

Abstract
We demonstrate that reactive oxygen species (ROS) plays an important role in the process of apoptosis in human peripheral blood mononuclear cell (PBMC) which is induced by the radiation of 900 MHz radiofrequency electromagnetic field (RFEMF) at a specific absorption rate (SAR) of ~0.4 W/kg when the exposure lasts longer than two hours. The apoptosis is induced through the mitochondrial pathway and mediated by activating ROS and caspase-3, and decreasing the mitochondrial potential. The activation of ROS is triggered by the conformation disturbance of lipids, protein, and DNA induced by the exposure of GSM RFEMF. Although human PBMC was found to have a self-protection mechanism of releasing carotenoid in response to oxidative stress to lessen the further increase of ROS, the imbalance between the antioxidant defenses and ROS formation still results in an increase of cell death with the exposure time and can cause about 37% human PBMC death in eight hours.
1. Introduction
Mobile phones have been widely used in popular telecommunication and medical telemetry systems. The tremendous use of mobile phone has drastically increased the amount of radiofrequency electromagnetic field (GSM RFEMF) exposure in our daily lives. To ensure telecommunication in anywhere, various kinds of mobile phone relay stations or devices need to be placed inside or near living/working and residential areas. It makes people have the possibility to be exposed to the RFEMF radiation almost every moment. Thus there is a major concern about the effects of RFEMF radiation exposure on human health. Despite previous studies, our knowledge on these effects is still inadequate and strong debates continue [1–5].
Among the various health effects of GSM RFEMF exposure, the formation of reactive oxygen species (ROS) and increased oxidative stress are those proposed mechanisms that can explain the link between RFEMF radiation and possible harmful effects on human health. It was found that RFEMF could induce ROS formation in animal brain, cortical neurons, spleen, blood serum, and human semen [6–10]. The purpose of this study was to investigate the extent of ROS formation and oxidative DNA damage as well as cell apoptosis caused by RFEMF on human peripheral blood mononuclear cell (PBMC). PBMC cells are a critical component in the immune system to fight infection and adapt to intruders. They also play significant roles in neurodegenerative diseases and aging [11–14]. Therefore, investigation of whether and how oxidative stress activates in PBMC under the exposure of RFEMF radiation can help to further clarify its effects on human health.
In this study, isolated fresh human peripheral blood mononuclear cells were exposed to the radiation of 900 MHz GSM RFEMF at a specific absorption rate (SAR) of 0.4 W/kg for 1 h, 2 h, 4 h, 6 h, and 8 h. The specific absorption rate was chosen to mimic the situation that people usually may absorb in an environment within a distance of 20 meters from mobile phone relay stations, or occupationally in an equipment room of microwave communication, or around a surveillance radar [15–17]. It is also the occupational exposure restriction suggested by the International Commission on Non-ionizing Radiation Protection and some national radiological protection boards [18, 19].
To detect the intracellular ROS activation in the exposed cells, fluorescent dye DCFH was used as the probe in flow cytometry. The caspase-3 activity of the cells was assessed by colorimetric assay, while the cell apoptosis was analyzed by flow cytometry with FITC-Annexin V/Propidium Iodide (PI) double staining. To assess DNA damage of human PBMC and reveal the mechanism of the effect of RFEMF radiation, confocal Raman microspectroscopy was also employed.
2. Material and Methods
2.1. Sample and Reagents
Study on blood of volunteers (providing informed written consent) was proved by Jinan University Animal Care and Use Committee conforming to the Chinese Public Health Service Police on Human Care and Use of Laboratory Animals.
Normal peripheral blood was obtained from healthy nonsmoking adult volunteers aging 25.3 ± 0.8 by venipuncture and poured into heparinized tubes. The blood samples were anticoagulated with heparin lithium. After centrifugation, the peripheral blood monocytes in the middle cloud layer were taken out, washed twice repeatedly, and then re-suspended. The cell survival rate was >98% estimated by Trypan blue staining.
Annexin V/PI double-staining kit was purchased from Bender Company, USA. The fluorescent dye DAPI was from Roche, USA. The mitochondrial membrane potential detection kit (JC-1), ROS detection kit, Bradford protein concentration assay kit, and caspase-3 colorimetric assay kit were all purchased from Beyotime Institute of Biotechnology, China.
