

Abstract
Regulated movements of the nucleus are essential during zygote formation, cell migrations, and differentiation of neurons. The nucleus moves along microtubules (MTs) and is repositioned on F-actin at the cellular cortex. Two families of nuclear envelope proteins, SUN and KASH, link the nucleus to the actin and MT cytoskeletons during nuclear movements. However, the role of actin nucleators in nuclear migration and positioning is poorly understood. We show that the branched actin nucleator, Arp2/3, affects nuclear movements throughout embryonic and larval development in C. elegans, including nuclear migrations in epidermal cells and neuronal precursors. In one-cell embryos the migration of the male pronucleus to meet the female pronucleus after fertilization requires Arp2/3. Loss of Arp2/3 or its activators changes the dynamics of non-muscle myosin, NMY-2, and alters the cortical accumulation of posterior PAR proteins. Reduced establishment of the posterior microtubule cytoskeleton in Arp2/3 mutants correlates with reduced male pronuclear migration. The UNC-84/SUN nuclear envelope protein that links the nucleus to the MT and actin cytoskeleton is known to regulate later nuclear migrations. We show here it also positions the male pronucleus. These studies demonstrate a global role for Arp2/3 in nuclear migrations. In the C. elegans one-cell embryo Arp2/3 promotes the establishment of anterior/posterior polarity and promotes MT growth that propels the anterior migration of the male pronucleus. In contrast with previous studies emphasizing pulling forces on the male pronucleus, we propose that robust MT nucleation pushes the male pronucleus anteriorly to join the female pronucleus.
Keywords: C. elegans, cell polarity, early embryonic development, actin nucleation, nuclear migration
INTRODUCTION
The nucleus, the largest organelle in the cell, assumes a distinct position during different cellular events. When a cell undergoes a polarized movement, the position of the nucleus is tightly controlled. During mammalian wound healing, fibroblast migration, and neuronal migrations, the nucleus is often reoriented so that the associated microtubule organizing center (MTOC) faces the direction of migration (reviewed in (Gundersen et al., 2005). Microtubules can also position the nucleus independently of centrosome position, for example in migrating granule cells in the mouse brain (Umeshima et al., 2007). In the lower eukaryote, Saccharomyces cerevisiae, when two cells of the opposite mating type form a mating projection or shmoo, the nucleus is moved toward the projection in preparation for karyogamy or nuclear fusion (reviewed in (Molk and Bloom, 2006; Rose, 1996).
The nucleus is attached to the microtubule (MT) cytoskeleton. MTs are often anchored at the cellular cortex, which is rich in filamentous actin (F-actin). How the actin and MT cytoskeletons work together to support movements of the nucleus is best understood in budding yeast during cell divisions. The spindle microtubules are captured by actin-associated cortical structures in the developing bud, allowing the spindle to align along the correct axis in preparation for cell division (reviewed in (Heil-Chapdelaine et al., 1999; Huisman and Segal, 2005; Irazoqui and Lew, 2004; Pruyne et al., 2004).
During C. elegans embryonic development it has been proposed that spindle polarity similarly relies on MT capture by factors associated with cortical actin. In the one-cell embryo actin becomes enriched at the anterior cortex, and this anterior cortical actin is proposed to recruit proteins that capture spindle MTs so that the spindle rotates to permit an anterior/posterior cell division (Hyman, 1989; White and Strome, 1996).
In the C. elegans one-cell embryo two nuclear migrations occur. The female pronucleus migrates posteriorly to meet the male pronucleus, while the male pronucleus first migrates to the posterior pole, makes contact with the cortex, and then migrates anteriorly to meet the female pronucleus. The stage when these pronuclear migrations happen is of particular interest because it is when the initial anterior/posterior (A/P) polarity of the embryo is established, in response to a cue from the sperm-derived centrosomes (Cowan and Hyman, 2004a; Galli and van den Heuvel, 2008; Goldstein and Hird, 1996; Munro and Bowerman, 2009; O’Connell et al., 2000; Sadler and Shakes, 2000; Wallenfang and Seydoux, 2000).
We have previously analyzed the role of WAVE/SCAR and Arp2/3 protein complexes during the movements of the epidermis to enclose the embryo in C. elegans (Patel et al., 2008; Soto et al., 2002). These complexes are responsible for assembling branched actin that provides the force to drive diverse cellular events including cellular protrusions at the plasma membrane during cell migrations (reviewed in (Chhabra and Higgs, 2007; Derivery and Gautreau, 2010; Kurisu and Takenawa, 2009; Pollard, 2007). The role of branched actin in nuclear migrations has not previously been examined.
We report here that the WAVE/SCAR and Arp2/3 protein complexes that promote branched actin nucleation have an essential role during nuclear migrations throughout embryonic and larval development. We describe a role for Arp2/3 during epidermal cell migrations, when lateral nuclear movements accompany the convergent extension-like movements of the dorsal epidermis. We also uncover a role for Arp2/3 during the nuclear migrations of the P neuroblasts that result in the differentiation of neuronal cells. Loss of Arp2/3 results in the death of neurons due to failed nuclear migrations. Finally, we find that the anterior migration of the male pronucleus in the one-cell embryo, during zygote formation, depends on Arp2/3. This nuclear migration occurs in the absence of cell migration and thus provides an ideal context in which to study the contribution of Arp2/3 to nuclear migrations. Our results show that Arp2/3-dependent actin nucleation promotes nuclear movements in the zygote by regulating two cytoskeletal systems. First, it contributes to actomyosin contractility that sets up A/P polarity, and second, it promotes MT growth that appears to facilitate the movement of the male pronucleus away from the posterior cortex. To investigate the molecules that mediate the Arp2/3-dependent F-actin role in nuclear movements, we compared the phenotypes of the known C. elegans nuclear migration mutants. Mutations in the UNC-84 SUN domain protein that localizes to the nuclear envelope (Malone et al., 1999) result in similar male pronuclear migration defects as loss of Arp2/3 and its regulators. These results strongly support a role for Arp2/3-dependent actin nucleation in promoting the growth of MTs that connect the nucleus to the cellular cortex during nuclear migrations.
MATERIALS AND METHODS
Nematode genetics
C. elegans strains were cultured as previously described (Brenner, 1974), if not mentioned otherwise. The strains used in this study include: nhr-25∷yfp, oxIs12 [unc-47∷gfp] (McIntire et al., 1997), OX361 [wve-1(ne350)/unc-101; oxIs12], OX362 [gex-3(zu196)/dnt-1; oxIs12], OX363 [unc-83(ku18); oxIs12]; OX364 [unc-84(e1410); oxIs12]; wve-1(ne350), gex-3(zu196), unc-83(ku18) (Starr et al., 2001), unc-84(e1410), anc-1(e1873), OD73(unc-119(ed3) III; ltIs38 [pAA1; pie-1p∷gfp∷ph(PLC1delta1); unc-119(+)]; ltIs24 [pAZ132; pie-1p∷gfp∷tba-2; unc-119(+)]) (Audhya et al., 2005), ltIs24 [pAZ132; pie-1p∷gfp∷tba-2; unc-119(+)] was isolated from OD73 by out crossing OD73 with N2 males, zuIs45[nmy-2p∷nmy-2∷gfp], BV67 [pie-1p∷lifeact∷gfp] (Pohl and Bao, 2010), itIs153 [pie-1p∷gfp∷par-2] (Cuenca et al., 2003), xnIs3 [par-6p∷par-6∷gfp; unc-119(+)] (Anderson et al., 2008), TH120 [gfp∷par-2; mCherry∷par-6] (Schonegg et al., 2007) and OX84 [nhr-25∷yfp; rrf-3] (Baugh et al., 2005; Patel et al., 2008). The temperature-sensitive strain zyg-12(or577) was maintained at the permissive temperature (15°C); larval stage 4 (L4) worms transferred to non-permissive 25°C gave rise to embryos exhibiting full penetrance of the detached centrosome phenotype (Malone et al., 2003). The par-2∷gfp (Cuenca et al., 2003) and par-6∷gfp strains were maintained at 25°C for optimal expression of the transgenes.
Monitoring RNAi in Gex and nuclear migration strains
All RNAi experiments were performed by feeding HT115 bacteria containing the respective gene of interest as described in (Timmons et al., 2001). Feeding RNAi constructs for gex genes as described (Patel et al., 2008); strains for unc-83, unc-84, and par-2 from the Ahringer RNAi library, verified by sequencing (Kamath and Ahringer, 2003). Effective RNAi against unc-83 and unc-84 resulted in ectopic nuclei in the dorsal cord of >95% of the L1 and L2 larval animals. Effective RNAi against gex genes and par-2 resulted in dead embryos with the expected Gex and Par embryonic phenotypes.
Analysis of nuclear migration in P cell derived motor neurons
The strains oxIs12 [unc-47∷gfp] (McIntire et al., 1997), OX361 [wve-1(ne350)/unc-101; oxIs12], OX362 [gex-3(zu196)/dnt-1; oxIs12], OX363 [unc-83(ku18); oxIs12]; OX364 [unc-84(e1410); oxIs12] were propagated at 25°C for analysis of nuclear migration defects in P cells. RNAi knock-down of wve-1, gex-3 or arp-2 in unc-47∷gfp worms was carried out by feeding synchronized larval stage 1 (L1) worms with the respective bacterial strains at 25°C. L4 animals were mounted onto 3% agarose pads for counting of the 16 P cell derived GFP positive nuclei in ventral nerve cord, excluding the nucleus in the tail and the nuclei cluster parallel to the pharynx region.
Imaging of embryos and larvae
One-cell embryos were dissected from adult hermaphrodites in M9 buffer on a cover slip. The cover slip was mounted on a 3% agarose pad and sealed with Vaseline during DIC or fluorescence time-lapse imaging. DIC movies were acquired at a single plane in 15 sec intervals; fluorescence time-lapse imaging was documented in 30 sec intervals each time point included a stack of 5 images at 1μm depth. Male pronuclear migration distance, pseudocleavage ingression or anterior shift of NMY-2∷GFP boundary was determined using the line tool in ImageJ. Embryo length and male pronuclear migration distance were measured from images captured at pronuclear meeting (PNM), the time when the two pronuclei meet and their adjacent membranes form a straight line. Only embryos where the positions of the male and female pronucleus were observed prior to PNM were included in the analysis. Imaging of ventral P nuclei in live unc-47∷gfp larvae (L4) and time-lapse imaging of one-cell embryos were performed on a Zeiss Axioskop 2 Plus microscope using a 40x oil objective with iVision 4.0 software driving a Cooke SensiCam QE camera, and analyzed using ImageJ software.
Laser spinning disc confocal microcopy
Live nhr-25∷yfp or gfp∷tba-2 embryos were mounted on poly-L-lysine-coated coverslips in embryo buffer. The coverslip was place on standard microscope slides and sealed with silicone oil. Embryos were imaged using a 40×1.3 NA oil immersion lens on a Nikon TE2000 inverted microscope fitted with a Yokogawa CSU21 spinning-disk confocal scanhead (Perkin Elmer), a Melles-Griot argon laser (514 nm excitation) controlled by a Neos programmable AOTF. Multidimensional datasets were acquired using the software MetaMorph on a Hamamatsu Orca-AG cooled CCD camera, and stereo QuickTimeVR movies were assembled from the raw data using a custom-written plug-in for the Java program ImageJ (http://rsb.info.nih.gov/ij/). Live lifeact∷gfp and TH120 [gfp∷par-2; mCherry∷par-6] embryos were prepared as described above prior to imaging using a 40x oil immersion lens on a Zeiss Cell Observer microscope with a Yokogawa CSU-X1 confocal scanning unit. Multidimensional time-lapse imaging was documented using Zeiss AxioVision software operating an Evolve EMCCD camera (Photometrics). Images taken at the time points of interest were analyzed using the software ImageJ.
Particle count and fluorescence intensity measurements
Cortical projection images of NMY-2∷GFP or Lifeact∷GFP at the respective time points were obtained by combining Z-stacks of 4-5 cortical images taken at 1μm intervals. To count the cortical NMY-2∷GFP or Lifeact∷GFP foci, the cytoplasmic background fluorescence was removed from the cortical projection images using the ImageJ plug-in ‘Subtract Background’ (with a ‘Rolling ball radius’ at 50.0 pixels for NMY∷GFP or 20 pixels for Lifeact∷GFP) before converting the processed images into binary images. Subsequently the binary images were used for counting of the cortical NMY-2∷GFP or Lifeact∷GFP foci using the ImageJ plug-in ‘Analyze Particles’.
