

Abstract
We investigated whether there were regional differences in the quadriceps enthesis and the patella bone structure that could suggest unequal force transmission to the patella. Quadriceps tendon enthesis was removed by cutting the patellae transversally in the middle and the quadriceps tendon approximately 1 cm from the bone. Tissues were post-fixed, decalcified, dehydrated through and embedded in paraffin wax. Serial longitudinal sections were cut, mounted on glass slides at 1-mm intervals and slides were stained. Trabecular architecture was analysed from digital images taken from the histological slides, and regional differences at the enthesis in the thickness of the uncalcified fibrocartilage and the cortical zone of calcified tissue (calcified cartilage and lamellar bone) were evaluated. At the quadriceps enthesis, the thickness of the cortical zone of calcified tissue was significantly greater in the central part of the enthesis than medially and laterally. The trabeculae were thicker in the central and lateral parts compared with the medial region. Similarly, the zone of uncalcified fibrocartilage was thicker laterally and centrally than medially. Bone structure and the thickness of uncalcified fibrocartilage presented a similarity between the centre and the lateral parts; however, the medial side was different. We suggest that the mechanical stress at the proximal quadriceps tendon enthesis is higher laterally and centrally compared with medially. This could induce a lateral patellar translation, which is potentially a risk factor for knee osteoarthritis.
Keywords: fibrocartilage, knee, osteoarthritis, patella, quadriceps
Introduction
Patellofemoral pain syndrome is related to a combination of factors involving malalignment of the knee extensor mechanism (Fredericson & Yoon, 2006; Balachandar et al. 2011). The common presentation is knee pain in association with positions of the knee that result in increased or misdirected mechanical forces between the kneecap and femur (Fredericson & Yoon, 2006). A common misconception is that the patella only moves in an up-and-down direction. In fact, it also tilts and rotates, so there are various points of contact between the undersurface of the patella and the femur (Koh et al. 1992). Repetitive contact associated with elevated joint stress at any of these areas, sometimes combined with maltracking of the patella, is the likely mechanism of retropatellar and peripatellar pain (Benjamin et al. 1991). Many theories have been proposed to explain the aetiology of the patella maltracking. These include biomechanical, muscular and overuse theories (Bolgla & Boling, 2011). It is still not known whether the problem primarily relates to a unique anatomical feature (i.e. alignment between the hip and the knee), to an unequal pattern of force transmission between the quadriceps muscles and the patella, or both (Bolgla & Boling, 2011).
It is generally accepted that changes in force transmission through cancellous bone result in a modification of its architecture. This is the principle underpinning Wolff’s law. It is widely assumed that analysis of trabecular bone architecture provides valuable information on stress patterns within cancellous bone (Pal & Routal, 1998). Tendons are capable of responding to exercise or immobilization by altering their tensile strength Woo & Buckwalter, 1988). The cells in tendons and ligaments are capable of detecting changes in mechanical load and co-ordinating their response to alter the composition of the extracellular matrix. One of the most obvious ways in which the extracellular matrix of tendons and ligaments is modified in response to load is by the formation of a fibrocartilaginous matrix at sites where the tendons or ligaments are under mechanical stress (Benjamin & Ralphs, 1998). There is more fibrocartilage at the horns of the more mobile lateral menisci of the knee joint than the medial menisci (Benjamin et al. 1991). Similarly, there is a greater quantity of fibrocartilage at the insertion of biceps than at brachialis or triceps (Benjamin et al. 1991, 1992), because although biceps, triceps and brachialis are all involved in flexion or extension of the elbow joint, biceps alone acts as a supinator and thus has a distinctive triplanar motion at its insertion site (Benjamin & Ralphs, 1998). Movement and load are the mechanical stimulus that triggers the metaplasia of fibroblasts to fibrocartilage cells, and there is a good correlation between the quantity of uncalcified fibrocartilage at an enthesis and the extent of movement that occurs between tendon/ligament and bone (Benjamin & Ralphs, 1998). Therefore, in order to evaluate the pattern of force transmission between the quadriceps muscles and the patella, we have described the fibrocartilaginous structure of the quadriceps tendon enthesis and the regional differences in the trabecular architecture of the patella. This may help to explain the patella instability, and would clarify the contribution of the different components of the quadriceps muscles in the alignment of the kneecap.