2.2. Exposure of Human PBMC Samples to RFEMF
200 μL of PBMC samples with cell density of 1.5 × 106/L was placed in each well of a culture plate. Then they were exposed to the radiation emitted by a VS401A RF RFEMF emitter (Shenzhen Weikete Technology Company, Ltd., China) at a specific absorption rate of 0.43 W/kg at 37°C for 1 h, 2 h, 4 h, 6 h, and 8 h. The radiation distributed uniformly on the sample and the SAR was determined using the conductivity of the PBMC sample σ, the RFEMF electric field strength E at the determined point, and the mass density of the sample ρ m in the follwoing form: SAR = σE 2/ρ m. In the experiment, σ was found to be 0.229 ± 0.001 (S/m), E was 43.42 (V/m), and ρ m was 1.011 ± 0.006 (g/mL). Therefore, SAR was estimated to be 0.43 W/kg.
2.3. Cell Apoptosis Detection
5 μL FITC-Annexin V and 10 μLPI were added to 100 μL cell suspension with cell concentration of 1 × 106/mL. The mixture was incubated for 15 minutes in dark at room temperature. Then they were washed with binding buffer twice and adjusted again to the cell concentration of 1 × 106/mL. The cell apoptosis was analyzed using an FACS Aria flow cytometry (BD company, USA) within 1 hour.
2.4. ROS Detection
The exposed cells were collected and the supernatant was removed by centrifugation. Thereafter the cells were resuspended and 5 × 105 cells were collected. They were centrifuged again to remove the supernatant and then added into 500 μL diluted DCFH-DA. The mixture was incubated for 20 minutes at 37°C and then washed twice. The samples were later analyzed with flow cytometer within 1 hour. An Ar+ laser with 488 nm wavelength was used as the excitation light and 525 nm was the receiving wavelength to obtain the proportion of the fluorescent cells.
2.5. Caspace-3 Activity Detection
The caspace-3 activity of the exposed cells was evaluated using the caspase-3 colorimetric assay kit and the assessment was performed according to the manufacturer's recommendations. The ratio of the OD value of the sample and that of the control group were taken to evaluate the caspase-3 activity.
2.6. Mitochondrial Membrane Potential Determination
(10–60) × 105 exposed cells were resuspended and mixed with the JC-1 staining working solution. The mixture was incubated at 37°C for 20 minutes and then centrifuged for 3-4 minutes to remove the supernatant. The mixture was washed twice with buffer solution and then the cells were resuspended with the buffer solution. The fluorescence of the cells was imaged using a Nikon TE300 inverted fluorescence microscope.
2.7. DNA Damage Detection by Raman Spectroscopy
The Raman spectra of PBMC were recorded by a JY RAM INV system using 514.2 nm excitation line from an Ar+ ion laser through an inverted Olympus optical microscope with a ×60 objective. The acquisition band was 600~1800 cm−1 with a spectrum resolution of 1 cm−1. At least 35 cells were measured for each group of the exposed PBMC sample.
2.8. Data Processing
The PBMC cells were from the blood samples of 6 volunteers (3 males and 3 females). Each sample contained 10000 cells. All data were averaged from the results of five parallel samples; each sample was detected three times. The final result is denoted by . SPSS 13.0 was used for statistical analysis of the data, in which P < 0.05 was regarded as significantly different.
3. Results
3.1. ROS Activation
The flow cytometric results of human PBMCs' ROS activation are shown in Figure 1. Figure 1(a) indicates the histograms of ROS-positive cells, and Figure 1(b) shows the histograms of mean DCF fluorescence intensity (indication of ROS level). Figure 2 shows how the ROS-positive cells and the ROS level vary with radiation time. We can see that just 1 h radiation can activate ROS in PBMC (P < 0.05, versus control). The ROS level continuously rose in the period from the 2nd h to the 6th h. After 6 h exposure, both the number of ROS-positive cells and ROS level reached their maximum and then declined.
Figure 1.
