

Abstract
Vasopressin is a neuropeptide that has been strongly implicated in the development and evolution of complex social relations and cognition in mammals. Recent studies in voles have shown that polymorphic variation in the promoter region of the arginine vasopressin V1a receptor gene (avpr1a) is associated with different dimensions of sociality. In humans, variation in a repetitive sequence element in the 5’ flanking region of the AVPR1A, known as RS3, have also been associated with variation in AVPR1a gene expression, brain activity and social behavior. Here, we examined the association of polymorphic variation in this same 5’ flanking region of the AVPR1A on subjective ratings of personality in a sample of 83 chimpanzees (Pan troglodytes). Initial analyses indicated that 34 females and 19 males were homozygous for the short allele, which lacks RS3(DupB−/−); while 18 females and 12 males were heterozygous and thus had one copy of the long allele containing RS3 (DupB+/−), yielding overall allelic frequencies of 0.82 for the DupB− allele and 0.18 for the DupB+ allele. DupB+/+ chimpanzees were excluded from the analysis due to the limited number of individuals. Results indicated no significant sex difference in personality between chimpanzees homozygous for the deletion of the RS3-containing DupB region (DupB−/−); however, among chimpanzees carrying one allele with the DupB present (DupB+/−), males had significantly higher dominance and lower conscientiousness scores than females. These findings are the first evidence showing that the AVPR1A gene plays a role in different aspects of personality in male and female chimpanzees.
Vasopressin is a neuropeptide with multiple physiological functions that has been strongly implicated in the development and evolution of complex social relations and cognition in mammals (Donaldson & Young, 2008, Goodson & Bass, 2001). Studies in several species have shown that one of three known AVP receptors, arginine vasopressin V1a receptor (AVPR1A), is expressed in the brain and plays a prominent role in producing diversity in social behavior, in addition to its role in regulation of vasoconstriction. For example, meadow and prairie voles, which differ dramatically in their pair bonding behavior, show pronounced differences in AVPR1A expression patterns in the brain (Lim et al., 2005, Lim et al., 2004). More recently, several studies in voles have examined variation in behavior and AVPR1A expression in relation to microsatellite length in the promoter region of the V1a receptor gene (avpr1a). There are both individual and species differences in avpr1a expression and these have been associated with variation in pair bonding and other dimensions of social behavior in voles (Hammock et al., 2005, Hammock & Young, 2005, Hammock & Young, 2006, Young & Wang, 2004). Variation in this polymorphic microsatellite have also been shown to affect avpr1a gene transcription in cell transcription from in vitro reporter assays (Hammock & Young, 2004).
In primates, less is known about the functional role of AVPR1A in social behavior but recent studies have shown considerable polymorphic variation in the AVPR1A gene analogous to that seen in voles, suggesting that individual and species differences in sociality may be associated with this gene (Babb et al., 2010, Donaldson et al., 2008, Rosso et al., 2008). In human AVPR1A, variation in repetitive microsatellite element RS3 (see Figure 1) located in the 5’ flanking region of the gene has been linked to variation in social behavior, including altruistic behavior, and pair bonding related behavior in males (Donaldson et al., 2008, Meyer-Lindenberg et al., 2011, Walum et al., 2008). It has also been suggested that AVPR1A may be a candidate susceptibility gene for autism, a spectrum of neurodevelopmental disorders marked by problems in the development of normal social relationships and socio-cognitive abilities (Melke, 2008). Likewise, AVPR1A variation in RS3 is related to activation of the amygdala during a face recognition task (Meyer-Lindenberg et al., 2008). Finally, variation in the length of RS3 has been associated with variation in AVPR1A gene expression in the human brain (Knafo et al., 2008) and in transcription reporter assays in cell culture (Tansey et al., 2011). Thus, our understanding of variation in sociality and cognitive processes as they relate to social cues and processes might benefit from further studies on the AVPR1A gene.
Figure 1.
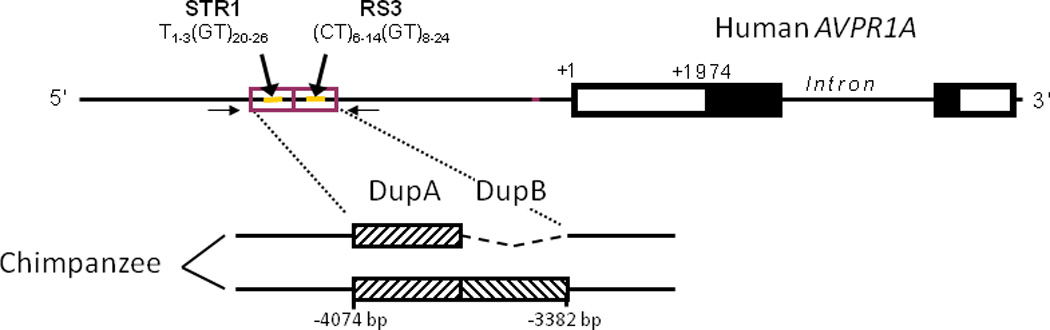
Schemative of the polymorphic microsatellite elements in the primate AVPR1A locus. Two repeat motifs (DupA and DupB) in tandem containing repetitive microsatellite elements (STR1 and RS3) are found in the 5’ flanking region of the human AVPR1A. Variation in the length of RS3 is associated with social behavior in humans, including social cognition in autism spectrum disorder (ASD). Chimpanzees are polymorphic at this site, with a significant proportion having the entire DupB, including RS3 deleted (short). Adapted from (Donaldson et al., 2008).
