

Abstract
Ornaments, weapons and aggressive behaviours may evolve in female animals by mate choice and intrasexual competition for mating opportunities—the standard forms of sexual selection in males. However, a growing body of evidence suggests that selection tends to operate in different ways in males and females, with female traits more often mediating competition for ecological resources, rather than mate acquisition. Two main solutions have been proposed to accommodate this disparity. One is to expand the concept of sexual selection to include all mechanisms related to fecundity; another is to adopt an alternative conceptual framework—the theory of social selection—in which sexual selection is one component of a more general form of selection resulting from all social interactions. In this study, we summarize the history of the debate about female ornaments and weapons, and discuss potential resolutions. We review the components of fitness driving ornamentation in a wide range of systems, and show that selection often falls outside the limits of traditional sexual selection theory, particularly in females. We conclude that the evolution of these traits in both sexes is best understood within the unifying framework of social selection.
Keywords: females, intrasexual competition, mate choice, ornamentation, social competition, weapons
1. Introduction
Explaining the evolution of exaggerated ornamentation and weaponry (hereafter ‘ornamental traits’) in male animals has preoccupied biologists since Darwin [1] published his ideas on selection in relation to sex. Darwin realized these traits must often arise via socially mediated mechanisms because they appear to impose costs without delivering any survival benefits. The central mechanisms he proposed—male–male competition and female choice—remain the foundation of sexual selection theory today. However, because Darwin's ideas were framed around competitive processes in males, they leave many questions unanswered about the nature of selection in females.
Recent research has increasingly focused on addressing this issue (reviewed in [2–6]), but instead of clarifying how selection operates within and between the sexes, the discussion has mainly generated controversy. In particular, the analysis of selection on females has highlighted a growing uncertainty about whether classical sexual selection can explain the evolution of ornamental traits in both sexes. This debate has major implications for our understanding of general patterns in biology, such as sexual dimorphism, sexual dichromatism and the expression of ornamental traits in both males and females of a species (e.g. mutual ornamentation).
The evolution of ornamental traits in males is almost universally ascribed to sexual selection, presumably because male–male competition for sexual resources is usually so overt. By contrast, similar traits expressed in females have been explained in three different ways. The original view, favoured by Darwin [1], is that ornamental traits in females arise from selection acting on males and are simply the result of correlated inheritance—what we might today call ‘shared genetic architecture’. The idea that exaggerated phenotypes are therefore adaptive in males but non-adaptive in females resurfaced in the genetic correlation hypothesis developed by Lande [7]. The main counter-argument is that such phenotypes are adaptive in females, arising through female–female competition for mates and breeding opportunities, as well as by male choice [2]. According to this second view, females—particularly those in sex-role-reversed and polyandrous species—are subject to forms of sexual selection analogous to those postulated to shape trait evolution in males. A third alternative is that females compete strongly for ecological resources rather than matings, and that female traits are therefore shaped by evolutionary forces that extend beyond the usual concept of sexual selection [6,8,9].
These three viewpoints have each received empirical and theoretical support. First, it makes sense that genetic correlation explains some degree of trait exaggeration in females given that much of the female genome is shared with conspecific males [4,10]. Second, numerous studies indicate that females gain fitness by deterring sexual rivals and attracting mates, indicating that ornamental trait evolution in females can be at least partly explained by sexual selection [2,5]. Third, females in many species clearly use ornamental traits during competition for non-sexual resources, implying that sexual selection may not be the only route to ornamental trait evolution [4–6,11,12]. There is currently no consensus about which of these explanations plays a dominant role.
The purpose of this review is to examine different types of selection on ornamental traits, and to document their prevalence in females. Female traits in polyandrous and sex-role-reversed species are only covered briefly as they are clearly sexually selected and are considered in detail elsewhere. We begin by addressing disagreements about the meaning of key terms, and then we move beyond semantics to ask whether competition for resources related to fecundity and survival can help us to explain patterns of ornamentation and weaponry in both sexes. Our main goal is to critically appraise existing frameworks for understanding selection on ornamental traits, with a particular focus on the theories of sexual selection and social selection [12,13].
2. What is sexual selection?
In The origin of species [14], Darwin gave a brief overview of sexual selection, stating that it depends ‘not on a struggle for existence, but on a struggle between the males for possession of the females; the result is not death to the unsuccessful competitor, but few or no offspring’ (p. 156). The emphasis here was on matings, although in The descent of man [1] Darwin later spoke of ‘the advantage which certain individuals have over others of the same sex and species, in exclusive relation to reproduction’ (p. 256). This latter phrase has often been presented as the original definition of sexual selection [15,16], and it can be interpreted as covering any aspect of breeding behaviour. However, it is taken out of context, and the rest of Darwin's detailed discussion suggests that he was always thinking more narrowly about fitness related directly to matings.
Although sexual selection began as a neatly circumscribed idea, its conceptual boundaries have been the subject of debate ever since [5,15,17]. To some extent, contrasting viewpoints have arisen because different authors have focused on different aspects of evolution, including phenotypic traits, components of fitness and general mechanisms of competition. This has led to confusion about whether contexts other than competition for matings can be included under the rubric of sexual selection. Although this debate is primarily semantic, it raises important questions about how we conceptualize evolutionary processes, and how we interpret the findings of empirical studies [15]. One of the main points of controversy relates to the array of social mechanisms influencing ornamental traits, and whether it is so broad that a more general theory than sexual selection is required to account for patterns of phenotypic evolution. Such questions have resurfaced periodically over the last century, generally inspired by conflicting views about how selection operates on ornamental traits in females [2–6,8,12,13,18–21].
3. The crux of the problem: understanding selection in females
Owing to differences between the sexes in reproductive constraints [22,23], competition among females does not map easily onto the classic Darwinian view of sexual selection. As noted by Rosvall [5], ‘a central tenet of mating systems and sexual selection theory is that the sexes differ fundamentally in their route to reproductive success, with male reproductive skew based on competition for mates, and female reproductive skew based on access to resources that affect fecundity’. There is no doubt that females tend to compete with both males and females during the course of reproduction—often using ornaments, weapons and aggressive behaviours—but this competition may often target ecological and social resources, including foraging territories, nest sites and the quality of paternal care, rather than matings [8,9,21,24]. Thus, according to Darwin's theoretical formulation, ornamental traits have been favoured by sexual selection in males, but not in females.
