

Abstract
The social brain hypothesis (an explanation for the evolution of brain size in primates) predicts that humans typically cannot maintain more than 150 relationships at any one time. The constraint is partly cognitive (ultimately determined by some aspect of brain volume) and partly one of time. Friendships (but not necessarily kin relationships) are maintained by investing time in them, and failure to do so results in an inexorable deterioration in the quality of a relationship. The Internet, and in particular the rise of social networking sites (SNSs), raises the possibility that digital media might allow us to circumvent some or all of these constraints. This allows us to test the importance of these constraints in limiting human sociality. Although the recency of SNSs means that there have been relatively few studies, those that are available suggest that, in general, the ability to broadcast to many individuals at once, and the possibilities this provides in terms of continuously updating our understanding of network members’ behaviour and thoughts, do not allow larger networks to be maintained. This may be because only relatively weak quality relationships can be maintained without face-to-face interaction.
Keywords: social brain hypothesis, social networks, Internet, social networking site, relationships
1. Introduction
During the last two decades, the Internet has opened up opportunities for communication that have revolutionized many aspects of our lives. Initially, email, bulletin boards and chat rooms markedly increased both the speed and the geographical reach of interaction, but these have been superseded in a marked fashion by the spectacular rise of Facebook and other social networking sites (SNSs) in the last half decade or so. The uptake of SNSs in particular has been little short of explosive: in the 8 years since it was launched in February 2004, Facebook alone has acquired more than 750 million active users, half of whom log in every day [http://www.facebook.com/press/info.php?statistics]. From the outset, these novel forms of digital communication were widely touted as offering the opportunity to expand our social horizons. The question is whether, in fact, they have been able to do so.
This is not a question of parochial interest. It bears on substantive issues that concern the nature of human social relationships and the cognitive and other mechanisms that underpin them. The social brain hypothesis [1] has argued that social group size in primates is constrained by some aspect of cognition (instantiated in hardware terms in some aspect of neuronal volume and connectivity), and this suggestion has been extended to humans with a specific prediction as to the number of relationships that the normal adult human can maintain as a coherent network [2]. This raises two important questions: (i) exactly what aspects of cognition and/or behaviour limit social group/network size and (ii) given that these limits exist, is it possible for technology to override these constraints and allow humans to maintain larger social groups? Internet services provide what is, in effect, a natural experiment that allows us to determine whether or not the constraints imposed by face-to-face interaction inevitably limit the size of human social networks. The answers to these questions have both historical relevance (how is it that humans differ from other great apes, and how did they come to be that way?) and implications for the design of future technology (how should we design digital communication technology to make better use of natural human capacities, and is it possible to do this in such a way that it allows us to increase the integration and cohesion of contemporary large-scale societies?).
In this paper, I first set the scene by briefly outlining the social brain hypothesis and the evidence for it, and summarize its implications for humans. I will then consider the structure of human social relationships in more detail in preparation for an examination of whether digital technology (i.e. the Internet) makes it possible to cut through the natural constraints that these processes impose so as to allow us to increase either the number or the quality of our social relationships. Finally, I return to the issue of whether the ways in which our relationships work limit our capacity to take full advantage of these novel modes of communication.
2. The social brain hypothesis
A number of alternative hypotheses have been suggested over the last four decades to explain the evolution of primates’ unusually large brains. The principal contenders have been a set of developmental hypotheses, a set of ecological hypotheses and several versions of the social brain hypothesis. The first category does not, strictly speaking, provide functional explanations (i.e. explanations that reflect the direct influence of selection), but rather constitute constraints that must be solved if large brains are to evolve. The ecological and social hypotheses provide functional explanations, but differ in which aspects of animals’ biology (specifically, survival rates versus birth rates) are regarded as rate-limiting (i.e. are more adversely affected and thus constitute the principal driver for evolutionary change). The various hypotheses and their functional consequences are summarized in figure 1.
Figure 1.

Alternative hypotheses for the evolution of large brains in primates. Hypotheses differ in whether their central claim is about ontogeny, ecological or social processes, on whether they view food or predation as the rate-limiting process in population dynamics, and on whether they view the fitness benefits from large brains as being direct or indirect. Adapted from Dunbar [3].
There has continued to be considerable confusion about the relationships between these various explanations, in part because most analyses focus on a single explanation and simply seek confirmatory evidence for that hypothesis. However, a second problem has been a consistent failure to distinguish between causes, consequences and constraints in biological explanations [4]. The persistent emphasis in the literature on developmental and life history correlates of brain volume is one reflection of this. It is not enough to observe that a species has evolved a large brain simply because it can do so, thanks to a change in body size, diet, the duration of gestation, lactation or longevity. Brain tissue is extremely expensive [5,6], and significant advantages must exist if encephalization is to occur in the face of so steep a selection gradient: that animals have the energetic capacity to encephalize does not mean that they must inevitably do so. There are two classes of explanations that provide the required benefit, namely ecological hypotheses and social brain hypotheses: both argue that larger brains are necessary to solve some problem that limits a species’ capacity to survive and reproduce successfully (i.e. its fitness).
Note that both kinds of functional explanations assume that the ultimate driver is ecology. They differ in whether the benefit of having large brains directly affects fitness (animals use their brains to solve fitness-determining problems on an individual trial-and-error basis, and it is this ecological problem-solving that is cognitively demanding) or indirectly via an intermediate social stage (individuals solve these ecological problems socially and it is the social component that is cognitively demanding). The second hypothesis (the social brain hypothesis) comes in two general forms that differ in what they consider to be the rate-limiting effect. One version assumes that it is longevity (or survival) that ultimately limits fitness and that this depends on how well animals can minimize predation risk by social means; the other in effect assumes that it is fertility that limits fitness and that this depends on how well the animals can maximize food intake by social means. Although passive aggregations can be used as a defence against predators (e.g. many ungulates), primates and some other mammalian taxa [7] rely on bonded relationships to ensure that groups stay together (so that individuals can protect each other). Animals might gather together passively in groups in response to foraging conditions, but such groups are invariably unstable. Bondedness appears to be critical in this context, but it remains a frustratingly difficult concept both to define and to study [8,9], even though there is high inter-observer agreement in identifying individuals or species that have bonded relationships (in primates, for example, such relationships are usually marked by frequent, intense social grooming). Occasional criticisms to the contrary notwithstanding, the fact that we do not always understand natural phenomena is not, and of course has never been, a bar to progress in science; indeed, it has often been the spur to novel developments.
If they have tested between competing hypotheses at all, most previous analyses [1,10] have used what amounts to multiple regression analysis to identify the best predictor of brain size. However, such an analysis is appropriate only when the factors included in the analysis are alternative explanations; it is quite inappropriate when they are of different logical and biological standing. The distinction between causes, consequences and constraints in evolutionary explanations is crucial if we are to avoid the kinds of egregious mistakes that have bedevilled much of the literature in this area. Path analysis provides one solution to this problem because it allows different types of relationship to be distinguished. A path analysis with group size as the dependent variable testing between alternative models of the causal relationships linking the different variables that are known to correlate with neocortex size in primates suggests (i) that the best model has activity pattern (nocturnal versus diurnal) and neocortex volume as the only predictors of group size, (ii) that foraging efficiency is a consequence of having a large brain (rather than its cause), and (iii) that the various life history and energetic variables that correlate with brain size are in fact constraints [11]. Predation risk remains the only selection factor for group size according to several analyses [12,13].