In this study, we examined the association of an indel containing RS3 upstream of the AVPR1A gene with individual differences in personality in captive chimpanzees (Pan troglodytes). Chimpanzees are an excellent model to explore the functional role of the RS3 polymorphism in the AVPR1A gene because there is a common indel resulting in a complete deletion of the RS3 sequence in 80% of the chimpanzee alleles (Donaldson et al., 2008). Within humans, the RS3 repeat region is housed within a larger, ~350 bp tandem duplicated region. The first of these duplicated regions, DupA, spans -3730 to -4074 bp relative to the transcription start site and contains a GT20-26 microsatellite, known as STR1. The second block, DupB, spans -3382 to -3729 bp and contains the complex microsatellite, RS3 ((CT)6–14(GT)8–24) (Figure 1). Chimpanzees are polymorphic for the presence of the RS3-containing DupB region, leading to a 357 bp difference between the DupB+ and Dup- alleles (Donaldson et al., 2008). The deletion of RS3 in some individuals makes this species ideal for assessing the potential role of the AVPR1a gene, and more specifically RS3, on sociality and related processes (i.e., social cognition).
To assess the association between the RS3 DupB indel and personality in the chimpanzees, we compared a sample of individuals who were either homozygous for the deletion (DupB−/−) or were heterozygous and had one DupB containing allele (DupB+/−) on subjective personality ratings. Previous studies have shown that chimpanzees, like humans, have different personalities, which include what has been described as the five-factor model (Digman, 1990) or “Big 5” personality dimensions in humans (extraversion, agreeableness, conscientiousness, neuroticism, & openness). In addition, chimpanzees have a sixth personality dimension, dominance, which is seemingly related to competitive prowess (King & Figueredo, 1997). Despite the recent comparative interest in personality among nonhuman animals (Freeman & Gosling, 2010, Gosling, 2001), to date the majority of studies have focused on the description and construction of personality factors in nonhuman animals (Gosling, 2001) whereas very few studies have examined the potential role of genetic factors on individual differences in personality. In the only study in chimpanzees, Hong et al. (2011) found that polymorphic variation in a gene that encodes tryptophan hydroxylase 2 (TPH2), an enzyme involved in the production of serotonin, was linked to variation in neuroticism.
In the current study, we sought to examine whether variation in personality was associated with the AVPR1A 5’ flanking region polymorphism, given its reported important role in sociality. This was accomplished by combining recently published data on personality in a sample of chimpanzees housed at the Yerkes National Primate Center (Weiss et al., 2007) with the vasopressin genotype data that was reported in a subsample of these same individuals (Donaldson et al., 2008). Thus, we combined the genetic data with the extant personality ratings to assess whether differences in personality were related to the presence or absence of the RS3-containing DupB element in the AVPR1A 5’ flanking region. If the AVPR1A 5’ flanking region plays a role in personality in chimpanzees, then significant differences should be evident in one or more of the traits in chimpanzees with different genotypes.
Methods
Subjects
All chimpanzees were members of the colony of apes housed at the Yerkes National Primate Research Center (YNPRC). DNA samples and associated personality rating scores were available in 83 adult and sub-adult chimpanzees including 52 females and 31 males.
Personality Assessment
We used the personality measures reported by Weiss, King and Hopkins (2007), which included chimpanzees housed at the YNPRC. Briefly, in theWeiss et al. (2007) paper, personality was assessed using the chimpanzee personality questionnaire (CPQ) which consists of a 43 adjective questionnaire items used by observers to rate their overall impressions of the chimpanzee’s behaviors (a freely available copy can be found at http://extras.springer.com/2011/978-1-4614-0175-9). For example, the adjective “cautious” is defined as subject that often seems attentive to harm or danger from its actions and avoids risky behaviors. Individual adjectives, reflecting traits in the behavior of the animals, were rated on a scale from 1 to 7, where 1 indicated the absence of that trait and 7 indicated high frequency in the expression of that trait. The CPQ has been shown to be both reliable and valid with interrater reliabilities ranging from 0.71 to 0.88 (King & Figueredo, 1997, King et al., 2005). Weiss et al (2007) described four personality factors in chimpanzees: dominance (DCH), extraversion (ECH), conscientiousness (CCH) and agreeableness (ACH) that were stable and replicable across different samples of chimpanzees including individuals living in zoos in comparison to those housed at the YNPRC.