One proposed solution has been to broaden the scope of sexual selection to include ‘all selection processes operating through intrasexual competition for breeding opportunities in either sex’ [3]. There are, however, at least three problems with this approach. First, the conceptual connections between sexual selection, ornamental traits and fitness via matings is so deeply entrenched that processes applying to males are often assumed to apply to females, usually without actually conducting the difficult work of understanding the mechanisms involved. Thus, a large number of recent studies cast female competition in the context of sexual selection, even though fitness components have not been thoroughly studied [25–27]. Second, by seeking a label that applies equally to males and females, and yet makes specific assumptions about fitness components, we risk obscuring some of the important differences between the sexes with respect to the underlying drivers of evolution. Finally, many ornamental traits are used in competition for resources that are not directly linked to reproduction, suggesting that the concept of sexual selection would have to stretch far beyond ‘breeding opportunities’ to capture all of the relevant processes.
By good fortune, we already have a perfectly adequate conceptual framework for dealing with all forms of social competition, including those operating in females: West-Eberhard's [12,13] social selection theory, first elaborated in 1979. This body of theory is hardly an unknown quantity as several previous authors have understood its implications and discussed its potential importance in some detail [4,6,28–30]. Nonetheless, the core ideas are still generally overlooked in studies of phenotypic evolution, even when the bigger picture is considered [31], or when the patterns investigated clearly point towards social competition for resources other than mates [32,33]. Some authors have suggested that the framework of social selection has been ignored owing to a deep-rooted historical preoccupation with sexual selection in its narrow sense [6]; others have warned that adopting the framework of social selection is ‘likely to generate more problems that it solves’ [3]. Overall, there appears to be much confusion over what exactly the term ‘social selection’ means, exacerbated by its unfortunate use to characterize largely unrelated ideas in a recent critique of sexual selection [34,35].
4. Social selection: what it is, and what it is not
In the second half of the twentieth century, researchers began to note parallels between standard sexual selection and selection exerted in other competitive contexts [20,36,37]. By building on these studies, West-Eberhard [12] proposed a theory of social selection, defining the central mechanism as ‘differential reproductive success (ultimately, differential gene replication) due to differential success in social competition, whatever the resource at stake’. Thus, social selection includes sexual selection as ‘the subset of social competition in which the resource at stake is mates’ [12]. It should be emphasized that ‘reproductive success’ is measured in this instance by the contribution of genes to the next generation, and may be the result of improved survival or better access to food in the non-breeding season, rather than the result of any resource associated explicitly with mates or mating opportunities.
Modern usage of the term ‘social selection’ has tended to emphasize this focus on non-sexual contexts, perhaps leading to the widespread misconception that the term specifically relates to the non-sexual category of social interactions. However, the original definition makes clear that social selection theory focuses on social interactions in their broadest sense, including any social context driving selection, whether it be sexual or non-sexual. In effect, the most accurate approach would be to partition social mechanisms into ‘sexual social selection’ and ‘non-sexual social selection’, but these terms are probably too cumbersome to be widely adopted. In the rest of this study, we use ‘sexual selection’ for the former and ‘social selection’ for the latter, while recognizing that the theory of social selection covers both processes (figure 1).
Figure 1.

Nested conceptual framework showing sexual selection (red) as a subset of social selection (blue), and social selection as a subset of natural selection (green). Although each concept is illustrated with a sharp boundary for convenience, different colours should be viewed as regions of a continuum. Category codes (listed to the right and further defined in table 1) are mapped onto this framework to show the function of ornamental traits reported in the literature for males and females. Code placement is subjective, based on our assessment of the strength of evidence for sexual, social or natural selection (see text). Unequivocal evidence for sexual selection is restricted to the red zone. Code size is scaled to the proportion of species in each category (n = 77). Data are potentially biased as they are based on previous studies, rather than a random sampling of species. Species are only included if traits have been studied in both sexes.
West-Eberhard [12] noted that social competition for non-sexual resources often involves the same sorts of traits produced by sexual selection—costly signals, conspicuous displays, weaponry and aggressive behaviours. Recent theoretical work confirms that sexual selection is not a prerequisite for the evolution of these traits: they will be favoured whenever they increase the fitness of the bearer by improving access to limiting resources via social interactions [30,38–40]. The underlying mechanisms of social selection are also similar to those associated with sexual selection, as competition and choice operate in contexts other than reproduction [41].
The most widespread social mechanism influencing ornamental trait evolution is direct competition. In many migratory bird species that flock in winter, individuals of both sexes resolve conflicts over food with plumage signals that enable them to determine fighting ability and establish social dominance without direct physical contests [42–44]. Many other species use vocal [9] or visual [24] signals to defend foraging territories during the non-breeding season. In all cases, the traits mediating competition have the same proximate function as traits used in sexual selection, i.e. they convey information about quality and condition, and thereby influence the outcome of interactions.
The most clear-cut contexts of social competition occur outside the breeding season, but many also apply to reproduction. West-Eberhard [12] drew attention to several forms of female reproductive competition that were not about mates, including pheromonal and aggressive control of reproduction in socially breeding taxa. She also noted that ornamental traits in females are often socially selected signals functioning in the defence of critical resources such as breeding territories and nest sites, and that there is ‘a correlation between monomorphically bright signal coloration and territoriality by both sexes’. In these cases of long-term social coalitions, it is possible that mate choice itself is partly driven by non-sexual forms of social selection as the pay-off for choosing ornamented partners can include the successful year-round defence of ecological resources.
As Darwin did before her, West-Eberhard [12] considered choice mechanisms to be forms of indirect competition, whereby one class of individuals competes to be chosen by another class of individuals. However, she was the first to point out that choice often extends beyond mating preferences, for example, in the case of parental choice, an indisputable mechanism of social selection. Parents are often in a position to show favouritism among their offspring through the allocation of resources that are critical to survival, such as food. If parents preferentially feed offspring that are adorned with a particularly conspicuous signal trait, and if such feeding enhances offspring fitness, then parental choice should drive elaboration of the offspring's signal [45]. The details of this process have been confirmed by experiments [46], and are implicit in much of the literature on begging signals used by offspring, and the offspring-provisioning rules used by parents [47].
