

Abstract
Eight isoline colonies of Anopheles peditaeniatus Leicester (Diptera: Culicidae) were established from wild-caught females collected from buffalo-baited traps at 8 localities in Thailand. They showed 2 types of X (X2, X3) and 4 types of Y (Y2, Y3, Y4, Y5) chromosomes based on the number and amount of major block(s) of heterochromatin present in the heterochromatic arm, and were tentatively designated as Forms B (X2, X3, Y2), C (X3, Y3), D (X3, Y4) and E (X2, X3, Y5). Form B was found in Nan, Ratchaburi, and Chumphon provinces; Form C was obtained in Chon Buri province; Form D was recovered in Kamphaeng Phet province; and Form E was acquired in Chiang Mai, Udon Thani, and Ubon Ratchathani provinces. Crossing studies among the 8 isoline colonies, which were representative of 4 karyotypic forms of An. peditaeniatus, revealed genetic compatibility in providing viable progenies and synaptic salivary gland polytene chromosomes through F2-generations, thus suggesting the conspecific nature of these karyotypic forms. These results were supported by the very low intraspecific sequence variations (0.0 – 1.1%) of the nucleotide sequences in ribosomal DNA (ITS2) and mitochondrial DNA (COI and COII) of the 4 forms.
Keywords : COI, COII, crossing experiment, Hyrcanus group, ITS2, metaphase karyotype
Introduction
The Hyrcanus group of the Myzorhynchus series of the subgenus Anopheles (Diptera: Culicidae) comprises a large number of species that occur widely in Asia. At least 8 species of this group, i.e. Anopheles argyropus Swellengrebel, An. crawfordi Reid, An. nigerrimus Gilles, An. nitidus Harrison, Scanlon and Reid, An. paraliae Sandosham, An. peditaeniatus Leicester, An. pursati Laveran, and An. sinensis Wiedemann are recorded in Thailand (Harrison and Scanlon 1975; Rattanarithikul et al. 2006). Among these, An. nigerrimus, An. peditaeniatus, and An. sinensis are suspected as vectors of Plasmodium vivax Grassi and Feletti in Thailand (Harrison and Scanlon 1975; Rattanarithikul et al. 1996), while An. sinensis has been incriminated as a natural vector of P. vivax in Korea (Chai 1999; Ree et al. 2001) and An. peditaeniatus as a secondary vector of Japanese encephalitis virus in China and India (Mourya et al. 1989; Zhang 1990; Kanojia et al. 2003). Although An. peditaeniatus has been found abundantly and widely distributed throughout Thailand, its status as a vector of the Japanese encephalitis virus remains a crucial question that needs to be clarified more thoroughly. Additionally, this species was also considered an economic pest of cattle because of its vicious biting-behavior and ability to transmit cervid filariae of the genus Setaria (Reid 1968; Harrison and Scanlon 1975).
Regarding the cytogenetic investigations of An. peditaeniatus, the results indicated that at least 3 types of X (X1, X2, X3) and 5 types of Y (Y1, Y2, Y3, Y4, Y5) chromosomes were found in both sympatric and/or allopatric populations in Chanthaburi, Chiang Mai and Phrae provinces (Baimai et al. 1993).
Chromosomes X1, X2 and X3 differ from each other in the number and amount of major block(s) of heterochromatin present in the heterochromatic arm, making them appear as metacentric X1, small submetacentric X2, and large submetacentric X3 chromosomes. Likewise, the evolution of Y chromosome types, i.e. very small telocentric Y1, medium telocentric Y2, large telocentric Y3, very large telocentric Y2, and submetacentric Y5 could have arisen via the process of gain, rather than loss, of major block(s) of heterochromatin (Baimai et al. 1993; Baimai 1998). Although marked genetic variation at the chromosomal level of An. peditaeniatus has obviously been illustrated, little is known about its genetic proximities. Accordingly, the chromosomal variant and/or distinction might be manifested as an important role in generating post-mating barrier and DNA sequence variation of some specific genomic regions. Thus, this paper presents the results of crossing experiments and comparative DNA sequencing of the ribosomal DNA (ITS2) and mitochondrial DNA (COI and COII) regions of 4 karyotypic forms of An. peditaeniatus strains from 8 localities in Thailand.