Fluorescence intensity of mCherry∷PAR-6 and GFP∷PAR-2 at the membrane was measured using the line tool in ImageJ. Fluorescence intensity in the anterior membranes was determined by placing the line tool at three short regions (approximately 5 μm each) of the anterior membrane of the one-cell embryo, and averaging the fluorescence intensity of these three lines to represent the anterior membrane signal. The fluorescence intensity of the posterior membrane was measured as described for the anterior membrane. The fluorescence intensity ratio between the anterior and the posterior membrane was plotted for the indicated time points.
GFP∷TBA-2 images were processed using the ImageJ plug-in ‘Subtract Background’ with a ‘Rolling ball radius’ at 50.0 pixels for all images and times points of interest prior to examination of the centrosomal MT size using the circle tool in ImageJ. A circle was placed around the centrosomal MT region to measure the size of the area marked by GFP∷TBA-2 fluorescence signal. The same adjustments were made for all of the images. The area size and fluorescence intensity of the centrosomal MT region decorated with GFP∷TBA-2 were determined using the ‘Measure’ command in ImageJ.
RESULTS
Nuclear migrations in the embryonic epidermis require the WAVE/SCAR and Arp2/3 complexes
The WAVE/SCAR and Arp2/3 complexes are essential during epidermal cell migrations in C. elegans. Loss of any component of WAVE/SCAR or Arp2/3 blocks the movements of the 6 rows of epidermal cells, leading to the Gex, or gut on the exterior, phenotype in which the epidermal cells fail to enclose the embryo. The ventral cells fail to initiate ventralward movements to enclose the embryo while the two dorsal rows fail to intercalate to elongate the epidermis along the anterior/posterior axis (Patel et al., 2008; Soto et al., 2002). While examining epidermal migrations in WAVE/SCAR and Arp2/3 mutants, we additionally detected a failure in the nuclear migrations of the two dorsal rows of epidermal cells. In wild-type embryos the two dorsal rows intercalate to form a single row. The nuclei of the cells in the right dorsal row migrate to the left, while the nuclei of the cells in the left dorsal row migrate to the right, eventually switching sides (Fig. 1A). In animals depleted of the WAVE/SCAR complex component GEX-2, the Sra-1/PIR121/CYFIP homolog, the two dorsal rows failed to intercalate, and the nuclear migrations also failed to occur, with the nuclei remaining at their original positions at the center of each cell (Fig. 1A; Movie 1). Similar defects were seen in other Gex mutants (data not shown). These defects suggested that Arp2/3-dependent actin nucleation may have a role in nuclear migrations.
Fig. 1. Nuclear migrations in the embryonic epidermis and larval neuronal precursors require the WAVE/SCAR and Arp2/3 complexes.
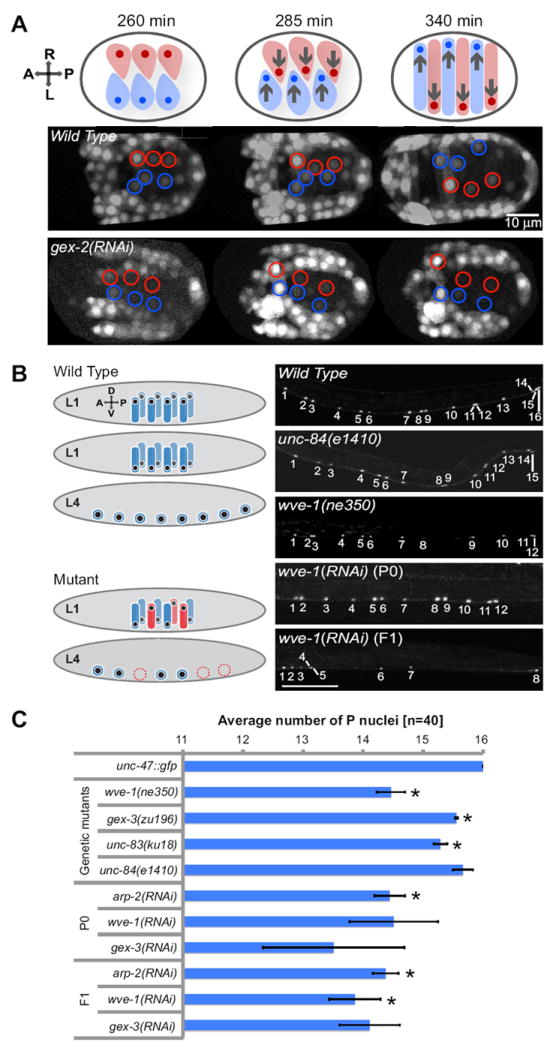
(A) During dorsal intercalation two dorsal rows of epidermal cells (hyp7) migrate and intercalate across the left-right body axis. Nuclei within the epidermal cells migrate in the direction of the cell migration and eventually to the opposite side (arrows). Fluorescent time-lapse images show nhr-25∷yfp in equally staged wild-type embryos undergoing morphogenesis and in embryos depleted via RNAi of gex-2, a component of the WAVE Complex (Patel et al., 2008). Embryos are oriented with anterior to the left and the dorsal side up. Epidermal cells and nuclei of the right and the left side of the body axis are highlighted in red and blue, respectively. Timing of developmental events after the first cell division is indicated in minutes. (B) During the first larval stage (L1) two rows of neuronal precursor cells (P cells) and their nuclei (P nuclei) migrate ventrally from the left and right side of the larval body to differentiate into P cell-derived motor neurons in the ventral nerve cord (wild type). The post-embryonic P cell differentiation is completed at the last larval stage (L4). Failure in migration of P nuclei often leads to elimination of the affected P cell (mutant) and reduced number of P cell-derived motor neurons in the ventral nerve cord. Wild type P nuclear migration is highlighted in blue. Mutant P nuclear migration and loss of P cell-derived motor neurons are shown in red. Right panels. Wild-type worms expressing the unc-47∷gfp marker at larval stage L4 exhibit 16 P nuclei in the posterior ventral nerve cord. The unc-84(e1410) and wve-1(ne350) mutations led to reduced number of nuclei in the ventral nerve cord. Depletion of wve-1 via RNAi resulted in loss of nuclei in the ventral nerve cord, observed both in the P0 generation and in the RNAi escapers, which developed through the F1 generation. Size bar represents 50μm. Worms were maintained at 25°C. (C) Average number of motor neuron nuclei in the L4 ventral nerve cord, detected with unc-47∷gfp transgene, of either wild type or animals missing WAVE/SCAR complex, SUN or KASH domain proteins. For each genotype 120 animals were analyzed. Error bars show standard error of the mean (SEM), which was calculated from the mean of three independent experiments for each strain. Asterisks mark statistical significance, p<0.05. See also Movie 1.
Nuclear migrations in neuronal precursors require the WAVE/SCAR and Arp2/3 complexes
Genetic screens in C. elegans have identified proteins specifically required for nuclear migrations (Hedgecock and Thomson, 1982; Horvitz and Sulston, 1980; Lee et al., 2002; Malone et al., 1999; McGee et al., 2006; Starr and Fischer, 2005; Starr and Fridolfsson, 2010; Starr and Han, 2002; Starr and Han, 2003; Starr et al., 2001). To test if the Arp2/3 defects were specific to the dorsal epidermis, or indicated a general requirement for nuclear migrations, we compared the Gex mutants to known nuclear migration genes focusing on another well characterized nuclear migration, that of neuronal precursors. In wild-type animals the P neuroblasts are born on the lateral surface of larvae and during the first larval stage the cells extend a protrusion towards the ventral cord that is followed by a nuclear migration. If cell migration fails, the P cells remain on the lateral side (Spencer et al., 2001). However, if the nuclear migration fails, the P cells often will die (Sulston and Horvitz, 1981) by a programmed-cell-death-independent mechanism (Malone et al., 1999) (Fig. 1B). This cell differentiation therefore requires a nuclear migration. P nuclei migration is temperature sensitive when nuclear migration proteins are compromised, with stronger defects at 25°C. The e1410 null allele of UNC-84, a known regulator of nuclear migrations, resulted in a 90% drop in the number of P nuclei descendants when total ventral cord neurons were counted at 25°C (Malone et al., 1999).
Defects in nuclear migration of P neuroblasts can be detected by monitoring the number of GABAergic neurons that differentiate at the ventral cord using the oxIs12 (unc-47∷gfp) transgenic strain (McIntire et al., 1997). In this line 19 nuclei express GFP in the ventral cord, and 13 of these are from P cell lineages (Starr et al., 2001). We focused on 16 posterior nuclei that are easiest to count, culturing the worms at 25°C (Fig. 1B). Using the null UNC-84/SUN allele, unc-84(e1410), we detected on average that 0.5/16 P neuroblast daughters were missing (a 3% drop). When we removed WAVE/SCAR or Arp2/3 components either by genetic mutations or RNAi, we noticed a stronger defect. Homozygous null mutations in wve-1, including ne350, are viable due to maternal rescue (Patel et al., 2008). Homozygous wve-1(ne350) animals had an average of 1.5/16 missing P nuclei (9%), while wild-type L1 larvae fed RNAi food to deplete wve-1, gex-3 or arp-2 also had an average of 1.5/16 missing P nuclei (9%) (Fig. 1B and C). This RNAi effect is remarkable because the migrations we are monitoring occur during the mid-L1 stage, so the RNAi has to take effect within 6 hours for this defect to be seen. To further monitor the outcome of depleting WAVE/SCAR or Arp2/3 components, we followed the progeny of these larvae. As expected, most of the progeny were dead embryos with the Gex (gut on the exterior) phenotype, precluding us from examining the nuclear migrations of their P nuclei. However, RNAi is not fully effective, especially against wve-1 (Patel et al., 2008), allowing us to monitor the P nuclei migrations in the RNAi escapers in the second generation (F1). These animals were missing an average of 1.5 to 2/16 (9-12.5%) of their P nuclei. Notably, some animals had a striking loss of up to 8/16 (50%) of their P nuclei (Fig. 1 B and C).
It is possible that branched actin could be contributing to P cell migrations, as opposed to P nuclei migrations. However, mutations that affect P cell migration, rather than P nuclear migration lead to P descendants that stay on the lateral side of the animal, and do not die (Spencer et al., 2001). The fact that there were no ectopic lateral P descendants in animals missing WAVE/SCAR or Arp2/3 components (n=600), but there were missing P cell descendents suggested that the P cell defect was due to inappropriate nuclear migration and cell death, not inappropriate cell migration.
Male pronuclear migrations in the one-cell embryo require the WAVE/SCAR and Arp2/3 complexes
Analysis of nuclear migrations in the dorsal epidermis and in the P nuclei is complicated by the fact that the nuclear migration is coupled to a cell migration. Therefore, the failure in nuclear migration may be a secondary consequence of the failure in cell migration. To focus more specifically on the behavior of the nucleus during nuclear migration, we looked for a simpler instance of an Arp2/3-dependent nuclear migration. Gex mutants do not display obvious developmental delays or defects during early embryonic development. Our previous analyses suggested that lethality of Gex embryos is caused by failures in epidermal cell movements at the 500-cell stage of embryonic development (Patel et al., 2008; Soto et al., 2002). We were therefore surprised to discover that the male pronuclear migration of the one-cell zygote is affected by loss of WAVE/SCAR and Arp2/3 components. In wild-type embryos the male pronucleus becomes visible via DIC microscopy shortly after fertilization. Regardless of the exact location of sperm entry, the male pronucleus moves to one pole and rests on the cortex. The pole to which the male pronucleus migrates becomes the posterior pole of the embryo, since the sperm-associated centrosomes establish posterior polarity in C. elegans (Goldstein and Hird, 1996). The male pronucleus then begins a migration towards the anterior pole to meet the female pronucleus. The female pronucleus, which is typically found at the opposite pole, simultaneously migrates toward the posterior pole to meet the male pronucleus. In Gex mutants the female pronuclear migration still occurs and begins at the opposite pole relative to the male pronucleus (Figure 2B, Movie 2). However, the male pronucleus shows a specific change: it migrates to the pole opposite to the female pronucleus, and associates with the cortex, but then does not migrate at all (in 21-29% of the Gex mutants), or undergoes a minimal migration away from the posterior pole (in an additional 33-73% of the Gex mutants) (Fig. 2; Table 1, Movie 2). Mutations in some proteins, including dynein, the minus-end directed microtubule motor, affect migrations of both pronuclei (Gonczy et al., 1999; Hamill et al., 2002; Malone et al., 2003; O’Connell et al., 2000; Schmidt et al., 2005).