More generally, a number of analyses have demonstrated uncontroversially that sociality correlates with absolute or relative brain (or neocortex) size across a wide range of mammalian and avian taxa, and over evolutionary time [7,14–16], though only in the case of primates is there a quantitative relationship between brain size and social group size [15]. Indeed, across the mammals as a whole, those lineages that have encephalized during the course of their evolutionary (i.e. fossil) history are precisely those whose descendent representatives today have bonded social groups [7]. More importantly, Barton [16] has been able to show rather convincingly that, among primates at least, ecological (including tool use) and social functions may depend on neural circuits that have separate foci in the cerebellum and the neocortex, respectively. This is not to say that each of these brain areas is never involved in the other function, but rather to suggest that the primary processing constraints for different tasks may lie in different parts of the brain.
Three additional comments need to be made here. First, although almost all previous analyses have attempted to relativize brain size in some way [16], this has mainly been driven by the demands of reviewers rather than any intrinsic biological or neurological logic. In reality, the same results (and, indeed, often cleaner results) are obtained by using absolute brain component volumes [3]. Second, some care needs to be taken with all of these kinds of analyses, because it cannot be assumed that different species simply represent scaled versions of each other. Gorillas provide a specific case in point: they have a large brain, but relative to other apes, their typical social group size correlates poorly with total brain size because they have an unusually large cerebellum. Instead, gorilla group size is almost exactly what would be predicted by their neocortex size [1,3]. The same is true of orangutans [3]. Third, it has long been understood that the brain functions in a very distributed manner: in humans, for example, the mentalizing skills that appear to underpin social network size involve a specific network of separate neuron bundles in the temporal and frontal lobes [17]. If we wish to test functional hypotheses, then we really need to isolate out the neural networks that are responsible for the function of interest. This is a particular issue with respect to the visual system, because this involves very substantial chunks of neocortex from the dedicated visual areas in the occipital lobe via the visual streams in the parietal lobes to the frontal lobes themselves [16], which are mainly involved in progressively complex processing of visual input. Excluding the brain regions whose primary function is visual processing invariably improves the fit between social indices and brain component volume [3,18] not because it, in some sense, implies that there are brain modules for specific social functions (pace [16]) but because doing so reduces the error variance that results from including brain regions that have well-known, quite specific, mainly low-level processing functions. Our problem is not the lack of understanding of how the brain works, but a desperate lack of sufficiently detailed brain data. Neuroimaging of live subjects is beginning to open up the possibility of undertaking more sophisticated analyses (see later text), but even this exciting new direction has its limitations. Moreover, it is a long way from providing data on a sufficient number of species to allow the kinds of comparative analyses that are the engine of evolutionary explanations.
It is perhaps equally important to be clear that the social brain hypothesis is not really about group size as such: it is about social complexity and the limits placed on animals’ abilities to interact in complex ways [4,7]. A number of studies have reported correlations between neocortex size and various indices of behavioural complexity in primates (for summaries, see [3,4]), but it seems to have been the evolution of ‘bonded’ relationships that proved cognitively so demanding [14,15]. A recent neuroimaging study of captive macaques [19] used neuroimaging to demonstrate that the volumes of core social cognition areas in the neocortex (in the frontal and temporal lobes, and in the amydgala) correlate not only with the size of group in which an individual animal has been housed but also with its dominance rank in the group. Social group size is best viewed as an emergent property of this behavioural complexity (although this point commonly seems to have been misunderstood: for example, see [9]). The social brain hypothesis assumes that some aspect of brain size determines, or sets limits on, core social cognitive abilities, and that it is these in turn that determine the number of relationships (and hence social group size) that animals can effectively maintain as coherent entities. This causal sequence has now been explicitly demonstrated in a neuroimaging study of humans (see below).
One reflection of this kind of behavioural complexity is the fact that the social systems of primates and at least some other mammals actually consist of a hierarchically embedded series of grouping layers [20]. Social interactions in such societies are not homogenous but highly structured, and it is this that creates the layering. More importantly, even though increasing group size is associated with increasing time spent in social interaction (specifically, grooming [21,22]), this does not mean that the animals interact with more individuals within the group. Paradoxically, the reverse is true: increasing group size is actually associated with a decrease in the number of social partners with whom an individual interacts, such that social groups become more highly structured [23,24]. In network terms, density and connectedness decrease, and the group becomes more cliquish. Functionally, this seems to reflect the need to ensure that one's alliances can be relied on as the stresses of group-living increase [4,25]. In these species, the cognitive demands of large groups seem to lie in the capacity to maintain cohesion on the large scale while simultaneously investing one's limited social capital (essentially grooming time) at a more intimate scale [26]. This may itself be associated with the capacity to maintain different relationships (or social layers) for different functional purposes [24,27].
3. Size and structure of human social networks
Humans are as much subject to the constraints of the social brain hypothesis as any other primate, although, with our disproportionately large neocortices, we inevitably have social groups that are considerably larger than those of any other primate. The equation relating ape social group size to relative neocortex size [1,3] predicts a ‘natural’ cognitive community size for humans of around 150 individuals [2]. There is now considerable evidence that groupings of this size occur frequently in human social organization [2,26], and that this is the normative limit on the size of personal social networks among adults [27,28]. More importantly, perhaps, five recent neuroimaging studies have reported correlations between individual differences in social network size and the volumes of core social cognition areas in the cortex and (to a lesser extent) the amygdala [29–33].
Unfortunately, some considerable confusion has been perpetuated in the literature by attempts to suggest that natural human community sizes are significantly larger or smaller than 150. One of these has been to equate the basic form of human social organization in hunter–gatherers with overnight camp groups (often referred to as bands), which typically number 30–50 in size [34,35]. Unfortunately, this claim ignores the fact that these band groupings are unstable and purely ecological in origin—and are clearly embedded in a stable larger community such that band members are drawn only from this community (occasional visitors excepted) [2,36–38]. The other has been to claim that the natural size of human communities must be much greater because we know many more than 150 individuals or because we can list many more individuals as Facebook friends [39]. In the latter case, the issue involves confusion between real social relationships and the capacity to remember faces/names. The limit on memory for faces is indeed much higher than 150, possibly in the order of 1500 individuals [26]. The distinction between these two categories hinges around the difference between friends (those individuals with whom you have a reciprocated relationship, one where knowledge is mutual and there is historical depth to the relationship) and acquaintances (relationships that are more casual or even asymmetrical—you know who they are, but they do not know who you are). This distinction is important in that it seems to differentiate between those who will, all else equal, come to your assistance unconditionally and those who will not (or will do so only on condition that the favour is reciprocated) [40].
All these grouping levels (and others) demonstrably exist, both in human social organization and in personal social networks [36,37]. The substantive issue is not whether one particular level of grouping has precedence or another, but whether there is an observed group size that matches the value of 150 predicted by the primate equations: uncontroversially, this is empirically the case [2,26].
