

Abstract
Background:
DNA methylation is an important epigenetic mechanism in prostate cancer (PCa) progression. Given the role of even-skipped homeobox 1 (EVX1) in the regulation of multiple genes during embryogenesis, we postulated that EVX1 methylation is altered in PCa progression.
Methods:
Bisulphite sequencing and quantitative MethyLight were used to assess methylation in human prostate epithelial cells, four PCa cell lines, liver, lung, spleen, kidney, 35 paired tumour and tumour-associated benign tissues, and 11 normal prostate tissues. Prostate cancer cell lines were treated with 5-azacytidine (AzaC) or trichostatin A (TSA), and expression of EVX1 transcript and variants was assessed by qPCR. Hypermethylation was compared with clinicopathological features in a validation set of 58 patients using microarray.
Results:
Even-skipped homeobox 1 hypermethylation was observed in all four PCa cell lines and 57% of tumours. High-grade tumours exhibited increased methylation compared with intermediate-grade tumours. Even-skipped homeobox 1 expression was induced in PCa cell lines after treatment with AzaC or TSA. In the validation set, 83% of tumours were hypermethylated and hypermethylation was associated with worse recurrence-free survival.
Conclusion:
In this first evaluation of EVX1 methylation in human cancer, EVX1 is one of the most commonly hypermethylated genes observed in PCa and predicted treatment failure in moderate risk patients.
Keywords: EVX1 , methylation, prostate cancer, PSA, prognosis
The predominant tools for predicting prostate cancer (PCa) outcomes consist of clinical and pathologic nomograms incorporating PSA, clinical stage, Gleason grade (GG) and biopsy results (Kattan et al, 1999). The biological behaviour of PCa, especially intermediate-grade cancer (GG 5-7), exhibits a wide range of aggressive behaviour (Stark et al, 2009). Clearly, identifying new biological markers that allow improved detection and prognosis of PCa is critical. Prostate cancer, like many other malignancies, arises through an interplay of genetic and epigenetic alterations (Schulz and Hoffmann, 2009). DNA alterations at CpGs occur early in PCa development and represent a promising area for screening and prognostic marker development (Bastian et al, 2004).
There are increasing number of reports detailing the epigenetic gene modifications in PCa pathogenesis (Yegnasubramanian et al, 2004; Hanson et al, 2006; Henrique et al, 2006; Aitchison et al, 2007; Kawamoto et al, 2007). The most well-known target, GSTP1, is subject to epigenetic silencing in PCa, but its clinical applications are currently limited by its low sensitivity (Ellinger et al, 2008). In addition to cancer detection, epigenetic markers have the potential to predict cancer progression for selected diseases (Hartmann et al, 2009; Tanaka et al, 2011; Sun et al, 2012). In PCa, altered methylation at specific loci has been reported to correlate with clinicopathological features, but its role in predicting outcomes, especially for intermediate-grade, remains unknown (Sathyanarayana et al, 2003; Enokida et al, 2004). In addition to improving PCa detection, diagnosis, staging and risk stratification, epigenetics provides an opportunity to expand our knowledge of cancer biology (Nelson et al, 2009).
Recent studies have found aberrant methylation of embryonic developmental genes in cancer, suggesting a link between the cancer epigenome and developmental programs (Baylin and Jones, 2011). The Wnt signalling pathway, which is critical for normal cell growth and differentiation, was found to have a role in the development of urological cancers (Costa et al, 2010). Even-skipped homeobox 1 (EVX1) is a developmental homeobox domain protein located at 7p15 that is closely related to the Drosophila gene even-skipped (eve). Even-skipped homeobox 1 protein is expressed in the visceral endoderm and is required for normal blastocyst development and differentiation (Spyropoulos and Capecchi, 1994). Its transcripts are detected in undifferentiated F9 stem cells and disappear upon differentiation into parietal endoderm cells after the administration of retinoic acid (Faiella et al, 1991). Most notably, EVX1 functions as a potent repressor of gene transcription at numerous promoters in mammalian cells, suggesting a central role in the maintenance of normal gene expression (Briata et al, 1995). Currently, the role of EVX1 in cancer development is completely unknown. Given its important role in embryogenesis as a widespread gene repressor, we hypothesise that aberrant EVX1 methylation may have a role in prostate tumourigenesis.
In a recent study, we identified aberrant downstream hypermethylation of the developmental gene EVX1 on genome-wide methylation arrays to be associated with an epigenetic field defect in PCa (Yang et al, submitted for publication). In the present study, we extensively examine EVX1 methylation profiles in PCa within the promoter, exon 1 and intron 1 using bisulphite sequencing. Here, we provide evidence that EVX1 is frequently methylated in PCa and that its expression is regulated by DNA methylation and histone modification. Furthermore, EVX1 hypermethylation is an independent predictor of PSA recurrence following prostatectomy in intermediate-grade cancer.
Materials and methods
Cell culture
LNCaP, PPC-1, DU145 and PC3 cells were purchased from the American Type Culture Collection and cultured in DMEM (Life Technologies, Inc., Grand Island, NY, USA) with 10% fetal bovine serum and 1% penicillin-streptomycin. Human prostate epithelial cells (HPEC) were established on collagen-coated dishes in Ham's F-12 supplemented medium containing 1% fetal bovine serum as previously described (Schwarze et al, 2001).
Tissue samples
A total of 11 normal prostate specimens, termed ‘non-tumour associated’ (NTA) tissue, were obtained from either organ donation or cystoprostatectomy and OCT-embedded. Histological evaluation ruled out the presence of cancer cells. We also obtained 35 prostate specimens from patients with PCa who underwent radical prostatectomy. Microdissection was performed under the direction of a genitourinary pathologist to obtain tumour and benign ‘tumour-associated’ (TA) tissue apart from the tumour foci as previously described (Bhusari et al, 2011). In total, 19 intermediate-grade and 16 high-grade paired specimens along with 11 controls were obtained for DNA methylation analysis. Tissues were obtained in accordance with institutional review board policies.