Materials and methods
This study meets the NHS Ethics Research Committee-provided approval in accordance with the principles in the declaration of Helsinki. Seven dissecting room cadavers (five male and five female) were donated to the Department of Anatomy for anatomical examination. The donors ranged in age from 68 to 78 years at the time of death; the mean age at death was 73 ± 5 years. None of the donors’ knees appeared to have undergone prior lower limb operative procedures. Details about the lifetime activities of the donors were not available. The cadavers had been perfused for 72 h with an embalming fluid containing 4% formaldehyde and 25% alcohol. In total, 20 patellae (patella on both sides in 10 cadavers) were removed from each cadaver for histological and morphometrical analysis.
Histology
The procedures used have been described by a previous study (Toumi et al. 2006). Quadriceps tendon enthesis was removed by cutting the patellae transversally in the middle and the quadriceps tendon approximately 1 cm from the bone. Each specimen was divided longitudinally into medial, central and lateral parts. For examining the quadriceps tendon enthesis, tissue was post-fixed in 10% neutral buffered formalin, decalcified with 5% nitric acid, dehydrated through a graded alcohol series, cleared in xylene and embedded in paraffin wax. Serial longitudinal sections were cut at 8 μm throughout the medial, central and lateral thirds of the enthesis, and 12 sections were mounted on glass slides at 1-mm intervals. Slides were stained with Hall and Brunt’s quadruple stain, Masson’s trichrome (for photography) and toluidine blue (for fibrocartilage metachromasia).
Trabeculae structure analysis
The histological slides were scanned using Epson scanner (Epson perfection 1250 Model G820A, Seiko Epson Corporation, Japan) with a high-resolution algorithm (1200 × 2400 dpi; Fig. 5A). The imaging parameters were identical for all the slides. Images were transferred to a personal computer and structural analyses of the bone were performed. Algorithms used to characterize bone architecture were all developed in the laboratory using Matlab Software.
The analyses used have been described previously by Toumi et al. (2006). The boundary between the thin cortical bone and the underlying trabecular bone was defined using an automatic contour detection algorithm. Subsequently, a segmentation process permitted separation of spicules from bone marrow. The segmentation was made with an edge detection using a LaPlacian–Gaussian filter. This includes both a smoothing filter (which convolutes the image by a Gaussian filter) and a second-order derivative filter. Tuning of this combined filter addresses the size of the smoothing window, but also the variance of the convolutive Gaussian filter. Zero-crossing detection in the resulting image provides a binary image in which dark regions represent the bone marrow and light regions represent trabeculae. Several variables were calculated on the binary images based on two-dimensional (2D) analysis – these were: apparent bone volume = trabecular bone/tissue volume; and apparent trabecular thickness = 2/(trabecular separation/apparent bone volume).
Morphometric analysis of histological sections
The method has been described previously by Toumi et al. (2006). At all three parts (medial, central and lateral) of the enthesis and following a method adopted originally by Benjamin et al. (1991), the thickness of total calcified tissue was determined by measuring the thickness of the cortical zone of calcified tissue (calcified cartilage and lamellar bone). The thickness of the zone of uncalcified fibrocartilage was estimated by measuring the distance from the tidemark to the furthest recognizable chondrocyte within the tendon. Five such measurements were taken at equal intervals across the attachment site on one slide at each 1-mm sample point.
Statistical analysis
Mean values reported are given ± standard deviations (SD). A repeated-measures analysis of variance (anova) with Fisher’s post-hoc tests was used to compare medial, central and lateral parts of the patella for each variable. The P-value for significance was set at 0.05.
Results
The results are summarized in Figs 1–7, and the most significant findings are detailed below.
Fig. 1.
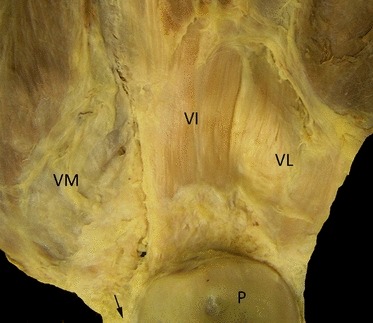
Gross anatomy of the patella and the quadriceps: posterior views. Note the VL and VI tendons runs vertically, and the VM inserts obliquely to the patella and becomes tendinous only a few millimetres before it inserts to the patella. Note the small region of direct continuity of the VM tendon (arrow) with the patellar tendon. P, patella; VI, vastus intermedius; VL, vastus lateralis; VM, vastus medialis.
Fig. 2.