5. Darwin's legacy—why we tend to link ornaments to matings
If social selection theory offers a broad explanation for the evolution of ornamental traits in all contexts from infancy to adulthood, then it is worth pausing to consider why Darwin attempted to explain such traits only in adults and only with respect to social competition for matings. West-Eberhard [12] suggested one possible answer: that Darwin emphasized survival as the driver of natural selection, and thus his theory of sexual selection [1] was designed specifically to deal with the evolution of conspicuous traits that are unlikely to arise simply because of the ‘struggle for existence’. He was therefore motivated to emphasize the sharpest contrasts between natural and sexual selection, as any consideration of less clear-cut forms of selection may have weakened his arguments.
A careful reading of The origin of species [14] and The descent of man [48] suggests that another factor may have been important. When the idea of sexual selection was first outlined in The origin of species, Darwin's focus was entirely on differences between the sexes in ‘structure, colour or ornament’. Thus, his concept of sexual selection was chiefly inspired by sexual dimorphism in secondary sexual characteristics [49]. He explored this topic at length in The descent of man, where he focused mainly on species with extreme sexual dimorphism—such as birds of paradise, pheasants and other lekking or highly polygynous species—perhaps because they provided the clearest support for his theory. Given that male fitness in those taxa is limited by access to females, it was logical for Darwin to frame sexual selection in terms of male competition for matings.
The subsequent disagreement between Darwin and Wallace about the evolution of sexual dimorphism sheds further light on the preconceptions of both men about selection in females [4,21]. Whereas Darwin maintained that dimorphism was due to sexual selection restricted to males, Wallace argued that it could be explained by natural selection for crypsis in females [50]. He supported this counter-theory with comparative data from birds showing that dimorphic species were more likely to have open nest types, while species with highly ornamented females tended to have concealed nests (and thus lower predation on females). Darwin's main rebuttal was that colourful females arise via sexual selection on males, after which natural selection then acts on nesting behaviour such that species with conspicuous females are more likely to build concealed nests. This explanation seems rather convoluted today, and it is telling that neither man ever seriously considered the possibility of widespread selection for ornamentation in females. Darwin ([48], p. 276) even described a theory similar to the modern view of mutual sexual selection [51,52], then dismissed it as far-fetched, on the assumption that males will always be more limited by mates (i.e. ‘more eager’) than females.
Overall, we believe that Darwin's focus on sexual dimorphism over-emphasized competition for matings as a general explanation for trait evolution, with important consequences for his treatment of the evidence. The legacy of his approach can be seen in current studies of ornament evolution, many of which still focus entirely on components of fitness linked to matings or fertilizations [15,53–56]. Adherence to this relatively narrow framework has had a huge impact on decades of biological research, with numerous studies overlooking the role of alternative social contexts. In §6, we review the literature to assess the function of ornamental traits in both sexes, and the validity of sexual selection and social selection as frameworks for understanding trait evolution.
6. Review of empirical studies
Several strands of evidence support Darwin's suggestion that female ornaments arise via two routes: either through standard sexual selection in sex-role-reversed species or else through genetic correlation, i.e. correlated evolution in females driven by selection restricted to males. Role-reversed pipefish (Syngnathus typhle) provide an example of the first route, as males prefer ornamented females [57]. Similarly, in two-spotted gobies (Gobiusculus flavescens), the preference of males for bright females [58] is best explained by dynamic sex roles rather than by mutual sexual selection, as females sometimes compete strongly for males [59]. Genetic correlation is more difficult to demonstrate, but theoretical models do predict that the elaboration of female traits in species with traditional sex roles can be driven by evolutionary processes in males, at least until natural selection corrects the trend by favouring less costly female phenotypes [7,10]. This tallies with several studies that find no evidence that male-like traits are adaptive in females [60–62].
So far, so good, for Darwin's worldview, but the picture changes markedly in the light of recent evidence that adaptive female ornamentation occurs in more than just role-reversed species. The most obvious examples are group-living animals with traditional sex roles and complex mating systems, wherein ornamented females are forced to compete against each other for mating opportunities or other resources, such as access to paternal care [3,63–66]. Thus, fertile female alpine accentors (Prunella collaris)—a polygynandrous passerine bird—apparently sing to attract males [67]. Similarly, in African starlings (Sturnidae), female–female competition over breeding status is thought to explain why females of group-living species have bright plumage similar to males, whereas females of socially monogamous species tend to be contrastingly dull [68].
Complex social systems, however, are not the key to female ornamentation because ornaments are expressed in both sexes in many socially monogamous species [6]. Again, ideas have changed significantly since Darwin's day, as it is now clear that such cases of monomorphic ornamentation can evolve via mutual mate choice [69–72]. Recent studies in a range of species have shown that males choose females on the basis of ornaments, and vice versa [52,73,74]. This reciprocal mechanism has generally been interpreted as evidence of sexual selection acting on both sexes, but it does not rule out a major role for social selection in driving ornamentation. It could be argued that mates are chosen on the basis of socially mediated badges of status, and that the main drivers of selection are pay-offs related to survival or fecundity. In addition, mutual ornaments may function in intrasexual competition for ecological resources, particularly in females.
(a). Competition for ecological resources
In theory, females maximize their own fecundity rather than their access to matings [22,23], and thus are more likely than males to compete directly or indirectly for the ecological resources needed to raise their offspring [2,3]. The greater value of ecological resources to the reproductive fitness of females—and of matings to the reproductive fitness of males—may have important implications for the dynamics of competition [75,76]. It may also underlie fundamental differences between the sexes in the mechanisms of selection promoting trait evolution.