DNA extraction and bisulphite conversion
DNA was isolated from prostate tissue samples and PCa cell lines using DNeasy Blood and Tissue Kit (QIAGEN, Valencia, CA, USA). Sodium bisulphite conversion was performed using Epitect Bisulfite Kit (QIAGEN).
Bisulphite sequencing
Three sets of primers were designed to amplify bisulphite-converted DNA spanning 25–450, 532–1018 and 1098–1484 bp downstream of the EVX1 transcriptional start site (TSS), using Methyl Primer Express Software (Applied Biosystems, Foster City, CA, USA). The following primer sets were used to amplify the three regions specified above: 5′-AGTGTGTGTGTTTTTTTGTTTG-3′ and 5′-AAAACCCCAAATAAATTCCTAA-3′ 5′-TTGGTATTTTGGTTGGTAAGAG-3′ and 5′-CAACCATCCTACCCTAAAAAA-3′ and 5′-GAGGGTGATAGTAGTGTTTTGA-3′ and 5′-AACAAAAACAATTAAACCATT-3′, respectively. Two separate HPEC DNA samples from individual patients (patients 1 and 2) and pooled HPEC DNA samples from multiple patients (patients 3–5) were bisulphite converted. PCR reactions were performed to amplify bisulphite-converted DNA using HotStarTaq (QIAGEN). The following cycling conditions were used: 95 °C for 15 min; 50 cycles of 95 °C for 15 s, 55 °C for 30 s, 72° for 90 s. PCR products were cloned into pCR2.1 using TOPO TA Cloning Kit (Invitrogen, Carlsbad, CA, USA). We collected 10–20 colonies for each of the three regions for each cell type being assessed. Plasmid isolation was performed using PureYield Plasmid Miniprep System (PROMEGA, Fitchburg, WI, USA) kit and sequenced. A total of 4–7 sequencing results per region per cell type were obtained and aligned using BiQ Analyzer (Bock et al, 2005).
In total, 96 CpGs within 2 CpG islands were analysed spanning a total of 1300 bp. The entire set of CpGs was compared using two-way analysis of variance (ANOVA) with Bonferroni post hoc testing for multiple comparisons. Fisher’s statistical analyses were performed using GraphPad Prism Version 5.03 (La Jolla, CA, USA).
Methylight assay
As we demonstrated with bisulphite sequencing that PCa cell lines and HPEC were differentially methylated, we designed MethyLight probes 460 bp downstream of the TSS in a region where PCa cell lines were highly methylated and HPEC were highly unmethylated. Probes sequences were designed using Beacon Designer (Premier Biosoft, Palo Alto, CA, USA).
The methylation-specific probe was tagged with 6-FAM and the unmethylated-specific probe was tagged with HEX. The following methylated-specific and unmethylated-specific probe sequences were synthesised: 5′-6-FAM-CCCTAAATCTCCAACGACCGAACGAATCT-3′ and 5′-HEX-CCCTAAATCTCCAACAACCAAACAAATCTT-3′, respectively (Integrated DNA Technologies, Coralville, IA, USA). Forward and reverse primers that do not contain any CpGs were designed: 5′-GTTTTATAAGGTGATTTTAGTTT-3′ and 5′-ACCCCAAATAAATTCCTAAC-3′, respectively. The primer annealing temperatures were optimised and 56 °C was ideal. With the exception of our optimised annealing temperature, the PCR reaction was carried out in duplicate using the Epitect MethyLight PCR Kit (QIAGEN) according to the manufacturer’s instructions. Real-time amplification and measurements of threshold cycles (Ct) for 6-FAM and HEX was performed using the CFX96 Real-Time PCR Detection System (BioRad, Hercules, CA, USA). The % methylation was calculated using the formula: 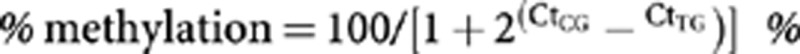 , where CtCG and CtTG are the measured Ct value from the real-time detection of 6-FAM and HEX, respectively (Eads et al, 2000). We used a t-test to compare groups. We generated a receiver operating characteristic (ROC) curve to determine the optimal cutoff value to define hypermethylation, which maximised the sum of sensitivity and specificity in predicting cancer in this cohort. This cutoff was then used to determine what percentage of tumour specimens were hypermethylated compared with the normal specimens. Receiver operating characteristic analysis was carried out using Graphpad Prism Version 5.03.
, where CtCG and CtTG are the measured Ct value from the real-time detection of 6-FAM and HEX, respectively (Eads et al, 2000). We used a t-test to compare groups. We generated a receiver operating characteristic (ROC) curve to determine the optimal cutoff value to define hypermethylation, which maximised the sum of sensitivity and specificity in predicting cancer in this cohort. This cutoff was then used to determine what percentage of tumour specimens were hypermethylated compared with the normal specimens. Receiver operating characteristic analysis was carried out using Graphpad Prism Version 5.03.
Experiments were performed for assay validation. DNA mixing of methylated LNCaP DNA and unmethylated kidney DNA was performed and was highly linear across a wide range of methylation (Supplemental Figure 1). Detection of methylation was highly accurate with as low as 40 pg of DNA template. Furthermore, the % methylation did not change with differing amounts of template DNA or between multiple experiments, demonstrating high efficiency and reproducibility of the MethyLight assay.
5-Azacytidine (AzaC) and trichostatin A (TSA) treatment
After performing a dose-ranging study, PCa cell lines were treated with 2.5 μℳ of the demethylating agent AzaC (Sigma-Aldrich, St Louis, MO, USA) for 48 h or 25 nℳ of the histone deacetylase TSA (Sigma-Aldrich) for 16 h. Cells were redosed with AzaC after the first 24 h of treatment. 5-Azacytidine stock solution was prepared in sterile water and TSA stock solution was prepared in DMSO, stored in aliquots at −20 °C.