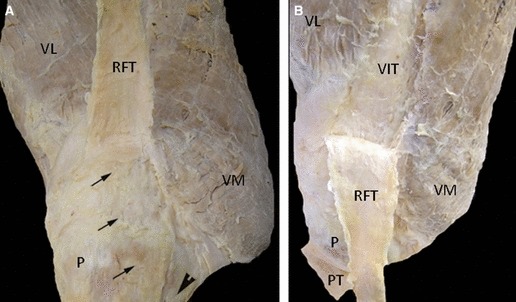
Gross anatomy of the patella and the quadriceps (A, B): anterior view views. Note the RF is layering superficially and centrally on the top of the quadriceps tendon. RFT could be easily separated from the rest of the quadriceps tendon. There are few superficial fibres that continue on the top of the patella (arrows) and join the patella tendon. P, patella; PT, patella tendon; RF, rectus femoris; RFT, rectus femoris tendon; VIT, vastus intermedius tendon; VL, vastus lateralis; VM, vastus medialis.
Fig. 4.
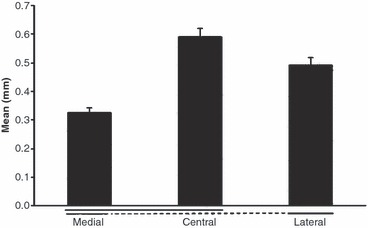
The thickness of the uncalcified fibrocartilage in the lateral, central and medial parts of the quadriceps tendon enthesis (mean values ± SD). Significant differences (P < 0.05) are underlined. Note the zone of uncalcified fibrocartilage was significantly thicker laterally and centrally than medially.
Fig. 5.
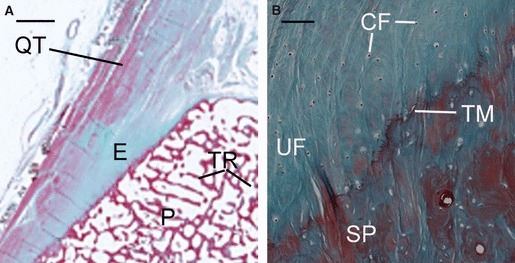
Histological sections of the quadriceps tendon enthesis. (A) A low-magnification view (Epson scanner) of a histological section of the superior pole of the patella (P) showing the quadriceps tendon (QT), the enthesis (E) and the trabeculae (TR). Scale bars: 2 mm. Masson’s trichrome. (B) A high-magnification view at the quadriceps tendon enthesis showing the uncalcified fibrocartilage (UF), the fibrocartilage cells (FC), the tide mark (TM) and the subchondral bone plate (SP). Scale bars: 0.1 mm. Masson’s trichrome.
Fig. 7.
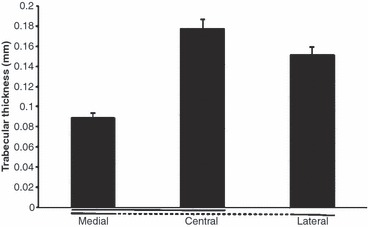
The trabeculae thickness in the lateral, central and medial patella facets expressed as a percentage of the total tissue volume (mean values ± SD). Significant differences (P < 0.05) are underlined. Values were significantly higher centrally and laterally than medially.
Gross anatomy (Figs 1 and 2): the attachment of the quadriceps tendon to the patella is shown in Figs 1 and 2. The cadaveric dissections showed that the quadriceps tendon is multilayered and formed by the convergence of all four muscles just proximal to the superior patella. The rectus femoris (RF) is the most superficial layer inserting on to the patella and becomes tendinous 3.2 ± 0.4 cm proximal to the patella. The RF tendon is layering superficially and centrally on the top of the quadriceps tendon. It could be easily separated from the rest of the quadriceps tendon simply by introducing the thumb between the RF tendon and the rest of the quadriceps tendon. There are a few superficial fibres that continue on the top of the patella and join the patella tendon. Vastus lateralis (VL) and vastus intermedius (VI) tendons run vertically and join together at 2.8 ± 0.5 cm proximal to its insertion into the patella. However, the vastus medialis (VM) inserts obliquely to the patella and becomes tendinous only a few millimetres before it inserts to the patella (1.1 ± 0.7 cm).
Histological analysis of the quadriceps tendon enthesis (Figs 3–5): the thickness of the cortical zone of calcified tissue (calcified cartilage and lamellar bone) was significantly greater in the central part of the enthesis than medially (P = 0.02) and laterally (P = 0.04). Values in millimetres were 1.27 ± 0.12 centrally, 0.93 ± 0.11 laterally and 0.72 ± 0.07 medially. There was also a significant difference between the medial and lateral side. The thickness of the calcified tissue was significantly higher laterally (P = 0.04). The zone of uncalcified fibrocartilage was significantly thicker laterally and centrally than medially. Values in millimetres were 0.59 ± 0.11 centrally, 0.49 ± 0.09 laterally and 0.32 ± 0.07 medially). However, no significant difference was observed between the central and lateral parts (P = 0.08).