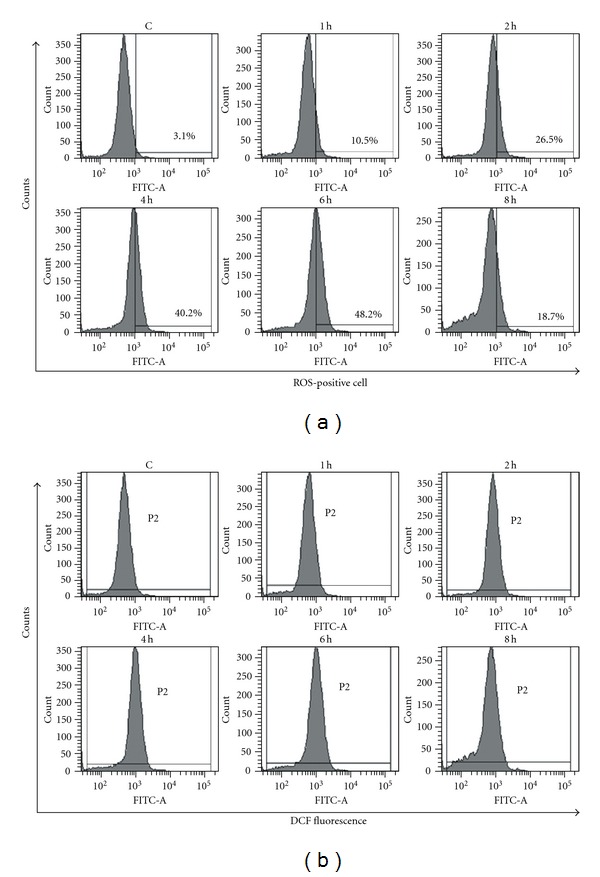
The flow cytometric results of human PBMCs' ROS activation. (a) Histograms of ROS-positive cell percentage. (b) Histograms of mean DCF fluorescence intensity.
Figure 2.
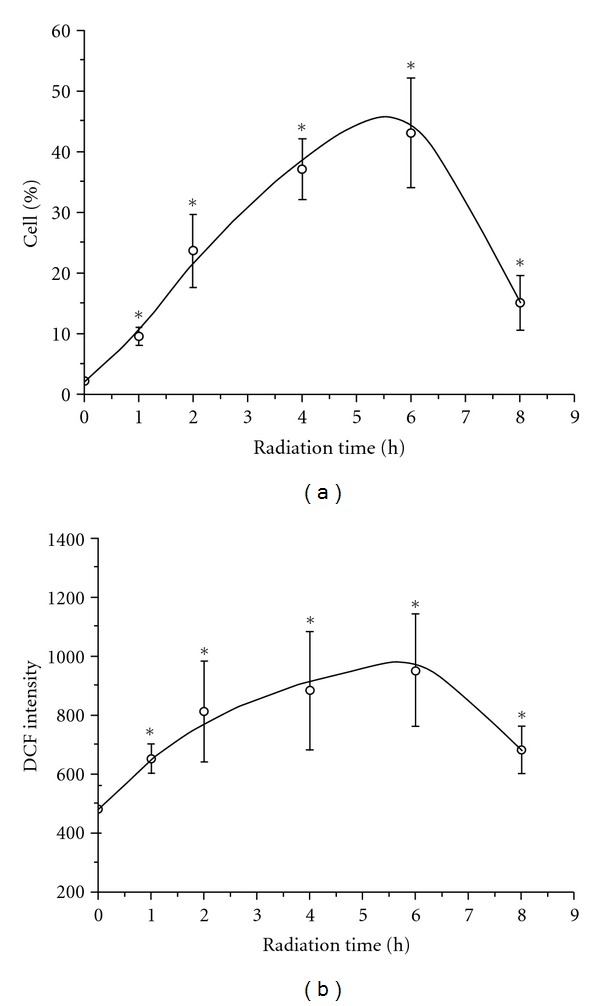
The number of ROS-positive cells (a) and DCF intensity (b) versus radiation time.
3.2. Caspace-3 Activity
The variation of caspacse-3 activity in human PBMCs with radiation time is shown in Figure 3. Within the first 2 h radiation, the change of caspase-3 content was not evident (P > 0.05 versus control). However, when the cells were radiated longer than 2 h, the caspase-3 activity became significantly increased (P < 0.05 versus control). The activity of caspase-3 at the 6th h was 6 times as that of control group. But at the 8th h, the caspase-3 activity declined significantly compared with that at the 6th h (P < 0.05).
Figure 3.
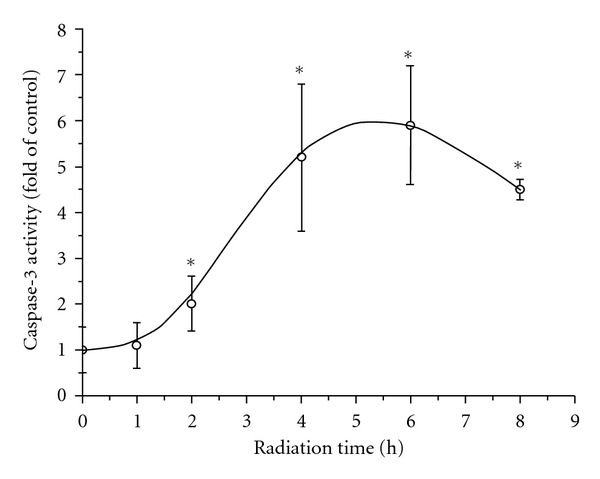
The variation of caspacse-3 activity in human PBMCs with radiation time.