Quantitative real-time PCR
Total RNA was isolated from drug-treated cells using the RNeasy RNA isolation kit (QIAGEN) and treated with DNase I (QIAGEN). The Omnicript Reverse Transcript Kit (QIAGEN) was used to synthesise cDNA using 200 ng of RNA per sample. Even-skipped homeobox 1 mRNA transcripts and variants were identified from expressed sequence tags (EST) found in the UCSC genome database (Dreszer et al, 2011). Primers were designed that spanned exons 1 and 2 (primer set 1) and exons 2 and 3 (primer set 2) of the forward expressing mRNA transcripts RefSeq NM_001989, EST AK314155 and EST-DC36605. We also designed a set of primers (primer set 3) that spanned exons 2 and 3 of EST-DB260732 and exons 1 and 2 of EST-DB455833, both of which are reverse-transcribing mRNA variants (Figure 4A). The following conditions were used for PCR amplification: 95 °C for 3 min; 40 cycles of 95 °C for 10 s, 55 °C for 30 s as described (Bhusari et al, 2011). All reactions were performed in duplicate. Quantification was performed by monitoring real-time fluorescence of Syto 9. Amplification and detection of PCR products was performed and threshold cycles (Ct) were measured using CFX96 Real-Time PCR Detection System (BioRad). Target gene expression was normalised to 18S expression and calculated to generate fold change. A t-test was used to compare the mean expression in treatment vs control groups.
Microarray
Kobayashi et al (2011) used the HumanMethylation27 (Illumina, San Diego, CA, USA) methylation microarray to identify hypermethylated genes in PCa. Using this patient population as a validation cohort, we assessed the association of clinicopathological features and the presence of EVX1 methylation. Using two probes, two CpGs were assayed, one located in the promoter region 70 bp upstream of the TSS (Microarray Probe no. 1) and one CpG in exon 1, 10 bp downstream of the TSS (Microarray Probe no. 2).
Association of clinical features before prostatectomy between the two groups of patients with or without hypermethylated tumours were assessed using Student’s t-test for normally distributed variables, whereas Wilcoxon rank sum test was used for non-normal distributed variables. For nominal variables, Fisher exact test was used. Kaplan–Meier survival analysis was performed to test for differences in time to biochemical recurrence between these two patient groups, using the Log-rank test. A multivariate Cox proportional hazards model was used to assess whether individual variables were independent predictors of biochemical recurrence using a forward selection method with a model entry criterion of P<0.05. Analyses were performed using SAS 9.3 for Windows (SAS Institute, Cary, NC, USA). All tests of significance were at the P<0.05 level, and two-tailed.
Results
Even-skipped homeobox 1 contains several CpG islands
We identified four CpG islands within the EVX1 gene using MethPrimer software (Li and Dahiya, 2002). Previously described CpG island criteria include a region greater than 200 bp with a observed/expected CpG ratio of >0.6 and CG percentage of >50% (Gardiner-Garden and Frommer, 1987). Three of the four CpG islands overlap exonic regions (Figure 1). We surveyed promoter methylation encompassing 96 CpGs found in CpG islands no. 1 and 2. In a validation set, microarray probes surrounding the TSS were used to compare methylation in tumour vs TA tissue. Even-skipped homeobox 1 CpG islands and the gene regions assayed are detailed in Figure 1.
Figure 1.

Genomic map of the EVX1 gene and its proximal promoter, and locations of methylation assays and probes. EVX1 is located at human 7p15 and has three exons. Three of the four CpG islands overlap exonic regions, and one CpG island is located within the first intron. Bisulphite sequencing was performed to assess methylation of the first two CpG islands. MethyLight probes were designed close to the transcription start site (TSS). A validation set consisted of two microarray probes, one probe located in the promoter region and one located 10 bp downstream of the TSS.
Bisulphite sequencing demonstrates EVX1 hypermethylation in PCa cell lines
Bisulphite conversion and cloning of DNA from PPC-1 and PC3 PCa cell lines was compared to HPECs cultured from non-tumour prostate tissue across 96 CpGs within CpG islands no 1 and 2, as described previously (Schwarze et al, 2001). Comparing the entire set of CpGs, PCa cell lines were significantly hypermethylated compared with HPEC lines (P<0.0001, two-way ANOVA with Bonferroni post-test). From CpGs no. 59–96, there was no difference between HPEC and PCa lines in 45/48 (94%) of the CpGs (P>0.05). However, in the region adjacent to the transcription start site, HPEC lines were significantly unmethylated. From CpGs no. 1–58, HPEC lines were hypomethylated in 54/58 (93%) of CpGs compared with PCa cell lines (P<0.05). There was no statistical difference between PPC-1 and PC3 (P=0.95). Thus, HPEC lines exhibited increasing methylation approaching intron 1, whereas PPC-1 and PC3 demonstrated a high density of methylation throughout the sequences analysed (Figure 2). This differential methylation between cancer lines and normal epithelial cells led to further analysis of the 5′ promoter region of CpG island no. 1.
Figure 2.

Bisulphite sequencing analysis of two CpG islands in prostate cancer (PCa) cell lines and normal human prostate epithelial cells (HPEC). Each circle indicates the average methylation of 4–7 sequenced clones at a single CpG location. The methylation profile of PC3 and PPC-1 cell lines was compared to that of two separate HPEC DNA samples from individual patients (patients 1 and 2) and pooled HPEC DNA samples from multiple patients (patients 3–5). Comparing the entire set of CpGs, PCa cell lines were significantly hypermethylated compared with HPEC lines (P<0.0001, two-way ANOVA with Bonferroni post-test). There was no difference between HPEC and PCa lines in downstream regions (P>0.05). However, in the region adjacent to the transcription start site, HPEC lines were significantly unmethylated (P<0.05). There was no statistical difference between PPC-1 and PC3 (P=0.95).