Materials and Methods
Field collections and the establishment of isoline colonies
Wild, fully engorged female An. peditaeniatus were collected from buffalo-baited traps from November 2007 to September 2008 at 8 localities in Thailand (Figure 1; Table 1). Eight isoline colonies were successfully established and maintained in an insectary using the techniques described by Kim et al. (2003). These isoline colonies were used for studies on metaphase karyotypes, crossing experiments, and molecular analyses.
Figure 1.

Map of Thailand showing 8 locations where mosquitoes were collected; northern region: Chiang Mai and Nan provinces; northeastern region: Udon Thani and Ubon Ratchathani provinces; central region: Kamphaeng Phet province; western region: Ratchaburi province; eastern region: Chonburi province; and southern region: Chumporn province. High quality figures are available online.
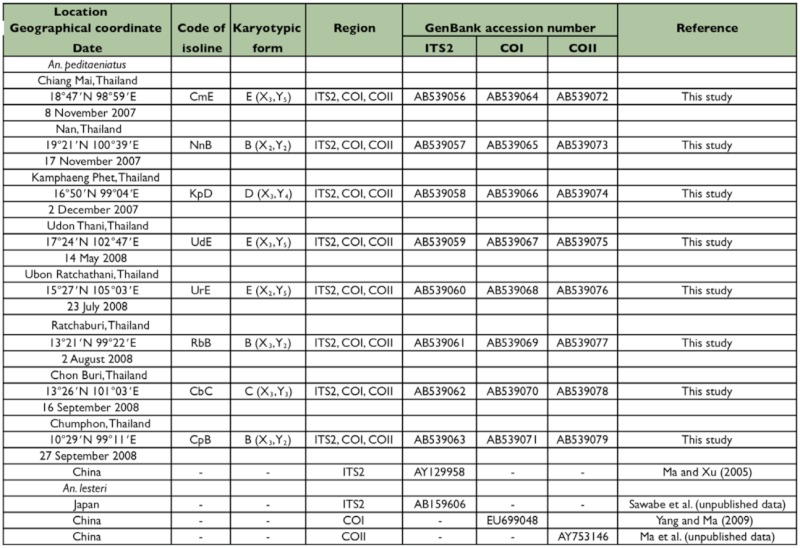
Table 1.
Locations, isoline colonies and karyotypic forms of Anopheles peditaeniatus and An. lesteri, and their GenBank accession numbers.
Metaphase chromosome preparation
Metaphase chromosome preparation was the technique used for chromosome preparation in adult mosquitoes, as described by Choochote et al. (2001). Briefly, newly emerged adult males of laboratory-raised An. peditaeniatus (aged about 6–12 hr) were intra-thoracically inoculated with 0.30 µl of 1% ethanol-extracted Gloriosa superba L. (Liliales: Colchicaceae) solution and held in an insectary at 27 ± 2°C, with 70–80% relative humidity for 3 hr. The excised testes were incubated in 1% hypotonic sodium citrate solution, fixed in Carnoy's fixative, stained with 10% Giemsa in phosphate buffer pH 7.2, mounted in Permount® (Fisher, www.fishersci.com), and examined under a compound microscope. Identification of types of sex chromosomes followed the cytotaxonomic key of Baimai et al. (1993).
Crossing experiments
In crossing experiments, the 8 laboratoryraised isoline colonies of An. peditaeniatus were representative of the 4 karyotypic forms, i.e. Forms B [Nan strain: NnB (X2, Y2), Ratchaburi strain: RbB (X3, Y2), Chumphon strain: CpB (X3, Y2)], C [Chon Buri strain: CbC (X3, Y3)], D [Kamphaeng Phet: KpD (X3, Y4)], and E [Chiang Mai strain: CmE (X3, Y5), Udon Thani strain: UdE (X3, Y5), Ubon Ratchathani strain: UrE (X2, Y5)] (Table 1). These isoline colonies were used for crossing experiments in order to determine post-mating reproductive isolation by employing the techniques previously reported by Thongsahuan et al. (2009). Experiments were carried out once for each crossing of karyotypic forms. The salivary gland polytene chromosomes of 4th instar larvae from the crosses were investigated using the techniques described by Kanda (1979).