3.3. Mitochondrial Potential
Figure 4 illustrates the mitochondrial staining of human PBMCs. The PBMCs in the control group (not exposed to electromagnetic radiation) emitted bright orange-red fluorescence with few emitting green fluorescence. The red fluorescence intensity of the cells weakened while the proportion of the green fluorescence cells increased in the images taken from the 2nd h to the 6th h, indicating a decline of mitochondrial potential in the cells during the period. However, it slightly went up at the 8th h.
Figure 4.
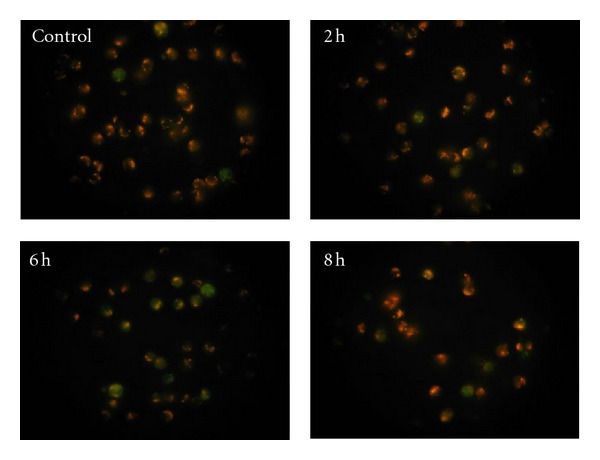
The mitochondrial staining images of human PBMCs.
3.4. Human PBMC Apoptosis
Figure 5 shows the flow cytometric analysis of apoptosis in human PBMC using FITC-annexin V and PI double staining, and Figure 6 illustrates the apoptotic rates of the exposed cells. It can be seen that neither early apoptosis (Annexin V+/PI−) nor late apoptosis (Annexin V+/PI+) was evident (P > 0.05 versus control) in the 1st h. When the exposure lasted longer than 2 h, the apoptotic rates increased evidently. The early apoptotic rate increased to 12.2% ± 3.3% and 21.5% ± 5.2% (P < 0.05 versus control), respectively, at the 2nd h and 4th h. At the same time, the late apoptotic rate increased to 2.2% ± 0.8% and 5.0% ± 1.6% (P < 0.05 versus control). Compared with the early apoptotic rate at the 4th h, there was no significant increase (P > 0.05 versus the 4th h) at the 6th h, whereas the late apoptotic rate began to decrease at the time. At the 8th h, the early apoptotic rate decreased from 21.2% ± 4.9% (at the 6th h) to 10.4% ± 5.0% (P < 0.05 versus the 6th h). Figures 6(a) and 6(b), respectively, show the detail information about the variations of early and late apoptotic rates of human PBMC with radiation time.
Figure 5.
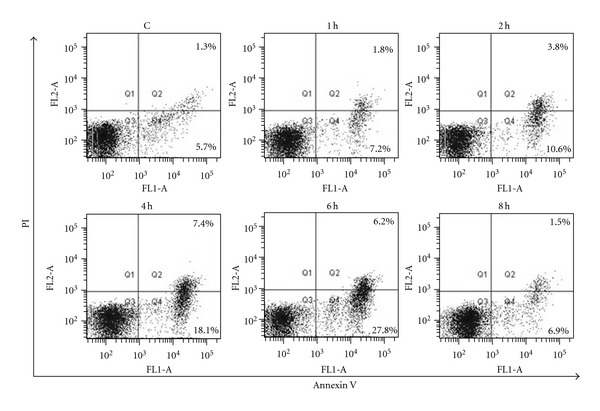
The flow cytometric analysis of apoptosis in human PBMCs using FITC-annexin V and PI double staining. Quadrant analysis of the gated cells in FL-1 versus FL-2 channels was from 10,000 events. Annexin V+/PI− (lower right quadrant) areas stand for early apoptotic cells, and Annexin V+/PI+ (upper right quadrant) areas stand for late apoptotic or necrotic cells.
Figure 6.