As in the case of non-human primates, the circle of 150 individuals that defines the average human personal social network is not homogenously structured. Rather, it is organized into a series of circles whose sizes scale with a ratio of about three [36,37]. (In their independent analysis of a very large database on hunter–gatherer grouping patterns, Hamilton et al. [37] arrived at a scaling factor of approximately 3.8—a value slightly higher than the 3.2 obtained by Zhou et al. [36]. This is because they anchored their Horton Order analysis on the individual (‘group’ size = 1) rather than the smallest actual group (group size ≈ 5): this has the effect of pulling the left-hand side of the regression line down, yielding a slightly steeper slope [20]. Anchoring the analysis on the smallest true grouping level yields a scaling value of approximately 3 as in Zhou et al. [36]).
The layers identified by these analyses occur at around 5, 15, 50 and 150 individuals, and indeed are known to extend beyond to at least 500 and 1500 [36]. (It is important to remember that these layers are inclusive, so that the 15 includes the inner 5, etc.) These layers can be identified in terms of both perceived emotional closeness and the frequency of interaction [28,41]. The layers can also be identified in terms of individuals’ expressed willingness to behave altruistically towards others [40] and in the functionality of the relationships they support (for more details, see [41]).
In addition to network layer, personal social networks are also differentiated in terms of kinship. In fact, it seems that our networks really consist of two quite separate sub-networks that are completely inter-digitated but which (contra [42]) have quite different dynamics and do not seem to intersect cognitively or behaviourally [40,43]. Kin relationships seem to be immune to failure to interact: emotional closeness to kin remains stable over time (at least across approximately 18 months) whether or not we see the individuals concerned. In contrast, emotional closeness to unrelated friends declines at a rate of about 15 percentage points on an analogue scale of emotional closeness if partners fail to interact over an 18-month period (compared with a gain of approximately 7 percentage points under the same circumstances in the case of kin) [43]. While conventional kin selection theory implies that there is some kind of motivational predisposition to favour kin, one additional proximate reason why kin relationships may be more robust in this respect is that kin networks tend to be more dense (i.e. have more internal interconnections) than friendship networks [44], and this has been shown to play a significant role in mediating altruism even among friends, at least within the inner two network layers [45]. Densely interconnected networks of this kind are probably more susceptible to reputation effects because knowledge about individuals’ behaviour is more likely to be spread around the network when members are interconnected than when the network is of a hub-with-spokes form (as happens more often in friendship networks) [44]. Kinship networks also benefit from the activities of ‘kin keepers’ who make it their business to keep everyone up to date on important events, but there seems to be no regular equivalent in friendship networks [44].
4. Cognitive and behavioural constraints on network size
There is now considerable evidence to suggest that individual differences in human social network size are correlated with some aspect of brain (and, in particular, neocortex) volume [30–33] and, more importantly, that this relationship is mediated by social cognitive competences, as indexed by the ability to handle multi-level intentionality tasks [30,46,47]. While this seems to involve both the temporal lobe and the prefrontal cortex (PFC), it is the PFC (and especially the orbitofrontal PFC) that seems to play a crucial role (see also [48]).
Note that, in contrast to all previous neuroimaging studies of mentalizing (which have focused simply on the question of which brain regions are active when solving false belief (i.e. second-order intentionality) tasks [17,48]), our neuroimaging studies demonstrate a three-way parametric (i.e. quantitative) relationship: the more friends one has, the more orders of intentionality one can handle and the larger are these brain regions [30,33,48]. We do not currently understand what the cognitive processes involved actually are—any more than we really understand what cognitive mechanisms are involved in conventional theory of mind [49], although there is increasing evidence that executive functions play a crucial role (see [48]). Indeed, the orders of intentionality that seem to be so central to understanding individual differences in social network size should probably be interpreted as some kind of index of underlying cognitive abilities rather than a module in itself [3]. Unfortunately, there has been a proliferation of terminology in relation to the mentalizing processes involved, with different authors referring to essentially the same cognitive process as high-order (or multi-level) intentionality [47], collective or group intentionality [48] and ‘we-mode’ processes [50]. All of these involve understanding the mental states of several individuals simultaneously so as to be able to coordinate behaviour strategically. One thing that is clear, however, is that, although memory must at some level be involved, mentalizing abilities are not just a memory problem, or at least not a short-term memory problem: in our neuroimaging studies ([30,33,46]; see also [47]), memory for the facts of the social vignettes is factored out.
Our data point to a critical role for orbitofrontal (but not dorsal) cortex in managing both high-order intentionality and the number of close relationships. Because orbitofrontal cortex (in humans, Brodman areas 10, 11 and 47) is typically associated with reward and emotion-processing, in contrast to dorsal PFC (which is more often associated with rational cognition), this suggests that mentalizing may be an implicit process (see also [48]). This is not, of course, to say that mentalizing may not also be an explicit process, but the precise relationship between explicit and implicit mentalizing, and the neurobiological processes that may be involved have not as yet been investigated in any detail [48].
The especially important role that the human PFC seems to play in this respect has inevitably invited comment on the size of this brain region relative to that of other monkeys and apes [48]. The fact that human frontal lobes scale the same (i.e. are on the same allometric regression line relative to overall brain volume) as those of other apes and monkeys has led some to claim that human frontal lobes are not disproportionately larger than those of other primates [16,51]. However, it is important to appreciate that the underlying relationship is not monotonic: neocortex volume generally, and frontal lobe volume in particular, has a scaling ratio relative to the overall brain size that has a slope significantly greater than b = 1 (see [3]), and this means that human frontal lobes are absolutely (and proportionally to total brain size) much larger than those of other apes.
In addition to this cognitive constraint, the time an individual has available to invest in relationships also seems to be an important constraint on the quality (and, perhaps, quantity) of relationships [27,42]. Time is a critical issue because it turns out that the strength or quality of friendships (as opposed to kin relationships) is a function of the time invested in interactions with that individual: perceived emotional closeness deteriorates when interaction frequency declines, at least in the case of friendships [43].
5. networks in the digital world
SNSs offer the potential to enlarge the size of our social circles because, in principle, they circumvent two of the key constraints that limit our everyday social interactions. One is essentially a knowledge management and memory issue. Part of the problem of maintaining friendships is simply being able to keep up with what they are doing. This is important in that friendships seem to depend critically on shared knowledge and interests [52]. Because at least some of these change over time, maintaining shared knowledge with respect to former partners may be critical in maintaining the quality of the relationship. SNSs allow us to monitor the activities and views of former friends under circumstances where this would not be possible by conventional face-to-face interaction (e.g. because one of the parties has moved away to a location that is sufficiently distant to make regular meetings difficult). The other possibility is a broadcast issue. Conventional social interaction is limited to small numbers of individuals at any one time. Conversation groups, for example, have an upper limit of around four [53], and since almost all human social engagement takes place in conversation groups, this inevitably limits how one's available social time can be distributed around the community as a whole, thereby limiting the number of individuals with whom one can build relationships of a desired intensity [42].
In offline networks, each layer is characterized by a distinct frequency of interaction and a correspondingly distinct sense of emotional closeness [41]; more importantly, declining frequencies of interaction result in declining emotional closeness [43]. Because SNSs (as well as email and texting) allow one to broadcast simultaneously to a much larger group (in the limit, everyone in your address book), they hold out the possibility of breaking through the glass ceiling that otherwise limits the size of face-to-face social networks. By making it possible to interact with the same frequency and intensity with everyone in the network, and even beyond, these two processes might, in principle, allow us to turn all our casual acquaintances into close friends, perhaps even intimates.