Fig. 3.
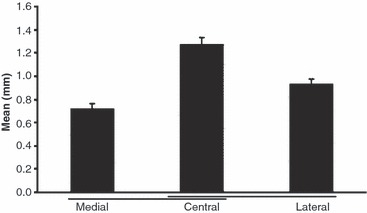
The thickness of the cortical zone of calcified tissue (calcified cartilage and lamellar bone) in the lateral, central and medial parts of the quadriceps tendon enthesis (mean values ± SD). Significant differences (P < 0.05) are underlined. Note values were significantly greater in the central part of the enthesis than medially (P = 0.02) and laterally (P = 0.04).
Bone structure analysis (Figs 6 and 7): essentially, in the proximal region of the patella, the apparent bone volume (corresponding to the sum of trabecular volume divided by the total tissue volume) was significantly higher centrally and laterally than it was medially (i.e. central: 0.47 ± 0.04; lateral: 0.43 ± 0.02; medial: 0.37 ± 0.02; Fig. 3), with no significant difference between the central and lateral parts (P = 0.2). There were similar differences in the apparent trabecular thickness. The trabeculae were thicker in the central and lateral parts compared with the medial region (i.e. central: 0.17 ± 0.05 mm; lateral: 0.15 ± 0.04 mm; medial: 0.08 ± 0.03 mm).
Fig. 6.
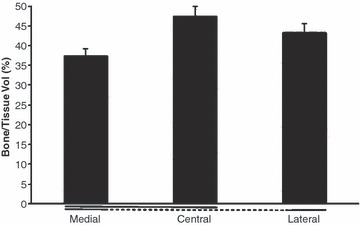
The apparent bone volume (corresponding to the sum of trabecular volume divided by the total tissue volume) in the lateral, central and medial patella facets expressed as a percentage of the total tissue volume (mean values ± SD). Significant differences (P < 0.05) are underlined. Values were significantly higher centrally and laterally than medially.
Discussion
The cells in tendons and ligaments are capable of detecting changes in mechanical load and coordinating their response to alter the composition of the fibrocartilaginous matrix at sites where the tendons or ligaments are under mechanical stress (Benjamin & Ralphs, 1998). Where a tendon is compressed or loaded in tension, there is an equal and opposite force acting on the bone (Benjamin & Ralphs, 1998), which results in a modification of its architecture. Thus, analysis of trabecular bone and enthesis architectures provides valuable information on stress patterns transmitted to the joint (Pal & Routal, 1998). In order to explain the patella instability and explore the different pattern of force transmission from quadriceps to the patella, the purpose of this study therefore was to analyse the regional variations in the fibrocartilage and the trabecular architecture of the quadriceps tendon enthesis.
The present results showed a regional variation in the fibrocartilage at the quadriceps tendon enthesis and the trabeculae architecture of the proximal region of the patella. Collectively, the data showed that a greater amount of total calcified and non-calcified tissues at the quadriceps tendon enthesis occurred centrally and laterally than medially. This suggests that it is these sites that are subjected to the greatest force. Furthermore, the close correspondence in the amount of bone at the proximal side of the patella (more bone volume laterally and centrally than medially) and the quantity of fibrocartilage at the quadriceps tendon enthesis (more fibrocartilage laterally and centrally than medially) provides further evidence to suggest that the load transferred from the quadriceps to the patella is greater laterally and centrally than medially. This could induce a lateral patellar translation that is potentially a risk factor for knee osteoarthritis.
The gross anatomy dissections showed that the quadriceps tendon is multilayered and formed by the convergence of all four muscles just proximal to the superior patella. The central and lateral layers represented by the RF, VI and VL insert centrally and laterally, and they become tendinous 2.8 ± 0.5 cm before the patella. At the enthesis, the trabecular thickness, the amount of bone and non-calcified fibrocartilage were similar (statistically not different) at the lateral and central parts. The parallel in these anatomical and fibrocartilaginous findings between the central and lateral parts suggests a comparable load at the central and lateral parts of the patella, but also a functional similarity of the lateral and central components of the quadriceps muscles. On the other hand, however, the VM inserts obliquely to the patella and becomes tendinous only 1.1 ± 0.7 cm before it inserts to the patella. Moreover, calcified fibrocartilage, trabecular thickness and the amount of bone were significantly different at the medial site compared with the lateral and central regions. Such results provide further evidence to suggest that the medial side of the patella receives a different load and the medial component of the quadriceps serves a different function(s) compared with the other components.