These differences are apparent even when both sexes compete for mates, which typically happens before pairing in males and after pairing in females. Paired females in many species aggressively deter rival females, presumably to defend the non-shareable portion of their mate's parental care and therefore to maximize food for their offspring [77–80]. Such competition over paternal care can involve ornamental traits—for example, plumage signals of female great tits (Parus major) signal social dominance in contests between rival females on breeding territories [81], whereas those of males influence female choice [82]. Likewise, female Hypocnemis antbirds use song to defend their position in long-term monogamous partnerships by repelling unpaired rivals from their territory [83], and jamming their mate's advertising signal [84]. These examples of female–female competition for paternal care are usually considered forms of sexual selection, but we emphasize that they are quite distinct from Darwin's original idea.
In other taxa, female fecundity is less influenced by paternal care than by direct access to food, leading to striking differences between the sexes in the fitness contexts of selection on ornamental traits. Both male and female dung beetles (Onthophagus sagittarius) use horns to compete intrasexually for food resources, but males use the food to attract mates, whereas females use it to raise offspring [85,86]. Similarly, facial markings mediate social dominance interactions in both sexes of social paper wasps (Polistes), but males compete for foraging territories visited by females who come to mate, whereas females (queens) compete for suitable sites for building colonies [87–89]. In neither beetles nor wasps do males contribute to the raising of offspring, suggesting that ornamental traits in males are sexually selected, whereas those in females function entirely in female–female competition for ecological resources.
Similar disparities in selective mechanisms abound in mammals. In reindeer (Rangifer rangifer), male antlers are used in contests for matings, whereas female antlers function in aggressive contests after calving, when they fight for access to prime foraging sites [90]. In Soay sheep (Ovis aries), the horns of males are used to gain access to mates, whereas those of females are to contest food during the lambing season [91]. Indeed, the most common forms of female–female territorial competition in ungulates involve the defence of offspring from infanticidal females, and the defence of food for offspring long after matings have occurred [92]. Comparative analyses provide further evidence that female weapons have evolved in this diverse group via competition for ecological resources associated with fecundity, rather than for mates [11].
The contrast between the sexes is also seen in many other mammalian clades, including rodents, pinnipeds and primates. In the southern flying squirrel (Glaucomys volans), for example, reverse sexual size dimorphism is related to female-only defence of foraging areas, such that ‘Females are clearly competing for the resources contained in the territories rather than for males’ [93]. Male elephant seals (Mirounga angustirostris) famously fight for access to females, whereas female–female competition at breeding colonies is focused on defending space for pups [94]. In chimpanzees (Pan troglodytes), female aggression and dominance is mostly related to competition for space and core foraging areas [95,96], whereas in chacma baboons (Papio ursinus) female aggression is used to achieve central positions in groups—where the foraging is good and predation risk is reduced ([97], but see [98]).
There is a huge literature dealing with the greater significance of ecological resources to female mammals, much of which relates to the evolution of social systems [99]. Females typically compete—among one another, as well as with males—for a broad array of such resources, often using weapons and aggressive behaviours (reviewed in [100,101]). Fitting the evolution of mammalian traits into the framework of sexual selection thus requires a major extension of Darwin's central proposal of competition for matings, at least where females are concerned.
Birds pose a similar challenge. Plumage signals in male starlings (Sturnus vulgaris) are shaped by female choice [102], whereas the same sorts of signals in females mediate dominance in interactions over food during the breeding season [103]. Likewise, bill colour in male American goldfinches (Spinus tristis) is influenced by female mating preferences [104], whereas female bill colour is a status signal during competition over food in the breeding season [105]. Moreover, females in numerous species use song mainly to defend breeding territories or to repel female rivals rather than to attract mates [106–109]. Where the limiting resource is a nest site, female–female competition is often particularly intense, involving high levels of aggression or extravagant traits [8,110–112].
In all these cases, the fitness component driving selection in females is not related to the acquisition of mates, yet potentially involves some aspect of reproduction linked to fecundity. However, signal traits in some female birds also mediate female competition outside the breeding season, and here the link with fecundity is much more tenuous. In numerous species, females compete for space and foraging territories year-round, including the non-breeding season [9,24,113,114]. In some species, such as Townsend's solitaire (Myadestes townsendi) [115] and European robin (Erithacus rubecula) [116], females sing only to defend solitary non-breeding territories against both males and females, and never sing during the breeding season. In a similar fashion, food-provisioning experiments demonstrate that plumage signals mediate female dominance in some flocking bird species even in the non-breeding season [117–119].
(b). Ecological competition and mutual ornamentation
The most widespread context of female ornamentation is when both sexes have similar phenotypes. In birds, this seems to occur mainly when males and females have similar roles in reproduction: biparental care coupled with cooperative defence of territories or breeding sites. Many such cases involve socially monogamous species with low rates of extra-pair paternity, so it seems likely that mate quality or parental investment patterns—rather than the number of mates—may drive selection on ornaments [51]. On the other hand, the dominant source of selection may be from between-pair rather than within-pair interactions, as mutual ornamentation often goes hand-in-hand with year-round territoriality [9,12]. The potential role of ecological competition in mutual ornamentation is highlighted by the fact that those socially monogamous passerine bird species in which females play no role in territory defence tend to be either monomorphically dull, or sexually dichromatic with relatively dull females.
Mutual ornaments involved in mate choice can also mediate competition in contexts far removed from breeding events. For example, plumage ornaments in the Gouldian finch (Erythrura gouldiae) function in mutual sexual selection [120], but also mediate social dominance and competition over food between females outside the breeding season [117]. Similarly, plumage ornaments in female rock sparrows (Petronia petronia) function in mate attraction [121,122] but also mediate intra-sexual competition for food during the non-breeding season [119]. In these cases, traits originally thought to evolve by mutual sexual selection are potentially influenced by other social mechanisms.
The evolution of mutual ornamentation in both sexual and non-sexual contexts makes sense from a physiological perspective. Previous studies reveal that female ornaments often correlate with measures of condition, such as immunocompetence, parasite load and egg mass [123–125]. Such ornamentation is also related to underlying levels of circulating testosterone [27,126], even fluctuating over short timeframes [127]. The link with testosterone is particularly enlightening as it helps to explain why the expression of female ornaments so often predicts dominance in contests [24,117,128–130]. In short, female ornaments are similar to most male ornaments in that they function as honest signals of condition, status and fighting ability. The main difference is that female traits may be under weaker selection from mate choice, and stronger selection via competition for ecological resources.