MethyLight demonstrates EVX1 promoter hypermethylation in PCa lines and PCa specimens
MethyLight is a quantitative assay utilising PCR amplification of bisulphite-converted DNA and binding of fluorescent labelled unmethylated and methylated probes (Eads et al, 2000). Prostate cancer cell lines PPC-1, DU145, PC3 and LNCaP demonstrated high levels of methylation (100.0%, 72.1%, 100.0% and 96%, respectively), whereas normal HPEC, liver, lung, spleen and kidney demonstrated low levels of methylation (26.3%, 22.8%, 22.8% and 24.9%, respectively) at the three CpGs assayed by the MethyLight probe (Figure 3A). A region in CpG island no. 1 was assayed by MethyLight (Figure 1). The HPEC DNA treated with SSSI methyltransferase, a cytosine-methylating enzyme, was used as a positive control.
Figure 3.

Quantitative MethyLight assessment of methylation in normal tissues, human prostate epithelial cells, prostate cancer cell lines and microdissected prostate tissue specimens. Bisulphite-treated DNA was analysed using PCR and binding of fluorescently labelled MethyLight probes distinguishing methylated and unmethylated CpGs. (A) Prostate cancer cell lines PPC-1, DU145, PC3 and LNCaP demonstrated high levels of methylation (100.0%, 72.1%, 100.0% and 96%, respectively), whereas normal HPEC, liver, lung, spleen and kidney demonstrated low levels of methylation (26.3%, 22.8%, 22.8% and 24.9%, respectively). HPEC DNA treated with a methylating agent (SSSI methylase) was used as a positive control. (B) Tumour specimens (T) demonstrated increased methylation compared with matched benign tumour-associated (TA) tissue, as well as normal peripheral prostate tissue from prostates not containing cancer (NTA; P=0.02 and P=0.001, respectively). When stratified according to grade, high-grade tumours (Gleason ⩾8) demonstrated increased methylation compared with intermediate-grade tumours (Gleason ⩽7; P=0.04). (C) A receiver operating characteristic (ROC) curve was constructed to determine the frequency of hypermethylation in tumour specimens (AUC 0.74, P=0.023). The optimum cutoff of 18.2% yielded a 55.9% sensitivity and 90.9% specificity for differentiating tumour from normal specimens, and was used to define hypermethylation. Using 18.2% as a cutoff, 20/35 (57%) of tumours were hypermethylated, while only 1/11 (9%) NTA tissues were hypermethylated (P=0.006).
Prostate cancer specimens were then analysed using MethyLight. Methylation was increased in tumour specimens compared to matched TA tissue, as well as normal peripheral prostate tissue from prostates not containing cancer (NTA; P=0.02 and P=0.001, respectively). We then stratified paired specimens according to grade and found that high-grade tumours (Gleason⩾8) demonstrated significantly higher methylation compared with intermediate-grade tumours (Gleason⩽7, P=0.04; Figure 3B). Patient characteristics for these prostate specimens are provided in Supplemental Table 1.
To determine the proportion of tumours specimens that were hypermethylated, a ROC curve was constructed yielding an area under the curve of 0.73 (P=0.023). The optimal cutoff of 18.2% methylation yielded a sensitivity of 55.9% and specificity of 90.9% for differentiating tumour vs NTA specimens, and was used to define hypermethylation (Figure 3C). Using 18.2% as a cutoff, 20/35 (57%) of tumours were hypermethylated, while only 1/11 (9%) NTA tissues were hypermethylated (P=0.006, two-tailed Fisher’s exact test).
Even-skipped homeobox 1 expression is regulated by DNA methylation and histone modification
To examine the effects of methylation on the expression of EVX1 mRNA and its variants, we treated PCa cell lines with the demethylating agent AzaC and the HDAC inhibitor TSA. A number of sense and antisense transcripts are expressed from the EVX1 gene (Figure 4A). 5-Azacytidine and TSA treatments were optimised for maximal gene expression and cell viability (data not shown). Using primer set 1, we found that AzaC treatment for 48 h resulted in a 2.6- and 3.9-fold induction of EVX1 in PPC-1 and PC3 cell lines (P=0.047 and P=0.014, respectively). 5-Azacytidine did not induce expression of downstream or reverse transcripts (Figure 4B). The HDAC inhibitor TSA induced the EVX1 expression with the use of primer set 1 in PPC-1 and PC3 (3.2- and 8.7-fold induction; P=0.048 and P=0.0095, respectively). Trichostatin A treatment also resulted in a 1.9-fold induction of the downstream transcript using primer set 2 (P=0.039) and 2.4-fold induction of the reverse transcripts using primer set 3 in PC3 cells (P=0.0023), an effect that was not observed with AzaC treatment (Figure 4C). Induction of transcripts was not significantly altered in LNCap and DU145 cells. 5-Azacytidine and TSA can often act synergistically to re-express certain genes (Cameron et al, 1999); however, this was not observed in any cell lines for EVX1 (data not shown).
Figure 4.

The EVX1 mRNA and splice variants and induction of EVX1 expression in human prostate cancer (PCa) cell lines. After drug exposure, RNA was isolated and expression quantitated utilising qPCR. (A) A select number of forward- and reverse-transcribing EVX1 transcripts were assayed using three different primer sets. (B) Re-expression of EVX1 mRNA was achieved after treating PPC-1 and PC3 cell lines with 2.5 μM of 5-azacytidine (AzaC) or 25 nM of trichostatin A (TSA; C) for 16 h. Using primer set 1, treatment of PPC-1 and PC3 cell lines with AzaC resulted in a 2.6-and 3.9-fold induction of EVX1 (P=0.047 and P=0.014, respectively), while TSA treatment resulted in a 3.2- and 8.7-fold induction (P=0.048 and P=0.0095, respectively). TSA treatment also resulted in a 1.9-fold induction of the downstream transcript using primer set 2 (P=0.039) and 2.4-fold induction of the reverse transcripts using primer set 3 (P=0.0023) in PC3 cells. *P<0.05, **P<0.01.