DNA extraction, amplification, sequencing and analysis
One individual F1-progeny adult female from each isoline colony of An. peditaeniatus forms was used for DNA extraction and amplification. Genomic DNA was extracted from individual adult mosquitoes using a RED Extract-N-Amp™ Tissue PCR Kit (Sigma-Aldrich). The ribosomal DNA (rDNA) internal transcribed spacer 2 (ITS2), and mitochondrial cytochrome c oxidase subunit I (COI) and subunit II (COII) were amplified using the primers described by Park et al. (2003), with minor modifications: 5.8S + 35 (5′-ACG CAT ATT GCA CGT CGT GG-3′) and 28S - 20 (5′-GGG TTG TCA CAC ATA ACT TGA GGC-3′) for ITS2; LCO1490 (5′GGT CAA CAA ATC ATA AAG ATA TTG G-3′) and HCO2198 (5′-TAA ACT TCA GGG TGA CCA AAA AAT CA-3′) for COI; AnoCO2+1 (5′-GAT TAG TGC AAT GAA TTT AAG C-3′) and AnoCO2END (5′-GAG ATC ATT ACT TGC TTT CAG TC-3′) for COIL The PCR condition, cloning, and sequencing followed the techniques previously reported by Park et al. (2008). The PCR products were purified using the QIAquick® Gel Extraction Kit (Qiagen, www.qiagen.com), and directly sequenced with an ABI PRISMH® 3700 DNA Analyzer (Applied Biosystems, www.appliedbiosystems.com) using a Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems). Both strands were sequenced and aligned using the ClustalX multiple alignment programs (Thompson et al. 1997). The nucleotide sequence data reported are in the DDBJ/EMBL/GenBank nucleotide sequence databases with the accession numbers AB539056-AB539079. Geographical type of specimens and their sequence accession numbers within GenBank are denoted in Table 1.
Results
Cytological observations of F1-progenies of 8 isoline colonies demonstrated 2 types of X (X2, X3) and 4 types of Y (Y2, Y3, Y4, Y5) chromosomes. Based on uniquely different characteristics of Y chromosome from each isoline colony, they were tentatively designated as Forms B (X2, X3, Y2), C (X3, Y3), D (X3, Y4), and E (X2, X3, Y5). Form B was detected in 3 isoline colonies from Nan (X2, Y2), Ratchaburi (X3, Y2), and Chumphon (X3, Y2) provinces. Form C was found in 1 isoline colony from Chonburi (X3, Y3) province. Form D was obtained in 1 isoline colony from Kamphaeng Phet (X3, Y4) province. Form E was recovered in 3 isoline colonies from Chiang Mai (X3, Y5), Udon Thani (X3, Y5) and Ubon Ratchathani (X2, Y5) provinces (Figure 2; Table 1).
Figure 2.

Metaphase karyotypes of Anopheles peditaeniatus. Form B: (A) Nan strain: showing X2, Y2 chromosomes; (B) Ratchaburi strain: showing X3, Y2 chromosomes; (C) Chumphon strain: showing X3, Y2 chromosomes. Form C: (D) Chonburi strain: showing X3, Y3 chromosomes. Form D: (E) Kamphaeng Phet strain: showing X3, Y4 chromosomes. Form E: (F) Chiang Mai strain: showing X3, Y5 chromosomes; (G) Udon Thani strain: showing X3, Y5 chromosomes; (H) Ubon Rathchathani strain: showing X2, Y5 chromosomes. High quality figures are available online.
For crossing experiments, details of hatchability, pupation, emergence, and adult sex-ratio of parental, reciprocal, and F1-hybrid crosses among the 8 isoline colonies of An. peditaeniatus Forms B (X2, X3, Y2), C (X3, Y3), D (X3, Y4), and E (X2, X3, Y5) are shown in Table 2. All crosses yielded viable progenies through F2-generations. No evidence of genetic incompatibility and/or post-mating reproductive isolation was observed among these crosses. The salivary gland polytene chromosomes of the 4th stage larvae from all crosses showed complete synapsis along the whole length of all autosomes and the X chromosome (Figure 3).
Table 2.
Crossing experiments among the 8 isoline colonies of Anopheles peditaeniatus forms.