The item loadings for each trait reported byWeiss et al. (2007) are shown in Table 1. The factor scores were unit weighted and converted into T-scores with a mean of 50 and a standard deviation of 10. Thus, T-scores over 50 indicate that a subject has more than the average dimension of that trait whereas values lower than 50 indicated that subjects had lower than average characteristics of that trait. The individual T-scores as well as the 43 item-specific ratings collected in the Weiss et al. study were used in the analyses in this study.
Table 1.
Item Traits Loading on E ach Personality Factor
| Trait | Dominance | Extraversion | Conscientiousness | Agreeableness |
|---|---|---|---|---|
| Positive Items | ||||
| Dominant | Active | Stable | Gentle | |
| Persistent | Playful | Predictable | Sociable | |
| Bullying | Sociable | Friendly | ||
| Independent | Inquisitive | Affectionate | ||
| Decisive | Friendly | Stable | ||
| Aggressive | Inventive | Predictable | ||
| Defiant | Affectionate | Sensitive | ||
| Stingy | Helpful | |||
| Manipulative | Protective | |||
| Intelligent | ||||
| Sympathetic | ||||
| Negative Items | ||||
| Submissive | Lazy | Aggressive | ||
| Dependent | Solitary | Defiant | ||
| Timid | Depressed | Depressed | ||
| Fearful | Unemotional | Erratic | ||
| Cautious | Impulsive | |||
| Excitable | ||||
| Jealous | ||||
| Disorganized | ||||
| Autistic | ||||
| Irritable | ||||
| Reckless | ||||
| Clumsy |
The data are taken from Weiss et al. (Weiss et al., 2007).
DNA Extraction, Genotyping, and Analysis
DNA samples were isolated from buccal swabs or blood samples using Puregene DNA Purification system (Gentra, Minneapolis, MN) as described by Donaldson et al (2008). Following extraction, stock DNA was separated into three aliquots: one for onsite storage at -80°C, one for offsite storage, and a working stock for genotyping. Samples were tracked via a secure Filemaker Pro 8 database that linked sample codes for each aliquot, demographics for each subject (e.g., subject number, birth date, sire, dam, etc.), DNA quantification and purity analysis results, and genotype data.
Each individual was genotyped for the AVPR1A DupA/B region using the primers and conditions reported in previous studies with slight modifications (Donaldson et al., 2008). Briefly, we used forward primer 5'-GCATGGTAGCCTCTCTTTAAT and a reverse primer of 5'-CATACACATGGAAAGCACCTAA with an annealing temperature of 57°C for 30 cycles: 95°C, 5 min; 30×(95°C, 30 sec; 57°C, 30 sec; 72°C, 3 min; 72°C, 10 min; 4°C, hold). PCR amplification was undertaken using the Epicentre Failsafe kit using premix H (Illumina Inc., Madison, WI) according to the manufacturer’s directions. Genotyping was performed in a volume of 20 microliters containing 20 ng target genomic DNA. PCR products were resolved on a 2% agarose gel (SeaKem Agarose LE, Lonza, Basel, Switzerland) at 100V for 45 min with a 100bp DNA ladder (New England Biolabs, Ipswich, MA) in TBE. The DupB containing allele resulted in a band of ~900 bp, while the DupB minus allele was ~570 bp long, and genotypes were visually assigned (Donaldson et al., 2008). All genotypes were run in duplicate with gel analysis and were checked by the authors before the data set was finalized (N = 130).
Results
Allelic Frequencies
Thirty-four females and 19 males were homozygous for the short allele (DupB−/−); while 18 females and 12 males were heterozygous and thus had the long allele(DupB+/−), yielding overall allelic frequencies of 0.64 for the DupB− allele and 0.36 for the DupB+ allele. These frequencies are consistent with those previously identified in wild-caught chimpanzees (Donaldson et al., 2008). Based on these allelic frequencies, only 3.24% of the population is expected to be homozygous for DupB+ allele. While we did not identify any homozygous DupB+/+ individuals in this particular population our distribution of genotypes is not significantly different from the number of individuals expected to carry these genotypes according to the Hardy Weinberg equation Χ2 (2, N=130)=.03, n.s.
AVPR1A Variation and Personality
We compared the factor scores using ANCOVA with sex (male, female) and DupB genotype (DupB+/−, DupB−/−) as between group factors while the relatedness coefficients served as a covariate. Relatedness coefficients were used to determine the degree of relatedness of each individual to all other individuals in the YNPRC chimpanzee pedigree analysis. A significant three-way interaction was found between DupB genotype, sex and personality factor F(3, 216) = 4.43, p < .005. To further breakdown this interaction, we performed univariate F-tests with each personality factor serving as a dependent measure while sex and genotype served as between group factors.