Even-skipped homeobox 1 hypermethylation is a strong independent predictor of PSA recurrence
Previously, genome-wide methylation profiling was conducted in 70 matched tumours and TA prostate tissues from patients that underwent radical prostatectomy for clinically localised PCa between 1998 and 2007 (Kobayashi et al, 2011). We analysed EVX1 methylation data at two different CpGs in 56 pairs where clinical data was available. These samples were primarily intermediate-grade (93%), comprising of GG six (19%) and GG seven (74%). Of paired samples assessed for EVX1 methylation differences, 48 out of 58 (83%) exhibited hypermethylation at one or both sites in tumour compared to adjacent tissue. Serum PSA was measured in these patients on follow-up visits after surgery (mean 56 months, median 49 months) and recurrence was defined as serum PSA>0.07 ng ml−1. Using the Kaplan–Meier analysis and the Log-rank test, we found that patients with hypermethylated tumours using the upstream Probe no. 1 had a significantly worse recurrence-free survival compared with patients without hypermethylation (51 vs 91% P=0.04; Figure 5). Furthermore, we analysed age, PSA, Gleason sum, SV involvement, extracapsular extension, peripheral node involvement, margins, pT category and EVX1 methylation at Probe no. 1 and 2 using multivariate Cox regression survival analysis and found that only pT category and EVX1 methylation (Probe no. 1) were independent predictors of PSA recurrence (Table 1). Probe no. 2 was not predictive of biochemical recurrence (P=0.6).
Figure 5.

A Kaplan–Meier survival analysis with respect to PSA recurrence was conducted in a validation set of 56 men that underwent prostatectomy for clinically localised prostate cancer. Of note, these samples were primarily intermediate-grade (93%), comprising of Gleason six (19%) and Gleason seven (74%). Patients with hypermethylated tumours had a significantly worse recurrence-free survival compared with patients without hypermethylation (51 vs 91% P=0.04, Log-rank test).
Table 1. Multivariate cox regression analysis for predicting biochemical recurrence.
| Variables a | Coefficent | s.e. | Wald | P | Odd ratio (95% CI) |
|---|---|---|---|---|---|
| Step 1: entry into model (P<0.05) | |||||
| pT category | 1.125 | 0.404 | 7.742 | 0.005 | 3.081 (1.395–6.807) |
| Step 2: entry into model (P<0.05) | |||||
| pT category | 1.296 | 0.452 | 8.241 | 0.004 | 3.656 (1.509–8.859) |
| EVX1 (Probe no. 1) | 2.114 | 1.061 | 3.967 | 0.046 | 8.282 (1.034–66.320) |
Variables analysed: age, PSA, Gleason sum, pT category, seminal vesical involvement, extracapsular extension, peripheral node involvement, margins, EVX1 probe no. 1, EVX1 probe no. 2
Discussion
Epigenetic alterations, such as gene promoter hypermethylation, show promise as potential tumour biomarkers. Even-skipped homeobox 1 (EVX1) is a gene important in blastocyst development and differentiation (Spyropoulos and Capecchi, 1994) that functions as a potent repressor of gene transcription in mammalian cells (Briata et al, 1995). Previously, a role for EVX1 in cancer had not been examined. In this study, EVX1 methylation was extensively surveyed across the promoter and 5′ regions in cell lines and tumour samples. We demonstrated that hypermethylation occurs in the majority of PCa samples, is more extensive in high-grade samples, and correlates with the risk of PSA failure in a validation cohort. This suggests a previously unrecognised biological role for this gene in PCa and as a marker for cancer and of risk of progression.
Mapping of methylation in cell lines demonstrated dense methylation of intron 1 and the 3′ region of exon 1 in HPECs, but absent methylation in upstream exonic regions including those surrounding the TSS. Prostate cancer cell lines were found to be uniformly densely methylated throughout both of these regions. These findings are consistent with the phenomenon of epigenetic spreading during aging and cancer in which CpG islands have the propensity to become methylated de novo by extension from surrounding methylated regions (Jones and Takai, 2001). Our data suggest this may occur early in the disease, as differential methylation at a downstream site was previously recognised in TA histologically normal tissues (i.e., field defect) when compared with prostate tissue from men without cancer (Yang et al, submitted for publication).
Using a quantitative MethyLight probe, we extended these findings and found that promoter hypermethylation occurred in over 57% of tumours compared to histologically normal tissues, and that hypermethylation at this locus is associated with a 6.2-fold increase in the presence of cancer, using a ROC curve. These results were confirmed by an independent validation set demonstrating that 83% of tumours exhibited hypermethylation. We observed that a higher frequency of methylation (83%) in this validation cohort, which may, in part, be explained by the close proximity of these probes to the TSS compared to our MethyLight probe (Figure 1). Alternatively, it may be due to the use of different assays. Gleason grade is a strong prognostic indicator and is linked to a number of important clinical end points, including pathological stage, progression to metastatic disease and survival (Egevad et al, 2002; Humphrey, 2004). Previously, Maruyama et al (2002) reported genes that had a qualitatively higher methylation in tumours with Gleason 7 or greater compared to Gleason 5–6. Liu et al (2011) found higher methylation of AKAP12 in Gleason 8–10 tumours compared with Gleason 4–7. Here, we demonstrate that EVX1 can quantitatively differentiate high-grade Gleason 8–10 vs intermediate-grade Gleason 6–7 tumours, thereby separating indolent from more aggressive tumours.