Fig. 2. Migration of the male pronucleus in the one-cell embryo requires WAVE/SCAR and Arp2/3 components and the SUN protein UNC-84.
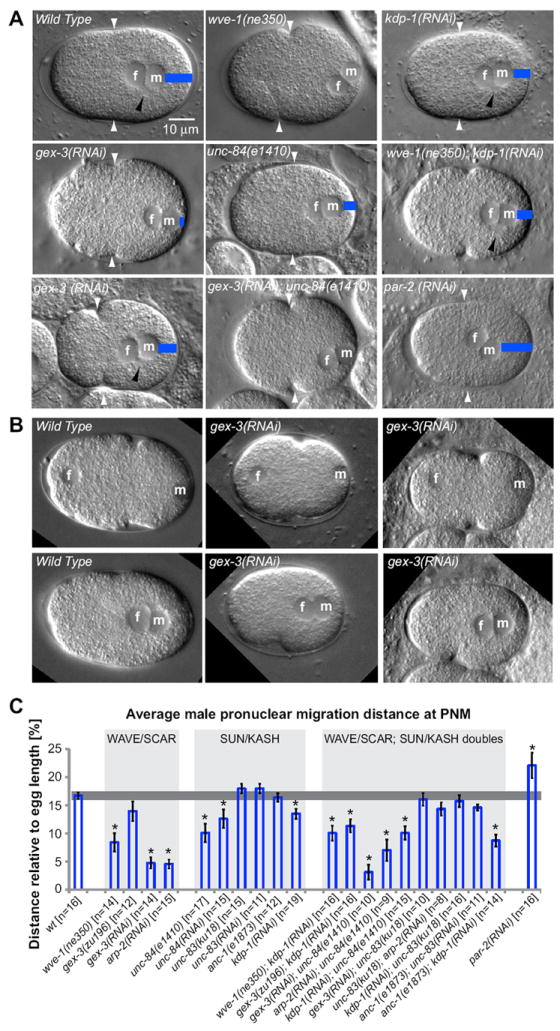
(A) DIC images of one-cell embryos are shown at the stage of pronuclear meeting (PNM). In all images the anterior pole is aligned on the left. The female and male pronuclei are marked by f and m, respectively. Blue bars indicate the distance of male pronuclear migration away from the posterior pole. White arrowheads point to the site of pseudo-cleavage formation. Black arrowheads point to the centrosomal region, visible by DIC in some focal planes as a clear circular region attached to the nucleus. Two representative gex-3(RNAi) embryos show the variation in embryo length and male pronuclear migration distance in Gex mutants. (B) Still images taken from Movie 2 show one wild-type and two gex-3(RNAi) embryos at -345 seconds before PNM (top panels) and at PNM (0 seconds, bottom panels). The female and male pronuclei are marked by f and m. (C) Average migration distance of the male pronucleus was examined in the WAVE/SCAR and SUN and KASH mutants as well as in the double mutants. Male pronuclear migration distance was determined relative to the total length of the one-cell embryo at PNM. Embryo length and male pronuclear migration distance were measured from images captured at PNM, the time when the two pronuclei meet and their adjacent membranes form a straight line. Only embryos where the positions of the male and female pronucleus were observed prior to PNM were included in the analysis. Double mutants were made by reducing one gene via RNAi in animals containing a mutation in the second gene, and monitoring RNAi in control animals. For example, unc-83; anc-1 doubles were made by depleting unc-83 via RNAi in anc-1(e1873) animals and monitoring RNAi effectiveness in unc-83(RNAi) single mutants by the hyp7 nuclear migration defect. Error bars indicate SEM; asterisks mark statistical significance, p<0.05. See also Table 1 and Movie 2. All movies and images of one-cell embryos were made at 22°C, except for those of zyg-12(or577ts), which needs to be viewed at 25°C (Malone et al., 2003).
Table 1.
Male pronuclear migration distance at pronuclear meetinga
| Genotype | Rangeb [% egg length] | Number of embryos (% of n)
|
|||
|---|---|---|---|---|---|
| Decreased [<14%] | wt [14-20%] | Increased [>20%] | |||
| wt [n=16] | 14-20 | 0 | 16 (100%) | 0 | |
|
| |||||
| WAVE/SCAR | wve-1(ne350) [n=14] | 0-16 | 10 (71%) | 4 (29%) | 0 |
| gex-3(zu196) [n=12] | 0-21 | 5 (42%) | 7 (58%) | 0 | |
| gex-3(RNAi) [n=14] | 0-9 | 14 (100%) | 0 | 0 | |
| arp-2(RNAi) [n=15] | 0-7 | 15 (100%) | 0 | 0 | |
|
| |||||
| SUNKASH | unc-84(e1410) [n=17] | 0-19 | 10 (59%) | 7 (41%) | 0 |
| unc-84(RNAi) [n=15] | 0-23 | 8 (53%) | 7 (47%) | 0 | |
| unc-83(ku18) [n=15] | 10-22 | 2 (13%) | 11 (73%) | 2 (13%) | |
| unc-83(RNAi) [n=11] | 13-21 | 1 (9%) | 8 (73%) | 2 (18%) | |
| anc-1(e1873) [n=12] | 13-22 | 2 (17%) | 9 (75%) | 1 (8%) | |
| kdp-1(RNAi) [n=17] | 7-24 | 7 (41%) | 9 (53%) | 1 (6%) | |
|
| |||||
| Double mutants | wve-1(ne350); kdp-1(RNAi) [n=16] | 1-18 | 12 (75%) | 4 (25%) | 0 |
| gex-3(zu196); kdp-1(RNAi) [n=16] | 1-17 | 10 (62%) | 6 (38%) | 0 | |
| gex-3(RNAi); unc-84(e1410) [n=10] | 0-9 | 10 (100%) | 0 | 0 | |
| arp-2(RNAi); unc-84(e1410) [n=9] | 0-19 | 8 (89%) | 1 (11%) | 0 | |
| kdp-1(RNAi); unc-84(e1410) [n=15] | 3-17 | 12 (80%) | 3 (20%) | 0 | |
| gex-3(RNAi); unc-83(ku18) [n=10] | 9-21 | 2 (20%) | 8 (80%) | 0 | |
| unc-83(ku18); arp-2(RNAi) [n=8] | 8-18 | 4 (50%) | 4 (50%) | 0 | |
| kdp-1(RNAi); unc-83(ku18) [n=16] | 4-23 | 5 (31%) | 10 (63%) | 1 (6%) | |
| anc-1(e1873); unc-83(RNAi) [n=11] | 12-18 | 3 (27%) | 8 (73%) | 0 | |
| anc-1(e1873); kdp-1(RNAi) [n=14] | 4-16 | 12 (86%) | 2 (14%) | 0 | |
|
| |||||
| par-2(RNAi) [n=16] | 0-33 | 2 (12%) | 2 (13%) | 12 (75%) | |
Male pronuclear migration distance indicates distance between posterior plasma membrane and posterior membrane of the male pronucleus relative to total egg length [%]
Range reflects male pronuclear migration distance relative to total egg length [%] for each genotype
The Gex male pronuclear migration defects in the one-cell zygote are not fully penetrant (Fig. 2C, Table 1), and they do not interfere with the overall asymmetry of the first cell division (Fig. 4; Movie 3). The existence of parallel pathways to ensure the robustness of polarity in early embryos is frequently observed (Lu and Mains, 2005; Page et al., 2007; Rose and Kemphues, 1998; Walston et al., 2004). Therefore, this early role for the GEX proteins likely works in parallel with other mechanisms to ensure proper pronuclear migrations and polarization during the one-cell stage.
Fig. 4. WAVE/SCAR and Arp2/3 components are required for posterior PAR cortical accumulation.
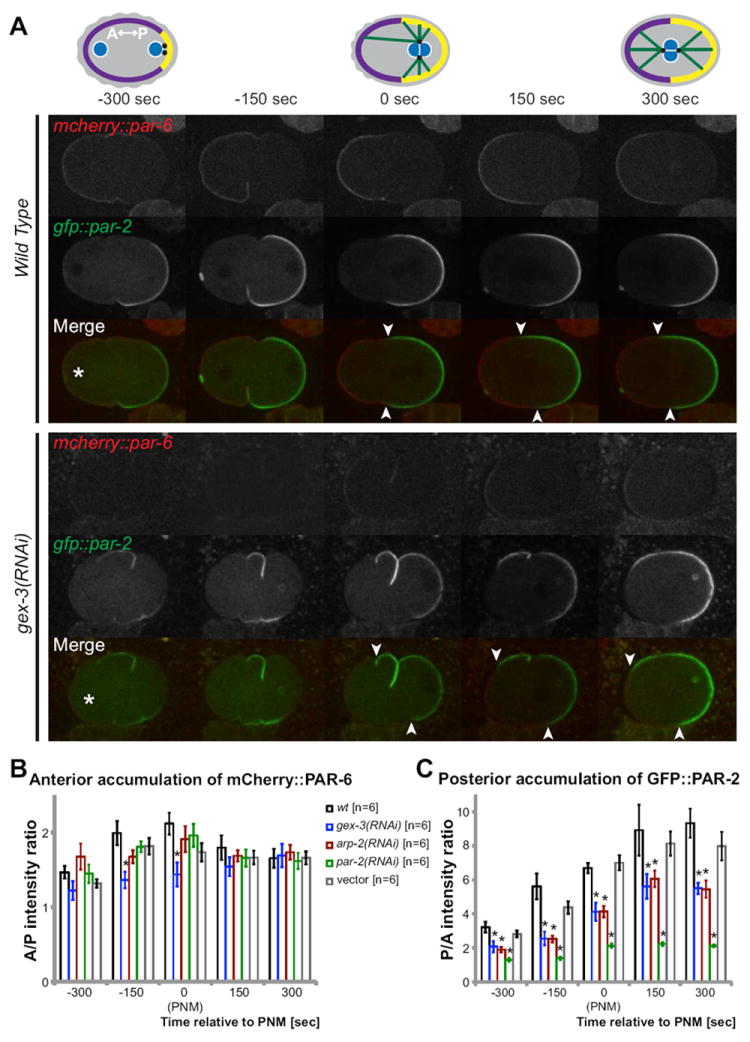
(A) Cartoon depicting the wild type cortical accumulation of PAR-6 (purple) at the anterior and PAR-2 (yellow) at the posterior half of the one-cell embryo. Pronuclei are marked blue, centrosomes black and microtubules green. Pronuclear meeting (PNM) is set as time 0 sec. Live imaging of mCherry∷PAR-6 (red) and GFP∷PAR-2 (green) in wild type and in embryos depleted of gex-3 via RNAi are shown at the indicated time points using the TH120 transgene (Schonegg et al., 2007). The initial position of the male and female pronucleus was recorded. Asterisks mark the location of the female pronucleus 300 seconds before pronuclear meeting. (B) Average fluorescence intensity ratio of mCherry∷PAR-6 at the anterior membrane relative to the posterior membrane was measured in live one-cell embryos. For each embryo three measurements were made at the anterior membrane and the posterior membrane using the line tool function of ImageJ. See Methods for details. The average fluorescence intensity ratio between the anterior and posterior membrane (A/P) was plotted. C) Average ratio of fluorescence intensity of GFP∷PAR-2 at the posterior membrane and in the anterior membrane (P/A) in live one-cell embryos, calculated as for mCherry∷PAR-6. Error bars represent SEM; asterisks mark statistical significance, p<0.05.