Significant two-way interactions between sex and DupB genotype were found for dominance F(1, 79)=5.133, p < .03 and conscientiousness F(1, 79) = 4.33, p < .05. The mean T-score for dominance and conscientiousness for male and female DupB+/− and DupB−/−chimpanzees are shown in Figure 2. Post-hoc analysis using Tukey’s HSD indicated that for dominance, no significant difference was found between DupB−/− males and females. However, DupB+/− males had significantly higher dominance T-scores than DupB+/− females. In contrast, for conscientiousness, post-hoc analysis revealed no significant difference between DupB−/− males and females. However, DupB+/− males showed significantly less conscientiousness than DupB+/− females. Thus, sex differences in these personality dimensions only emerged in chimpanzees carrying the RS3 allele. No significant main effects or interactions were found for extraversion or agreeableness.
Figure 2.
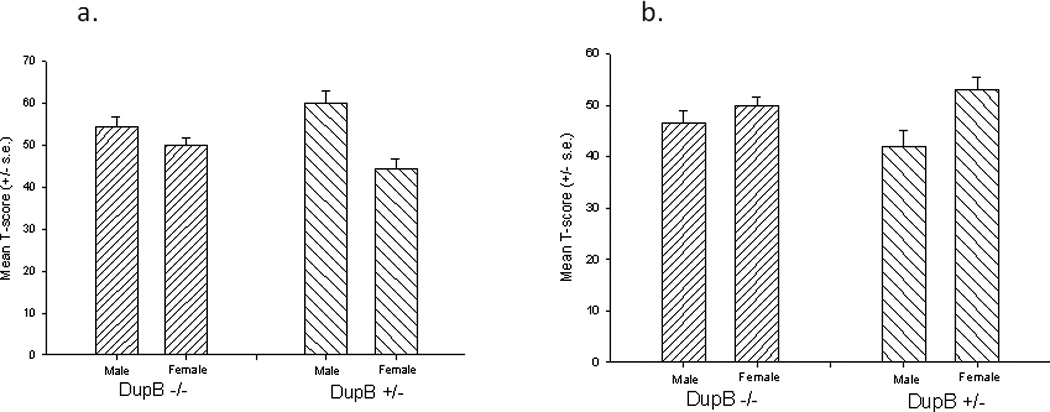
Mean T-scores (+/− s.e.) for the a) Dominance and b) Conscientiousness Traits for Male and Female DupB+/− and DupB−/− Chimpanzees
Item Rankings and AVPR1A
We next considered whether sex and genotype differences were evident for each item on the chimpanzee personality questionnaire. When considering the individual personality item rankings, we compared males and females within each genotype using a Mann-Whitey U-test because the data were on a ranked scale of measurement (see Table 2). For the DupB −/− chimpanzees, significant sex differences were found for only three items including fearful, active and aggressive. Females were ranked as more fearful while males were ranked as more active and aggressive. For the DupB+/− chimpanzees, significant sex differences were found for 18 items. Females were found to be more fearful, cautious, timid, sympathetic, submissive, and dependent than males. In contrast, males were found to be more dominant, persistent, reckless, playful, active, bullying, aggressive, manipulative, excitable, impulsive, defiant, and erratic than females.
Table 2.