(c). Sociality
Species that live in groups also frequently display mutual ornamentation or weapons. As with socially monogamous systems, sexual selection is often assumed to be a driving force of ornamental trait evolution in more complex societies, yet there is often little evidence that males choose females, or that matings are limiting for females [2,66]. In some cases, exaggerated female traits may evolve via competition among females for reproductive dominance or breeding opportunities, i.e. competition over who gets to breed [68,131]. However, assigning these cases of fecundity-based selection to sexual selection is not always as straightforward as it first appears. To understand why, it is worth considering an extreme example: all-female social insect colonies in which females compete over reproduction would be considered hotbeds of sexual selection were it not for the fact that no males are involved [12,89]. Thus, female–female competition within social groups may fit more neatly into the framework of social selection.
Ornamental traits in group-living animals may be used in interactions with neighbours, and the most influential social dynamics may be between groups rather than within groups. Current evidence suggests that group-living females are under socially mediated selection to contribute to cooperative defence of ecological resources such as territories, e.g. the roaring of female lions (Panthera leo) [132], and the mixed-sex choruses of birds such as mesites (Mesitornithidae) [133] and trumpeters (Psophidae) [134]. In all these group-territorial systems, cooperative defence of ecological resources may be a major driver of ornamental trait evolution in females [12]. Again, we suspect that social selection theory may provide a more general explanation for evolutionary patterns of ornamental traits in group-living animals.
(d). Other assorted mechanisms and their implication for selection in females
Several other ideas have been forwarded to explain female ornamentation, but none of these is completely encompassed by the traditional view of sexual selection. The good-parent hypothesis, for example, was originally proposed as a mechanism for honest signalling in males [135]. Although there is some evidence that females choose males on the basis of their quality as parents [136], and that paternal quality may be reflected in male ornaments [137,138], there is little support for the idea that the same process operates in reverse, selecting for ornaments in females [119,139]. Some correlational studies suggest that mutual choice of mates that are good parents may be mediated by ornaments in socially monogamous species [140] but this needs experimental corroboration.
Another possibility is that ornaments reduce sexual harassment for females. This has been proposed for reptiles because some female lizards (e.g. Holbrookia maculate [141]) are relatively dull when attracting males but bright after copulation. The mechanism driving these inverse patterns has not yet been identified, but it is thought that such post-copulatory signals may reduce costly harassment by advertising that the females are no longer fertilizable [142,143]. Sexual mimicry of males by females is thought to be adaptive for similar reasons, at least in polymorphic insects [144]. In the damselfly Ischnura ramburi, for example, some females express vivid male-like coloration, and as a result attract a lower number of copulation attempts than dull-coloured females [145]. Given that copulations last up to 3 h in this species, increasing the risk of predation, ornamented females may benefit from conserving time and energy, as well as reducing predation risk [145]. In all such cases, the fitness component could be classified as representing either natural selection (as the real pay-off may be foraging efficiency and survival rather than matings) or sexual conflict. Note that sexual conflict involves a number of mechanisms that extend far beyond Darwin's original concept of sexual selection [146], but all of them fall neatly within the broader framework of social selection.
Finally, some cases of apparent mutual ornamentation may also be driven by natural selection. For example, in the dart poison frog Dendrobates pumilio, the anti-predator function of a bright aposematic signal is shared by both sexes [147], but that same signal serves as a sexual ornament in males but not females [148]. Similarly, the elaborate tail of the turquoise-browed motmot (Eumomota superciliosa) functions in pursuit-deterrent anti-predator displays in both sexes [149], but appears to be sexually selected only in males [150]. These examples suggest that exaggerated traits shaped primarily by natural selection can be co-opted as sexual ornaments, or vice versa. Either way, the sexual function is restricted to males, whereas the exaggerated trait appears to be naturally selected in females.
(e). Mechanisms of selection in males and females
We have argued that selection on ornamental traits in females takes many forms, and often contrasts with selection on similar traits in males. To gain a better understanding of these patterns, and the relative importance of different processes, we gathered data on the function of ornamental traits (table 1). Because we are interested in comparisons between the sexes, we restricted our search to species with information from both males and females. Thus, a large number of studies focusing on male-only (or female-only) sexual selection are omitted from this compilation.
Table 1.
Aspects of fitness thought to drive selection on ornamental traits in males and females.