Nuclear envelope proteins, including UNC-84/SUN, influence the migration of the male pronucleus
Since Gex mutants share P nuclei migration defects with known nuclear migration mutants, we looked at the male pronuclear migrations in those mutants. C. elegans has two SUN proteins, UNC-84 and SUN-1. SUN-1 is required in the one-cell embryo to link the centrosome to the nuclear envelope through its KASH and HOOK protein partner, ZYG-12 (Malone et al., 2003). Loss of sun-1 or zyg-12 resulted in detached centrosomes and pronuclei that often failed to meet at all before the first cell division began. In contrast, loss of WAVE/SCAR or Arp2/3 components did not result in detached centrosomes (n=100+). Therefore the severity of the centrosome attachment defects in embryos missing SUN-1 or its KASH partner ZYG-12 prevented us from making a direct comparison to Gex defects.
Unlike mutations in sun-1, mutations in unc-84, the second SUN domain protein, showed defects in male pronuclear migration similar to those in WAVE/SCAR or Arp2/3 mutants. UNC-84 has two KASH domain partners, UNC-83 and ANC-1. UNC-83/KASH connects the nucleus to the microtubule cytoskeleton via direct interactions with kinesin-1 (Meyerzon et al., 2009) and two dynein-regulating complexes (Fridolfsson et al., 2009; Fridolfsson and Starr, 2010). ANC-1 instead connects the nuclear envelope to the actin cytoskeleton through a N-terminal actin binding domain (Starr and Han, 2002). However, neither unc-84, unc-83, nor anc-1 mutants have been reported to have early embryonic phenotypes. UNC-84 has not been detected in early embryos using transgenes or antibodies (Lee et al., 2002; Malone et al., 1999). To our surprise, loss of UNC-84 via mutation or RNAi resulted in approximately 50% of embryos with decreased male pronuclear migration at pronuclear meeting (n=32), a defect almost as penetrant and severe as loss of WAVE/SCAR or Arp2/3 components (Fig. 2, Table 1). To identify the KASH partner of UNC-84 that supports the role in male pronuclear migration, we removed UNC-83 and ANC-1. The strong loss of function mutation in anc-1, (e1873) (Starr and Han, 2002) resulted in mild male pronuclear migration defects, as did loss of UNC-83 through mutations or RNAi (Table 1). The anc-1(e1873); unc-83(RNAi) double mutants did not display a significant increase in pronuclear migration defects (Table 1). A third KASH protein, KDP-1, was identified in a yeast two-hybrid screen for UNC-84 partners, but is thought to interact in vivo with SUN-1 (McGee et al., 2009). Mutations in kdp-1 often cause severe germline and cell cycle defects (McGee et al., 2009) never seen in unc-84 or arp-2 mutations (our unpublished data). Depletion of kdp-1 via RNAi resulted in 41% (n=17) of the embryos with decreased male pronuclear migration, almost as strong as in unc-84 mutants. kdp-1; anc-1 double mutants (86%, n=14) and kdp-1; unc-84 double mutants (80%, n=15) resulted in increased defects in male pronuclear migration, stronger than in the single mutants. These results suggest that in addition to the roles previously described for KDP-1 in germline development (McGee et al., 2009), that in the one-cell embryo KDP-1 could be a KASH partner of UNC-84 required for male pronuclear migration, possibly in combination with ANC-1. Alternatively, KDP-1 may act in parallel to UNC-84 and ANC-1 during male pronuclear migration.
WAVE/SCAR and Arp2/3 regulate the polarized distribution of actomyosin in the zygote
To better understand how removing the WAVE/SCAR complex or Arp2/3 affects cytoskeletal dynamics in the one-cell embryo, we made movies with transgenes that illustrate the dynamics of the actin and microtubule cytoskeletons. The movies were acquired at 22°C and aligned temporally relative to pronuclear meeting (PNM, 0 seconds). The nmy-2∷gfp transgene illustrates non-muscle myosin distribution and movements in the early embryo (Munro et al., 2004). Dynamic NMY-2∷GFP puncta initially form all over the oocyte cortex. Upon fertilization and the establishment of the posterior pole, NMY-2∷GFP moves away from the posterior pole, setting up an anterior cap rich in actomyosin contractility. In wild-type embryos the movement of NMY-2∷GFP away from the posterior pole was rapid and highly dynamic, with most of the NMY-2∷GFP disappearing from the posterior pole by 150 seconds before pronuclear meeting and becoming highly enriched at the anterior-most cortex by 150 seconds after pronuclear meeting (Fig. 3). Upon depletion of WAVE/SCAR or Arp2/3 components, NMY-2∷GFP still moved away from the posterior pole, but the dynamics were altered. It took an additional 300 seconds for NMY-2∷GFP to disappear from the posterior half of the embryo (Fig. 3B). Once the anterior cap of NMY-2∷GFP is established, a further event illustrated by NMY-2∷GFP is the disassembly of large actomyosin foci at the cortex (Munro et al., 2004). In embryos depleted of WAVE/SCAR or Arp2/3 components, the formation and dissolution of puncta was altered (Fig. 3C). For example, we counted an average of 58 large puncta on one surface of wild-type embryos at 300 seconds before pronuclear meeting (n=6 for all genotypes and time points). gex-3 mutant embryos at the same time point had an average of just 30 large puncta on one surface. At 150 seconds wild-type embryos had an average of one punctum, while Gex embryos had an average of 20 puncta. These results suggest slower overall dynamics of both actomyosin contractility and turnover that result in slower clearing of NMY-2∷GFP from the posterior half of the embryo and slower dissolution of large foci. unc-84 mutant embryos did not display these changes in NMY-2∷GFP contractility (0/6 movies; data not shown). Embryos depleted of WAVE/SCAR and Arp2/3 still went on to divide at approximately the correct time (Fig. 5B, Movie 3). Therefore we analyzed when actomyosin contractility finally reaches the anterior of the cortex. By 450 seconds after pronuclear meeting, actomyosin contractility reached the anterior cortex in all of the WAVE/SCAR and Arp2/3 mutant strains, compared to 150 seconds after pronuclear meeting in wild type (Fig. 3B). Therefore actomyosin dynamics are delayed, but not enough to interfere with the first cell division (Fig. 3; Movie 3) or subsequent cell divisions that we examined (data not shown). The change in NMY-2∷GFP dynamics is penetrant, visible in 100% of the NMY-2∷GFP movies made with Gex mutants (n= 12).
Fig. 3. WAVE/SCAR and Arp2/3 regulate the polarized distribution of actomyosin in the zygote.
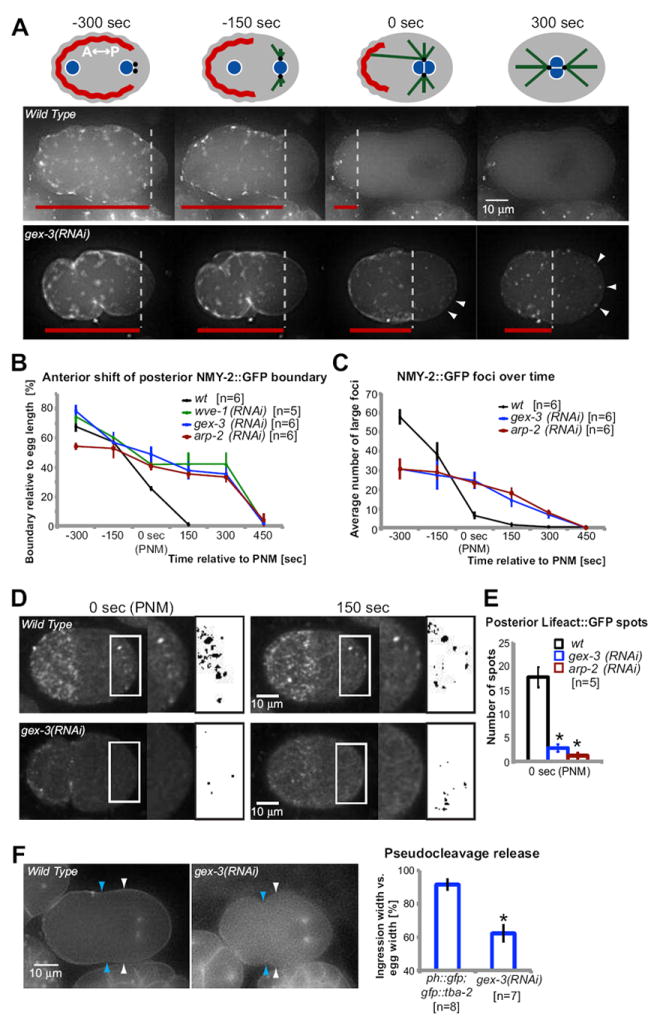
(A) Schematic illustration of cortical NMY-2∷GFP migration towards the anterior pole of one-cell embryos. Meeting of female and male pronuclei is defined as time 0, timing of other developmental stages are shown in seconds relative to time of pronuclear meeting (PNM). NMY-2∷GFP is marked red, pronuclei blue, centrosomes black and microtubules green. Time-lapse imaging of NMY-2∷GFP in one-cell embryos. The posterior boundary of cortical NMY-2∷GFP contractile filaments is labeled with dotted lines. Red lines indicate the distance of the posterior NMY-2∷GFP boundary relative to the anterior pole. Arrowheads mark posterior NMY-2∷GFP foci. (B) Average distance of the posterior boundary of NMY-2∷GFP filaments relative to the total length of the embryo, over time, in wild type and WAVE/SCAR mutants. (C) Abundance of cortical actomyosin contraction was measured as the average number of large NMY-2∷GFP foci found at the indicated time points in wild type and WAVE/SCAR mutants. Number of NMY-2∷GFP foci was determined using the ImageJ plug-in ‘Analyze Particles’. (D) Spinning disc confocal time-lapse imaging of Lifeact∷GFP in one-cell embryos. White boxes highlight the posterior region in which the number of actin foci was counted after converting that region into a binary image (black boxes). See Methods for details. (E) Average number of posterior actin foci in wild type and WAVE/SCAR mutants at the indicated time points. (F) Degree of pseudo-cleavage release at pronuclear meeting was investigated using GFP∷PH [OD73 pie-1p∷gfp∷ph(plc1δ1)]. The absolute width (white arrowheads) and the width of pseudocleavage ingression (blue arrowheads) were measured at the stage of pronuclear meeting in GFP∷PH and in GFP∷PH embryos depleted of gex-3 via RNAi. The average pseudo-cleavage width relative to the average width of the one-cell embryo is graphed on right. In all panels error bars represent SEM; timing (sec) are relative to pronuclear meeting (PNM); asterisks mark statistical significance, p<0.05.
Fig. 5. Events in the zygote thought to depend on MT-cortex interactions are delayed by the loss of Arp2/3 and SUN proteins.
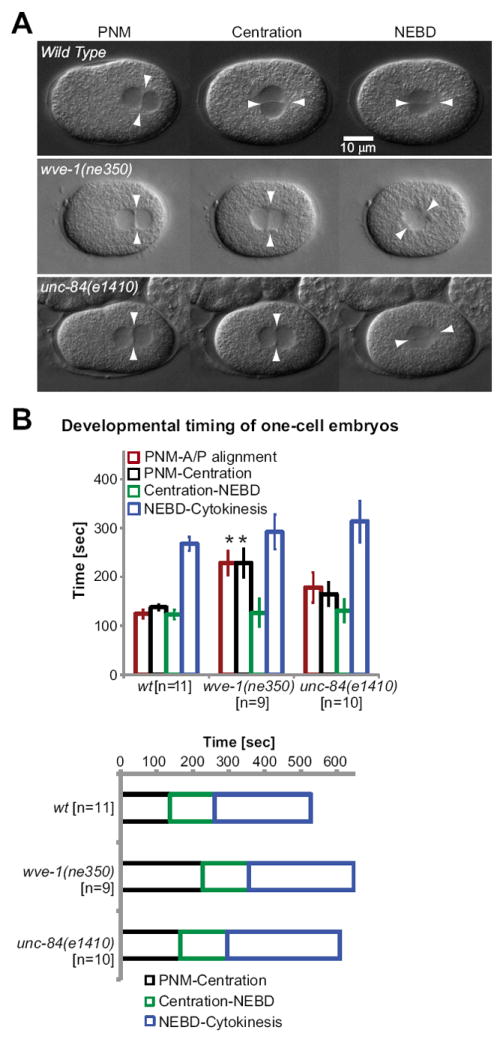
(A) In wild-type embryos at pronuclear meeting (PNM), the midplane, formed by the female and male pronuclei, is aligned perpendicular to the anteroposterior axis of the one-cell embryo (arrowheads). At pronuclear centration, pronuclei rotated and aligned along the anteroposterior axis in wild-type embryos. In contrast, pronuclear rotation was not observed during pronuclear centration in gex-3(zu196) or unc-84(e1410) embryos. Instead, the rotation leading to anteroposterior alignment of pronuclei occurred simultaneously with nuclear envelope breakdown (NEBD). Arrowheads mark the alignment of pronuclear midplane relative to the anteroposterior axis. See also Movie 3. (B) Comparison of developmental timing of wild-type, wve-1(ne350) and unc-84(e1410) one-cell embryos. Average time lapse between the developmental events are indicated in seconds. The timing of the following developmental events were determined at 22°C: from pronuclear meeting (PNM) to nuclear-centrosomal rotation onto the anterior/posterior axis (NC Rotation), from PNM to pronuclear centration, from centration to nuclear envelope breakdown (NEBD) and from NEBD to completion of cytokinesis of the one-cell embryo.