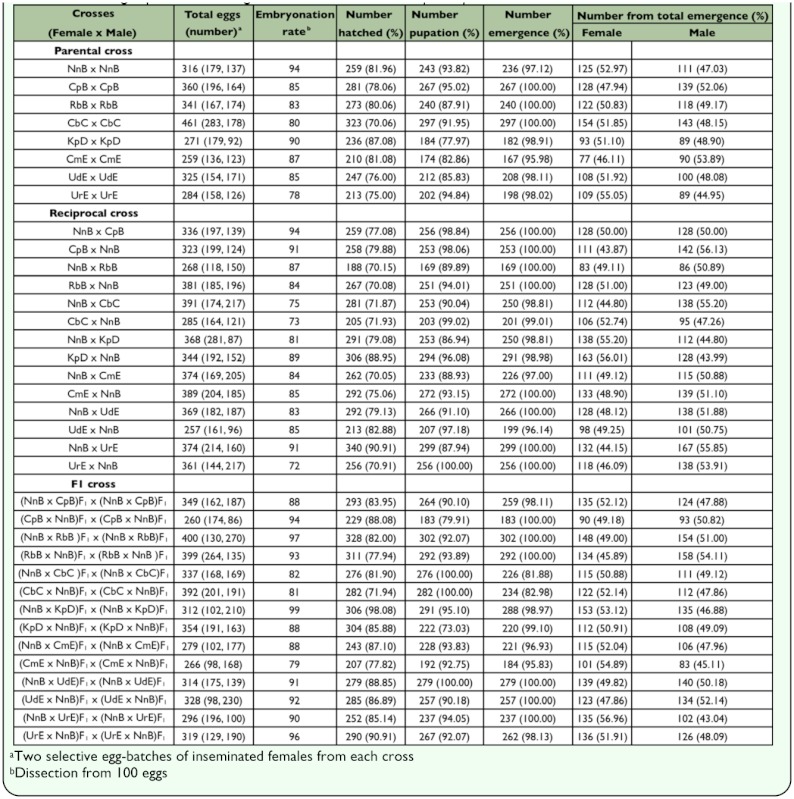
Figure 3.

Complete synapsis in all arms of salivary gland polytene chromosome of F1-hybrid 4th stage larvae of Anopheles peditaeniatus. (A) NnB female × CpB male; (B) NnB female × RbB male, note: small gap of homosequential asynapsis was found on chromosome 3R; (C) NnB female × CbC male; (D) NnB female × KpD male, note: small gap of homosequential asynapsis was found on chromosome 3L; (E) NnB female × CmE male; (F) NnB female × UdE male; (G) NnB female × UrE male. High quality figures are available online.
In the DNA sequence analysis, DNA sequences were determined and analyzed for the ITS2, COI, and COII regions from 8 isoline colonies representative of 4 karyotypic forms of An. Peditaeniatus. In these, all sequences of the ITS2 region were found to be completely identical with a length of 463 bp, but in comparison with An. lesteri they had a very high interspecific sequence variation of 35.4%. The results of comparative sequences of COI and COII regions revealed 548 bp for COI with 0.0 – 1.1% intraspecific sequence variations, and 672 bp for COII with 0.0 – 0.8% intraspecific sequence variations, and seven variable sites were observed from both (Figure 4). Interspecific sequence variations between An. peditaeniatus and An. lesteri in COI and COII were 3.6 – 4.0% and 3.1 – 3.5%, respectively.
Figure 4.

Variable sites in the sequence alignment of the COI and COII sequences. Bases are numbered relative to the alignment. Only those positions differing from the consensus are shown. A dot indicates a base pair identical to that of the NnB sequence. Sequence names are defined in Table 1. High quality figures are available online.
Discussion
Karyotypic variation, due to the addition of an extra block of herterochromatin on sex chromosome (X, Y), is an important mechanism in the speciation process of anopheline mosquitoes and/or other dipteran insects. It could be used as a primary marker for further investigations of sibling species or subspecies status in natural populations of mosquitoes, particularly in those that have heteromorphic sex chromosomes as anophelines (Baimai 1998; Subbarao 1998). Nonetheless, limitation in use should be kept in mind since either markedly different or identical metaphase karyotypes could be cytological characteristics of sibling species or subspecies (cytological races). For example, An. minimus Theobald (minimus species A) has uniquely submetacentric X1, medium submetacentric X2, and submetacentric Y1 chromosomes. Anopheles harrisoni Harbach and Manguin (minimus species C) has unique large submetacentric X3 and large submetacentric Y2 chromosomes (Baimai et al. 1996). Anopheles barbirostris Van der Wulp species A1, A2, A3, and A4 share common characteristics of medium submetacentric X2 and subtelocentric Y1 chromosomes, whereas submetacentric X1, large submetacentric X3, submetacentric Y2, and large submetacentric Y3 chromosomes were common phenomena of the karyotypic variation of An. barbirostris species A1 (Suwannamit et al. 2009).