Although there has been considerable interest in how the digital world might influence social relationships, the relatively short time-span currently available, combined with the variety of research perspectives motivating those studies that have been done, has yielded a somewhat ambiguous picture. Broadly speaking, two opposing camps have emerged: ‘cyberoptimists’ hold that the Internet's effects have been largely beneficial, whereas ‘cyberpessimists’ argue that, at best, the online social world has not had a significant effect on our social behaviour, and may even have been detrimental.
Cyberoptimists tend to see the Internet as having positive effects on building and maintaining close relationships [54,55], leading to an increase in ‘social capital’ [56–59] and a positive impact on well-being [60–63]. They argue that if maintaining relationships via SNSs is more efficient and cognitively less demanding than other modes of communication, SNS use may allow weak relationships to be maintained at higher levels of trust [64]. Cyberpessimists, by contrast, point out that interactions via digital channels typically tend to be with existing friends; although it obviously does happen occasionally, striking up new satisfying relationships with strangers is relatively rare by comparison [65]. Several studies have shown that use of social media can have a negative impact on social relationships and well-being [66–70]. Nie [69], in particular, has argued that, because time is inelastic, time spent on the Internet inevitably displaces time spent on socializing face-to-face (notably with family members; see also [71]). Other studies have even suggested that use of SNSs can give rise to novel constraints on relationship maintenance, in part due to the difficulties caused by having to manage the flow of information when it is openly available to several different groups of ‘friends’ [72–74].
So does the use of SNSs enlarge one's social network? We undertook two studies to explore this. In one, 115 individuals (63% female, mean age 28 years, 86% students) were asked to complete a lengthy questionnaire detailing all the individuals with whom they felt they had offline relationships, and answered a questionnaire relating to Internet and SNS use, the number of online friends they had, how often they contacted them and their emotional closeness to each listed individual (on a 10-point analogue scale) [75]. In the second study, 270 individuals (80% female, mean age 23 years, 49% students) completed a shorter questionnaire detailing just the people they felt especially close to (‘would greatly regret losing contact with’), frequency of Facebook use, emotional closeness to each listed individual and whether or not they were in a romantic relationship [76]. In neither study was there an association between the use of SNSs and either the size of ‘offline’ social networks or emotional closeness to network members. In fact, Facebook's own data suggest that the claim that large numbers of friends is the norm is, at best, an exaggeration: while the distribution is certainly ‘fat-tailed’, the average and modal number of friends is in fact approximately 130 [http://www.facebook.com/press/info.php?statistics]. Similarly, an analysis of Twitter communities (i.e. the followers of a particular tweeter who interact with each other online) likewise suggests typical community sizes of 100–200 [77].
More importantly, perhaps, our data suggest that digital technology may be especially disadvantageous for men because, unlike women (who maintain their relationships predominantly through conversation), men service their relationships mainly by engaging in joint social or physical activities [43]. In an 18-month longitudinal study of 30 high school students (15 female, average age at start of study 18 years), subjects were asked to rate their frequency of contact (face-to-face or by phone/email etc), the frequency with which they did things together (rating five core social activities) and their emotional closeness to each of the members of their social network at nine month intervals (for details, see [43]). Figure 2 plots the change in mean emotional closeness to individual network members as a function of whether subjects spent more or less time engaged in joint activities versus conversations over the first nine month period (the period when the subjects moved away from home to go to university) for the two sexes separately. There is a striking sex difference in what best allows a relationship to maintain its intensity: for males it is being able to do activities together; for females it is the opportunity to talk together, with opportunities to do things (as opposed to just talk) actually having a negative effect on their relationships. In other words, electronic communication is well suited to women's preferred mode of servicing relationships, but not necessarily to men's who need to interact directly to be able to do so.
Figure 2.
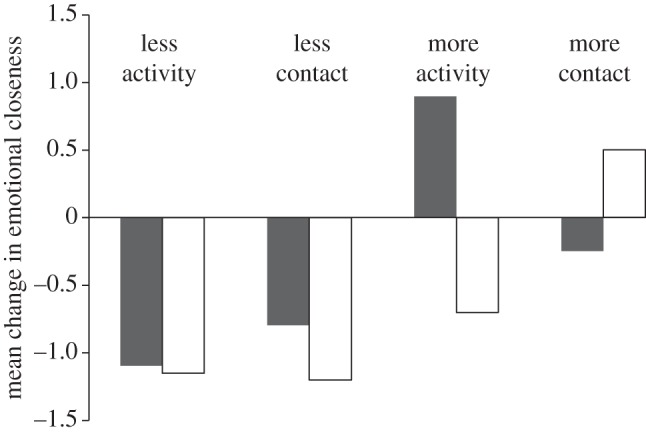
Mean change in self-rated emotional closeness (1–10 analogue scale) towards all unrelated friends for male (filled bars) and female (open bars) subjects as a function of their spending less or more time engaged in activities (shopping, going out, helping, going away) or in conversation (indexed as time since last contact face-to-face or by phone) at month 9 than they had at month 0. Adapted from Roberts & Dunbar [43].
The quality of communication has also been highlighted as a potential limiting factor that might make it difficult to build strong, emotionally intense relationships over digital media [60,68,78,79]. By comparison with face-to-face interaction, most digital media are relatively impoverished because—with the partial exception of Skype—they typically involve only one sensory modality (sound or sight). In a diary study in which 41 adults (61% women, mean age 24.5 years) self-evaluated daily the quality of their individual interactions with their five closest friends through six different modes of communication (face-to-face, telephone, Skype, instant messaging, SNSs and email/texting) over a two-week period, we found that subjects rated face-to-face and Skype interactions significantly more satisfying than interactions via the other media [80]. This suggests that media naturalness (the sense of co-presence and instantaneous feedback) plays an important role in how satisfied we feel with an interaction. Subjects also recorded whether or not laughter occurred (in vivo or virtually in the form of emoticons or other symbolic forms), and the results showed a very striking effect for laughter to positively influence the perceived satisfaction with an interaction, irrespective of medium but especially so in face-to-face interactions (figure 3).
Figure 3.
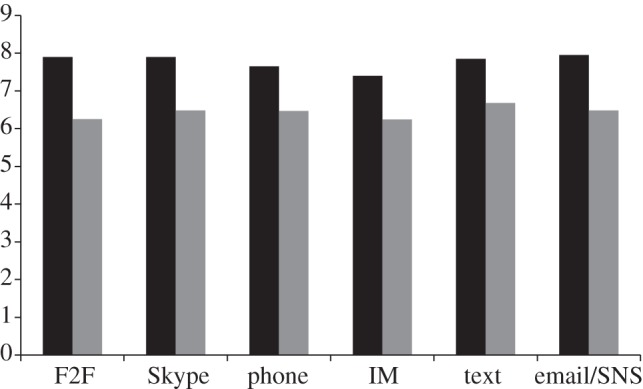
Mean rated happiness score (scale 0–10) for interactions with five best friends through six different communication media as a function of whether laughter occurred during the interaction (black bars) or not (grey bars). Laughter was recorded as both actual or virtual (e.g. the use of emoticons or acronyms such as LOL (‘laugh out loud’)). Interaction satisfaction was rated using Fordyce's [81] Happiness Measure on an 11-point Likert scale. Adapted Vlahovic et al. [80]. F2F, face-to-face; IN, instant messaging.