The fibrocartilage analysis of the quadriceps tendon enthesis showed that the uncalcified fibrocartilage increases from medial to lateral. There was a greater quantity of uncalcified fibrocartilage centrally and laterally than medially. Fibrocartilage is found in areas of the body subject to high mechanical load (Benjamin et al. 1991). It can dissipate stress concentration at the bony interface (Benjamin et al. 1986; Woo & Buckwalter, 1988) by promoting a gradual bending of collagen fibres. It acts like a rubber grommet on an electrical plug, protecting the tendon from compression (Benjamin & Ralphs, 1998), and enables the highly directed forces in the tendon to be redistributed over a larger area. The degree of fibrocartilage differentiation in a tendon varies according to the distance from the bony surface against which the tendon is compressed (Merrilees & Flint, 1980). The finite element analyses of Giori et al. (1993) show that regions where fibrocartilage is present in the rabbit flexor digitorum profundus tendon correlate well with areas of high compressive stress. The quadriceps muscle group functions as a knee extensor when the leg is elevated. When the foot is on the ground, contraction of the quadriceps stabilizes the knee, functioning as a decelerator. The patella provides a significant mechanical advantage to the knee extensor mechanism, allowing the knee to extend with a smaller contractile force of the quadriceps. In addition, the patella redirects the force exerted by the quadriceps, resulting in a large compressive stress on the patellofemoral joint. According to our results, there is more fibrocartilage at the central and lateral parts of the quadriceps enthesis than medially, thus it is plausible to suggest that during the knee extensor mechanism the patella is more compressed centrally and laterally than medially. Then again, there is also a good correlation between the quantity of uncalcified fibrocartilage at an enthesis and the extent of movement that occurs between tendon and bone (Benjamin & Ralphs, 1998). Our results showed that VL and VI tendons (2.8 ± 0.5 cm) are longer than the VM tendon (1.1 ± 0.7 cm). This supposes a possible association between the amount of fibrocartilage and tendon length. An argument could be made that a longer tendon allows more range of movements, which in turn results in a greater amount of fibrocartilage. This could explain why more uncalcified fibrocartilage was found laterally and centrally than medially.
In a same context, Gao & Messner (1996) have suggested that the shape and surface area of the calcified fibrocartilage–bone interface at ligament insertions is determined by the tensile loads to which a ligament is subjected around puberty, but the thickness of its subchondral plate may respond to loads beyond that time and reflect motion at the hard–soft tissue interface. This is in line with the earlier suggestion of Evans et al. (1990) that differences in the thickness of the subchondral plate at the enthesis relate to regional variations in tensile loading. Furthermore, the thickness of the zone of calcified fibrocartilage and the extent of the interface that it provides for the bone may be related to the physiological strength and loading of the tendon or ligament (Pal & Routal, 1998; Fredericson & Yoon, 2006). In a recent study by Prescott et al. (2009), which was carried out on 103 patients average age between 51 and 71 years, it was found that both VI and VL have a higher volume compared with VM. Prior studies showed that a decrease in anatomical and physiological muscle cross-sectional area measured on magnetic resonance imaging is related to loss of strength (Chen et al. 2011). In the present study the specimen’s age was between 68 and 78 years, compared with the study mentioned above where the age was between 51 and 71 years. We can nevertheless hypothesize that if the differences between the VL, VM and VI could be mirrored in our population this could explain the difference in the quantity of bone at the medial, central and lateral parts of the quadriceps enthesis. Equally, we could suggest that a higher physiological strength for the VI and VL compared with the VM generates more mechanical stress at the central and lateral sides of the patella than at the medial part, which in turn would account for the observed differences in the structures of the fibrocartilage and the subchondral bone plate.
In conclusion, our study demonstrates that there is a variation in the trabecular architecture of the patella and in the structure of the proximal enthesis of the quadriceps tendon. Bone structure and the thickness of uncalcified fibrocartilage presented a similarity between the centre and the lateral parts; however, the medial side was different. We suggest that the mechanical stress at the proximal quadriceps tendon enthesis is higher laterally and centrally compared with medially. Consequently, the patella does not remain within the central groove of the femur (thigh bone), and the kneecap tends to tilt and glide towards the outside of the knee (subluxation). This is in line with the particular site of pathology at the lateral femoral condyle and the posterior medial surface of the patella. Differences in the physiological strength between the different components of the quadriceps muscles could be an option to explain the variations in the structure of the quadriceps tendon enthesis and the patella.