In wild-type embryos a constriction of the cortex termed pseudocleavage begins approximately 200 seconds before pronuclear meeting. This constriction, which is thought to result from differences in actomyosin contractility between the anterior and posterior of the embryo (Cowan and Hyman, 2004b), normally recedes by the time of pronuclear meeting. In Gex mutant embryos pseudocleavage lasted longer, still being visible 150 seconds beyond when it recedes in wild type. To better measure the extent of the pseudocleavage furrow ingression, we used a strain carrying the gfp∷plc-δ-ph transgene (GFP∷PH), which expresses a protein enriched at the cell membrane (Audhya et al., 2005; Stauffer et al., 1998; Varnai and Balla, 1998). Using this strain to measure the pseudocleavage ingression, we found that in wild-type embryos at pronuclear meeting the ingression constricted the embryo by approximately 10% of the embryo width, while in the mutants at pronuclear meeting the ingression constricted the embryo by as much as 50% of embryo width (Fig. 3F). Together with the changed number of large NMY-2∷GFP puncta, and the slower dynamics of anterior movement of NMY-2∷GFP to the anterior, these results suggest that Arp2/3-dependent actin nucleation has a role in promoting turnover of NMY-2∷GFP puncta that normally limits the extent of pseudocleavage ingression.
WAVE/SCAR and Arp2/3 regulate F-actin in the zygote
To directly monitor the effects of WAVE/SCAR or Arp2/3 depletion on F-actin in the zygote, we used a lifeact∷gfp transgene expressed under the early embryonic pie-1 promoter (Pohl and Bao, 2010). This transgene highlights a population of F-actin that is enriched at the cellular cortex in the one cell embryo, and that becomes enriched at the anterior cortex as actomyosin contractility remodels the cortex (Fig. 3D). The posterior of the embryo contains a small population of actomyosin puncta that are detected when nmy-2∷gfp is imaged with spinning disc confocal microscopy (Goulding et al., 2007; Munro et al., 2004). Using spinning disc microscopy on lifeact∷gfp we detected transient actin-rich puncta at the posterior cortex of wild-type embryos, most visible between pronuclear meeting (0 seconds) and 150 seconds after. Removal of arp-2 or gex-3 resulted in an overall drop in the level of Lifeact∷GFP, and a change and reduction in the posterior puncta at pronuclear meeting (Fig. 3D and E). This result suggests that Arp2/3 is required for the overall abundance of F-actin that is recognized by Lifeact∷GFP, including a small population of posterior actin present in the one-cell embryo.
Loss of WAVE/SCAR or Arp2/3 components does not affect PAR-6 distribution
The movement of NMY-2 to the anterior pole recruits the anterior PAR proteins to the anterior pole (Munro et al., 2004). Therefore changes in NMY-2 and actin dynamics could alter the establishment of anterior/posterior polarity through the PARs. We examined the accumulation of the anterior PAR protein PAR-6 by use of two transgenes, par-6∷gfp and mCherry∷par-6 (Cuenca et al., 2003; Schonegg et al., 2007). In wild type, mCherry∷PAR-6 was found all over the cortex of the unfertilized, unpolarized oocyte. Upon recruitment by NMY-2, PAR-6 becomes enriched at the cortex in the anterior half of the embryo and disappears from the posterior half by the time of pronuclear meeting. In embryos depleted of WAVE/SCAR or of Arp2/3 components we were unable to detect changes in the distribution of PAR-6: there was no measurable change in the cortical depletion of mCherry∷PAR-6 from the posterior cortex, and mCherry∷PAR-6 was asymmetrically enriched at the anterior cortex by 150 seconds before pronuclear meeting (Fig. 4A and B). Similar results were obtained with the par-6∷gfp transgene (data not shown).
WAVE/SCAR and Arp2/3 are required for cortical accumulation of PAR-2 at the posterior of the zygote
It is necessary to remove the anterior PAR proteins from the posterior so that posterior PARs can accumulate at the posterior cortex as the anterior and posterior PAR complexes help set up and maintain each other s boundaries (Cuenca et al., 2003; Kemphues, 2000). We therefore examined if the loss of WAVE/SCAR and Arp2/3 affected the posterior PARs, by analyzing two different gfp∷par-2 transgenes (Cuenca et al., 2003; Schonegg et al., 2007). In wild-type embryos GFP∷PAR-2 accumulated on the cortex of the posterior half of the embryo by the time of pronuclear meeting. By 150 seconds after pronuclear meeting GFP∷PAR-2 was highly enriched at the posterior cortex, and reduced in the anterior cytoplasm. In contrast, loss of WAVE/SCAR or Arp2/3 components decreased GFP∷PAR-2 accumulation at the posterior cortex. The earliest we could detect GFP∷PAR-2 enrichment at the posterior cortex in mutants was at 300 sec before pronuclear meeting, at levels below those seen in wild type (Fig. 4A). This defect in GFP∷PAR-2 accumulation was fully penetrant, affecting 100% of gex-3 and arp-2 embryos examined (n=12). Similar results were obtained with a second par-2∷gfp transgene (data not shown). For comparison, depletion of par-2 via RNAi more severely reduced GFP∷PAR-2 accumulation at the cortex (Fig. 4C). This suggests that there is a reduction but not complete loss of PAR-2 at the posterior cortex in embryos depleted of Arp2/3. The boundary between anterior and posterior PAR proteins is usually similar all around the transverse circumference of the embryo (white arrow heads in Fig. 4A). Loss of gex-3 or arp-2 via RNAi led to more skewed distribution of the anterior and posterior PARs, affecting 4/6 arp-2 embryos and 3/6 gex-3 embryos. This result further suggests that Arp2/3 regulates cortical polarity.
Events in the zygote thought to depend on MT-cortex interactions are delayed by the loss of Arp2/3 and SUN proteins
The movies of early zygotes revealed that mutations in WAVE/SCAR, Arp2/3, or UNC-84 lead to other defects that are thought to be related to movements of the nucleus by the microtubules. In wild-type animals, the male and female pronuclei meet at the posterior, then travel together to the center of the one-cell embryo, in a process known as centration. Simultaneously, the pronuclei undergo a nuclear-centrosomal rotation that permits the first cell division to occur along the anterior/posterior axis (White and Strome, 1996). Wild-type centration and rotation are thought to depend on astral microtubules (Cowan and Hyman, 2004a) through undescribed connections to the cortex, and on the anterior enrichment of NMY-2 by cortical Gα factors GOA-1/GPA-16 (Goulding et al., 2007). In animals depleted of WAVE/SCAR, Arp2/3, or UNC-84, just as in wild type, the two pronuclei met and then migrated together to the center of the cell, and the first cell division occurred along the A/P axis and was asymmetrical. However, in these movies, made at 22°C, we noted slight delays in the first cell cycle due to delays in centration and in the nuclear-centrosomal rotation (Fig. 5; Movie 3). Wild type embryos complete centration by 140 seconds after pronuclear meeting, while in wve-1 and unc-84 embryos centration occurs, on average, 90 and 48 seconds later, respectively (Fig. 5B). In wild-type embryos nuclear-centrosomal rotation began as soon as the two pronuclei met, and was completed by 150 seconds after pronuclear meeting, by the time centration had occurred (Fig. 5; Movie 3). In wve-1 and unc-84 embryos nuclear-centrosomal rotation was delayed by variable amounts, with an average delay of 84 and 55 seconds, respectively, so that the nuclear-centrosomal rotation often occurred after centration (Fig. 5A). Both centration and nuclear-centrosomal rotation are thought to depend on the connection of astral microtubules to proteins at the actin-rich anterior cortex (Hyman, 1989; White and Strome, 1996). Therefore the delayed centration and nuclear-centrosomal rotation suggested that there is a general defect in the link between the microtubule and actin cytoskeletons.
Loss of WAVE/SCAR or Arp2/3 components reduces centrosomal MT assembly
One important posterior event occurring at the time of PAR-2 posterior accumulation is the maturation of the centrosomes that are brought in by the sperm and that associate with the male pronucleus (Cowan and Hyman, 2004b; Galli and van den Heuvel, 2008; Goldstein and Hird, 1996; O’Connell et al., 2000; Sadler and Shakes, 2000; Wallenfang and Seydoux, 2000). We used an alpha-tubulin transgene, gfp∷ tba-2 (Audhya et al., 2005) to view tubulin accumulation in one-cell embryos in the presence or absence of WAVE/SCAR or Arp2/3 components. Live imaging of embryos expressing GFP∷TBA-2 showed that by the time of pronuclear meeting, there was significant accumulation of α-tubulin at the centrosomes. Tubulin intensity at the centrosomes peaked at the time of bipolar spindle assembly (300 sec after pronuclear meeting), right before the first cell division (Fig. 6). Embryos depleted of gex-3 or arp-2 via RNAi showed decreased centrosomal accumulation of GFP∷TBA-2 at all stages. For example, the peak intensity still occurred at 300 seconds, but the level was decreased by 17% and 33% in gex-3 and arp-2 RNAi animals, respectively. This defect affected 100% of the Gex embryos (n=11) (Fig. 6A). These results indicated that branched actin is needed to support centrosomal MT growth during the first cell cycle. To determine if the loss of tubulin accumulation reflects a general need for branched actin, or a requirement for branched actin only during the first cell cycle we measured centrosomal MT growth, as monitored by GFP∷TBA-2, in these same embryos, during the second cell division. We did not find a defect in GFP∷TBA-2 accumulation at the centrosomes at the second cell division in arp-2 mutant embryos (n=6), which suggested that there is a specialized requirement for branched actin to help centrosomal MT growth during the first cell cycle.
Fig. 6. Loss of WAVE/SCAR or Arp2/3 components reduces centrosomal MT assembly.
