

Abstract
A recombinant modified vaccinia Ankara (MVA) virus expressing mature viral protein 2 (VP2) of the infectious bursal disease virus (IBDV) was constructed to develop MVA-based vaccines for poultry. We demonstrated that this recombinant virus was able to induce a specific immune response by observing the production of anti-IBDV-seroneutralizing antibodies in specific pathogen-free chickens. Besides, as the epitopes of VP2 responsible to induce IBDV-neutralizing antibodies are discontinuous, our results suggest that VP2 protein expressed from MVA-VP2 maintained the correct conformational structure. To our knowledge, this is the first report on the usefulness of MVA-based vectors for developing recombinant vaccines for poultry.
Keywords: chicken immunity, IBDV, MVA, VP2 protein
Infectious bursal disease virus (IBDV) is the causative agent of infectious bursal disease (IBD), a highly contagious and severe disease in young chickens responsible for economic losses in the poultry industry worldwide. IBDV destroys B lymphocyte precursors found within the bursa of Fabricious, thereby inducing atrophy, immunosuppression, and/or mortality in unprotected flocks [1]. Viral mature protein 2 (VP2), main component of the icosahedral IBDV capsid, is known as the only host-protective antigen. The function of VP2 is highly conformation-dependent with at least three independent epitopes responsible for inducing the production of neutralizing antibodies [11]. The modified vaccinia Ankara (MVA) virus is highly attenuated and used as an efficient non-replicative vaccine vector. Its inability to replicate in most mammalian cells provides excellent host protection. Antigens encoded by foreign genes inserted into the recombinant vectors are expressed and presented to the immune system, thus triggering humoral and cellular immune responses in the host [8]. In the present report, we describe the construction of a recombinant MVA virus expressing the VP2 protein of IBDV and the evaluation of its capability to induce immunity in specific pathogen-free (SPF) chickens.
Firstly, we constructed a transfer vector (denominated VT-MTK-GUS-VP2) containing foreign genes flanked by sequences of the MVA086R gene (NCBI accession No. U94848). This plasmid will recombine with the MVA genome through homolog sequences to produce recombinant MVA viruses. The selected genes included the nucleotide sequence encoding the VP2 mature protein of the Argentinian IBDV isolate LD-847-04 (vp2 gene) and the uid A gene (encoding the β-glucuronidase enzyme [GUS]) under the regulation of poxviral synthetic pE/L and pH6 promoters, respectively. For this purpose, the VTM-TK-GUS plasmid described previuosly [3] was used. The vp2 gene (from nucleotide 1 to 1,323 of segment A, NCBI accession No. JF965438) was amplified by RT-PCR from total RNA of infected chicken embryos using specific primers (VP2 Forward 5'GCTAGCCGCCGCCATGAC AAACCTGCAAGATC and VP2 Reverse 5'AGATCTGC TCCTGCAATCTTCAGG). Next, the vp2 gene was cloned into the VT-MTK-GUS plasmid to obtain the transfer vector VT-MTK-GUS-VP2. The correct nucleotide sequence and orientation of the vp2 gene in the transfer vector were verified by DNA sequencing using an ABI PRISM 3130 genetic analyzer (Applied Biosystems, USA).
Recombinant MVA viruses expressing the VP2 protein were obtained by transfecting the VT-MTK-GUS-VP2 construct into primary chicken embryo fibroblasts (CEFs) previously infected with MVA at a multiplicity of infection (moi) of 0.05 plaque-forming units (PFU) per cell. The embryonated eggs for the production of CEFs monolayers were purchased at the Instituto Rosenbusch (Argentina). The expression of GUS allowed plaque lysis purification of recombinant viruses by screening in the presence of β-glucoronidase substrate (X-Gluc, 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid; Inalco, Italy). The purity of MVA-VP2 viruses and presence of the vp2 gene were confirmed by PCR after five rounds of screening (data not shown).
In order to evaluate the expression of VP2 protein after infection with the recombinant viruses, Western blotting (WB) and immunofluorescence assays (IFA) were performed using polyclonal and monoclonal anti-VP2 antibodies, respectively, provided by Dr. J. F. Rodríguez (Centro Nacional de Biotecnología, Spain). Briefly, DF1 cells (CRL-12203; ATCC, USA) were infected with MVA or MVA-VP2 at a moi of 0.6 PFU per cell. At 24 h post-infection, the cells were collected by centrifugation and resuspended in Laemmli's sample buffer (62.5 mM Tris-HCl, pH 6.8; 2% sodium dodecyl sulfate, SDS; 0.25% bromophenol blue; 5% glycerol and 50 mM dithiothreitol) or fixed and permeabilized with methanol at -20℃. In the WB, a specific band with an apparent molecular weight of 37 kDa corresponding to mature VP2 protein was only detected in protein extracted from MVA-VP2-infected cells (data not shown). In addition, expression of the VP2 protein was detected by IFA in the cytoplasm of cells infected with the recombinant virus. For this, the fixed cells were blocked with 5% fetal calf serum in phosphate buffered saline (PBS) (14 mM NaCl; 3 mM KCl; 8 mM Na2HPO4; 1.5 mM KH2PO4; pH 7.2), and then incubated with a monoclonal anti-VP2 (mouse 17G2; INGENASA, Spain) followed by incubation with secondary antibody Alexa 488 goat anti-mouse (Invitrogen, USA). The nuclei were stained with 4', 6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich, USA). Fluorescent signals were detected by a Zeiss Axiovert 200 confocal laser scanning microscope (Germany). Images were captured using the Laser Sharp software package (Bio-Rad, USA). As shown in the Fig. 1, the expression of VP2 protein was only evidenced in the cytoplasm of MVA-VP2 infected cells indicating the capability of these recombinant viruses to express the heterologous protein. Genetic stability of the MVA-VP2 virus was confirmed by PCR and WB assays after 10 blind passages in CEFs at a low moi (0.05~0.1).