We extended our analysis to evaluate whether EVX1 has prognostic significance using an independent validation cohort. Notably, this cohort consisted primarily of intermediate-grade cancers (Table 2), a group that demonstrates biological variability (Stark et al, 2009), and in which it is more difficult to predict behaviour. We found that 83% of tumours were hypermethylated and that patients with EVX1 hypermethylation had only a 51% PSA recurrence-free survival compared with 91% in patients without hypermethylation. The high proportion of patients without recurrence (91%) when hypermethylation is absent indicates that EVX1 is a highly specific methylation marker for detecting recurrence, compared to other methylation markers such as AIM1 (Rosenbaum et al, 2011). Even in a cohort consisting of 93% intermediate-grade tumours (Gleason ⩽7), pT category and EVX1 methylation were independent predictors of biochemical recurrence with odds ratios of 7.742 and 3.967, respectively, using a multivariate Cox proportional hazards model. In another study, pT category and PGP9.5 methylation were found to be independent predictors of biochemical recurrence with odds ratios of 5.352 and 3.526, respectively (Mitsui et al, 2012). In this cohort, 29% of the patients were high-grade (Gleason >7). Previously, HOXD3 was reported to interact with stage (pT) in for predicting recurrence, but was not an independent predictor on multivariate analysis (Kron et al, 2010). Incorporation of EVX1 as a predictive marker into clinical practice may help to differentiate aggressive from indolent tumours in biopsy specimens, as well as provide a more accurate prediction of PSA recurrence following prostatectomy in intermediate-grade patients.
Table 2. Validation cohort characteristics.
| Number | 58 |
|---|---|
| Mean age | 61 |
| Mean PSA | 6.9 |
| Mean follow-up (months) | 57 |
| Gleason | |
| 3+3 | 11 (19%) |
| 3+4 | 35 (60%) |
| 4+3 | 8 (14%) |
| 4+4 | 1 (2%) |
| 4+5 | 3 (5%) |
| Stage | |
| T2 | 2 |
| T2a | 1 |
| T2b | 44 |
| T3a | 8 |
| T3b | 3 |
| PSA recurrence | 14 (24%) |
| Lymph nodes (+) | 40 (69%) |
| Surgical margins (+) | 5 (9%) |
| Extracapsular extension | 11 (19%) |
| Seminal vesicle (+) | 4 (7%) |
| Hypermethylated tumour | 46 (83%) |
Finally, treatment of PCa cell lines with AzaC or TSA can induce EVX1 expression. However, despite dose-ranging studies, generating re-expression in Du145 and LNCaP was difficult. These genes may contain mutations or other alterations that lead to gene inactivation. Unlike genetic mutations, the reversibility of these epigenetic changes makes them attractive targets for cancer therapy. Previous groups have shown that reversal of aberrant methylation with restoration of silenced gene expression using demethylating agents and HDAC inhibitors can decrease tumour growth in vivo and in vitro (Butler et al, 2001; Kuefer et al, 2004; Li et al, 2005; Rokhlin et al, 2006). 5-Azacytidine is currently in phase II clinical trials for the treatment of PCa (Jerónimo et al, 2011). The drawback of these agents is that they modulate the epigenome through widespread, non-specific changes in gene expression. Improving our understanding of the biology of these aberrantly methylated genes would allow for the therapeutic targeting of specific genes and signalling pathways.
In summary, our data indicate that EVX1 is epigenetically altered in PCa and may serve as a biological marker of the disease that can differentiate high-grade and intermediate-grade tumours and independently predict biochemical recurrence. Furthermore, these epigenetic alterations may influence the expression of EVX1, a gene important in embryological development. Future studies will focus on elucidating the biological role of this aberrantly-methylated gene in PCa.
Acknowledgments
We would like to thank Dr Wei Huang for pathological review of specimens and Pushpa Weeratunga for technical assistance. MT was funded by the Institute for Clinical and Translational Research Training Grant (NIH KL2 RR02012) and the University of Wisconsin School of Medicine Shapiro Grant. BY and DJ were funded by NIH 5RO1CA 097131.
Footnotes
Supplementary Information accompanies the paper on British Journal of Cancer website (http://www.nature.com/bjc)
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License.
Supplementary Material
References
- Aitchison A, Warren A, Neal D, Rabbitts P (2007) RASSF1A promoter methylation is frequently detected in both pre-malignant and non-malignant microdissected prostatic epithelial tissues. Prostate 67(6): 638–644 [DOI] [PubMed] [Google Scholar]
- Bastian PJ, Yegnasubramanian S, Palapattu GS, Rogers CG, Lin X, De Marzo AM, Nelson WG (2004) Molecular biomarker in prostate cancer: the role of CpG island hypermethylation. Eur Urol 46(6): 698–708 [DOI] [PubMed] [Google Scholar]
- Baylin SB, Jones PA (2011) A decade of exploring the cancer epigenome - biological and translational implications. Nat Rev Cancer 11(10): 726–734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhusari S, Yang B, Kueck J, Huang W, Jarrard DF (2011) Insulin-like growth factor-2 (IGF2) loss of imprinting marks a field defect within human prostates containing cancer. Prostate 71(15): 1621–1630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bock C, Reither S, Mikeska T, Paulsen M, Walter J, Lengauer T (2005) BiQ Analyzer: visualization and quality control for DNA methylation data from bisulfite sequencing. Bioinformatics 21(21): 4067–4068 [DOI] [PubMed] [Google Scholar]