Acknowledgments
This study has been made in the course of the authors’ employment; no external or internal specific funds have been used, and all the authors contributed substantially to this work.
References
- Balachandar V, Twycross-Lewis R, Morrissey D. EMG mapping of the quadriceps in patellofemoral pain syndrome during functional activities: a pilot study. Br J Sports Med. 2011;45:A14–A15. [Google Scholar]
- Benjamin M, Evans EJ, Copp L. The histology of tendon attachments to bone in man. J Anat. 1986;149:89–100. [PMC free article] [PubMed] [Google Scholar]
- Benjamin M, Ralphs JR. Fibrocartilage in tendons and ligaments – an adaptation to compressive load. J Anat. 1998;193:481–494. doi: 10.1046/j.1469-7580.1998.19340481.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamin M, Tyers RN, Ralphs JR. Age-related changes in tendon fibrocartilage. J Anat. 1991;179:127–136. [PMC free article] [PubMed] [Google Scholar]
- Benjamin M, Newell RL, Evans EJ, et al. The structure of the insertions of the tendons of biceps brachii, triceps and brachialis in elderly dissecting room cadavers. J Anat. 1992;180:327–332. [PMC free article] [PubMed] [Google Scholar]
- Bolgla LA, Boling MC. An update for the conservative management of patellofemoral pain syndrome: a systematic review of the literature from 2000 to 2010. Int J Sports Phys Ther. 2011;6:112–125. [PMC free article] [PubMed] [Google Scholar]
- Chen BB, Shih TT, Hsu CY, et al. Thigh muscle volume predicted by anthropometric measurements and correlated with physical function in the older adults. J Nutr Health Aging. 2011;15:433–438. doi: 10.1007/s12603-010-0281-9. [DOI] [PubMed] [Google Scholar]
- Evans EJ, Benjamin M, Pemberton DJ. Fibrocartilage in the attachment zones of the quadriceps tendon and patellar ligament of man. J Anat. 1990;171:155–162. [PMC free article] [PubMed] [Google Scholar]
- Fredericson M, Yoon K. Physical examination and patellofemoral pain syndrome. Am J Phys Med Rehabil. 2006;85:234–243. doi: 10.1097/01.phm.0000200390.67408.f0. [DOI] [PubMed] [Google Scholar]
- Gao J, Messner K. Quantitative comparison of soft tissue–bone interface at chondral ligament insertions in the rabbit knee joint. J Anat. 1996;188:367–373. [PMC free article] [PubMed] [Google Scholar]
- Giori NJ, Beaupré GS, Carter DR. Cellular shape and pressure may mediate mechanical control of tissue composition in tendons. J Orthop Res. 1993;11:581–591. doi: 10.1002/jor.1100110413. [DOI] [PubMed] [Google Scholar]
- Koh TJ, Grabiner MD, De Swart RJ. In vivo tracking of the human patella. J Biomech. 1992;25:637–643. doi: 10.1016/0021-9290(92)90105-a. [DOI] [PubMed] [Google Scholar]
- Merrilees MJ, Flint MH. Ultrastructural study of tension and pressure zones in a rabbit flexor tendon. Am J Anat. 1980;157:87–106. doi: 10.1002/aja.1001570109. [DOI] [PubMed] [Google Scholar]
- Pal GP, Routal RV. Architecture of the cancellous bone of the human talus. Anat Rec. 1998;252:185–193. doi: 10.1002/(SICI)1097-0185(199810)252:2<185::AID-AR4>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- Prescott JW, Priddy M, Best TM, et al. An automated method to detect interstitial adipose tissue in thigh muscles for patients with osteoarthritis. Conf Proc IEEE Eng Med Biol Soc. 2009;2009:6360–6363. doi: 10.1109/IEMBS.2009.5333260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toumi H, Higashiyama I, Suzuki D, et al. Regional variations in human patellar trabecular architecture and the structure of the proximal patellar tendon enthesis. J Anat. 2006;208:47–57. doi: 10.1111/j.1469-7580.2006.00501.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo SL, Buckwalter JA. Injury and repair of the musculoskeletal soft tissues. J Orthop Res. 1988;6:907–931. doi: 10.1002/jor.1100060615. [DOI] [PubMed] [Google Scholar]