Fig. 1.
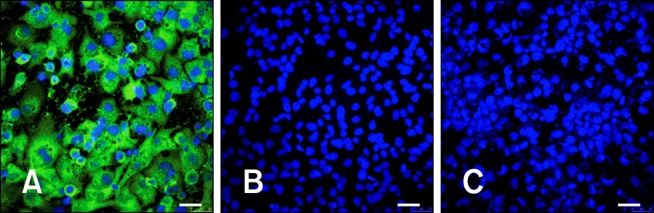
Expression of mature viral mature protein 2 (VP2) from modified vaccinia Ankara-VP2 recombinant virus. (A) The cytoplasmic expression of the VP2 protein was evaluated by an immunofluorescence assay in DF1 cells infected with MVA-VP2. As a control, uninfected (B) or MVA-infected DF1 cells (C) were included. Scale bars = 25 µm.
Once expression of the VP2 protein following infection with MVA-VP2 recombinant virus was confirmed, immunogenicity was assessed in SPF White Leghorn chickens purchased from Instituto Rosenbusch and housed in animal facilities at the Biotechnology Institute, Instituto Nacional de Tecnología Agropecuaria (Argentina). All experiments were done in compliance with international and institutional guidelines for the care and use of laboratory animals. Groups of five birds (11 days old) were intramuscularly (i.m.) immunized with homogenates of CEFs infected with MVA or MVA-VP2 (2~4 × 107 PFU/bird). As a control, groups of five animals were i.m. inoculated with a homogenate of non-infected CEFs (CEF-NI) or vaccinated orally with D78 live vaccine (2.7 × 103 PFU/bird) or 50 µL of PBS. All chickens were boosted when they were 25 and 39 days old with the same inocula. Blood samples were collected from the wing vein when the birds were 11 days of age (pre-immune) and 2 weeks after each immunization (25, 39, and 53 days old). Antibodies specific for IBDV were analyzed by an in vitro virus neutralization test using a constant virus-varying serum procedure previously described by Jackwood et al. [5]. IBDV-specific antibodies were not detected in the sera of birds vaccinated with CEF-NI, MVA, or PBS. On the other hand, IBDV-seroneutralizing antibodies were detected in serum samples from chickens immunized with MVA-VP2 or the D78 vaccine. In the group of birds vaccinated with MVA-VP2, the seroneutralizing antibody titers increased after each booster. The commercial IBDV-D78 live vaccine resulted in higher levels of neutralizing antibodies than the recombinant MVA-VP2 immunogen (Table 1).
Table 1.
IBDV-neutralizing antibody titers in the sera of vaccinated birds

Seroneutralizing titers are expressed as the reciprocal of the last serum dilution that completely neutralized the virus. MVA-VP2: recombinant MVA virus expressing IBDV VP2 mature protein, IBDV-D78: live commercial IBDV vaccine (strain D78).
Currently, IBD is mainly controlled by administration of attenuated IBDV vaccines in the poultry industry. These vaccines often fail to protect chickens with maternal antibodies and many cause moderate bursal atrophy, which may facilitate both infections by opportunistic pathogens and poor immune responses to other vaccines. In order to overcome the problems caused by conventional vaccines against IBDV, a variety of different systems have been used to express VP2 protein as a subunit immunogen or by in vivo expression from live viral vectors [4,7,10]. In all cases, IBDV-neutralizing antibody production was induced in the treated chickens. However, different levels of protection against viral challenge were reported depending on certain factors such as the vector used, dose and route of administration, and chicken genotype [10,11].