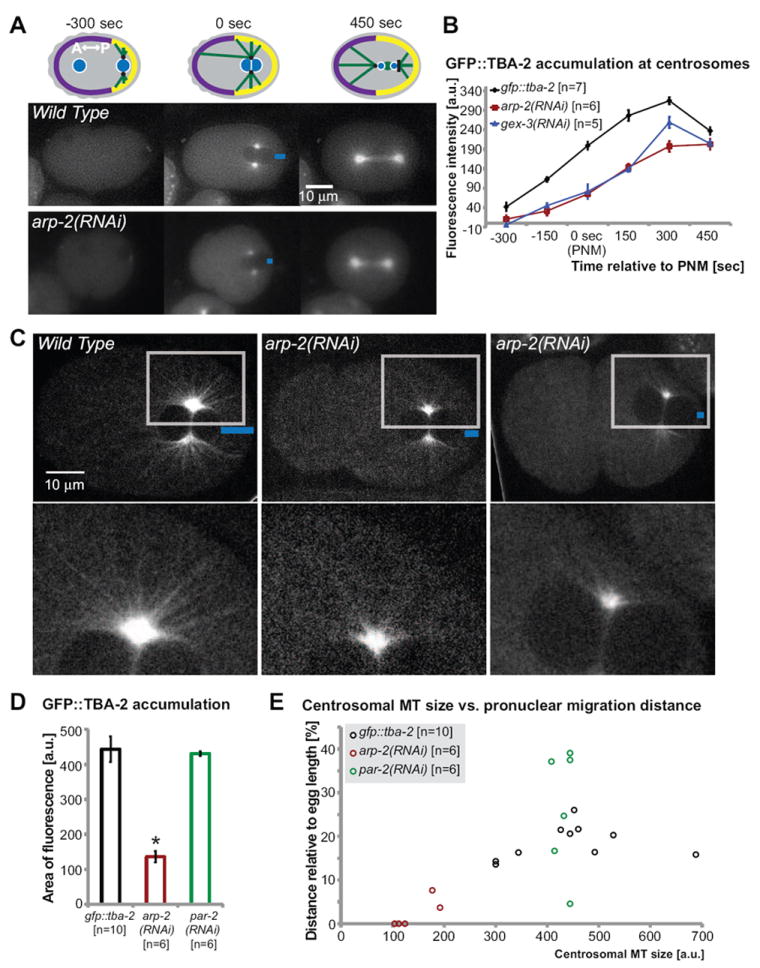
(A) Schematic illustration highlights the centrosomes and microtubules in the one-cell embryo. Centrosomes are shown in black, microtubules in green and pronuclei in blue. GFP∷TBA-2 accumulation in zygotes was examined by fluorescence time lapse imaging in wild-type embryos and in embryos depleted of arp-2 via RNAi. Blue bars indicate the migration distance of the male pronucleus from the posterior cortex. (B) Average fluorescence intensity at the centrosomes was determined by outlining the area of GFP∷TBA-2 fluorescence above an arbitrarily chosen threshold intensity, using the circle tool in ImageJ (see Methods for details). The average GFP∷TBA-2 fluorescence intensity in the cytoplasm was subtracted from the average fluorescence intensity at the centrosomes, and plotted in arbitrary units (a.u.). (C) Spinning disc confocal micrographs of gfp∷tba-2 (Audhya et al., 2005) and gfp∷tba-2 depleted of arp-2 via RNAi demonstrating microtubule abundance and organization in the one-cell embryo. Boxes indicate the region of magnification (lower panel) highlighting the microtubule filaments emerging from centrosomes. Distance of pronuclear migration is marked with blue bars. Two arp-2 RNAi embryos are shown, one with partial migration and one with almost no migration away from the posterior pole. (D) Average GFP∷TBA-2 accumulation at the centrosomes at pronuclear meeting was determined by measuring the fluorescent area around the centrosome decorated with GFP∷TBA-2 pixels above an arbitrarily chosen threshold intensity, using the circle tool in the program ImageJ, shown in arbitrary units (a.u.). (E) The area of high GFP∷TBA-2 accumulation around the centrosomes in individual embryos at pronuclear meeting, in arbitrary units (a.u.), is plotted against male pronuclear migration distance relative to total embryo length. Error bars indicate SEM; asterisks mark statistical significance, p<0.05.
To better characterize the assembly of MTs we used spinning disc confocal microscopy to make GFP∷TBA-2 movies in wild-type and arp-2 RNAi embryos captured at 15-second intervals. These faster movies supported the conclusions from Fig. 6A that there was decreased centrosomal MT density in arp-2 RNAi embryos. In addition, these movies showed fewer and shorter MTs emanating from the centrosomes (Fig. 6B; Movie 4). 100% of the arp-2 RNAi embryos showed these defects (n=6 movies). These results suggested that wild-type levels of MT intensity, length and abundance in the one-cell embryo depend on Arp2/3.
We compared the size of the area of high MT accumulation around the centrosomes to the distance migrated by the male pronucleus, and found that the larger the centrosomal MT region, the larger the migration distance, measured as a percent of total egg length (Fig. 6C). In arp-2(RNAi) embryos at pronuclear meeting (0 seconds), the centrosomal MT region was significantly smaller, and the male pronucleus either completely failed to migrate, or it migrated a reduced distance compared to its migration in wild-type embryos (Fig. 6C). This correlation is reminiscent of studies by Tsai and Ahringer (Tsai and Ahringer, 2007) that showed that the size of the sperm aster determined the time required to establish cortical polarity, with smaller asters taking longer to establish cortical polarity. Although the embryos depleted of Arp2/3 and its regulators did not show significantly delayed initiation of A/P polarity, as measured by asymmetrical accumulation of PAR-6∷GFP, they were defective in recruiting PAR-2 to the posterior cortex, and delayed in assembling the microtubule cytoskeleton at the posterior. PAR-2 has been reported to bind to MTs and this binding supports PAR-2 accumulation at the cortex (Zonies et al., 2010). Therefore the decreased astral MT assembly in embryos depleted of Arp2/3 could contribute to the decrease in PAR-2 accumulation at the posterior cortex (Fig. 4). To test if PAR-2 also affects MT growth, we measured GFP∷TBA-2 accumulation at comparable stages in embryos depleted of PAR-2 via RNAi. Loss of PAR-2 does not affect GFP∷TBA-2 accumulation (Fig. 6D), but showed instead, on average, slightly increased male pronuclear migration at pronuclear meeting (Fig. 2, Table 1). Therefore while PAR-2 does not regulate MT growth, it may influence MT interactions with the cortex required for male pronuclear migration.
DISCUSSION
The actin nucleator, Arp2/3, is shown here to contribute to three distinct nuclear migrations in three different tissues and at three different times in development. The effect is as strong or stronger than what is seen in null mutations of known nuclear migration genes of the SUN and KASH domain families. Our results indicate that there is a general requirement for the regulators of branched actin during nuclear migrations. Interestingly, plant researchers have suggested that WAVE and Arp2/3 support MT organization (Saedler et al., 2004; Zhang et al., 2005).
Understanding how the actin and microtubule cytoskeletons work together to position the nucleus during key events in development has been a challenging problem. Complete elimination of any important component of the cytoskeleton, such as occurs in studies that depolymerize or stabilize all microtubules or all actin, often gives results that are difficult to interpret. In the present study, we used genetic methods to remove specific cytoskeletal regulators in order to study their contribution to nuclear movements. The use of the one-cell zygote provided several advantages. The nuclear migrations in the zygote are not coupled to a cell migration, allowing us to focus on nuclear movements. In addition, the robustness of early embryos, that employ multiple mechanisms to ensure proper polarization (Rose and Kemphues, 1998), allowed us to remove Arp2/3-dependent actin nucleation and monitor several polarization events, in the context of embryos that maintained their overall polarity and continued cell divisions.
A role for Arp2/3 in symmetry breaking in the one-cell zygote
Anterior/posterior polarity in C. elegans requires a signal both from the sperm-derived centrosomes (Goldstein and Hird, 1996; Sadler and Shakes, 2000) and from microtubules (Tsai and Ahringer, 2007; Wallenfang and Seydoux, 2000). We report here that the generation of that signal requires Arp2/3. The sperm pronucleus reaches the future posterior pole through an Arp2/3-independent mechanism, and is also held at the posterior pole by an Arp2/3-independent mechanism. However, the next step in the establishment of anterior-posterior polarity requires Arp2/3 and its activators. Recent studies suggest that there are two parallel polarization pathways operating in the zygote. In one pathway, a centrosomal cue leads to a clearing of actomyosin contractility from the posterior pole (Jenkins et al., 2006; Motegi and Sugimoto, 2006; Schonegg and Hyman, 2006). This first pathway is delayed in Arp2/3 mutants, suggesting Arp2/3 contributes to the dynamics of actomyosin contractility (Fig. 3). A second polarization pathway, only visible under special conditions, (e.g. in mutants containing hypomorphic ect-2 alleles), requires the presence of the PAR proteins (Zonies et al., 2010). Loss of Arp2/3 also affects this second pathway, as cortical GFP∷PAR-2 establishment is reduced. Although some PAR-2 is recruited to the posterior cortex in embryos depleted of arp-2 (Fig. 4), some PAR-2-dependent functions appear compromised. For example, there are ectopic posterior NMY-2∷GFP puncta in embryos depleted of arp-2 (Fig. 3A), just as is seen in embryos depleted of PAR-2 (Munro et al., 2004). Therefore, the maintenance function of PAR-2 that keeps NMY-2∷GFP from returning to the posterior cortex may be compromised in arp-2 RNAi embryos. Further, loss of Arp2/3 leads to a delay in centrosomal MT growth at the male pronucleus. Since accumulation of PAR-2 at the posterior cortex has been proposed to be one of the symmetry-breaking events that signal the position of the centrosome to the cortex, and since PAR-2 accumulation on the posterior cortex requires that it bind MTs (Zonies et al., 2010), Arp2/3 appears to support this symmetry-breaking event.
Arp2/3 regulates MT growth required for nuclear migrations in the zygote
The delayed removal of actomyosin at the posterior and delayed accumulation of posterior polarity determinants correlates with decreased microtubule polymerization at the centrosomes. As a result the force that moves the nucleus away from the posterior pole appears to be compromised. Previous studies suggested that during male pronuclear migration, pulling forces rather than pushing forces on the MTs are dominant (Kimura and Onami, 2005). The “pulling mechanism” was proposed to use length-dependent pulling force generated by minus-end-directed motors anchored throughout the cytoplasm to position the male pronucleus (Kimura and Onami, 2005). We note that dynein-dependent pulling movements, including centrosome separation and the migration of the female pronucleus towards the male pronucleus (Gonczy et al., 1999), occur in embryos depleted of UNC-84 or Arp2/3, albeit with some changes. Our results do not discount a need for pulling forces. However, our results are consistent with a model where pushing forces at the posterior move the male pronucleus away from the posterior pole.
The “pushing mechanism” was proposed to use pushing force resulting from microtubule polymerization to position the male pronucleus (Kimura and Onami, 2005). The microtubules emanating from the centrosomes require cortical stiffness to push against the cortex (Kimura and Onami, 2005). These pushing forces would be stronger when the nucleus is closest to the posterior cortex, and could have been missed in previous studies, which appear to begin measuring nuclear movement at a later stage, once the nucleus has moved away from the cortex (Kimura and Onami, 2005). One simple explanation for how both UNC-84 and Arp2/3 may contribute to pushing forces is that both may provide stiffness to the two membranes needed for the male nuclear migration, the nuclear envelope membrane and the cortical membrane, so that the force created by MT polymerization can be converted into nuclear displacement rather than membrane deformation. Alternatively, both UNC-84, through its KASH partners, and Arp2/3, through unknown partners, may support factors that promote pushing forces, as has been shown for CLIP170 in promoting pushing forces at the plus end of MTs during nuclear centration in S. pombe (Daga et al., 2006). The distance that the male pronucleus travels to meet the female pronucleus, approximately 10um, is short enough that MTs would not buckle and pushing forces could be sufficient (Tolic-Norrelykke, 2008). Finally, an actin-rich cortical meshwork has been proposed to anchor microtubule motors (Brito et al., 2005).
Where is the branched actin that positions the male pronucleus?
F-actin becomes enriched at the anterior cortex as the zygote becomes polarized (Munro and Bowerman, 2009). However, the results shown here suggest that there is a population of posterior F-actin in the zygote that depends on Arp2/3. This actin could be at (1) the nuclear periphery, (2) the centrosomes, (3) the posterior cortex, or in all of these places. (1) Actin at the nuclear periphery could be recruited by ANC-1, a KASH domain protein that is anchored in the outer nuclear membrane and can bind to actin. ANC-1 is large enough (0.5um) that some models predict a direct link from the nucleus to the cell cortex via ANC-1 (Starr and Han, 2002; Starr and Han, 2003). We show here that loss of ANC-1 in combination with loss of KDP-1 results in a strong defect in movement away from the posterior cortex (Fig. 2). In addition, perinuclear actin was shown to form part of the TAN (transmembrane actin-associated nuclear) lines that form a bridge between the nuclear envelope and the cytoskeleton in a tissue culture system (Luxton et al., 2010). (2) The presence of actin at the centrosome is supported by mammalian tissue culture studies showing that Arp2/3 components are part of the centrosome during mitosis (Molli et al., 2010). (3) Other studies predict the presence of posterior cortical actin. When actin was depleted from C. elegans embryos via RNAi the male pronucleus detached prematurely from the posterior cortex (Velarde et al., 2007), which suggests that there is cortical actin at the posterior that positions the male pronucleus prior to migration. Imaging of NMY-2∷GFP reveals a small population of NMY-2 at the posterior of the zygote (Goulding et al., 2007; Munro et al., 2004). Our imaging results with Lifeact∷GFP support that there is a posterior cortical population of F-actin that depends on Arp2/3 (Fig. 3D) while leaving open the possibility that actin that has yet to be detected at the centrosome or around the nucleus could also be playing a role. Posterior cortical actin is ideally located to mediate the events shown here that depend on Arp2/3: generation of the polarity cue from the centrosome; promotion of MT plus end interactions with the cortex; and nuclear-centrosomal rotation in preparation for cell division. Further studies will be needed to better characterize the populations of actin in the zygote and to identify the molecules that link the population of posterior Arp2/3-dependent F-actin with the nucleus.