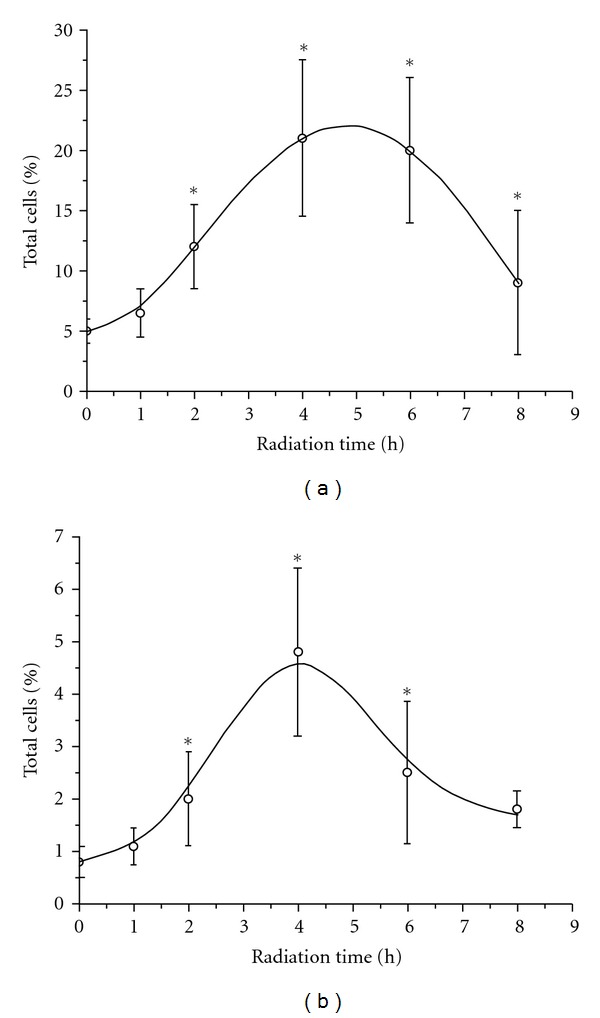
The apoptotic rates of exposed human PBMCs:(a) Early apoptotic cells and (b) late apoptotic cells.
3.5. Raman Spectra
Two kinds of Raman spectra were obtained from the exposed cells. One contains weak signal of carotenoid but the other one contains strong signal of carotenoid. Both of them are shown in Figure 7, with peaks at 1157 and 1525 cm−1 being the bands of carotenoid. The spectra with strong signal of carotenoid were observed only in the samples being exposed longer than one hour but not found in the control group. The proportion of the spectra with strong signal of carotenoid thereafter increased with radiation time and become 60% of all the observed ones in the 4th h. This indicates that the carotenoid releasing in the exposed cells was a reaction to the exposure.
Figure 7.
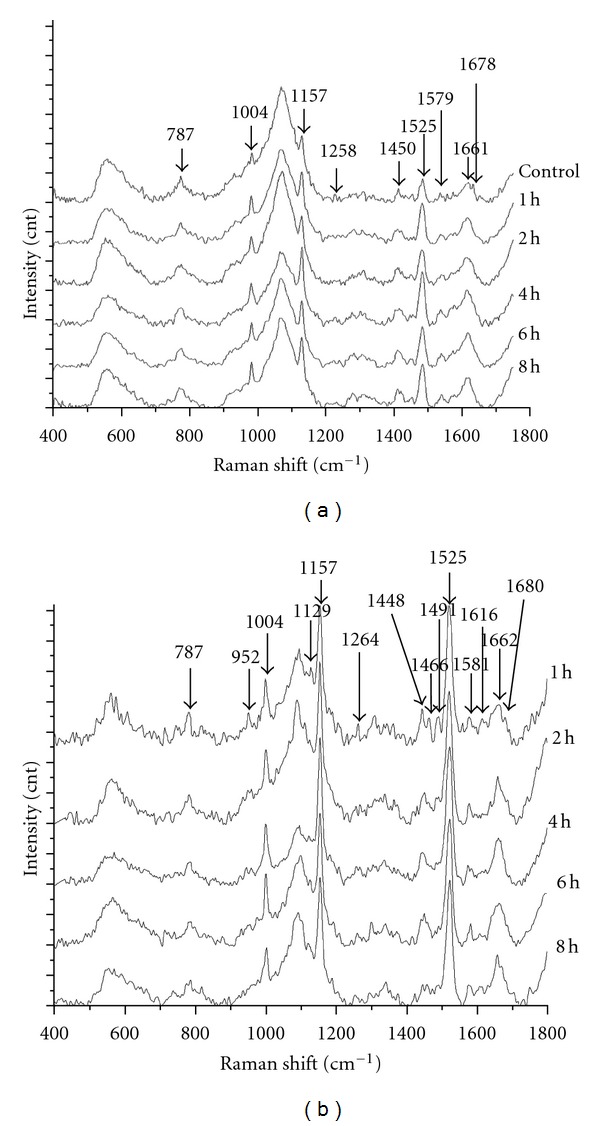
The Raman spectra of PBMC which were exposed for different time. (a) Raman spectra with weak signal of carotenoid. (b) Raman spectra with strong signal of carotenoid.