- Briata P, Van De Werken R, Airoldi I, Ilengo C, Di Blas E, Boncinelli E, Corte G (1995) Transcriptional repression by the human homeobox protein EVX1 in transfected mammalian cells. J Biol Chem 270(46): 27695–27701 [DOI] [PubMed] [Google Scholar]
- Butler LM, Webb Y, Agus DB, Higgins B, Tolentino TR, Kutko MC, LaQuaglia MP, Drobnjak M, Cordon-Cardo C, Scher HI, Breslow R, Richon VM, Rifkind RA, Marks PA (2001) Inhibition of transformed cell growth and induction of cellular differentiation by pyroxamide, an inhibitor of histone deacetylase. Clin Cancer Res 7(4): 962–970 [PubMed] [Google Scholar]
- Cameron EE, Bachman KE, Myöhänen S, Herman JG, Baylin SB (1999) Synergy of demethylation and histone deacetylase inhibition in the re-expression of genes silenced in cancer. Nat Genet 21(1): 103–107 [DOI] [PubMed] [Google Scholar]
- Costa VL, Henrique R, Ribeiro FR, Carvalho JR, Oliveira J, Lobo F, Teixeira MR, Jerónimo C (2010) Epigenetic regulation of Wnt signaling pathway in urological cancer. Epigenetics 5(4): 343–351 [DOI] [PubMed] [Google Scholar]
- Dreszer TR, Karolchik D, Zweig AS, Hinrichs AS, Raney BJ, Kuhn RM, Meyer LR, Wong M, Sloan CA, Rosenbloom KR, Roe G, Rhead B, Pohl A, Malladi VS, Li CH, Learned K, Kirkup V, Hsu F, Harte RA, Guruvadoo L, Goldman M, Giardine BM, Fujita PA, Diekhans M, Cline MS, Clawson H, Barber GP, Haussler D, James Kent W (2011) The UCSC Genome Browser database: extensions and updates 2011. Nucleic Acids Res 40(D1): D918–D923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eads CA, Danenberg KD, Kawakami K, Saltz LB, Blake C, Shibata D, Danenberg PV, Laird PW (2000) MethyLight: a high-throughput assay to measure DNA methylation. Nucleic Acids Res 28(8): E32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egevad L, Granfors T, Karlberg L, Bergh A, Stattin P (2002) Prognostic value of the Gleason score in prostate cancer. BJU Int 89(6): 538–542 [DOI] [PubMed] [Google Scholar]
- Ellinger J, Haan K, Heukamp LC, Kahl P, Büttner R, Müller SC, von Ruecker A, Bastian PJ (2008) CpG island hypermethylation in cell-free serum DNA identifies patients with localized prostate cancer. Prostate 68(1): 42–49 [DOI] [PubMed] [Google Scholar]
- Enokida H, Shiina H, Igawa M, Ogishima T, Kawakami T, Bassett WW, Anast JW, Li LC, Urakami S, Terashima M, Verma M, Kawahara M, Nakagawa M, Kane CJ, Carroll PR, Dahiya R (2004) CpG hypermethylation of MDR1 gene contributes to the pathogenesis and progression of human prostate cancer. Cancer Res 64(17): 5956–5962 [DOI] [PubMed] [Google Scholar]
- Faiella A, D'Esposito M, Rambaldi M, Acampora D, Balsofiore S, Stornaiuolo A, Mallamaci A, Migliaccio E, Gulisano M, Simeone A (1991) Isolation and mapping of EVX1, a human homeobox gene homologous to even-skipped, localized at the 5′ end of HOX1 locus on chromosome 7. Nucleic Acids Res 19(23): 6541–6545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardiner-Garden M, Frommer M (1987) CpG islands in vertebrate genomes. J Mol Biol 196(2): 261–282 [DOI] [PubMed] [Google Scholar]
- Hanson JA, Gillespie JW, Grover A, Tangrea MA, Chuaqui RF, Emmert-Buck MR, Tangrea JA, Libutti SK, Linehan WM, Woodson KG (2006) Gene promoter methylation in prostate tumor-associated stromal cells. J Natl Cancer Inst 98(4): 255–261 [DOI] [PubMed] [Google Scholar]
- Hartmann O, Spyratos F, Harbeck N, Dietrich D, Fassbender A, Schmitt M, Eppenberger-Castori S, Vuaroqueaux V, Lerebours F, Welzel K, Maier S, Plum A, Niemann S, Foekens JA, Lesche R, Martens JW (2009) DNA methylation markers predict outcome in node-positive, estrogen receptor-positive breast cancer with adjuvant anthracycline-based chemotherapy. Clin Cancer Res 15(1): 315–323 [DOI] [PubMed] [Google Scholar]
- Henrique R, Costa VL, Cerveira N, Carvalho AL, Hoque MO, Ribeiro FR, Oliveira J, Teixeira MR, Sidransky D, Jerónimo C (2006) Hypermethylation of Cyclin D2 is associated with loss of mRNA expression and tumor development in prostate cancer. J Mol Med (Berl) 84(11): 911–918 [DOI] [PubMed] [Google Scholar]
- Humphrey PA (2004) Gleason grading and prognostic factors in carcinoma of the prostate. Mod Pathol 17(3): 292–306 [DOI] [PubMed] [Google Scholar]
- Jerónimo C, Bastian PJ, Bjartell A, Carbone GM, Catto JW, Clark SJ, Henrique R, Nelson WG, Shariat SF (2011) Epigenetics in prostate cancer: biologic and clinical relevance. Eur Urol 60(4): 753–766 [DOI] [PubMed] [Google Scholar]
- Jones PA, Takai D (2001) The role of DNA methylation in mammalian epigenetics. Science 293(5532): 1068–1070 [DOI] [PubMed] [Google Scholar]
- Kattan MW, Wheeler TM, Scardino PT (1999) Postoperative nomogram for disease recurrence after radical prostatectomy for prostate cancer. J Clin Oncol 17(5): 1499–1507 [DOI] [PubMed] [Google Scholar]