Supplementary Material
NHR-25∷YFP is expressed in the nuclei and cytoplasm of epidermal cells during morphogenesis. In wild-type nhr-25∷gfp embryos, the two dorsal rows of epidermal cells migrate to form one row during dorsal intercalation, and the nuclei within the cytoplasm of those cells migrate to the opposite side of the body midline. In contrast, embryos depleted of gex-2 failed in dorsal intercalation. In addition, the nuclei in the epidermis of gex-2 depleted embryos also failed to migrate to the opposite side of the body midline. Body axes of embryos are aligned with anterior to the left, dorsal up, the left side on the bottom and the right on top. Timing (minutes) relative to the first cell division. All movies of embryos made at 22°C.
Live DIC imaging of one wild-type and two gex-3(RNAi) embryos from 345 seconds before pronuclear meeting (PNM) to 90 seconds after PNM. The migrations of both the female pronucleus and of the male pronucleus can be compared. Embryos are oriented with anterior to the left, posterior to the right. Timing (sec) relative to pronuclear meeting (0 sec).
Live DIC imaging of wild-type, gex-3(zu196) or unc-84(e1410) embryos aligned at pronuclear meeting (0 seconds) shows the relative timing of events during the first cell cycle. Mutant embryos complete all of the events of the first cell cycle, with small delays in specific steps including delayed centration, and delayed nuclear-centrosomal rotation. However, the first cell division is asymmetrical, as in wild type.
Live fluorescence imaging of wild-type tba-2∷gfp one-cell embryos and embryos depleted of arp-2 by RNAi. Embryos are oriented with anterior to the left. Timing (sec) relative to pronuclear meeting (0 sec). The scale on the right indicates fluorescence intensity from low (blue) to high (white).
Acknowledgments
We would like to thank the NCRR-funded Caenorhabditis Genetics center, Anjon Audhya, Karen Oegema, Martin Srayko, Tony Hyman, Ken Kemphues, Kevin O Connell and Zhirong Bao for strains. We thank our colleagues for helpful suggestions on the manuscript: Matthew Buechner and Lesilee Rose. We would like to thank Kevin O’Connell, Ed Munro and Dan Starr for helpful discussions. Thanks to Jerry Rackowski at Zeiss for the demonstration of the Zeiss Cell Observer Microscope with Yokogawa CSU-X1 Laser Spinning Disc system. This research was funded by a postdoctoral grant from the New Jersey Commission for Cancer Research to H.X. and by grants from the NIH (GM081670) and NSF (0641123) to M.C.S.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Anderson DC, Gill JS, Cinalli RM, Nance J. Polarization of the C. elegans embryo by RhoGAP-mediated exclusion of PAR-6 from cell contacts. Science. 2008;320:1771–4. doi: 10.1126/science.1156063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Audhya A, Hyndman F, McLeod IX, Maddox AS, Yates JR, 3rd, Desai A, Oegema K. A complex containing the Sm protein CAR-1 and the RNA helicase CGH-1 is required for embryonic cytokinesis in Caenorhabditis elegans. J Cell Biol. 2005;171:267–79. doi: 10.1083/jcb.200506124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baugh LR, Hill AA, Claggett JM, Hill-Harfe K, Wen JC, Slonim DK, Brown EL, Hunter CP. The homeodomain protein PAL-1 specifies a lineage-specific regulatory network in the C. elegans embryo. Development. 2005;132:1843–54. doi: 10.1242/dev.01782. [DOI] [PubMed] [Google Scholar]
- Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77:71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brito DA, Strauss J, Magidson V, Tikhonenko I, Khodjakov A, Koonce MP. Pushing forces drive the comet-like motility of microtubule arrays in Dictyostelium. Mol Biol Cell. 2005;16:3334–40. doi: 10.1091/mbc.E05-01-0057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chhabra ES, Higgs HN. The many faces of actin: matching assembly factors with cellular structures. Nat Cell Biol. 2007;9:1110–21. doi: 10.1038/ncb1007-1110. [DOI] [PubMed] [Google Scholar]
- Cowan CR, Hyman AA. Asymmetric cell division in C. elegans: cortical polarity and spindle positioning. Annu Rev Cell Dev Biol. 2004a;20:427–53. doi: 10.1146/annurev.cellbio.19.111301.113823. [DOI] [PubMed] [Google Scholar]
- Cowan CR, Hyman AA. Centrosomes direct cell polarity independently of microtubule assembly in C. elegans embryos. Nature. 2004b;431:92–6. doi: 10.1038/nature02825. [DOI] [PubMed] [Google Scholar]
- Cuenca AA, Schetter A, Aceto D, Kemphues K, Seydoux G. Polarization of the C. elegans zygote proceeds via distinct establishment and maintenance phases. Development. 2003;130:1255–65. doi: 10.1242/dev.00284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daga RR, Yonetani A, Chang F. Asymmetric microtubule pushing forces in nuclear centering. Curr Biol. 2006;16:1544–50. doi: 10.1016/j.cub.2006.06.026. [DOI] [PubMed] [Google Scholar]
- Derivery E, Gautreau A. Generation of branched actin networks: assembly and regulation of the N-WASP and WAVE molecular machines. Bioessays. 2010;32:119–31. doi: 10.1002/bies.200900123. [DOI] [PubMed] [Google Scholar]
- Fridolfsson HN, Ly N, Meyerzon M, Starr DA. UNC-83 coordinates kinesin-1 and dynein activities at the nuclear envelope during nuclear migration. Dev Biol. 2009;338:237–50. doi: 10.1016/j.ydbio.2009.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridolfsson HN, Starr DA. Kinesin-1 and dynein at the nuclear envelope mediate the bidirectional migrations of nuclei. J Cell Biol. 2010;191:115–28. doi: 10.1083/jcb.201004118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galli M, van den Heuvel S. Determination of the cleavage plane in early C. elegans embryos. Annu Rev Genet. 2008;42:389–411. doi: 10.1146/annurev.genet.40.110405.090523. [DOI] [PubMed] [Google Scholar]
- Goldstein B, Hird SN. Specification of the anteroposterior axis in Caenorhabditis elegans. Development. 1996;122:1467–74. doi: 10.1242/dev.122.5.1467. [DOI] [PubMed] [Google Scholar]
- Gonczy P, Pichler S, Kirkham M, Hyman AA. Cytoplasmic dynein is required for distinct aspects of MTOC positioning, including centrosome separation, in the one cell stage Caenorhabditis elegans embryo. J Cell Biol. 1999;147:135–50. doi: 10.1083/jcb.147.1.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goulding MB, Canman JC, Senning EN, Marcus AH, Bowerman B. Control of nuclear centration in the C. elegans zygote by receptor-independent Galpha signaling and myosin II. J Cell Biol. 2007;178:1177–91. doi: 10.1083/jcb.200703159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gundersen GG, Wen Y, Eng CH, Schmoranzer J, Cabrera-Poch N, Morris EJ, Chen M, Gomes ER. Regulation of microtubules by Rho GTPases in migrating cells. Novartis Found Symp. 2005;269:106–16. discussion 116-26, 223-30. [PubMed] [Google Scholar]
- Hamill DR, Severson AF, Carter JC, Bowerman B. Centrosome maturation and mitotic spindle assembly in C. elegans require SPD-5, a protein with multiple coiled-coil domains. Dev Cell. 2002;3:673–84. doi: 10.1016/s1534-5807(02)00327-1. [DOI] [PubMed] [Google Scholar]
- Hedgecock EM, Thomson JN. A gene required for nuclear and mitochondrial attachment in the nematode Caenorhabditis elegans. Cell. 1982;30:321–30. doi: 10.1016/0092-8674(82)90038-1. [DOI] [PubMed] [Google Scholar]
- Heil-Chapdelaine RA, Adames NR, Cooper JA. Formin’ the connection between microtubules and the cell cortex. J Cell Biol. 1999;144:809–11. doi: 10.1083/jcb.144.5.809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horvitz HR, Sulston JE. Isolation and genetic characterization of cell-lineage mutants of the nematode Caenorhabditis elegans. Genetics. 1980;96:435–54. doi: 10.1093/genetics/96.2.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huisman SM, Segal M. Cortical capture of microtubules and spindle polarity in budding yeast - where’s the catch? J Cell Sci. 2005;118:463–71. doi: 10.1242/jcs.01650. [DOI] [PubMed] [Google Scholar]
- Hyman AA. Centrosome movement in the early divisions of Caenorhabditis elegans: a cortical site determining centrosome position. J Cell Biol. 1989;109:1185–93. doi: 10.1083/jcb.109.3.1185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irazoqui JE, Lew DJ. Polarity establishment in yeast. J Cell Sci. 2004;117:2169–71. doi: 10.1242/jcs.00953. [DOI] [PubMed] [Google Scholar]
- Jenkins N, Saam JR, Mango SE. CYK-4/GAP provides a localized cue to initiate anteroposterior polarity upon fertilization. Science. 2006;313:1298–301. doi: 10.1126/science.1130291. [DOI] [PubMed] [Google Scholar]
- Kamath RS, Ahringer J. Genome-wide RNAi screening in Caenorhabditis elegans. Methods. 2003;30:313–21. doi: 10.1016/s1046-2023(03)00050-1. [DOI] [PubMed] [Google Scholar]
- Kemphues K. PARsing embryonic polarity. Cell. 2000;101:345–8. doi: 10.1016/s0092-8674(00)80844-2. [DOI] [PubMed] [Google Scholar]
- Kimura A, Onami S. Computer simulations and image processing reveal length-dependent pulling force as the primary mechanism for C. elegans male pronuclear migration. Dev Cell. 2005;8:765–75. doi: 10.1016/j.devcel.2005.03.007. [DOI] [PubMed] [Google Scholar]
- Kurisu S, Takenawa T. The WASP and WAVE family proteins. Genome Biol. 2009;10:226. doi: 10.1186/gb-2009-10-6-226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee KK, Starr D, Cohen M, Liu J, Han M, Wilson KL, Gruenbaum Y. Lamin-dependent localization of UNC-84, a protein required for nuclear migration in Caenorhabditis elegans. Mol Biol Cell. 2002;13:892–901. doi: 10.1091/mbc.01-06-0294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu C, Mains PE. Mutations of a redundant alpha-tubulin gene affect Caenorhabditis elegans early embryonic cleavage via MEI-1/katanin-dependent and -independent pathways. Genetics. 2005;170:115–26. doi: 10.1534/genetics.104.030106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luxton GW, Gomes ER, Folker ES, Vintinner E, Gundersen GG. Linear arrays of nuclear envelope proteins harness retrograde actin flow for nuclear movement. Science. 2010;329:956–9. doi: 10.1126/science.1189072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malone CJ, Fixsen WD, Horvitz HR, Han M. UNC-84 localizes to the nuclear envelope and is required for nuclear migration and anchoring during C. elegans development. Development. 1999;126:3171–81. doi: 10.1242/dev.126.14.3171. [DOI] [PubMed] [Google Scholar]
- Malone CJ, Misner L, Le Bot N, Tsai MC, Campbell JM, Ahringer J, White JG. The C. elegans hook protein, ZYG-12, mediates the essential attachment between the centrosome and nucleus. Cell. 2003;115:825–36. doi: 10.1016/s0092-8674(03)00985-1. [DOI] [PubMed] [Google Scholar]
- McGee MD, Rillo R, Anderson AS, Starr DA. UNC-83 IS a KASH protein required for nuclear migration and is recruited to the outer nuclear membrane by a physical interaction with the SUN protein UNC-84. Mol Biol Cell. 2006;17:1790–801. doi: 10.1091/mbc.E05-09-0894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGee MD, Stagljar I, Starr DA. KDP-1 is a nuclear envelope KASH protein required for cell-cycle progression. J Cell Sci. 2009;122:2895–905. doi: 10.1242/jcs.051607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIntire SL, Reimer RJ, Schuske K, Edwards RH, Jorgensen EM. Identification and characterization of the vesicular GABA transporter. Nature. 1997;389:870–6. doi: 10.1038/39908. [DOI] [PubMed] [Google Scholar]