Mean Rank Score on Each of the 43 Personality Items in Male and Female Chimpanzees with the Dup −/−and Dup +/− Genotype
| Dup −/− | Dup +/− | |||
|---|---|---|---|---|
| Item | Female | Male | Female | Male |
| Fearful | 3.78 | 2.89 | 3.78 | 2.42 |
| Dominant | 3.67 | 4.18 | 2.94 | 5.13 |
| Personable | 4.51 | 4.55 | 3.86 | 4.92 |
| Cautious | 4.10 | 3.55 | 4.97 | 2.54 |
| Stable | 3.99 | 3.74 | 4.11 | 3.63 |
| Autistic | 2.03 | 2.24 | 2.50 | 3.67 |
| Stingy | 4.07 | 3.58 | 3.22 | 4.08 |
| Jealous | 3.74 | 4.42 | 3.58 | 4.38 |
| Reckless | 3.29 | 3.68 | 2.14 | 4.38 |
| Sociable | 3.93 | 4.11 | 4.31 | 4.00 |
| Timid | 3.54 | 2.79 | 3.94 | 2.54 |
| Sympathetic | 3.24 | 3.16 | 4.19 | 3.13 |
| Playful | 3.19 | 4.03 | 3.06 | 4.17 |
| Solitary | 3.90 | 3.13 | 3.53 | 3.58 |
| Active | 3.63 | 4.68 | 3.28 | 4.58 |
| Helpful | 3.25 | 3.74 | 3.94 | 3.50 |
| Bullying | 3.49 | 4.08 | 2.61 | 4.54 |
| Aggressive | 2.94 | 3.76 | 2.11 | 4.58 |
| Manipulative | 4.01 | 3.74 | 3.17 | 4.42 |
| Gentle | 3.93 | 3.66 | 4.69 | 3.96 |
| Affectionate | 3.81 | 3.84 | 4.14 | 3.92 |
| Excitable | 3.75 | 4.05 | 3.47 | 4.83 |
| Impulsive | 3.34 | 3.26 | 2.83 | 4.50 |
| Inquisitive | 3.84 | 4.11 | 3.78 | 4.46 |
| Submissive | 3.72 | 3.11 | 4.36 | 2.29 |
| Dependent | 3.84 | 3.16 | 4.06 | 2.25 |
| Irritable | 3.59 | 3.34 | 2.86 | 3.79 |
| Predictable | 4.65 | 4.32 | 5.22 | 4.58 |
| Decisive | 4.63 | 4.42 | 4.00 | 4.42 |
| Depressed | 3.10 | 2.66 | 3.33 | 3.08 |
| Sensitive | 4.15 | 4.05 | 4.39 | 3.79 |
| Defiant | 3.50 | 3.55 | 2.64 | 4.46 |
| Intelligent | 4.34 | 4.03 | 4.39 | 4.29 |
| Protective | 3.44 | 3.39 | 3.83 | 3.62 |
| Inventive | 3.22 | 3.71 | 3.08 | 4.40 |
| Clumsy | 2.69 | 2.62 | 2.86 | 2.83 |
| Erratic | 2.88 | 3.45 | 2.58 | 3.73 |
| Friendly | 3.88 | 4.18 | 4.53 | 3.88 |
| Lazy | 3.41 | 2.97 | 4.00 | 3.13 |
| Disorganized | 2.79 | 3.34 | 3.06 | 3.25 |
| Unemotional | 3.50 | 2.82 | 3.14 | 2.67 |
| Imitative | 3.21 | 3.13 | 3.58 | 2.29 |
| Independent | 4.32 | 4.82 | 4.19 | 5.13 |
Bolded values indicate a significant sex difference as revealed by a Mann-Whitney U-test at p < .05.
Discussion
Chimpanzees with different polymorphisms in the 5’ flanking region of the AVPR1A promoter region show significant differences in the personality traits of dominance and conscientiousness. Moreover, the influence of this polymorphism on personality differs in males and females. DupB+/− males had higher dominance and lower conscientiousness scores than DupB+/− females. No significant differences in dominance or conscientiousness were found between DupB−/− females and males. Thus the sex differences in these traits depend on the presence of the RS3 containing DupB element. Not surprisingly, the DupB genotype by sex interaction on personality generalized to several items that loaded on the dominance and conscientiousness factors (see Table 2). In short, significant sex differences were found in 18 traits within the Dup +/− chimpanzees whereas only 3 items differed significantly between males and females within the Dup −/− subjects. Moreover, the sex differences found within the Dup −/− cohort were similarly found in the Dup +/− chimpanzees. Thus, the sex differences in personality found for the remaining 15 items were evident only in the Dup +/− cohort of chimpanzees.
Dominance in chimpanzees is complicated, and a number of factors or “styles” of behavioral interactions can influence the development and maintenance of rank (Foster et al., 2009). Dominant males are typically more aggressive and show higher levels of testosterone then lower ranking males, particularly in the presence of estrous females (Pusey et al., 1997). Notwithstanding, smaller males can achieve high ranking status by engaging in a significant amount of socially affiliative behaviors, notably grooming with many members of the group (Foster et al., 2009). Thus, at least in males, no single behavioral trait or style necessarily predicts whether an individual will eventually achieve a high or low dominance rank. In female chimpanzees, no obvious linear dominance hierarchies are always found, which is in contrast to other group living primates (Pusey et al., 1997, Wittig & Boesch, 2003). Thus, in chimpanzees, it is likely that the mechanisms that underlie dominance differ between males and females. Our results suggest that polymorphic variation in the AVPR1A promoter regions may be an important variable in the mediation of dominance behaviors chimpanzees, particularly among males.
Direct studies examining the role of AVPR1A variation and personality in humans, at least as measured by comparable tests to those used in this study with the chimpanzees, are lacking and therefore direct comparisons between species cannot be performed. However, several studies in humans have shown that polymorphisms in the RS3 region of AVPR1A are linked to some dimensions of social behavior such as novelty seeking and harm avoidance (Ebstein, 2006), male pair bonding and relationship quality (Meyer-Lindenberg et al., 2008, Prichard et al., 2007, Walum et al., 2008), altruism (Avinum et al., 2011), expressive dance and communication, as well as interests in music (Bachner-Melman et al., 2005, Ukkola-Vuoti et al., 2011).