Besides the bands assigned to carotenoid, the bands assigned to DNA (787, 1258, and 1579 cm−1 in Figure 7(a); 787, 952, 1491, and 1581 cm−1 in Figure 7(b)) and the bands assigned to protein (1004, 1450, 1616, 1661, and 1678 cm−1) also change evidently with radiation time.
3.6. Cell Counting
Figure 8 illustrates the result of cell counting on the exposed human PBMC as a function of radiation time. We can see that the number of cells constantly decreases within six hours and then significantly reduces from the 6th h to the 8th h. At the 8th h, about 37% of the exposed cells had died.
Figure 8.
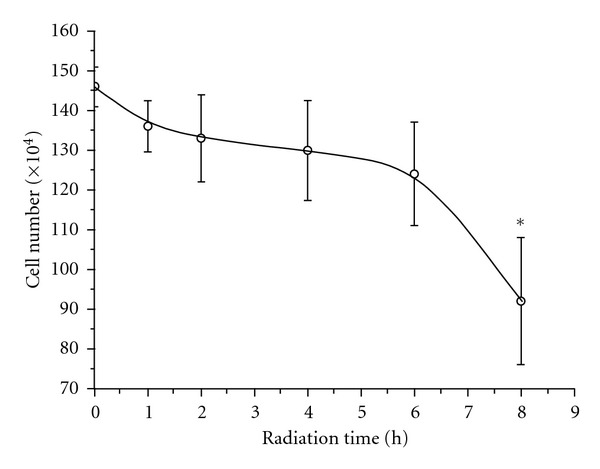
The cell counting of the exposed human PBMC versus radiation time.
4. Discussion
From the results we can see obviously that cell apoptosis can be induced in human peripheral blood mononuclear cell (PBMC) by the radiation of 900 MHz GSM RFEMF at a specific absorption rate of ~0.4 W/kg when the exposure lasts longer than two hours. Using the data about ROS activation, caspacse-3 activity, mitochondrial potential and the Raman spectra of DNA and proteins, we can figure out the mechanism of the cell apoptosis as follows. The exposure to the radiation of 900 MHz GSM RFEMF can induce a series changes in the protein, lipid, and DNA structure. These changes include (1) broken carbon-hydrogen bond of lipid and protein (indicated by the intensity decrease at 1130 cm−1 in the Raman spectra), (2) damage of the protein side chain (Phe, Tyr, indicated by the intensity decrease at 1616 cm−1 in the Raman spectra), (3) destruction of the protein secondary structure such as reducing α-helix and β-sheet and increasing random coil (indicated by the intensity decreases at 1264 cm−1 and 1678/1680 cm−1 in the Raman spectra), and (4) DNA damage (indicated by the intensity decreases at 952 cm−1 and 1491 cm−1 and intensity increase at 1579 cm−1 in the Raman spectra). All these changes influence the stability of the protein conformation, so that the proteins cannot perform their normal function to get rid of the excess ROS. The imbalance between ROS formation and antioxidant defenses results in oxidative stress in human PBMC, thus inducing mitochondrial permeability transition pore (mPTP) opening [20, 21]. The opening of mPTP declines the mitochondrial potential, thereby triggering the caspacse-3 activity and finally inducing cell apoptosis [22–26]. The apoptosis was mainly early apoptosis, with less than 6% of the cells being late apoptosis.
This is the so-called mitochondrial pathway of apoptosis and has been demonstrated step by step by our experimental results. As described previously, the ROS activation was induced by DNA damage and the disturbance on protein and lipid conformation, suggesting that DNA, protein, and lipid probably are the targets of the GSM RFEMF radiation on human PBMC. On the other hand, human PBMC seems to have a self-protection mechanism of releasing carotenoid in response to oxidative stress to inhibit the further increase of ROS. However, it cannot stop the process of cell death if the exposure continues. The number of cells even decreased faster in the period from the 6th h to the 8th h as shown in Figure 8. A possibility is that the amount of releasing carotenoid was not enough to against the excessive ROS generation. Another possibility is that, besides cell apoptosis, human PBMC has another cell death process induced by the GSM RFEMF exposure. It is oncosis [27–29] and was proved by our experiment of cell morphological observation on the exposed human PBMC (data not shown). Therefore, the cell number continuously decreased from the 6th h to the 8th h even though the number of apoptotic cells had already decreased in the period. We will not discuss the mechanism of the cell oncosis in human PBMC here but will leave it for a future paper. Finally, we strongly ask for more concern on the possible hazardous health effects of exposure to the radiation of GSM RFEMF emitted from the mobile phone relay stations or devices as it can cause 37% human PBMC death in eight hours.