Crossing experiments for determining hybrid non-viability, sterility, or breakdown are still a useful tool used in the recognition of anopheline species complexes. Detailed genetic incompatibility, including lack of insemination, embryonation, hatchability, larval survival, pupation, emergence, adult sex distortion, abnormal morphology, and reproductive system are useful criteria for elucidating sibling species or subspecies status (Baimai et al. 1987, 1988; Sawadipanich et al. 1990; Subbarao 1998). However, a point worth noting is that an isoline colony established from the combinative characters of morphological and/or cytological markers has to be seriously considered. A laboratoryraised colony established from a naturally mixed population should be omitted, since it may be a mixture of cryptic species or sibling species. Several intra-taxa of the anopheline species that were primarily detected with cytological differences and/or variations that led to doubt of the status of sibling species or subspecies were subsequently confirmed by crossing experiments. These crossing experiments were for sibling species, e.g. An. dirus Peyton and Harrison complex (Baimai et al. 1987, 1988; Sawadipanich et al. 1990), An. maculatus Theobald complex (Chabpunnarat 1988, Thongwat et al. 2008), and An. minimus complex (Choochote et al. 2002b; Somboon et al. 2005); and subspecies (cytological races), e.g. An. sinensis Wiedemann Forms A and B (Park et al. 2008), An. vagus Doenitz Forms A and B (Choochote et al. 2002a), An. pullus Yamada Forms A and B (Park et al. 2003), An. aconitus Doenitz Forms B and C (Junkum et al. 2005), and An. campestris-like Form B, E, and F (Thongsahuan et al. 2009).
In this study, 4 tentative karyotypic forms of An. peditaeniatus, i.e. Form B (X2, X3, Y2), C (X3, Y3), D (X3, Y4), and E (X2, X3, Y5) were obtained from natural populations in Thailand. It is interesting to note that the ancestral Form A (X2, Y1), reported by Baimai et al. (1993), was not detected in any isoline colonies, as only a few samples appeared to be used in the current study. Even though Form A (X2, Y1) was not detected in the present investigation markedly distinct characteristics, particularly the Y chromosomes among the 4 karyotypic forms, were enough to perform their genetic proximity thoroughly. Accordingly, the crossing experiments were carried out among the 4 karyotypic forms in order to determine the degree of genetic proximity. In addition, their comparative DNA sequences of ITS2, COI, and COII were included in this study. The results of no post-mating reproductive isolation among the 4 karyotypic forms, by yielding viable progenies and synaptic salivary gland polytene chromosomes through F2-generations, suggested their conspecific nature. The very low intraspecific sequence variations (0.0 – 1.1%) of the nucleotide sequences of ribosomal DNA (ITS2) and mitochondrial DNA (COI and COII) of the 4 karyotypic forms were strong supportive evidence. Additionally, the length (463 bp) and sequences of ITS2 regions of An. peditaeniatus forms obtained in this study were identical to that of a previous report (Ma and Xu 2005). Similar results have been reported in An. sinensis Forms A and B (Park et al. 2008), An. vagus Forms A and B (Choochote et al. 2002a), An. pullus Forms A and B (Park et al. 2003), An. aconitus Forms B and C (Junkum et al. 2005), and An. campestris-like Forms B, E, and F (Thongsahuan et al. 2009).
Acknowledgements
I thank the Thailand Research Fund (TRF: BRG5380021), and Research and Diagnostic Center for Emerging Infectious Diseases, Khon Kaen Univerysity, Thailand for financially supporting this research project; Assoc. Prof. Dr. Niwes Nantachit, Dean of the Faculty of Medicine, Chiang Mai University, for his interest in this research; and the Faculty of Medicine Endowment Fund for Research Publication for its financial support in defraying publication costs.