So how is it that many claim that digital media do yield larger social networks [39,82]? It seems likely that there are two quite separate issues here. One concerns those who list very large numbers (greater than 1000) of friends: we suspect that these are mainly professional users (journalists and artists using Facebook as a fan base or to access information, in effect as a form of Twitter). The other is specific to those who list between 200 and 500 friends: we suspect that these are simply dipping into their 500 layer and including acquaintances rather than true friends. It is important to appreciate that in neither case are meaningful relationships being established. Indeed, Facebook's own data suggest that the number of close relationships (indexed by reciprocated exchanges) does not increase in proportion to the total number of friends listed: users who list only 50 friends typically contact only three to four of them regularly, compared with 10–16 (around three times as many) for those who list 500 or more friends (an order of magnitude greater) [83].
On balance, then, the evidence to date suggests that: (i) the Internet does not significantly increase one's actual social network size (though it can lead to the impression of doing so by allowing one to include acquaintances under the rather muddied rubric of ‘friends’), (ii) it can, nonetheless, be a valuable means of maintaining contact with friends when geographical distance prevents interaction. However, this last point can, in itself, have two adverse consequences. First, time spent on pursuing relationships with old friends that would, in the normal course of events, have withered away through a lack of contact must inevitably detract from time devoted to making new friends, especially after moving to a new location. This may have significant social consequences, because only friends in the physical vicinity can provide serious emotional, social or financial help. It will also have the effect of restricting the size of your local face-to-face network because of the time constraints on the opportunity to make and service new offline relationships. Second, in the contemporary economically mobile environment, this will tend to result in networks becoming fragmented as individuals build successive sets of face-to-face sub-networks at each new location where they spend time, while maintaining old ones online; since geography prevents the interaction of these sub-networks, such networks may lack the level of integration and social cohesion that face-to-face communities necessarily develop precisely because all the members are co-located and know each other. That, in turn, may have consequences for the level of emotional and instrumental support that individuals can call upon.
6. Why the internet wont work … yet
The current technical state of digital media seems to have two distinct drawbacks. First, the lack of a sense of physical co-presence tends to make interactions less satisfying than direct face-to-face interaction. Second, social media sites have, at least until very recently, failed to differentiate between different kinds of relationships. This ambiguity, in part orchestrated by Facebook's use of the term ‘friend’ as a catch-all for any kind of relationship, has been the root cause of the claim that the Internet can broaden our social horizons. This is possible only by failing to recognize those relationships in, say, the 1500 acquaintances layer are of a radically different kind to those in the more intimate 15 or 50 layers. By the same token, we should not confuse media such as Twitter with the capacity to form and service relationships: they are good for coordinating activities or spreading opinions, but that is all. Merely being part of a Twitter-created crowd does not constitute having more friends, no matter how intoxicating it may seem at the time.
In the end, we run up against the fact that relationships depend on two key elements: a cognitive mechanism that creates relationships of trust and obligation, and an emotional component that creates the underpinnings for this [41], just as it does in other social primates [4,26]. Both require time to build, and the level to which they are developed, given the limited time available, ultimately influences their quality (and, hence, the network layer they lie in [41]). In this respect, time does seem to be genuinely inelastic and unsharable: quality relationships require focused time, not the anonymized chatter of a broadcast medium such as Twitter.
One important issue here is that the emotional component of a relationship appears to come through an endorphin surge that, in primates as much as humans, is normally triggered by physical contact (e.g. social grooming) [84,85]. These activities are essentially one-on-one, and cannot easily be generalized to create intense relationships simultaneously between many individuals—except in the special case of laughter. Laughter has been shown to trigger endorphin activation, and to do so especially in groups [86]. It seems to do this by virtue of the fact that relaxed social laughter is physically exhausting for the diaphragm and chest wall muscles, and it is this kind of stress on the body that seems to be so good at triggering endorphins. However, to function as a real bonding device, even laughter is probably constrained to the comparatively modest scale of the conversation group of three to five individuals [25] and may be most effective in face-to-face contexts.
In conclusion, on the evidence available so far, it seems unlikely that the digital media will significantly change our social lives, at least in terms of the number and intensity of different kinds of relationships. Our cognition, inherited as it is from our primate ancestry, seems to make that impossible. This is not to say that the digital media serve no social function, but rather that the function may not be those originally envisaged by their advocates. Indeed, this much is implied by the very recent appearance of SNSs that radically restrict the number of members one can have in one's circle (e.g. Path.com, which limits the number of friends to 50 or 150 in explicit recognition that some social functions can only be supported by a very restricted number of partners [87]). Ultimately, the benefits of the digital revolution seem to be to increase the frequency with which we can continue to interact with those whom we can no longer see so easily face-to-face and in maintaining a form of intermittent, casual ‘poking’ of those we can see but are temporarily separated from (cf. http://en.wikipedia.org/wiki/Path_(social_network)).
References
- 1.Dunbar R. I. M. 1992. Neocortex size as a constraint on group size in primates. J. Hum. Evol. 22, 469–493 10.1016/0047-2484(92)90081-J (doi:10.1016/0047-2484(92)90081-J) [DOI] [Google Scholar]