Vasopressin systems within the brain are sexually dimorphic, and vasopressin is expressed at higher levels in males than in females (De Vries et al., 1992). Vasopressin is thought to regulate behaviors in sex-specific manner, typically regulating male typical social behaviors. Thus one would predict that genetic variation in AVPR1 would result in sex-specific effects on social behaviors. In at least one study, the effect of RS3 polymorphisms on human relationship quality was sex-specific, with the association being present in males but not females (Wallum et al., 2008). It has also been shown that intranasal administration of vasopressin increases agonistic facial expressions in response to unfamiliar faces and decreases perceptions of friendliness in men whereas in women, vasopressin administration elicits the opposite pattern of response (Thompson et al., 2006). Furthermore, intranasal vasopressin selectively impairs emotion recognition in men (Uzefovsky et al., 2012). This result is particularly intriguing in light of our finding of sexually dimorphic effects of DupB on several traits in apes.
Studies in voles have shown that individual differences in the avpr1a 5’ flanking region are associated with approach to novel odors, and the formation of pair bonds and partner preferences (Hammock & Young, 2005). Furthermore, central administration of vasopressin in male prairie voles selectively increases aggression and territoriality (Winslow et al., 1993), traits that seems to overlap with the items loading on the dominance factor in chimpanzees (see Tables 1 & 2). Vasopressin appears to have a conserved role in modulating the response to social cues in a sex-specific manner in a variety of species, regardless of the modality of those cues (Goodson and Bass, 2001; Donaldson and Young, 2008).
The extent to which traits, such as dominance and conscientiousness, are manifest in these types of social behaviors is unclear but this should be the focus of additional studies. Certainly one could argue that lower scores on certain adjectives such as “fearful” or “cautious” in the DupB−/− chimpanzees would suggest that DupB+/− individuals exhibit more of these traits which would manifest itself as more exploratory to novelty but, as previously stated, this awaits future testing using behavioral paradigms.
In summary, as far as we know, these are the first evidence of the influence of polymorphisms in the AVPR1A gene on behavioral profiles of chimpanzees and, in fact, nonhuman primates. The findings reported here should be viewed as an initial report that warrants further study, with a specific emphasis on overt behavioral responses and profiles of individual chimpanzees with different AVPR1A polymorphisms. It would be particularly interesting to determine whether DupB+/− and DupB−/− chimpanzees differ in the expression of the AVPR1a gene in the brain, since polymorphisms in the RS3 have been associated with variation in gene expression in the human brain as well as in in vitro transcription reporter assays(Knafo et al., 2008, Tansey et al., 2011). At this point we cannot conclude that the differences that we see in behaviors in our study are due directly to differences in gene expression caused by the presence or absence of the DupB region. It is possible that these polymorphisms are linked to other polymorphisms in the AVPR1A that are directly affecting AVPR1A expression or functionality. Regardless, our data does support the hypothesis that genetic variation at this locus is associated with variation in social behaviors. Moreover, as the chimpanzee genome has been mapped and publicly available for 6 years (Consortium, 2005) and recently updated (CGSC 2.1.3 panTro3), the potential for evaluating the influence of additional genes on social behavior of chimpanzees as they relate to human typical and atypical behavior should be a focus of continuing research efforts.
Acknowledgments
All aspects of this research adhered to the American Psychological Associations guidelines for the ethical treatment of animals in research. We would like to acknowledge support from NIMH grants MH56897 and MH64692 to LJY, an HHMI predoctoral fellowship to ZRD, NIMH grant MH-92923 to WDH and NIH RR00165 to YNPRC.