References
- Baimai V. Heterochromatin accumulation and karyotypic evolution in some Dipteran insects. Zoological Studies. 1998;32:75–88. [Google Scholar]
- Baimai V, Andre RG, Harrison BA, Kijchalao U, Panthusiri L. Crossing and chromosomal evidence for two additional sibling species within the taxon Anopheles dirus Peyton and Harrison (Diptera: Culicidae) in Thailand. Proceedings of the Entomological Society of Washington. 1987;89:157–166. [Google Scholar]
- Baimai V, Harbach RE, Kijchalao U. Cytogenetic evidence for a fifth species within the taxon Anopheles dirus in Thailand. Journal of the American Mosquito Control Association. 1988;4:333–338. [PubMed] [Google Scholar]
- Baimai V, Kijchalao U, Rattanarithikul R. Metaphase karyotypes of Anopheles of Thailand and Southeast Asia: V. Myzomyia series, Subgenus Cellia (Diptera: Culicidae). Journal of the American Mosquito Control Association. 1996;12:97–105. [PubMed] [Google Scholar]
- Baimai V, Rattanarithikul R, Kijchalao U. Metaphase karyotypes of Anopheles of Thailand and Southeast Asia: I. The hyrcanus group. Journal of the American Mosquito Control Association. 1993;9:59–67. [PubMed] [Google Scholar]
- Chabpunnarat S. Cytogenetic study of the Anopheles maculatus complex. M.Sc. Thesis, Mahidol University Press; 1988. [Google Scholar]
- Chai JY. Re-emerging Plasmodium vivax malaria in the Republic of Korea. The Korean Journal of Parasitology. 1999;37:129–143. doi: 10.3347/kjp.1999.37.3.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choochote W, Jitpakdi A, Sukontason KL, Chaithong U, Wongkamchai S, Pitasawat B, Jariyapan N, Suntaravitun T, Rattanachanpichai E, Sukontason K, Leemingsawat S, Rongsriyam Y. Intraspecific hybridization of two karyotypic forms of Anopheles vagus (Diptera: Culicidae) and the related egg surface topography. Southeast Asian Journal of Tropical Medicine and Public Health. 2002a;33(3):29–35. [PubMed] [Google Scholar]
- Choochote W, Pitasawat B, Jitpakdi A, Rattanachanpichai E, Riyong D, Leemingsawat S, Wongkamchai S. The application of ethanol-extracted Gloriosa superba for metaphase chromosome preparation in mosquitoes. Southeast Asian Journal of Tropical Medicine and Public Health. 2001;32:76–82. [PubMed] [Google Scholar]
- Choochote W, Rongsriyam Y, Leemingsawat S, Jitpakdi A, Komalamisra N, Surathin K, Somboon P, Chen B, Wongkamchai S, Jariyapan N, Tippawangkosol P, Pitasawat B, Riyong D. Intraspecific hybridization of Anopheles minimus (Diptera: Culicidae) species A and C in Thailand. Southeast Asian Journal of Tropical Medicine and Public Health. 2002b;33(3):23–28. [PubMed] [Google Scholar]
- Harrison BA, Scanlon JE. Medical entomology studies II. The subgenus Anopheles in Thailand (Diptera: Culicidae). Contributions of the American Entomological Institute. 1975;12:78. [Google Scholar]
- Junkum A, Komalamisra N, Jitpakdi A, Jariyapan N, Min GS, Park MH, Cho KH, Somboon P, Bates PA, Choochote W. Evidence to support two conspecific cytological races on Anopheles aconitus in Thailand. Journal of Vector Ecology. 2005;30:213–224. [PubMed] [Google Scholar]
- Kanda T. Improved techniques for the preparation of polytene chromosome for some anopheline mosquitoes. Mosquito News. 1979;39:568–574. [Google Scholar]
- Kanojia PC, Shetty PS, Geevarghese G. A long-term study on vector abundance & seasonal prevalence in relation to the occurrence of Japanese encephalitis in Gorakhpur district, Uttar Pradesh. Indian Journal of Medical Research. 2003;117:104–110. [PubMed] [Google Scholar]
- Kim SJ, Choochote W, Jitpakdi A, Junkum A, Park SJ, Min GS. Establishment of a self-mating mosquito colony of Anopheles sinensis from Korea. The Korean Journal of Parasitology. 2003;33:267–271. [Google Scholar]
- Ma YJ, Xu JN. The Hyrcanus group of Anopheles (Anopheles) in China (Diptera: Culicidae): Species discrimination and phylogenetic relationships inferred by ribosomal DNA internal transcribed spacer 2 sequences. Journal of Medical Entomology. 2005;42:610–619. [PubMed] [Google Scholar]
