103, 13345–13350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cravatt B. F.; Giang D. K.; Mayfield S. P.; Boger D. L.; Lerner R. A.; Gilula N. B. (1996) Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 384, 83–87. [DOI] [PubMed] [Google Scholar]
- Giang D. K.; Cravatt B. F. (1997) Molecular characterization of human and mouse fatty acid amide hydrolases. Proc. Natl. Acad. Sci. U.S.A. 94, 2238–2242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cravatt B. F.; Lichtman A. H. (2002) The enzymatic inactivation of the fatty acid amide class of signaling lipids. Chem. Phys. Lipids 121, 135–148. [DOI] [PubMed] [Google Scholar]
- Moriyama T.; Urade R.; Kito M. (1999) Purification and characterization of diacylglycerol lipase from human platelets. J. Biochem. 125, 1077–1085. [DOI] [PubMed] [Google Scholar]
- Goparaju S. K.; Ueda N.; Taniguchi K.; Yamamoto S. (1999) Enzymes of porcine brain hydrolyzing 2-arachidonoylglycerol, an endogenous ligand of cannabinoid receptors. Biochem. Pharmacol. 57, 417–423. [DOI] [PubMed] [Google Scholar]
- Pertwee R. G.; Howlett A. C.; Abood M. E.; Alexander S. P.; Di Marzo V.; Elphick M. R.; Greasley P. J.; Hansen H. S.; Kunos G.; Mackie K.; Mechoulam R.; Ross R. A. (2010) International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB and CB. Pharmacol. Rev. 62, 588–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans J. P.; Florman H. M. (2002) The state of the union: the cell biology of fertilization. Nature Cell Biol. 4, S57–S63. [DOI] [PubMed] [Google Scholar]
- Wassarman P. M.; Jovine L.; Litscher E. S. (2001) A profile of fertilization in mammals. Nature Cell Biol. 3, E59–E64. [DOI] [PubMed] [Google Scholar]
- Rossant J.; Tam P. P. (2004) Emerging asymmetry and embryonic patterning in early mouse development. Dev. Cell 7, 155–164. [DOI] [PubMed] [Google Scholar]
- Rossant J. (2004) Lineage development and polar asymmetries in the peri-implantation mouse blastocyst. Semin. Cell Dev. Biol. 15, 573–581. [DOI] [PubMed] [Google Scholar]
- Paria B. C.; Das S. K.; Dey S. K. (1995) The preimplantation mouse embryo is a target for cannabinoid ligand-receptor signaling. Proc. Natl. Acad. Sci. U.S.A. 92, 9460–9464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das S. K.; Paria B. C.; Chakraborty I.; Dey S. K. (1995) Cannabinoid ligand-receptor signaling in the mouse uterus. Proc. Natl. Acad. Sci. U.S.A. 92, 4332–4336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H.; Guo Y.; Wang D.; Kingsley P. J.; Marnett L. J.; Das S. K.; DuBois R. N.; Dey S. K. (2004) Aberrant cannabinoid signaling impairs oviductal transport of embryos. Nature Med. 10, 1074–1080. [DOI] [PubMed] [Google Scholar]
- Paria B. C.; Ma W.; Andrenyak D. M.; Schmid P. C.; Schmid H. H.; Moody D. E.; Deng H.; Makriyannis A.; Dey S. K. (1998) Effects of cannabinoids on preimplantation mouse embryo development and implantation are mediated by brain-type cannabinoid receptors. Biol. Reprod. 58, 1490–1495. [DOI] [PubMed] [Google Scholar]
- Yang Z. M.; Paria B. C.; Dey S. K. (1996) Activation of brain-type cannabinoid receptors interferes with preimplantation mouse embryo development. Biol. Reprod. 55, 756–761. [DOI] [PubMed] [Google Scholar]
- Schmid P. C.; Paria B. C.; Krebsbach R. J.; Schmid H. H.; Dey S. K. (1997) Changes in anandamide levels in mouse uterus are associated with uterine receptivity for embryo implantation. Proc. Natl. Acad. Sci. U.S.A. 94, 4188–4192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paria B. C.; Song H.; Wang X.; Schmid P. C.; Krebsbach R. J.; Schmid H. H.; Bonner T. I.; Zimmer A.; Dey S. K. (2001) Dysregulated cannabinoid signaling disrupts uterine receptivity for embryo implantation. J. Biol. Chem. 276, 20523–20528. [DOI] [PubMed] [Google Scholar]
- Wang H.; Dey S. K.; Maccarrone M. (2006) Jekyll and hyde: two faces of cannabinoid signaling in male and female fertility. Endocr. Rev. 27, 427–448. [DOI] [PubMed] [Google Scholar]
- Wang J.; Paria B. C.; Dey S. K.; Armant D. R. (1999) Stage-specific excitation of cannabinoid receptor exhibits differential effects on mouse embryonic development. Biol. Reprod. 60, 839–844. [DOI] [PubMed] [Google Scholar]
- Sharov A. A.; Piao Y.; Matoba R.; Dudekula D. B.; Qian Y.; VanBuren V.; Falco G.; Martin P. R.; Stagg C. A.; Bassey U. C.; Wang Y.; Carter M. G.; Hamatani T.; Aiba K.; Akutsu H.; Sharova L.; Tanaka T. S.; Kimber W. L.; Yoshikawa T.; Jaradat S. A.; Pantano S.; Nagaraja R.; Boheler K. R.; Taub D.; Hodes R. J.; Longo D. L.; Schlessinger D.; Keller J.; Klotz E.; Kelsoe G.; Umezawa A.; Vescovi A. L.; Rossant J.; Kunath T.; Hogan B. L.; Curci A.; D’Urso M.; Kelso J.; Hide W.; Ko M. S. (2003) Transcriptome analysis of mouse stem cells and early embryos. PLoS Biol. 1, E74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H., and Dey S. K., unpublished data.