Literature Cited
- Avinum R, Israel S, Shalev I, Gritsenko I, Bornstein G, Ebstein RP, Knafo A. AVPR1A variant associated with preschoolers' lower altruistic behavior. PLosONE. 2011;6:E25274. doi: 10.1371/journal.pone.0025274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babb PL, Fernandez-Duque E, Schurr TG. AVPR1A sequence variation in monogomous owl monkeys (Aotus azarai) and its implications for the evolution of platyrrhine social behavior. J. Mol. Evol. 2010;71:279–297. doi: 10.1007/s00239-010-9383-6. [DOI] [PubMed] [Google Scholar]
- Bachner-Melman R, Dina C, Zohar AH, Constantini N, Lerer E, Hoch S, Sella S, Nemanov L, Gritsenko I, Lichtenberg P, Granot R, Ebstein RP. AVPR1a and SLC6A4 gene poloymorphisms are associated with creative dance performance. PlosGenetics. 2005;1:e42. doi: 10.1371/journal.pgen.0010042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Consortium TCSaA. Initial sequence of the chimpanzee genome and comparison with the human genome. Nature. 2005;437:69–87. doi: 10.1038/nature04072. [DOI] [PubMed] [Google Scholar]
- De Vries GJ, Crenshaw BD, Al-Shamma HA. Oxytocin in maternal, sexual and social behaviors: Gonadal steroid modulation of vasopresson pathways. Ann. N.Y. Acad. Sci. 1992;652:387–396. doi: 10.1111/j.1749-6632.1992.tb34369.x. [DOI] [PubMed] [Google Scholar]
- Donaldson ZR, Bai Y, Kondrashov FA, Stoinski TL, Hammock EAD, Young LJ. Evolution of a behavior-linked microsatellite-containing element of the 5' flanking region of the primate AVPR1A gene. BMC Evol. Biol. 2008;8:180–188. doi: 10.1186/1471-2148-8-180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donaldson ZR, Young LJ. Oxytocin, vasopressin and the neurogenetics of sociality. Science. 2008;322:900–904. doi: 10.1126/science.1158668. [DOI] [PubMed] [Google Scholar]
- Ebstein RP. The molecular genetic architecture of human personality: beyond self-report questionnaires. Mol. Psychiatry. 2006;11:427–445. doi: 10.1038/sj.mp.4001814. [DOI] [PubMed] [Google Scholar]
- Foster MW, Gilby IC, Murray CM, Johnson A, Wroblewski EE, Pusey AE. Alpha male grooming patterns: Implications for dominance "style". Am. J. Primatol. 2009;71:136–144. doi: 10.1002/ajp.20632. [DOI] [PubMed] [Google Scholar]
- Freeman HD, Gosling SD. Personality in nonhuman primates: a review and evaluation of past research. Am. J. Primatol. 2010;72:653–671. doi: 10.1002/ajp.20833. [DOI] [PubMed] [Google Scholar]
- Goodson JL, Bass AH. Social behavior functions and related anatomical characteristics of vasotocin/vasopressin systems in vertebrates. Brain Res. Rev. 2001;35:246–265. doi: 10.1016/s0165-0173(01)00043-1. [DOI] [PubMed] [Google Scholar]
- Gosling SD. From mice to men: what can we learn about personality from animal research? Psychol Bull. 2001;127:45–86. doi: 10.1037/0033-2909.127.1.45. [DOI] [PubMed] [Google Scholar]
- Hammock EA, Lim MM, Nair HP, Young LJ. Assocation of vasopressin 1a receptor levels with a regulatory microsattelite and behavior. Genes, Brain and Behavior. 2005;4:289–301. doi: 10.1111/j.1601-183X.2005.00119.x. [DOI] [PubMed] [Google Scholar]
- Hammock EA, Young LJ. Micosatellite instability generates diversity in brain and sociobehavioral traits. Science. 2005;308:1630–1634. doi: 10.1126/science.1111427. [DOI] [PubMed] [Google Scholar]
- Hammock EA, Young LJ. Oxytocin, vasopressin and pair bonding: implications for autism. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 2006;361:2187–2198. doi: 10.1098/rstb.2006.1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammock EA, Young LY. Functional microsatellite polymorphisms associated with divergent social structure in vole species. Mol. Biol. Evol. 2004;21:1057–1063. doi: 10.1093/molbev/msh104. [DOI] [PubMed] [Google Scholar]
- Hong KW, Weiss A, Morimura N, Udono T, Hayasaka I, Humle T, Murayama Y, Ito S, UInoue-Murayama M. Polymoprphism of the tryptophan hydroxylase 2 (TPH2) gene is associated with chimpanzee neuroticism. PlosOne. 2011;6:e22144. doi: 10.1371/journal.pone.0022144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King JE, Figueredo AJ. The five-factor model plus dominance in chimpanzee personality. Journal of Research on Personality. 1997;31:257–271. [Google Scholar]
- King JE, Weiss A, Farmer KH. A chimpanzee (Pan troglodytes) analogue of cross-natioanl generalization of personality structure: zoological parks and African santuary. Journal of Personality. 2005;73:389–410. doi: 10.1111/j.1467-6494.2005.00313.x. [DOI] [PubMed] [Google Scholar]