- Kawamoto K, Okino ST, Place RF, Urakami S, Hirata H, Kikuno N, Kawakami T, Tanaka Y, Pookot D, Chen Z, Majid S, Enokida H, Nakagawa M, Dahiya R (2007) Epigenetic modifications of RASSF1A gene through chromatin remodeling in prostate cancer. Clin Cancer Res 13(9): 2541–2548 [DOI] [PubMed] [Google Scholar]
- Kobayashi Y, Absher DM, Gulzar ZG, Young SR, McKenney JK, Peehl DM, Brooks JD, Myers RM, Sherlock G (2011) DNA methylation profiling reveals novel biomarkers and important roles for DNA methyltransferases in prostate cancer. Genome Res 21(7): 1017–1027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kron KJ, Liu L, Pethe VV, Demetrashvili N, Nesbitt ME, Trachtenberg J, Ozcelik H, Fleshner NE, Briollais L, van der Kwast TH, Bapat B (2010) DNA methylation of HOXD3 as a marker of prostate cancer progression. Lab Invest 90(7): 1060–1067 [DOI] [PubMed] [Google Scholar]
- Kuefer R, Hofer MD, Altug V, Zorn C, Genze F, Kunzi-Rapp K, Hautmann RE, Gschwend JE (2004) Sodium butyrate and tributyrin induce in vivo growth inhibition and apoptosis in human prostate cancer. Br J Cancer 90(2): 535–541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li LC, Carroll PR, Dahiya R (2005) Epigenetic changes in prostate cancer: implication for diagnosis and treatment. J Natl Cancer Inst 97(2): 103–115 [DOI] [PubMed] [Google Scholar]
- Li LC, Dahiya R (2002) MethPrimer: designing primers for methylation PCRs. Bioinformatics 18(11): 1427–1431 [DOI] [PubMed] [Google Scholar]
- Liu W, Gong J, Hu J, Hu T, Sun Y, Du J, Sun C, Guan M, Jiang H, Lu Y (2011) Quantitative assessment of AKAP12 promoter methylation in human prostate cancer using methylation-sensitive high-resolution melting: correlation with Gleason score. Urology 77(41006): e1–e7 [DOI] [PubMed] [Google Scholar]
- Maruyama R, Toyooka S, Toyooka KO, Virmani AK, Zöchbauer-Müller S, Farinas AJ, Minna JD, McConnell J, Frenkel EP, Gazdar AF (2002) Aberrant promoter methylation profile of prostate cancers and its relationship to clinicopathological features. Clin Cancer Res 8(2): 514–519 [PubMed] [Google Scholar]
- Mitsui Y, Shiina H, Hiraki M, Arichi N, Hiraoka T, Sumura M, Honda S, Yasumoto H, Igawa M (2012) Tumor suppressor function of PGP9.5 is associated with epigenetic regulation in prostate cancer--novel predictor of biochemical recurrence after radical surgery. Cancer Epidemiol Biomarkers Prev 21(3): 487–496 [DOI] [PubMed] [Google Scholar]
- Nelson WG, De Marzo AM, Yegnasubramanian S (2009) Epigenetic alterations in human prostate cancers. Endocrinology 150(9): 3991–4002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rokhlin OW, Glover RB, Guseva NV, Taghiyev AF, Kohlgraf KG, Cohen MB (2006) Mechanisms of cell death induced by histone deacetylase inhibitors in androgen receptor-positive prostate cancer cells. Mol Cancer Res 4(2): 113–123 [DOI] [PubMed] [Google Scholar]
- Rosenbaum E, Begum S, Brait M, Zahurak M, Maldonado L, Mangold LA, Eisenberger MA, Epstein JI, Partin AW, Sidransky D, Hoque MO (2011) AIM1 promoter hypermethylation as a predictor of decreased risk of recurrence following radical prostatectomy. Prostate; e-pub ahead of print 29 November 2011; doi:10.1002/pros.22461 [DOI] [PMC free article] [PubMed]
- Sathyanarayana UG, Padar A, Suzuki M, Maruyama R, Shigematsu H, Hsieh JT, Frenkel EP, Gazdar AF (2003) Aberrant promoter methylation of laminin-5-encoding genes in prostate cancers and its relationship to clinicopathological features. Clin Cancer Res 9(17): 6395–6400 [PubMed] [Google Scholar]
- Schulz WA, Hoffmann MJ (2009) Epigenetic mechanisms in the biology of prostate cancer. Semin Cancer Biol 19(3): 172–180 [DOI] [PubMed] [Google Scholar]
- Schwarze SR, Shi Y, Fu VX, Watson PA, Jarrard DF (2001) Role of cyclin-dependent kinase inhibitors in the growth arrest at senescence in human prostate epithelial and uroepithelial cells. Oncogene 20(57): 8184–8192 [DOI] [PubMed] [Google Scholar]
- Spyropoulos DD, Capecchi MR (1994) Targeted disruption of the even-skipped gene, evx1, causes early postimplantation lethality of the mouse conceptus. Genes Dev 8(16): 1949–1961 [DOI] [PubMed] [Google Scholar]
- Stark JR, Perner S, Stampfer MJ, Sinnott JA, Finn S, Eisenstein AS, Ma J, Fiorentino M, Kurth T, Loda M, Giovannucci EL, Rubin MA, Mucci LA (2009) Gleason score and lethal prostate cancer: does 3+4=4+3? J Clin Oncol 27(21): 3459–3464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun W, Zaboli D, Wang H, Liu Y, Arnaoutakis D, Khan T, Khan Z, Koch WM, Califano JA (2012) Detection of TIMP3 promoter hypermethylation in salivary rinse as an independent predictor of local recurrence-free survival in head and neck cancer. Clin Cancer Res 18(4): 1082–1091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, Chang P, Li Y, Li D, Overman M, Maru DM, Sethi S, Phillips J, Bland GL, Abbruzzese JL, Eng C (2011) Association of CHFR promoter methylation with disease recurrence in locally advanced colon cancer. Clin Cancer Res 17(13): 4531–4540 [DOI] [PubMed] [Google Scholar]
- Yegnasubramanian S, Kowalski J, Gonzalgo ML, Zahurak M, Piantadosi S, Walsh PC, Bova GS, De Marzo AM, Isaacs WB, Nelson WG (2004) Hypermethylation of CpG islands in primary and metastatic human prostate cancer. Cancer Res 64(6): 1975–1986 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.