- Pisarska M. D.; Carson S. A.; Buster J. E. (1998) Ectopic pregnancy. Lancet 351, 1115–1120. [DOI] [PubMed] [Google Scholar]
- Farquhar C. M. (2005) Ectopic pregnancy. Lancet 366, 583–591. [DOI] [PubMed] [Google Scholar]
- Wang H.; Xie H.; Guo Y.; Zhang H.; Takahashi T.; Kingsley P. J.; Marnett L. J.; Das S. K.; Cravatt B. F.; Dey S. K. (2006) Fatty acid amide hydrolase deficiency limits early pregnancy events. J. Clin. Invest. 116, 2122–2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo Y.; Wang H.; Okamoto Y.; Ueda N.; Kingsley P. J.; Marnett L. J.; Schmid H. H.; Das S. K.; Dey S. K. (2005) N-acylphosphatidylethanolamine-hydrolyzing phospholipase D is an important determinant of uterine anandamide levels during implantation. J. Biol. Chem. 280, 23429–23432. [DOI] [PubMed] [Google Scholar]
- Heilman R. D.; Reo R. R.; Hahn D. W. (1976) Changes in the sensitivity of adrenergic receptors in the oviduct during early gestation in the rabbit. Fertil. Steril. 27, 426–430. [PubMed] [Google Scholar]
- Howe G. R.; Black D. L. (1973) Autonomic nervous system and oviduct function in the rabbit. I. Hormones and contraction. J. Reprod. Fertil. 33, 425–430. [DOI] [PubMed] [Google Scholar]
- Paria B. C.; Huet-Hudson Y. M.; Dey S. K. (1993) Blastocyst’s state of activity determines the “window” of implantation in the receptive mouse uterus. Proc. Natl. Acad. Sci. U.S.A. 90, 10159–10162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshinaga K. (1988) Uterine receptivity for blastocyst implantation. Ann. N.Y. Acad. Sci. 541, 424–431. [DOI] [PubMed] [Google Scholar]
- Dey S. K.; Lim H.; Das S. K.; Reese J.; Paria B. C.; Daikoku T.; Wang H. (2004) Molecular cues to implantation. Endocr. Rev. 25, 341–373. [DOI] [PubMed] [Google Scholar]
- Wang H.; Dey S. K. (2006) Roadmap to embryo implantation: clues from mouse models. Nature Rev. 7, 185–199. [DOI] [PubMed] [Google Scholar]
- Wang H.; Xie H.; Sun X.; Kingsley P. J.; Marnett L. J.; Cravatt B. F.; Dey S. K. (2007) Differential regulation of endocannabinoid synthesis and degradation in the uterus during embryo implantation. Prostaglandins Other Lipid Mediators 83, 62–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor A. H.; Abbas M. S.; Habiba M. A.; Konje J. C. (2010) Histomorphometric evaluation of cannabinoid receptor and anandamide modulating enzyme expression in the human endometrium through the menstrual cycle. Histochem. Cell. Biol. 133, 557–565. [DOI] [PubMed] [Google Scholar]
- Maccarrone M.; DeFelici M.; Klinger F. G.; Battista N.; Fezza F.; Dainese E.; Siracusa G.; Finazzi-Agro A. (2004) Mouse blastocysts release a lipid which activates anandamide hydrolase in intact uterus. Mol. Hum. Reprod. 10, 215–221. [DOI] [PubMed] [Google Scholar]
- Wang H.; Matsumoto H.; Guo Y.; Paria B. C.; Roberts R. L.; Dey S. K. (2003) Differential G protein-coupled cannabinoid receptor signaling by anandamide directs blastocyst activation for implantation. Proc. Natl. Acad. Sci. U.S.A. 100, 14914–14919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maccarrone M.; Valensise H.; Bari M.; Lazzarin N.; Romanini C.; Finazzi-Agro A. (2000) Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet 355, 1326–1329. [DOI] [PubMed] [Google Scholar]