- Mourya DT, Ilkal MA, Mishra AC, Jacob PG, Pant U, Ramanaujam S, Mavale MS, Bhat HR, Dhanda V. Isolation of Japanese encephalitis virus from mosquitoes collected in Karnataka State, India from 1985 to 1987. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1989;83:550–552. doi: 10.1016/0035-9203(89)90288-5. [DOI] [PubMed] [Google Scholar]
- Park SJ, Choochote W, Jitpakdi A, Junkum A, Kim SJ, Jariyapan N. Evidence for a conspecific relationship between two morphologically and cytologically different forms of Korean Anopheles pullus mosquito. Molecules and Cells. 2003;16:354–360. [PubMed] [Google Scholar]
- Park MH, Choochote W, Kim SJ, Somboon P, Saeung A, Tuetan B, Tsuda Y, Takagi M, Joshi D, Ma YJ, Min GS. Nonreproductive isolation among four allopatric strains of Anopheles sinensis in Asia. Journal of the American Mosquito Control Association. 2008;24:489–495. doi: 10.2987/08-5753.1. [DOI] [PubMed] [Google Scholar]
- Rattanarithikul R, Harrison BA, Harbach RE, Panthusiri P, Coleman RE. Illustrated keys to the mosquitoes of Thailand IV. Anopheles. Southeast Asian Journal of Tropical Medicine and Public Health. 2006;37(2):1–128. [PubMed] [Google Scholar]
- Rattanarithikul R, Konishi E, Linthicum KJ. Detection of Plasmodium vivax and Plasmodium falciparum circumsporozoites antigen in anopheline mosquitoes collected in southern Thailand. American Journal of Tropical Medicine and Hygiene. 1996;54:114–121. doi: 10.4269/ajtmh.1996.54.114. [DOI] [PubMed] [Google Scholar]
- Ree HI, Hwang UW, Lee IY, Kim TE. Daily survival and human blood index of Anopheles sinensis, the vector species of malaria in Korea. Journal of the American Mosquito Control Association. 2001;17:67–72. [PubMed] [Google Scholar]
- Reid JA. Anopheline mosquitoes of Malaya and Borneo. Studies from the Institute for Medical Research Malaysia. 1968;31:1–520. [Google Scholar]
- Sawadipanich Y, Baimai V, Harrison BA. Anopheles dirus species E: chromosomal and crossing evidence for another member of the dirus complex. Journal of the American Mosquito Control Association. 1990;6:477–481. [PubMed] [Google Scholar]
- Somboon P, Thongwat D, Choochote W, Walton C, Takagi M. Crossing experiments of Anopheles minimus species C and putative species E. Journal of the American Mosquito Control Association. 2005;21:5–9. doi: 10.2987/8756-971X(2005)21[5:CEOAMS]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Subbarao SK. Anopheline species complexes in South-East Asia. World Health Organization Technical Publication Searo. 1998;18:1–82. [Google Scholar]
- Suwannamit S, Baimai V, Otsuka Y, Saeung A, Thongsahuan S, Tuetun B, Apiwathnasorn C, Jariyapan N, Somboon P, Takaoka H, Choochote W. Cytogenetic and molecular evidence for an additional new species within the taxon Anopheles barbirostris (Diptera: Culicidae) in Thailand. Parasitology Research. 2009;104:905–918. doi: 10.1007/s00436-008-1272-1. [DOI] [PubMed] [Google Scholar]
- Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997;25:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thongsahuan S, Baimai V, Otsuka Y, Saeung A, Tuetun B, Jariyapan N, Suwannamit S, Somboon P, Jitpakdi A, Takaoka H, Choochote W. Karyotypic variation and geographic distribution of Anopheles campestris-like (Diptera: Culicidae) in Thailand. Memorias do Instituto Oswaldo Cruz. 2009;104:558–566. doi: 10.1590/s0074-02762009000400004. [DOI] [PubMed] [Google Scholar]
- Thongwat D, Morgan K, O'loughlin MS, Walton C, Choochote W, Somboon P. Crossing experiment supporting the specific status of Anopheles maculatus chromosomal form K. Journal of the American Mosquito Control Association. 2008;24:194–202. doi: 10.2987/5668.1. [DOI] [PubMed] [Google Scholar]
- Yang MN, Ma YJ. Molecular population genetic structure of Anopheles lesteri (Diptera: Culicidae) based on mtDNACOI gene sequences. Acta Entomologica Sinica. 2009;52:1000–1007. [Google Scholar]
- Zhang HL. The natural infection rate of mosquitoes by Japanese encephalitis B virus in Yunnan Province. Zhonghua yufangyixue zazhi. 1990;24:265–267. [PubMed] [Google Scholar]