5. Conclusions
We have demonstrated that cell apoptosis can be induced in human PBMC by the radiation of 900 MHz GSM RFEMF at a specific absorption rate of ~0.4 W/kg when the exposure lasts longer than two hours. The apoptosis is induced through the mitochondrial pathway and mediated by activating ROS and caspase-3, and decreasing the mitochondrial potential. The activation of ROS is triggered by the conformation disturbance of lipids, protein, and DNA induced by the exposure to GSM RFEMF. Although human PBMC has a self-protection mechanism of releasing carotenoid to inhibit further increase of ROS, if the exposure continues, the imbalance between the antioxidant defenses and ROS formation still results in an increase of cell death with the exposure time. These findings not only clarify the effect of GSM RFEMF on human health but also reveal its mechanism. We hope that it will help people to realize the possible hazardous health effects of exposure to GSM RFEMF radiation emitted from the mobile phone relay stations or devices in their living/occupational environment.
References
- 1.Hardell L, Sage C. Biological effects from electromagnetic field exposure and public exposure standards. Biomedicine and Pharmacotherapy. 2008;62(2):104–109. doi: 10.1016/j.biopha.2007.12.004. [DOI] [PubMed] [Google Scholar]
- 2.Khurana VG, Teo C, Kundi M, Hardell L, Carlberg M. Cell phones and brain tumors: a review including the long-term epidemiologic data. Surgical Neurology. 2009;72(3):205–214. doi: 10.1016/j.surneu.2009.01.019. [DOI] [PubMed] [Google Scholar]
- 3.Salama N, Kishimoto T, Kanayama HO. Effects of exposure to a mobile phone on testicular function and structure in adult rabbit. International Journal of Andrology. 2010;33(1):88–94. doi: 10.1111/j.1365-2605.2008.00940.x. [DOI] [PubMed] [Google Scholar]
- 4.Abdus-salam A, Elumelu T, Adenipekun A. Mobile phone radiation and the risk of cancer; a review. African Journal of Medicine and Medical Sciences. 2008;37(2):107–118. [PubMed] [Google Scholar]
- 5.Krewski D, Glickman BW, Habash RWY, et al. Recent advances in research on radiofrequency fields and health: 2001–2003. Journal of Toxicology and Environmental Health—Part B. 2007;10(4):287–318. doi: 10.1080/15287390600974973. [DOI] [PubMed] [Google Scholar]
- 6.Agarwal A, Desai NR, Makker K, et al. Effects of radiofrequency electromagnetic waves (RF-EMW) from cellular phones on human ejaculated semen: an in vitro pilot study. Fertility and Sterility. 2009;92(4):1318–1325. doi: 10.1016/j.fertnstert.2008.08.022. [DOI] [PubMed] [Google Scholar]
- 7.De Iuliis GN, Newey RJ, King BV, Aitken RJ. Mobile phone radiation induces reactive oxygen species production and DNA damage in human spermatozoa in vitro. PLoS ONE. 2009;4(7) doi: 10.1371/journal.pone.0006446.e6446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sokolovic D, Djindjic B, Nikolic J, et al. Melatonin reduces oxidative stress induced by chronic exposure of microwave radiation from mobile phones in rat brain. Journal of Radiation Research. 2008;49(6):579–586. doi: 10.1269/jrr.07077. [DOI] [PubMed] [Google Scholar]
- 9.Stankiewicz W, Dabrowski MP, Kubacki R, Sobiczewska E, Szmigielski S. Immunotropic influence of 900 MHz microwave GSM signal on human blood immune cells activated in vitro. Electromagnetic Biology and Medicine. 2006;25(1):45–51. doi: 10.1080/15368370600572961. [DOI] [PubMed] [Google Scholar]
- 10.Dasdag S, Bilgin HM, Akdag MZ, Celik H, Aksen F. Effect of long term mobile phone exposure on oxidative-antioxidative processes and nitric oxide in rats. Biotechnology and Biotechnological Equipment. 2008;22(4):992–997. [Google Scholar]
- 11.Colombo C, Cosentino M, Marino F, et al. Dopaminergic modulation of apoptosis in human peripheral blood mononuclear cells. Annals of the New York Academy of Sciences. 2003;1010:679–682. doi: 10.1196/annals.1299.124. [DOI] [PubMed] [Google Scholar]