- Maccarrone M.; Bisogno T.; Valensise H.; Lazzarin N.; Fezza F.; Manna C.; Di Marzo V.; Finazzi-Agro A. (2002) Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol. Hum. Reprod. 8, 188–195. [DOI] [PubMed] [Google Scholar]
- El-Talatini M. R.; Taylor A. H.; Konje J. C. (2009) Fluctuation in anandamide levels from ovulation to early pregnancy in in-vitro fertilization-embryo transfer women, and its hormonal regulation. Hum. Reprod. 24, 1989–1998. [DOI] [PubMed] [Google Scholar]
- Cross J. C.; Baczyk D.; Dobric N.; Hemberger M.; Hughes M.; Simmons D. G.; Yamamoto H.; Kingdom J. C. (2003) Genes, development and evolution of the placenta. Placenta 24, 123–130. [DOI] [PubMed] [Google Scholar]
- Sun X.; Xie H.; Yang J.; Wang H.; Bradshaw H. B.; Dey S. K. (2010) Endocannabinoid signaling directs differentiation of trophoblast cell lineages and placentation. Proc. Natl. Acad. Sci. U.S.A. 107, 16887–16892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park B.; Gibbons H. M.; Mitchell M. D.; Glass M. (2003) Identification of the CB1 cannabinoid receptor and fatty acid amide hydrolase (FAAH) in the human placenta. Placenta 24, 990–995. [DOI] [PubMed] [Google Scholar]
- Habayeb O. M.; Taylor A. H.; Bell S. C.; Taylor D. J.; Konje J. C. (2008) Expression of the endocannabinoid system in human first trimester placenta and its role in trophoblast proliferation. Endocrinology 149, 5052–5060. [DOI] [PubMed] [Google Scholar]
- Cella M.; Leguizamon G. F.; Sordelli M. S.; Cervini M.; Guadagnoli T.; Ribeiro M. L.; Franchi A. M.; Farina M. G. (2008) Dual effect of anandamide on rat placenta nitric oxide synthesis. Placenta 29, 699–707. [DOI] [PubMed] [Google Scholar]
- Guillemot F.; Nagy A.; Auerbach A.; Rossant J.; Joyner A. L. (1994) Essential role of Mash-2 in extraembryonic development. Nature 371, 333–336. [DOI] [PubMed] [Google Scholar]
- Riley P.; Anson-Cartwright L.; Cross J. C. (1998) The Hand1 bHLH transcription factor is essential for placentation and cardiac morphogenesis. Nat. Genet. 18, 271–275. [DOI] [PubMed] [Google Scholar]
- Cross J. C.; Hemberger M.; Lu Y.; Nozaki T.; Whiteley K.; Masutani M.; Adamson S. L. (2002) Trophoblast functions, angiogenesis and remodeling of the maternal vasculature in the placenta. Mol. Cell. Endocrinol. 187, 207–212. [DOI] [PubMed] [Google Scholar]
- Kelley L., and Faddis R., Eds. (1999) Reducing Perinatal and Neonatal Mortality, a Child Health Research Project Special Report, pp 1–48, Johns Hopkins University Office of Design and Publications, Baltimore, MD. [Google Scholar]
- Mathews T. J.; MacDorman M. F. (2006) Infant mortality statistics from the 2003 period linked birth/infant death data set. Natl. Vital Stat. Rep. 54, 1–29. [PubMed] [Google Scholar]
- Mesiano S.; Welsh T. N. (2007) Steroid hormone control of myometrial contractility and parturition. Semin. Cell Dev. Biol. 18, 321–331. [DOI] [PubMed] [Google Scholar]
- Wang H.; Xie H.; Dey S. K. (2008) Loss of cannabinoid receptor CB1 induces preterm birth. PLoS ONE 3, e3320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H.; Tranguch S.; Xie H.; Hanley G.; Das S. K.; Dey S. K. (2005) Variation in commercial rodent diets induces disparate molecular and physiological changes in the mouse uterus. Proc. Natl. Acad. Sci. U.S.A. 102, 9960–9965. [DOI] [PMC free article] [PubMed] [Google Scholar]