| taxon | species | phenotypic trait | fitness component in femalesa | fitness component in malesa | emphasis on ecological competitionb | source |
|---|---|---|---|---|---|---|
| bird | Aethia cristatella | crest size | M, NS | M, NS | F = M | Jones & Hunter [52], Jones & Hunter [151] |
| bird | Aethia pusilla | overall plumage colour | SD | SD | F = M | Jones [152] |
| bird | Agelaius phoeniceus | epaulets | NF | BT | U | Johnsen et al. [153] |
| bird | Aimophila ruficauda | vocalization rate | BT, NBT | BT, NBT | F > Mc | Illes & Yunes-Jimenez [109] |
| bird | Aptenodytes patagonicus | colour of plumage patch | NS | M, NS | F > M | Dobson et al. [154] |
| bird | Bubulcus ibis | body plume length | GRC, M | GRC, M | F = M | Krebs and Putland [45] |
| bird | Calonectris diomedia | vocalization | M, NS | M, NS | F = M | Bretagnolle & Lequette [155], Bretagnolle [156] |
| bird | Coracias garrulus | overall plumage colour | M | M | F = M | Silva et al. [157] |
| bird | Cyanistes cyaneus | colour of plumage | BT, FNR, FR, M, PC | BT, FNR, FR, M | F = M | Kempenaers [78], Hunt et al. [158], Remy et al. [159], Vedder et al. [160], Midamegbe et al. [129] |
| bird | Cygnus atratus | wing plumage | SD | SD | F = M | Kraaijeveld et al. [161] |
| bird | Eclectus roratus | overall plumage colour | NS | M | F > M | Heinsohn et al. [111] |
| bird | Erithacus rubecula | vocalization | NBT | BT, M | F > M | Kriner & Schwabl [126], Tobias [116] |
| bird | Erythrura gouldiae | colour of plumage patch | FNR, M, SD | FNR, M, SD | F = M | Pryke & Griffith [162], Pryke [117], Pryke & Griffith [120] |
| bird | Eumomota superciliosa | tail ornament | AP | BS, M, RS | F > M | Murphy [149,150] |
| bird | Ficedula hypoleuca | wing patch size, aggression | BT, FR, PC | BT, M | F > M | Garamszegi et al. [163], Hegyi et al. [164], De Heij et al. [165] |
| bird | Fulica americana | body size | RS | U | F > M | Lyon [166] |
| bird | Gallinago media | aggression, vocalization | M | M | F = Md | Sæther et al. [167], Sæther et al. [168] |
| bird | Gallinula chloropus | aggression, body size | M, RS | U | U | Petrie [169,170] |
| bird | Gallinula tenebrosa | frontal shield | SD | SD | F = M | Crowley & Magrath [130] |
| bird | Gymnorhinus cyanocephalus | colour of plumage patch | FR, NS, SD | FR, M, SD | F > M | Johnson [128,171] |
| bird | Hirundo rustica | tail streamer length | Ue | B, M | U | Møller [172], Cuervo et al. [61], Møller et al. [173] |
| bird | Hylophylax naeivioides | vocalization, aggression, colour of plumage patch | BT, NBT | BT, NBT | F < M | Hau et al. [114] |
| bird | Hypocnemis peruviana | vocalization | BS, BT, M, NBT | BS, BT, M, NBT | F > M | Seddon & Tobias [83], Tobias & Seddon [84,174], Tobias et al. [9] |
| bird | Icteria virens | aggression | BT, PC? | BT, M | F > M | Mays & Hopper [175] |
| bird | Icterus pustulatus | overall plumage colour | BT, NBT | BT, NBT | F > Mc | Price et al. [108], Murphy et al. [24] |
| bird | Larosterna inca | facial plumes | BS, GRC | BS, GRC | F = M | Velando et al. [26] |
| bird | Luscinia svecica | colour of plumage patch | M | M, GRC | F = M | Amundsen et al. [73], Johnsen et al. [176] |
| bird | Malurus cyaneus | vocalization | BT, NBT | BT, NBT, M | F > M | Cooney & Cockburn [107] |
| bird | Melospiza melodia | aggression | FNR | FNR | F = M | Arcese & Smith [177] |
| bird | Myadestes townsendi | vocalization | NBT | BT, M, NBT | F > M | George [115] |
| bird | Myrmeciza longipes | vocalization | BT, NBT | BT, NBT | F = M | Fedy & Stutchbury [113] |
| bird | Pachyptila belcheri | vocalization | M, NS | M, NS | F = M | Bretagnolle [156], Bretagnolle et al. [178] |
| bird | Parus major | overall plumage colour, aggression | SD, FNR, FR | SD, FNR, M | F > M | Norris [82], Slagsvold [81], Jarvi & Bakken [179] |
| bird | Petronia petronia | colour of plumage patch | BS, FR, M | BS, FR, M | F = M | Griggio et al. [119,121,122,180] |
| bird | Phalacrocorax aristotelis | crest size | BS, M, GRC | BS, M, GRC | F = M | Daunt et al. [25] |
| bird | Prunella collaris | vocalization | M | BT, M | F < M | Langmore et al. [67] |
| bird | Sialia sialis | overall plumage colour, aggression | FR, NS | FR, M | F > M | Gowaty [181], Siefferman & Hill [182] |
| bird | Spinus tristis | bill colour | FR | M | F > M | Murphy et al. [105], Johnson et al. [104] |
| bird | Stagonopleura guttata | flank plumage patterng | FNR, FR | M | F > M | Crowhurst et al. [118] |
| bird | Sturnus vulgaris | chest plumage colour | FR | M | F > M | Swaddle & Witter [103], Bennett et al. [102] |
| bird | Sula nebouxii | foot colour | M | M | F = M | Torres & Velando [74] |
| bird | Tachycineta bicolor | aggression | NS | M | F > M | Rosvall [110] |
| bird | Turdus migratorius | overall plumage colour | M, RS | M, RS | F = M | Rowe & Weatherhead [183] |
| bird | Tyto alba | plumage patterns | M | M | F = M | Roulin [184], Roulin et al. [185] |
| bird | Vanellus vanellus | aggression | GRC | U | U | Liker & Szekely [77] |
| bird | Zonotrichia atricapilla | overall plumage colour | FNR | FNR | F = M | Chaine et al. [186] |
| bird | Zonotrichia querula | overall plumage colour | FNR | U | F > M | Watt [187] |
| mammal | Glaucomys volans | aggression | BT, NBT | M | F > M | Madden [93] |
| mammal | Mirounga angustirostris | aggression | BT | M | F > M | Christenson & Leboeuf [94] |
| mammal | Oreotragus oreotragus | scent mark | BT, M, NBT | BT, M, NBT | F > M | Roberts & Dunbar [188] |
| mammal | Ovis aries | horns | FR | M | F > M | Robinson & Kruuk [91] |
| mammal | Pan troglodytes | aggression | FNR, FR | M | F > M | Kahlenberg et al. [95] |
| mammal | Papio ursinus | aggression | AP, SD | M | F > M | Ron et al. [97], Huchard & Cowlinshaw [98] |
| mammal | Rangifer tarandus | horns | FR | M | F > M | Espmark [90] |
| fish | Gobiusculus flavescens | colour patchf | M | M | F = M | Amundsen & Forsgren [58], Forsgren et al. [59] |
| fish | Knipowitschia panizzae | colour | M, RS | U | U | Pizzolon et al. [189] |
| fish | Lamprologus ocellatus | aggression, body size | BT, GRC | U | U | Brandtmann et al. [190] |
| fish | Paragobiodon xanthosomus | aggression | FR, PC | U | U | Wong et al. [191] |
| fish | Pelvicachromis taeniatus | colour patches | M, PC | M, PC | F = M | Baldauf et al. [192] |
| amphibian | Dendrobates auratus | aggression | PC | M | F > M | Summers [80] |
| amphibian | Dendrobates pumilio | colour pattern | AP | AP, M | F > M | Daly & Myers [147], Summers et al. [148] |
| reptile | Chamaeleo chamaeleon | colour | AH?, RS | Signal absent | U | Cuadrado [193] |
| reptile | Crotophytus collaris | colour, aggression | BS, FR | M | F > M | Baird & Sloan [194], Baird [195], Lappin & Husak [196] |
| reptile | Ctenophorous maculosus | colour | AH, BS | Signal absent | U | Chan et al. [142] |