- 2.Dunbar R. I. M. 1992. Coevolution of neocortex size, group size and language in humans. Behav. Brain Sci. 16, 681–735 10.1017/S0140525X00032325 (doi:10.1017/S0140525X00032325) [DOI] [Google Scholar]
- 3.Dunbar R. I. M. 2011. Evolutionary basis of the social brain. In Oxford handbook of social neuroscience (eds Decety J., Cacioppo J.) pp. 28–28 Oxford, UK: Oxford University Press [Google Scholar]
- 4.Dunbar R. I. M. 2009. Brain and behaviour in primate evolution. In Mind the gap: tracing the origins of human universals (eds Kappeler P. H., Silk J.). pp. 315–330 Berlin, Germany: Springer [Google Scholar]
- 5.Mink J. W., Blumenschine R. J., Adams D. B. 1981. Ratio of central nervous system to body metabolism in vertebrates—its constancy and functional basis. Am. J. Physiol. 241, R203–R212 [DOI] [PubMed] [Google Scholar]
- 6.Karbowski J. 2007. Global and regional brain metabolic scaling and its functional consequences. BMC Biol. 5, 18–46 10.1186/1741-7007-5-18 (doi:10.1186/1741-7007-5-18) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Shultz S., Dunbar R. I. M. 2010. Encephalisation is not a universal macroevolutionary phenomenon in mammals but is associated with sociality. Proc. Natl Acad. Sci. USA 107, 21 582–21 586 10.1073/pnas.1005246107 (doi:10.1073/pnas.1005246107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Silk J. B. 2002. The ‘F’-word in primatology. Behaviour 139, 421–446 10.1163/156853902760102735 (doi:10.1163/156853902760102735) [DOI] [Google Scholar]
- 9.Dunbar R. I. M., Shultz S. 2010. Bondedness and sociality. Behaviour 147, 775–803 10.1163/000579510X501151 (doi:10.1163/000579510X501151) [DOI] [Google Scholar]
- 10.Reader S. M., Hager Y., Laland K. N. 2011. The evolution of primate general and cultural intelligence. Phil. Trans. R. Soc. B 366, 1017–1027 10.1098/rstb.2010.0342 (doi:10.1098/rstb.2010.0342) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dunbar R. I. M., Shultz S. 2007. Understanding primate brain evolution. Phil. Trans. R. Soc. B 362, 649–658 10.1098/rstb.2006.2001 (doi:10.1098/rstb.2006.2001) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shultz S., Noe R., McGraw S., Dunbar R. I. M. 2004. A community-level evaluation of the impact of prey behavioural and ecological characteristics on predator diet composition. Proc. R. Soc. B 271, 725–732 10.1098/rspb.2003.2626 (doi:10.1098/rspb.2003.2626) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shultz S., Finlayson L. V. 2010. Predator diet choices are associated with prey life history and behavioural characteristics in terrestrial mammal communities. Behav. Ecol. 21, 1073–1079 10.1093/beheco/arq108 (doi:10.1093/beheco/arq108) [DOI] [Google Scholar]
- 14.Pérez-Barbería J., Shultz S., Dunbar R. I. M. 2007. Evidence for intense coevolution of sociality and brain size in three orders of mammals. Evolution 61, 2811–2821 10.1111/j.1558-5646.2007.00229.x (doi:10.1111/j.1558-5646.2007.00229.x) [DOI] [PubMed] [Google Scholar]
- 15.Shultz S., Dunbar R. I. M. 2007. The evolution of the social brain: Anthropoid primates contrast with other vertebrates. Proc. R. Soc. B 274, 2429–2436 10.1098/rspb.2007.0693 (doi:10.1098/rspb.2007.0693) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Barton R. A. 2012. Embodied cognitive evolution and the cerebellum. Phil. Trans. R. Soc. B 367, 2097–2107 10.1098/rstb.2012.0112 (doi:10.1098/rstb.2012.0112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gallagher H. L., Frith C. D. 2003. Functional imaging of ‘theory of mind’. Trends Cogn. Sci. 7, 77–83 10.1016/S1364-6613(02)00025-6 (doi:10.1016/S1364-6613(02)00025-6) [DOI] [PubMed] [Google Scholar]
- 18.Joffe T. R., Dunbar R. I. M. 1997. Visual and socio-cognitive information processing in primate brain evolution. Proc. R. Soc. Lond. B 264, 1303–1307 10.1098/rspb.1997.0180 (doi:10.1098/rspb.1997.0180) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sallet J., et al. 2011. Social network size affects neural circuits in macaques. Science 334, 697–700 10.1126/science.1210027 (doi:10.1126/science.1210027) [DOI] [PubMed] [Google Scholar]
- 20.Hill R. A., Bentley A., Dunbar R. I. M. 2008. Network scaling reveals consistent fractal pattern in hierarchical mammalian societies. Biol. Lett. 4, 748–751 10.1098/rsbl.2008.0393 (doi:10.1098/rsbl.2008.0393) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lehmann J., Korstjens A. H., Dunbar R. I. M. 2007. Group size, grooming and social cohesion in primates. Anim. Behav. 74, 1617–1629 10.1016/j.anbehav.2006.10.025 (doi:10.1016/j.anbehav.2006.10.025) [DOI] [Google Scholar]
- 22.Dunbar R. I. M. 1991. Functional significance of social grooming in primates. Folia Primatol. 57, 121–131 10.1159/000156574 (doi:10.1159/000156574) [DOI] [Google Scholar]
- 23.Kudo H., Dunbar R. I. M. 2001. Neocortex size and social network size in primates. Anim. Behav. 62, 711–722 10.1006/anbe.2001.1808 (doi:10.1006/anbe.2001.1808) [DOI] [Google Scholar]
- 24.Lehmann J., Dunbar R. I. M. 2009. Network cohesion, group size and neocortex size in female-bonded Old World primates. Proc. R. Soc. B 276, 4417–4422 10.1098/rspb.2009.1409 (doi:10.1098/rspb.2009.1409) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dunbar R. I. M. 2012. Social cognition on the Internet: testing constraints on social network size. Phil. Trans. R. Soc. B 367, 2192–2201 10.1098/rstb.2011.0217 (doi:10.1098/rstb.2011.0217) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dunbar R. I. M. 2008. Mind the gap: or why humans aren't just great apes. Proc. Br. Acad. 154, 403–423 [Google Scholar]
- 27.Roberts S. B. G., Dunbar R. I. M., Pollet T., Kuppens T. 2009. Exploring variations in active network size: constraints and ego characteristics. Soc. Netw. 31, 138–146 10.1016/j.socnet.2008.12.002 (doi:10.1016/j.socnet.2008.12.002) [DOI] [Google Scholar]
- 28.Hill R. A., Dunbar R. I. M. 2003. Social network size in humans. Hum. Nat. 14, 53–72 10.1007/s12110-003-1016-y (doi:10.1007/s12110-003-1016-y) [DOI] [PubMed] [Google Scholar]
- 29.Bickart K., Wright C., Dautoff R., Dickerson B., Feldman Barrett L. 2010. Amygdala volume and social network size in humans. Nat. Neurosci. 14, 163–164 10.1038/nn.2724 (doi:10.1038/nn.2724) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lewis P., Rezaie R., Browne R., Roberts N., Dunbar R. I. M. 2011. Ventromedial prefrontal volume predicts understanding of others and social network size. NeuroImage 57, 1624–1629 10.1016/j.neuroimage.2011.05.030 (doi:10.1016/j.neuroimage.2011.05.030) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Horváth K., Martos J., Mihalik B., Bódizs R. 2011. Is the social brain theory applicable to human individual differences? Relationship between sociability personality dimension and brain size. Evol. Psychol. 9, 244–257 [PubMed] [Google Scholar]