- Meyerzon M, Fridolfsson HN, Ly N, McNally FJ, Starr DA. UNC-83 is a nuclear-specific cargo adaptor for kinesin-1-mediated nuclear migration. Development. 2009;136:2725–33. doi: 10.1242/dev.038596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molk JN, Bloom K. Microtubule dynamics in the budding yeast mating pathway. J Cell Sci. 2006;119:3485–90. doi: 10.1242/jcs.03193. [DOI] [PubMed] [Google Scholar]
- Molli PR, Li DQ, Bagheri-Yarmand R, Pakala SB, Katayama H, Sen S, Iyer J, Chernoff J, Tsai MY, Nair SS, Kumar R. Arpc1b, a centrosomal protein, is both an activator and substrate of Aurora A. J Cell Biol. 2010;190:101–14. doi: 10.1083/jcb.200908050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motegi F, Sugimoto A. Sequential functioning of the ECT-2 RhoGEF, RHO-1 and CDC-42 establishes cell polarity in Caenorhabditis elegans embryos. Nat Cell Biol. 2006;8:978–85. doi: 10.1038/ncb1459. [DOI] [PubMed] [Google Scholar]
- Munro E, Bowerman B. Cellular symmetry breaking during Caenorhabditis elegans development. Cold Spring Harb Perspect Biol. 2009;1:a003400. doi: 10.1101/cshperspect.a003400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munro E, Nance J, Priess JR. Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo. Dev Cell. 2004;7:413–24. doi: 10.1016/j.devcel.2004.08.001. [DOI] [PubMed] [Google Scholar]
- O’Connell KF, Maxwell KN, White JG. The spd-2 gene is required for polarization of the anteroposterior axis and formation of the sperm asters in the Caenorhabditis elegans zygote. Dev Biol. 2000;222:55–70. doi: 10.1006/dbio.2000.9714. [DOI] [PubMed] [Google Scholar]
- Page BD, Diede SJ, Tenlen JR, Ferguson EL. EEL-1, a Hect E3 ubiquitin ligase, controls asymmetry and persistence of the SKN-1 transcription factor in the early C. elegans embryo. Development. 2007;134:2303–14. doi: 10.1242/dev.02855. [DOI] [PubMed] [Google Scholar]
- Patel FB, Bernadskaya YY, Chen E, Jobanputra A, Pooladi Z, Freeman KL, Gally C, Mohler WA, Soto MC. The WAVE/SCAR complex promotes polarized cell movements and actin enrichment in epithelia during C. elegans embryogenesis. Dev Biol. 2008;324:297–309. doi: 10.1016/j.ydbio.2008.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pohl C, Bao Z. Chiral forces organize left-right patterning in C. elegans by uncoupling midline and anteroposterior axis. Dev Cell. 2010;19:402–12. doi: 10.1016/j.devcel.2010.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollard TD. Regulation of actin filament assembly by Arp2/3 complex and formins. Annu Rev Biophys Biomol Struct. 2007;36:451–77. doi: 10.1146/annurev.biophys.35.040405.101936. [DOI] [PubMed] [Google Scholar]
- Pruyne D, Gao L, Bi E, Bretscher A. Stable and dynamic axes of polarity use distinct formin isoforms in budding yeast. Mol Biol Cell. 2004;15:4971–89. doi: 10.1091/mbc.E04-04-0296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose LS, Kemphues KJ. Early patterning of the C. elegans embryo. Annu Rev Genet. 1998;32:521–45. doi: 10.1146/annurev.genet.32.1.521. [DOI] [PubMed] [Google Scholar]
- Rose MD. Nuclear fusion in the yeast Saccharomyces cerevisiae. Annu Rev Cell Dev Biol. 1996;12:663–95. doi: 10.1146/annurev.cellbio.12.1.663. [DOI] [PubMed] [Google Scholar]
- Sadler PL, Shakes DC. Anucleate Caenorhabditis elegans sperm can crawl, fertilize oocytes and direct anterior-posterior polarization of the 1-cell embryo. Development. 2000;127:355–66. doi: 10.1242/dev.127.2.355. [DOI] [PubMed] [Google Scholar]
- Saedler R, Mathur N, Srinivas BP, Kernebeck B, Hulskamp M, Mathur J. Actin control over microtubules suggested by DISTORTED2 encoding the Arabidopsis ARPC2 subunit homolog. Plant Cell Physiol. 2004;45:813–22. doi: 10.1093/pcp/pch103. [DOI] [PubMed] [Google Scholar]
- Schmidt DJ, Rose DJ, Saxton WM, Strome S. Functional analysis of cytoplasmic dynein heavy chain in Caenorhabditis elegans with fast-acting temperature-sensitive mutations. Mol Biol Cell. 2005;16:1200–12. doi: 10.1091/mbc.E04-06-0523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schonegg S, Constantinescu AT, Hoege C, Hyman AA. The Rho GTPase-activating proteins RGA-3 and RGA-4 are required to set the initial size of PAR domains in Caenorhabditis elegans one-cell embryos. Proc Natl Acad Sci U S A. 2007;104:14976–81. doi: 10.1073/pnas.0706941104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schonegg S, Hyman AA. CDC-42 and RHO-1 coordinate acto-myosin contractility and PAR protein localization during polarity establishment in C. elegans embryos. Development. 2006;133:3507–16. doi: 10.1242/dev.02527. [DOI] [PubMed] [Google Scholar]
- Soto MC, Qadota H, Kasuya K, Inoue M, Tsuboi D, Mello CC, Kaibuchi K. The GEX-2 and GEX-3 proteins are required for tissue morphogenesis and cell migrations in C. elegans. Genes Dev. 2002;16:620–32. doi: 10.1101/gad.955702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer AG, Orita S, Malone CJ, Han M. A RHO GTPase-mediated pathway is required during P cell migration in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2001;98:13132–7. doi: 10.1073/pnas.241504098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Starr DA, Fischer JA. KASH ‘n Karry: the KASH domain family of cargo-specific cytoskeletal adaptor proteins. Bioessays. 2005;27:1136–46. doi: 10.1002/bies.20312. [DOI] [PubMed] [Google Scholar]
- Starr DA, Fridolfsson HN. Interactions between nuclei and the cytoskeleton are mediated by SUN-KASH nuclear-envelope bridges. Annu Rev Cell Dev Biol. 2010;26:421–44. doi: 10.1146/annurev-cellbio-100109-104037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Starr DA, Han M. Role of ANC-1 in tethering nuclei to the actin cytoskeleton. Science. 2002;298:406–9. doi: 10.1126/science.1075119. [DOI] [PubMed] [Google Scholar]
- Starr DA, Han M. ANChors away: an actin based mechanism of nuclear positioning. J Cell Sci. 2003;116:211–6. doi: 10.1242/jcs.00248. [DOI] [PubMed] [Google Scholar]
- Starr DA, Hermann GJ, Malone CJ, Fixsen W, Priess JR, Horvitz HR, Han M. unc-83 encodes a novel component of the nuclear envelope and is essential for proper nuclear migration. Development. 2001;128:5039–50. doi: 10.1242/dev.128.24.5039. [DOI] [PubMed] [Google Scholar]
- Stauffer TP, Ahn S, Meyer T. Receptor-induced transient reduction in plasma membrane PtdIns(4,5)P2 concentration monitored in living cells. Curr Biol. 1998;8:343–6. doi: 10.1016/s0960-9822(98)70135-6. [DOI] [PubMed] [Google Scholar]
- Sulston JE, Horvitz HR. Abnormal cell lineages in mutants of the nematode Caenorhabditis elegans. Dev Biol. 1981;82:41–55. doi: 10.1016/0012-1606(81)90427-9. [DOI] [PubMed] [Google Scholar]
- Timmons L, Court DL, Fire A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene. 2001;263:103–12. doi: 10.1016/s0378-1119(00)00579-5. [DOI] [PubMed] [Google Scholar]
- Tolic-Norrelykke IM. Push-me-pull-you: how microtubules organize the cell interior. Eur Biophys J. 2008;37:1271–8. doi: 10.1007/s00249-008-0321-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai MC, Ahringer J. Microtubules are involved in anterior-posterior axis formation in C. elegans embryos. J Cell Biol. 2007;179:397–402. doi: 10.1083/jcb.200708101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umeshima H, Hirano T, Kengaku M. Microtubule-based nuclear movement occurs independently of centrosome positioning in migrating neurons. Proc Natl Acad Sci U S A. 2007;104:16182–7. doi: 10.1073/pnas.0708047104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varnai P, Balla T. Visualization of phosphoinositides that bind pleckstrin homology domains: calcium- and agonist-induced dynamic changes and relationship to myo-[3H]inositol-labeled phosphoinositide pools. J Cell Biol. 1998;143:501–10. doi: 10.1083/jcb.143.2.501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velarde N, Gunsalus KC, Piano F. Diverse roles of actin in C. elegans early embryogenesis. BMC Dev Biol. 2007;7:142. doi: 10.1186/1471-213X-7-142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallenfang MR, Seydoux G. Polarization of the anterior-posterior axis of C. elegans is a microtubule-directed process. Nature. 2000;408:89–92. doi: 10.1038/35040562. [DOI] [PubMed] [Google Scholar]
- Walston T, Tuskey C, Edgar L, Hawkins N, Ellis G, Bowerman B, Wood W, Hardin J. Multiple Wnt signaling pathways converge to orient the mitotic spindle in early C. elegans embryos. Dev Cell. 2004;7:831–41. doi: 10.1016/j.devcel.2004.10.008. [DOI] [PubMed] [Google Scholar]
- White J, Strome S. Cleavage plane specification in C. elegans: how to divide the spoils. Cell. 1996;84:195–8. doi: 10.1016/s0092-8674(00)80974-5. [DOI] [PubMed] [Google Scholar]
- Zhang X, Dyachok J, Krishnakumar S, Smith LG, Oppenheimer DG. IRREGULAR TRICHOME BRANCH1 in Arabidopsis encodes a plant homolog of the actin-related protein2/3 complex activator Scar/WAVE that regulates actin and microtubule organization. Plant Cell. 2005;17:2314–26. doi: 10.1105/tpc.104.028670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zonies S, Motegi F, Hao Y, Seydoux G. Symmetry breaking and polarization of the C. elegans zygote by the polarity protein PAR-2. Development. 2010;137:1669–77. doi: 10.1242/dev.045823. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
NHR-25∷YFP is expressed in the nuclei and cytoplasm of epidermal cells during morphogenesis. In wild-type nhr-25∷gfp embryos, the two dorsal rows of epidermal cells migrate to form one row during dorsal intercalation, and the nuclei within the cytoplasm of those cells migrate to the opposite side of the body midline. In contrast, embryos depleted of gex-2 failed in dorsal intercalation. In addition, the nuclei in the epidermis of gex-2 depleted embryos also failed to migrate to the opposite side of the body midline. Body axes of embryos are aligned with anterior to the left, dorsal up, the left side on the bottom and the right on top. Timing (minutes) relative to the first cell division. All movies of embryos made at 22°C.
Live DIC imaging of one wild-type and two gex-3(RNAi) embryos from 345 seconds before pronuclear meeting (PNM) to 90 seconds after PNM. The migrations of both the female pronucleus and of the male pronucleus can be compared. Embryos are oriented with anterior to the left, posterior to the right. Timing (sec) relative to pronuclear meeting (0 sec).
Live DIC imaging of wild-type, gex-3(zu196) or unc-84(e1410) embryos aligned at pronuclear meeting (0 seconds) shows the relative timing of events during the first cell cycle. Mutant embryos complete all of the events of the first cell cycle, with small delays in specific steps including delayed centration, and delayed nuclear-centrosomal rotation. However, the first cell division is asymmetrical, as in wild type.
Live fluorescence imaging of wild-type tba-2∷gfp one-cell embryos and embryos depleted of arp-2 by RNAi. Embryos are oriented with anterior to the left. Timing (sec) relative to pronuclear meeting (0 sec). The scale on the right indicates fluorescence intensity from low (blue) to high (white).