- Knafo A, Israel S, Darvasi A, Bachner0Melman R, Uzefovsky F, Cohen L, Feldman E, Lerer E, Laiba E, Raz Y, Nemanov L, Gritsenko I, Dina C, Agam G, Dean B, Bornstein G, Ebstein RP. Individual differences in allocation of funds in the Dictator Game and post-mortem hippocampal mRNA levels are correlated with length of the arginine vasopressin 1 a receptor (AVPR1A) RS3 promotor region repeat. Genes, Brain and Behavior. 2008;7:266–275. doi: 10.1111/j.1601-183X.2007.00341.x. [DOI] [PubMed] [Google Scholar]
- Lim MM, Nair HP, Young LJ. Species and sex differences in brain distribution of CRF receptor subtypes 1 and 2 in monogomous and promiscuous vole species. J. Comp. Neurol. 2005;487:75–92. doi: 10.1002/cne.20532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim MM, Wang Z, Olazabel DE, Ren X, Terwillinger EF, Young LJ. Enhanced partner prefernce in a promiscuous species by manipulating the expression of a single gene. Nature. 2004:754–757. doi: 10.1038/nature02539. [DOI] [PubMed] [Google Scholar]
- Melke J. Autism: Which genes are involved? Clinical Neuropsychiatry. 2008;5:63–69. [Google Scholar]
- Meyer-Lindenberg A, Domes G, Kirsch P, Heinrichs M. Oxytocin and vasopressin in the human brain: social neuropepetides for translational medicine. Nature Neuroscience Reviews. 2011;12:524–538. doi: 10.1038/nrn3044. [DOI] [PubMed] [Google Scholar]
- Meyer-Lindenberg A, Kolachana BS, Gold B, Olsh A, Nicodemus KK, Mattay V, Dean M, Weinberger DM. Genetic variants in AVPR1A linked to autism predict amygdala activation and personality traits in healthy humans. Mol. Psychiatry. 2008:1–8. doi: 10.1038/mp.2008.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prichard ZM, Mackinnon AJ, Jorm AF, Eastel S. AVPR1A and OXTR polymorphisms are associated with sexual and reproductive behavioral phenotypes in humans. Hum. Mutat. 2007;981:1–6. doi: 10.1002/humu.9510. [DOI] [PubMed] [Google Scholar]
- Pusey AE, Williams J, Goodall J. The influence of domiance rank on the reproductive success of female chimpanzees. Science. 1997;277:828–831. doi: 10.1126/science.277.5327.828. [DOI] [PubMed] [Google Scholar]
- Rosso L, Keller L, Kaessmann H, Hammond RL. Matng systems and avpr1a promoter variation in primates. Biology Letters. 2008;4:375–378. doi: 10.1098/rsbl.2008.0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tansey KE, Hill MJ, Cochrane LE, Gill M, Anney RJL, Gallagher L. Functionality of promoter microsatellites of arginine vasopressin receptor 1A (AVPR1A): implications for autism. Molecular Austism. 2011;2:1–8. doi: 10.1186/2040-2392-2-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson RR, George K, Walton JC, Orr SP, Benson J. Sex-specific influences of vasopresson on human social communication. Proceedings of the National Academy of Sciences. 2006;103:7889–7894. doi: 10.1073/pnas.0600406103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ukkola-Vuoti L, Oikkonen J, Onkamo P, Karma K, Raijas P, Jarvela I. Association of the argine vasopressin receptor 1A (AVPR1A) haplotypes with listening to music. J. Hum. Genet. 2011;56:324–329. doi: 10.1038/jhg.2011.13. [DOI] [PubMed] [Google Scholar]
- Uzefovsky F, Shalev I, Israel S, Knafo A, Ebstein RP. Vasopressin selectively impairs emotion recognition in men. Psychoneuroendocrinology. 2012;37:576–580. doi: 10.1016/j.psyneuen.2011.07.018. [DOI] [PubMed] [Google Scholar]
- Walum H, Westberg L, Henningsson S, Neiderhiser JM, Reiss D, Igl W, Ganiban JM, Spotts EL, Pedersen NL, Eriksson E, Lichenstein P. Genetic variation in the vasopressin receptor 1a gene (AVPR1A) associates with pair bonding behavior in humans. Proceedings of the National Academy of Sciences. 2008;105:14153–14156. doi: 10.1073/pnas.0803081105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss A, King JE, Hopkins WD. A cross-setting study of chimpanzee (Pan troglodytes) personality structure and development: Zoological parks and Yerkes National Primate Research Center. Am. J. Primatol. 2007;69:1264–1277. doi: 10.1002/ajp.20428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winslow JT, Hastings N, Carter CS, Harbaugh CR, Insel TR. The role of central vasopressin in pair bonding in monogamous praire voles. Nature. 1993;365:545–548. doi: 10.1038/365545a0. [DOI] [PubMed] [Google Scholar]
- Wittig RM, Boesch C. Food competition and linear dominance hierarchy among female chimpanzees of the Tai National Park. Int. J. Primatol. 2003;24:848–867. [Google Scholar]
- Young LJ, Wang Z. The neurobiology of pair bonding. Nature Neuroscience Reviews. 2004;7:1048–1054. doi: 10.1038/nn1327. [DOI] [PubMed] [Google Scholar]