- 32.Kanai R., Bahrami B., Roylance R., Rees G. 2011. Online social network size is reflected in human brain structure. Proc. R. Soc. B 279, 1327–1334 10.1098/rspb.2011.1959 (doi:10.1098/rspb.2011.1959) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Powell J., Lewis P. A., Roberts N., García-Fiñana M., Dunbar R. I. M. In press Orbital prefrontal cortex volume predicts social network size: an imaging study of individual differences in humans. Proc. R. Soc. B 279, 2157–2162 10.1098/rspb.2011.2574 (doi:10.1098/rspb.2011.2574) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Marlowe F. 2005. Hunter-gatherers and human evolution. Evol. Anthropol. 14, 54–67 10.1002/evan.20046 (doi:10.1002/evan.20046) [DOI] [PubMed] [Google Scholar]
- 35.de Ruiter J., Weston G., Lyon S. M. 2011. Dunbar's Number: group size and brain physiology in humans re-examined. Am. Anthropol. 113, 557–568 10.1111/j.1548-1433.2011.01369.x (doi:10.1111/j.1548-1433.2011.01369.x) [DOI] [PubMed] [Google Scholar]
- 36.Zhou W-X., Sornette D., Hill R. A., Dunbar R. I. M. 2005. Discrete hierarchical organization of social group sizes. Proc. R. Soc. B 272, 439–444 10.1098/rspb.2004.2970 (doi:10.1098/rspb.2004.2970) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hamilton M. J., Milne B. T., Walker R. S., Burger O., Brown J. H. 2007. The complex structure of hunter–gatherer social networks. Proc. R. Soc. B 274, 2195–2202 10.1098/rspb.2007.0564 (doi:10.1098/rspb.2007.0564) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Korstjens A. H., Layton R., van Schaik C. P., Boesch C., Dunbar R. I. M. 2008. Implications of fission–fusion dynamics for the evolution of complex human societies. Curr. Anthropol. 49, 638–640 [Google Scholar]
- 39.Wellman B. 2012. Is Dunbar's number up? Br. J. Psychol . 103, 174–176 10.1111/j.2044-8295.2011.02075.x (doi:10.1111/j.2044-8295.2011.02075.x) [DOI] [PubMed] [Google Scholar]
- 40.Curry O., Roberts S. B. G., Dunbar R. I. M. In press. Altruism in networks: evidence for a kin premium. Br. J. Psychol . [DOI] [PubMed] [Google Scholar]
- 41.Sutcliffe A. J., Dunbar R. I. M., Binder J., Arrow H. 2012. Relationships and the social brain: integrating psychological and evolutionary perspectives. Br. J. Psychol. 103, 149–168 10.1111/j.2044-8295.2011.02061.x (doi:10.1111/j.2044-8295.2011.02061.x) [DOI] [PubMed] [Google Scholar]
- 42.Ackerman J. M., Kenrick D. T., Schaller M. 2007. Is friendship akin to kinship? Evol. Hum. Behav. 28, 365–374 10.1016/j.evolhumbehav.2007.04.004 (doi:10.1016/j.evolhumbehav.2007.04.004) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Roberts S. B. G., Dunbar R. I. M. 2011. The costs of family and friends: an 18-month longitudinal study of relationship maintenance and decay. Evol. Hum. Behav. 32, 186–197 10.1016/j.evolhumbehav.2010.08.005 (doi:10.1016/j.evolhumbehav.2010.08.005) [DOI] [Google Scholar]
- 44.Roberts S. B. G. 2009. Constraints on social networks. In Social brain, distributed mind (eds Dunbar R. I. M., Gamble C., Gowlett J. A. J.), pp. 117–138 Oxford, UK: Oxford University Press [Google Scholar]
- 45.Curry O., Dunbar R. I. M. 2011. Altruism in networks: the effect of connections. Biol. Lett. 3, 651–653 10.1098/rsbl.2010.1202 (doi:10.1098/rsbl.2010.1202) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Powell J., Lewis P., Dunbar R. I. M., García-Fiñana M., Roberts N. 2010. Orbital prefrontal cortex volume correlates with social cognitive competence. Neuropsychologia 48, 3554–3562 10.1016/j.neuropsychologia.2010.08.004 (doi:10.1016/j.neuropsychologia.2010.08.004) [DOI] [PubMed] [Google Scholar]
- 47.Stiller J., Dunbar R. I. M. 2007. Perspective-taking and memory capacity predict social network size. Soc. Netw. 29, 93–104 10.1016/j.socnet.2006.04.001 (doi:10.1016/j.socnet.2006.04.001) [DOI] [Google Scholar]
- 48.Frith C. D. 2012. The role of metacognition in human social interactions. Phil. Trans. R. Soc. B 367, 2213–2223 10.1098/rstb.2012.0123 (doi:10.1098/rstb.2012.0123) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Roth D., Leslie A. M. 1998. Solving belief problems: toward a task analysis. Cognition 66, 1–31 10.1016/S0010-0277(98)00005-5 (doi:10.1016/S0010-0277(98)00005-5) [DOI] [PubMed] [Google Scholar]
- 50.Tomasello M. 2009. Why we cooperate. Cambridge, MA: The MIT Press [Google Scholar]
- 51.Semendeferi K., Armstrong E., Schleicher A., Zilles K., Van Hoesen G. W. 2001. Prefrontal cortex in humans and apes: a comparative study of area 10. Am. J. Phys. Anthropol. 114, 224–241 10.1002/1096-8644(200103)114:3<224::AID-AJPA1022>3.0.CO;2-I (doi:10.1002/1096-8644(200103)114:3<224::AID-AJPA1022>3.0.CO;2-I) [DOI] [PubMed] [Google Scholar]
- 52.McPherson M., Smith-Lovin L., Cook J. M. 2001. Birds of a feather: homophily in social networks. Annu. Rev. Sociol. 27, 415–444 10.1146/annurev.soc.27.1.415 (doi:10.1146/annurev.soc.27.1.415) [DOI] [Google Scholar]
- 53.Dunbar R. I. M., Duncan N., Nettle D. 1995. Size and structure of freely forming conversational groups. Hum. Nat. 6, 67–78 10.1007/BF02734136 (doi:10.1007/BF02734136) [DOI] [PubMed] [Google Scholar]
- 54.Bargh J. A., McKenna K. Y. A. 2004. The internet and social life. Annu. Rev. Psychol. 55, 573–590 10.1146/annurev.psych.55.090902.141922 (doi:10.1146/annurev.psych.55.090902.141922) [DOI] [PubMed] [Google Scholar]
- 55.Valkenburg P., Peter J. 2009. Social consequences of the internet for adolescents: a decade of research. Curr. Dir. Psychol. Sci. 18, 1–5 10.1111/j.1467-8721.2009.01595.x (doi:10.1111/j.1467-8721.2009.01595.x) [DOI] [Google Scholar]
- 56.Ellison N. B., Steinfield C., Lampe C. 2007. The benefits of Facebook ‘friends:’ social capital and college students’ use of online social network sites. J. Comput. Mediat. Commun. 12, 1143–1168 10.1111/j.1083-6101.2007.00367.x (doi:10.1111/j.1083-6101.2007.00367.x) [DOI] [Google Scholar]
- 57.Tong S. T., Van Der Heide B., Langwell L., Walther J. B. 2008. Too much of a good thing? The relationship between number of friends and interpersonal impressions on Facebook. J. Comput. Mediat. Commun. 13, 531–549 10.1111/j.1083-6101.2008.00409.x (doi:10.1111/j.1083-6101.2008.00409.x) [DOI] [Google Scholar]
- 58.Valenzuela S., Park N., Kee K. F. 2009. Is there social capital in a social network site? Facebook use and college students' life satisfaction, trust, and participation. J. Comput. Mediat. Commun. 14, 875–901 10.1111/j.1083-6101.2009.01474.x (doi:10.1111/j.1083-6101.2009.01474.x) [DOI] [Google Scholar]
- 59.Zywica J., Danowski J. 2008. The faces of Facebookers: investigating social enhancement and social compensation hypotheses; predicting Facebook™ and offline popularity from sociability and self esteem, and mapping the meanings of Popularity with semantic networks. J. Comput. Mediat. Commun. 14, 1–34 10.1111/j.1083-6101.2008.01429.x (doi:10.1111/j.1083-6101.2008.01429.x) [DOI] [Google Scholar]