- 12.Jenny M, Klieber M, Zaknun D, et al. In vitro testing for anti-inflammatory properties of compounds employing peripheral blood mononuclear cells freshly isolated from healthy donors. Inflammation Research. 2011;60(2):127–135. doi: 10.1007/s00011-010-0244-y. [DOI] [PubMed] [Google Scholar]
- 13.Prigione A, Begni B, Galbussera A, et al. Oxidative stress in peripheral blood mononuclear cells from patients with Parkinson’s disease: negative correlation with levodopa dosage. Neurobiology of Disease. 2006;23(1):36–43. doi: 10.1016/j.nbd.2006.01.013. [DOI] [PubMed] [Google Scholar]
- 14.Zhu H, Belcher M, van der Harst P. Healthy aging and disease: role for telomere biology? Clinical Science. 2011;120(10):427–440. doi: 10.1042/CS20100385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gupta JP. Microwave radiation hazards from radars and other high power microwave generators. Defence Science Journal. 1988;38(3):287–292. [Google Scholar]
- 16.Puranen L, Jokela K. Radiation hazard assessment of pulsed microwave radars. Journal of Microwave Power and Electromagnetic Energy. 1996;31(3):165–177. doi: 10.1080/08327823.1996.11688307. [DOI] [PubMed] [Google Scholar]
- 17.Petersen RC. Electromagnetic radiation from selected telecommunications systems. Proceedings of the IEEE. 1980;68(1):21–24. [Google Scholar]
- 18. International Commission on Non-ionizing Radiation Protection, ICNIRP Guidelines, 1998.
- 19.Mann SM. Exposure to Radio Waves Near Mobile Phone Base Stations. Oxon, UK: National Radiological Protection Board; 2000. [Google Scholar]
- 20.Cao XH, Zhao SS, Liu DY, et al. ROS-Ca2+ is associated with mitochondria permeability transition pore involved in surfactin-induced MCF-7 cells apoptosis. Chemico-Biological Interactions. 2011;190(1):16–27. doi: 10.1016/j.cbi.2011.01.010. [DOI] [PubMed] [Google Scholar]
- 21.Zorov DB, Filburn CR, Klotz LO, Zweier JL, Sollott SJ. Reactive oxygen species (ROS)-induced ROS release: a new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. Journal of Experimental Medicine. 2000;192(7):1001–1014. doi: 10.1084/jem.192.7.1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Breckenridge DG, Xue D. Regulation of mitochondrial membrane permeabilization by BCL-2 family proteins and caspases. Current Opinion in Cell Biology. 2004;16(6):647–652. doi: 10.1016/j.ceb.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 23.Green DR, Reed JC. Mitochondria and apoptosis. Science. 1998;281(5381):1309–1312. doi: 10.1126/science.281.5381.1309. [DOI] [PubMed] [Google Scholar]
- 24.Mohamad N, Gutiérrez A, Núñez M, et al. Mitochondrial apoptotic pathways. Biocell. 2005;29(2):149–161. [PubMed] [Google Scholar]
- 25.Spierings D, McStay G, Saleh M, et al. Connected to death: the (unexpurgated) mitochondrial pathway of apoptosis. Science. 2005;310(5745):66–67. doi: 10.1126/science.1117105. [DOI] [PubMed] [Google Scholar]
- 26.Verma SP, Singhal A. Low levels of the pesticide, δ-hexachlorocyclohexane, lyses human erythrocytes and alters the organization of membrane lipids and proteins as revealed by Raman spectroscopy. Biochimica et Biophysica Acta. 1991;1070(1):265–273. doi: 10.1016/0005-2736(91)90174-7. [DOI] [PubMed] [Google Scholar]
- 27.Levin S. Apoptosis, necrosis, or oncosis: what is your diagnosis? A report from the cell death nomenclature committee of the society of toxicologie pathologists 1. Toxicological Sciences. 1998;41(2):155–156. doi: 10.1006/toxs.1997.2432. [DOI] [PubMed] [Google Scholar]
- 28.Majno G, Joris I. Apoptosis, oncosis, and necrosis: an overview of cell death. American Journal of Pathology. 1995;146(1):3–15. [PMC free article] [PubMed] [Google Scholar]
- 29.Scarfì S, Magnone M, Ferraris C, et al. Ascorbic acid pre-treated quartz stimulates TNF-α release in RAW 264.7 murine macrophages through ROS production and membrane lipid peroxidation. Respiratory Research. 2009;10(1, article 25) doi: 10.1186/1465-9921-10-25. [DOI] [PMC free article] [PubMed] [Google Scholar]