The usefulness of MVA-based vectors to develop novel vaccines for use in mammals has been previously demonstrated [2,9] albeit these have not been evaluated in chickens. Consequently, we constructed a recombinant MVA virus expressing the VP2 protein and evaluated its immunogenicity in SPF chickens. Expression of the heterologous VP2 protein was demonstrated in MVA-VP2-infected cells by IFA and WB assays. Detection of the VP2 protein in the immunoassays does not determine the in vivo immunogenicity of the protein because the neutralizing VP2 epitopes are discontinuous. For this reason, the presence of IBDV-neutralizing antibodies in the serum of birds vaccinated with MVA-VP2 indicated that the VP2 protein expressed by this recombinant virus had the correct conformational structure. We also demonstrated that responses elicited by immunization with the viral vector would not interfere with subsequent immunizations since the IBDV-seroneutralizing titers increased four-fold after each booster. Although neutralizing antibody titers achieved with MVA-VP2 were lower than those obtained with a replicative D78 live vaccine, we did not evaluate the potential cellular immunity induced by a recombinant poxvirus that may also participate in immune responses against IBDV infections [6]. Further experiments will be performed to confirm the induction of cellular immune response and to assess the protection afforded by vaccination with MVA-VP2 in chickens (with and without anti-IBDV maternal antibodies). In summary, our findings demonstrated that MVA-based vectors may be useful to develop effective recombinant vaccines against poultry diseases.
Acknowledgments
The authors are grateful to Mrs. M. J. Mónaco, Mr. S. Díaz, and Dr. O. Zabal for technical assistance. The authors also thank Dr. J. F. Rodríguez (CNB, Spain) for assisting with the IFA. This work was supported by grants (AERG 232141 and PICT 2008-0400) from the Instituto Nacional de Tecnología Agropecuaria (INTA) and Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), respectively. FZ was supported by an external fellowship for junior researchers from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina.
References
- 1.Becht H. Infectious bursal disease virus. Curr Top Microbiol Immunol. 1980;90:107–121. doi: 10.1007/978-3-642-67717-5_5. [DOI] [PubMed] [Google Scholar]
- 2.Breathnach CC, Clark HJ, Clark RC, Olsen CW, Townsend HG, Lunn DP. Immunization with recombinant modified vaccinia Ankara (rMVA) constructs encoding the HA or NP gene protects ponies from equine influenza virus challenge. Vaccine. 2006;24:1180–1190. doi: 10.1016/j.vaccine.2005.08.091. [DOI] [PubMed] [Google Scholar]
- 3.Ferrer MF, Zanetti FA, Calamante G. Design and construction of transfer vectors in order to obtain recombinant modified vaccinia virus Ankara (MVA) Rev Argent Microbiol. 2007;39:138–142. [PubMed] [Google Scholar]
- 4.Francois A, Chevalier C, Delmas B, Eterradossi N, Toquin D, Rivallan G, Langlois P. Avian adenovirus CELO recombinants expressing VP2 of infectious bursal disease virus induce protection against bursal disease in chickens. Vaccine. 2004;22:2351–2360. doi: 10.1016/j.vaccine.2003.10.039. [DOI] [PubMed] [Google Scholar]
- 5.Jackwood DJ, Saif YM, Moorhead PD, Dearth RN. Infectious bursal disease virus and Alcaligenes faecalis infections in turkeys. Avian Dis. 1982;26:365–374. [PubMed] [Google Scholar]
- 6.Kim IJ, You SK, Kim H, Yeh HY, Sharma JM. Characteristics of bursal T lymphocytes induced by infectious bursal disease virus. J Virol. 2000;74:8884–8892. doi: 10.1128/jvi.74.19.8884-8892.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lai SY, Lee MS, Chen HC, Shen PC, Jinn TR, Kao SS, Wang MY. Production and purification of immunogenic virus-like particles formed by the chimeric infectious bursal disease virus structural protein, rVP2H, in insect larvae. Process Biochem. 2004;39:571–577. [Google Scholar]
- 8.Moss B, Carroll MW, Wyatt LS, Bennink JR, Hirsch VM, Goldstein S, Elkins WR, Fuerst TR, Lifson JD, Piatak M, Restifo NP, Overwijk W, Chamberlain R, Rosenberg SA, Sutter G. Host range restricted, non-replicating vaccinia virus vectors as vaccine candidates. Adv Exp Med Biol. 1996;397:7–13. doi: 10.1007/978-1-4899-1382-1_2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Porter DW, Thompson FM, Berthoud TK, Hutchings CL, Andrews L, Biswas S, Poulton I, Prieur E, Correa S, Rowland R, Lang T, Williams J, Gilbert SC, Sinden RE, Todryk S, Hill AV. A human Phase I/IIa malaria challenge trial of a polyprotein malaria vaccine. Vaccine. 2011;29:7514–7522. doi: 10.1016/j.vaccine.2011.03.083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shaw I, Davison TF. Protection from IBDV-induced bursal damage by a recombinant fowlpox vaccine, fpIBD1, is dependent on the titre of challenge virus and chicken genotype. Vaccine. 2000;18:3230–3241. doi: 10.1016/s0264-410x(00)00133-x. [DOI] [PubMed] [Google Scholar]
- 11.Vakharia VN, He J, Ahamed B, Snyder DB. Molecular basis of antigenic variation in infectious bursal disease virus. Virus Res. 1994;31:265–273. doi: 10.1016/0168-1702(94)90009-4. [DOI] [PubMed] [Google Scholar]