- 60.Boneva B. S., Quinn A., Kraut R. E., Kiesler S., Shklovski I. 2006. Teenage communication in the instant messaging era. In Computers, phones, and the internet: domesticating information technology (eds Kraut R., Brynin M., Kiesler S.). New York, NY: Oxford University Press [Google Scholar]
- 61.LaRose R., Eastin M. S., Gregg J. 2001. Reformulating the internet paradox: social cognitive explanations of internet use and depression. J. Online Behav. 1 http://www.behavior.net/JOB/v1n2/paradox.html [Google Scholar]
- 62.Liu X., LaRose R. 2008. Does using the internet make people more satisfied with their lives? The effects of the internet on college students’ school life satisfaction. CyberPsychol. Behav. 11, 310–320 10.1089/cpb.2007.0040 (doi:10.1089/cpb.2007.0040) [DOI] [PubMed] [Google Scholar]
- 63.Valkenburg P., Peter J. 2007. Online communication and adolescent well-being: testing the stimulation versus the displacement hypothesis. J. Comput. Mediat. Commun. 12, 1169–1182 10.1111/j.1083-6101.2007.00368.x (doi:10.1111/j.1083-6101.2007.00368.x) [DOI] [Google Scholar]
- 64.Donath J. 2008. Signals in social supernets. J. Comput. Mediat. Commun. 13, 231–251 10.1111/j.1083-6101.2007.00394.x (doi:10.1111/j.1083-6101.2007.00394.x) [DOI] [Google Scholar]
- 65.Boyd D. M., Ellison N. B. 2008. Social network sites: definition, history and scholarship. J. Comput. Mediat. Commun. 13, 210–238 10.1111/j.1083-6101.2007.00393.x (doi:10.1111/j.1083-6101.2007.00393.x) [DOI] [Google Scholar]
- 66.Schiffrin H., Edelman A., Falkenstern M., Stewart C. 2009. The associations among computer-mediated communication, relationships, and well-being. Cyberpsychol. Behav. Soc. Netw. 13, 299–306 10.1089/cyber.2009.0173 (doi:10.1089/cyber.2009.0173) [DOI] [PubMed] [Google Scholar]
- 67.van den Eijnden R., Meerkerk G. J., Vermulst A. A., Spijkerman R., Engels R. 2008. Online ommunication, compulsive internet use, and psychosocial well-being among adolescents: a longitudinal study. Dev. Psychol. 44, 655–665 10.1037/0012-1649.44.3.655 (doi:10.1037/0012-1649.44.3.655) [DOI] [PubMed] [Google Scholar]
- 68.Wolak J., Mitchell K. J., Finkelhor D. 2003. Escaping or connecting? Characteristics of youth who form close online relationships. J. Adolescence 26, 105–119 10.1016/S0140-1971(02)00114-8 (doi:10.1016/S0140-1971(02)00114-8) [DOI] [PubMed] [Google Scholar]
- 69.Nie N. H. 2001. Sociability, interpersonal relations, and the internet: reconciling conflicting findings. Am. Behav. Sci. 45, 420–435 10.1177/00027640121957277 (doi:10.1177/00027640121957277) [DOI] [Google Scholar]
- 70.Mesch G. S. 2006. Family relations and the internet: exploring a family boundaries approach. J. Family Commun. 6, 119–138 10.1207/s15327698jfc0602_2 (doi:10.1207/s15327698jfc0602_2) [DOI] [Google Scholar]
- 71.Nie N. H., Hillygus D. S. 2002. The impact of internet use on sociability: time-diary findings. IT Soc. 1, 1–20 [Google Scholar]
- 72.Binder J., Howes A., Sutcliffe A. 2009. The problem of conflicting social spheres: effects of network structure on experienced tension in social network sites. In Proc. 27th Int. Conf. on Human Factors in Computing Systems, Boston, MA, 4–9 April 2009, pp. 965–974 New York, NY: ACM [Google Scholar]
- 73.Joinson A. N., Houghton D. J., Vasalou A., Marder B. L. 2011. Digital crowding: privacy, self-disclosure and technology. In Privacy online: perspectives on privacy and self-disclosure in the social web (eds Trepte S., Reinecke L.), pp. 31–44 New York, NY: Springer [Google Scholar]
- 74.Houghton D. J., Joinson A. N. 2010. Privacy, social network sites, and social relations. J. Technol. Hum. Serv. 28, 74–94 10.1080/15228831003770775 (doi:10.1080/15228831003770775) [DOI] [Google Scholar]
- 75.Pollet T. V., Roberts S. G. B., Dunbar R. I. M. 2011. Use of social network sites and instant messaging does not lead to increased offline social network size, or to emotionally closer relationships with offline network members. Cyberpsychol. Behav. Soc. Netw. 14, 253–258 10.1089/cyber.2010.0161 (doi:10.1089/cyber.2010.0161) [DOI] [PubMed] [Google Scholar]
- 76.Veldkamp C., Pollet T., Roberts S. B. G., Dunbar R. I. M. Submitted Active communication over Facebook is not associated with larger offline social networks, or with emotionally closer relationships with network members. [Google Scholar]
- 77.Gonçalves B., Perra N., Vespignani A. 2011. Validation of Dunbar's number in Twitter conversations. PLoS ONE 6, e22656 10.1371/journal.pone.0022656 (doi:10.1371/journal.pone.0022656) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Cummings N. J., Butler B., Kraut R. 2002. The quality of online social relationships. Commun. ACM 45, 103–108 10.1145/514236.514242 (doi:10.1145/514236.514242) [DOI] [Google Scholar]
- 79.Gross E. F., Juvonen J., Gable S. L. 2002. Internet use and well-being in adolescence. J. Soc. Issues 58, 75–90 10.1111/1540-4560.00249 (doi:10.1111/1540-4560.00249) [DOI] [Google Scholar]
- 80.Vlahovic T., Roberts S. B. G., Dunbar R. I. M. In press Effects of duration and laughter on subjective happiness within different modes of communication. J. Comp. Med. Commun. [Google Scholar]
- 81.Fordyce M. W. 1988. A review of research on the happiness measures: a sixty second index of happiness and mental health. Soc. Indicators Res. 20, 355–381 10.1007/BF00302333 (doi:10.1007/BF00302333) [DOI] [Google Scholar]
- 82.Boase J. 2008. Personal networks and the personal communication system: using multiple communication media to connect with personal networks. Inform. Commun. Soc. 11, 490–508 10.1080/13691180801999001 (doi:10.1080/13691180801999001) [DOI] [Google Scholar]
- 83.Marlow C. 2011. Maintained relationships on Facebook. See http://overstated.net/. [Google Scholar]
- 84.Dunbar R. I. M. 2009. The social role of touch in humans and primates: behavioural function and neurobiological mechanisms. Neurosci. Biobehav. Rev. 34, 260–268 10.1016/j.neubiorev.2008.07.001 (doi:10.1016/j.neubiorev.2008.07.001) [DOI] [PubMed] [Google Scholar]
- 85.Machin A., Dunbar R. I. M. 2011. The brain opioid theory of social attachment: a review of the evidence. Behaviour 148, 985–1025 10.1163/000579511X596624 (doi:10.1163/000579511X596624) [DOI] [Google Scholar]
- 86.Dunbar R. I. M., et al. 2011. Social laughter is correlated with an elevated pain threshold. Proc. R. Soc. B 279, 1161–1167 10.1098/rspb.2011.1373 (doi:10.1098/rspb.2011.1373) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Horst H., Miller D. 2005. From kinship to link-up: cell phones and social networking in Jamaica. Curr. Anthropol. 45, 755–758 10.1086/432650 (doi:10.1086/432650) [DOI] [Google Scholar]