| reptile | Ctenophorus ornatus | colour | M, | U | U | LeBas & Marshall [143] |
| reptile | Holbrookia propinqua | colour | AH, BS | U | Ug | Cooper & Crews [197], Hager [141] |
| reptile | Microlophus occipitalis | colour | AH, BS | Signal absent | Ug | Watkins [198] |
| reptile | Sceloporus virgatus | colour | M | BT, M | F < M | Weiss [199] |
| reptile | Uta stansburiana | tail | SD | U | F > M | Fox et al. [200] |
| insect | Cyrtodiopsis dalmanni | eyestalk length | NF | FNR, FR, M | F < M | Panhuis & Wilkinson [201], Al-kairulla et al. [202] |
| insect | Drosophila melanogaster | aggression | FNR, FR | M, FNR, FR | F > M | Nilsen et al. [76], Vrontou et al. [203] |
| insect | Drosophila serrata | cuticular hydrocarbons | M | M | F = M | Chenoweth & Blows [63] |
| insect | Gryllus campestris | aggression | M, PC | U | U | Rillich et al. [204] |
| insect | Onthophagus sagittarius | horns | FR | FR, M | F > M | Watson & Simmons [85,86] |
| insect | Polistes exclamans | facial pattern | FR, NS | M | F > M | Lin [85], Thornhill & Alcock [89], Tibbetts & Sheehan [87] |
| insect | Rhamphomyia longicauda | ornamented wings, tibia | M | M | F = M | Bussiere et al. [205] |
| insect | Sabethes cyaneus | thoracic appendage size | M | U | F = M | South & Arnqvist [206] |
aAspects of fitness: AH, anti-harassment; AP, anti-predator; BS, breeding status (e.g. stage of reproduction, perhaps to minimize harassment); BT, breeding territory; FNR, food, but not related to reproduction; FR, food for reproduction/offspring; M, matings (including suppression of reproduction in competitors, mate-guarding, etc.); NBT, non-breeding territory; NS, nest/egg site; PC, parental care (either signalling parental quality or maintaining high parental care from partner by excluding additional mates), RS, reproductive success; SD, social dominance; GRC, general reproductive competition where fitness component is not known (dominance, mates, reproductive status); NF, trait studied but no function identified; U, unknown.
bCodes indicate whether competition for ecological rather than sexual resources was biased towards females (F > M), males (F < M) or neither sex (F = M). Some examples are as follows: species in which both sexes use signals to attract mates, but only females use them to defend ecological resources are listed as F > M; species in which males use signals to attract mates and defend ecological resources, whereas female signals appeared to be non-adaptive, are listed as F < M; and species in which both sexes use signals for mate attraction (or resource defence) with equal intensity are listed as F = M. In some cases, the emphasis was unknown (U).
cFitness components similar, but female more aggressive in defence of breeding territory.
dNo known ecological component to fitness.
eEmpirical study shows that the signal correlates with condition or reproductive success, but signal function remains unclear.
fTrait differs between sexes, e.g. different colour or location.
gOrnamented females reject males.
Our survey is based mainly on publications cited in (or citing) recent reviews on female and mutual ornamentation [5,6,9], and is not by any means exhaustive. The information is also unevenly distributed across taxonomic groups, with a distinct avian bias. This reflects our own research interests, coupled with the fact that many bird species are conspicuously ornamented, and attract a disproportionate amount of research attention. Nonetheless, we feel that this summary provides at least a qualitatively accurate window on the distribution of fitness components influencing the evolution of ornamental traits used in social interactions.
We compiled data on broad categories of fitness (e.g. reproductive success), direct components of fitness (e.g. matings) and potential routes to fitness (e.g. breeding territories; table 1). These aspects varied widely in the extent to which they corresponded to definitions of sexual selection, ranging from competition for mates (pure sexual selection) to avoidance of predation (natural selection). In each case, we recorded whether traits functioned in competition targeting ecological resources such as nest sites and parental care. As we were interested in the extent to which resources are linked to reproduction, we noted whether competition for territories and food was restricted to the breeding season.
To summarize the tabulated information, we constructed a concept map (figure 1) showing the relationship between the three main classes of selection, i.e. sexual selection nested within other types of social selection, and social selection nested within other types of natural selection. Some authors have suggested that sexual selection belongs in a category distinct from natural selection [16], but most accept that it is a form of natural selection—although a special one [48,54]. Our concept map combines this latter viewpoint and West-Eberhard's [12] definition of social selection, wherein sexual selection is a special form of social selection [41].
Placing aspects of fitness within this framework is necessarily somewhat subjective. Some aspects are easily assigned, such as competition for mates, which falls squarely within sexual selection, and which clearly occurs in both sexes. Greyer areas relate to variation within categories. Thus, the defence of breeding territories may involve different processes of selection depending on whether the territories are solely for mate attraction (sexual selection) or food (social selection). We highlight these ambiguous cases by placing them over the boundary between sexual and non-sexual social selection. Illustrating aspects of fitness on this framework raise two key points: sexual and non-sexual selection are often impossible to disentangle using the information currently available, and social selection is better at capturing most forms of selection on ornamental traits.
On the concept map (figure 1), we also varied the size of codes in relation to the number of cases identified in our survey, revealing a strong contrast between the sexes. In line with traditional sexual selection theory [22,23,48], male traits often function in competition for matings, whereas female traits mediate competition for a range of resources, associated mainly with reproduction. In particular, competition for mates was twice as frequent in males compared with females (51 versus 27 cases), whereas females were more likely than males to compete for resources such as nest sites (nine versus four cases) and parental care (eight versus one case). Again, the implication is clear—the framework of sexual selection has shortcomings in both sexes, but it works better for males than for females.
Finally, we classified each case according to differences between the sexes in their overall emphasis on ecological competition rather than matings, focusing solely on the function of ornamental traits, and taking into account all aspects of fitness reported in each study. This classification reveals that it is more common for ecological competition to be important for females than for males (figure 2). Moreover, this difference between the sexes would probably be further accentuated if we included cases of male-only ornamentation, rather than restricting the survey to traits expressed in both sexes.
Figure 2.
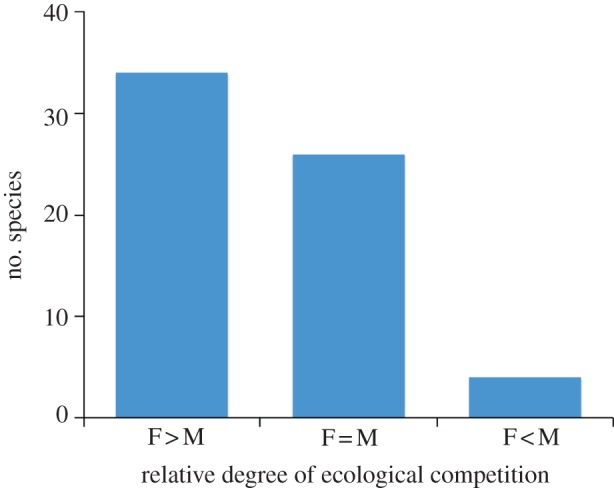
Summary of sex differences in the relative importance of ecological competition, based on data in table 1 (excluding uncertain cases). Species (n = 64) are divided into categories denoting whether the emphasis on competition for ecological resources was biased towards females (F > M), males (F < M), or neither sex (F = M). The sexual bias of emphasis on ecological competition was determined either by the difference between males and females in ecological versus sexual components of fitness (figure 1), or by the strength of competition within categories. See table 1 for further details of assignment to categories.
The main conclusions we draw from this exercise (figures 1 and 2) are that (i) Darwin's concept of sexual selection can regularly be applied to females as well as males, (ii) the function of ornamental traits varies widely in both sexes, (iii) most aspects of fitness are difficult to assign with any confidence to sexual selection, (iv) many selective mechanisms are a better fit with West-Eberhard's theory of social selection and (v) social selection is especially relevant to females, as they more often compete over ecological rather than sexual resources. We therefore argue that the long-standing narrow focus on sexual selection may have clouded our view, and that social selection provides a better framework for understanding the complex array of processes driving ornamental trait evolution in both sexes, particularly in females.
(f). The selection continuum
The scattered distribution of codes in figure 1 reflects the fact that ornamental traits have a variety of functions, and that their link with reproduction ranges from direct to very indirect. Some traits have multiple functions in both sexes, from mate attraction to predator deterrence or avoidance [159,180,207–208]. Others have a single function that is difficult to assign to any particular theory of selection. Overall, given the role of ornamental traits in numerous contexts unrelated to reproduction [42,209,210], selection on such traits is best characterized as a continuum running from sexual selection to natural selection (figure 3). One of the main implications of this continuum is that evidence for sexual selection does not rule out the importance of other processes. Thus, even though sexual selection is widespread in females, it should not be inferred without considering contexts that favour non-sexual forms of social selection.
Figure 3.

Female traits mediate competition in different social contexts that span a continuum from pure Darwinian sexual selection over matings (left, red) to pure non-sexual social competition over ecological resources like food and winter territories (right, blue), as well as natural selection in the form of predator-avoidance. This continuum runs from ornaments on the left to weapons (and anti-predator signals) on the right, though many traits have multiple functions at different points along the continuum. Clockwise from upper left: Irediparra gallinacea, Aethia cristatella, Icterus pustulatus, Ovis aries, Eumomota superciliosa, Erithacus rubecula, Onthophagus sagittarius, Lamprotornis superbus, Sceloporus virgatus and Rhamphomyia longicuada. See table 1 for fitness contexts for all species, except Irediparra gallinacea [211], where females compete for males, and Lamprotornis superbus [68], where females may compete for mates, reproductive opportunities or territories. Placement of some species along the sexual–non-sexual continuum is speculative because the fitness components underlying competition are poorly known. See acknowledgements for photo credits.
The tendency to interpret ambiguous results in the light of sexual selection is particularly widespread in studies of female or mutual ornamentation [19,25,26,212–214]. Most researchers assume that, if ornaments correlate with condition or breeding success, they must function in mate choice, ignoring the fact that those traits may function mainly in social competition over ecological resources. Likewise, aggression in the breeding season is often assumed to be about mates or mating opportunities. Research on chacma baboons, for example, revealed that female–female aggression increases when more females are fertile, leading to the conclusion that aggression is ‘associated with access to mates’ [98]. However, the data do not rule out competition for fecundity via reproductive suppression, or indeed hormonally induced aggression unrelated to matings [215].
It is also clear that mechanisms underlying ornamental traits should not be assumed to relate exclusively to reproduction. For example, studies of female ornaments in dark-eyed juncos (Junco hyemalis) assume that the main route to fitness is increased reproductive success [27], but the same signals are also known to mediate dominance interactions over food during the non-breeding season [216]. Similarly, patterns of variation in male eyestalks of diopsid flies are generally thought to be driven entirely by sexual selection [217], even though provisioning experiments show that they also increase access to food [201]. Even in Drosophila melanogaster, one of the most intensively studied organisms, behavioural traits long assumed to be sexual displays actually relate to aggressive defence of food [218]. It seems likely that the predominance of the sexual selection paradigm has resulted in an overemphasis on the role of sexual interactions in the study of such ornamental traits, particularly in females.
Evidence for mate choice also needs to be interpreted cautiously because individuals may choose mates on the basis of competitive ability. In particular, when cooperative defence of resources is key to maximizing fitness, it may be critical to choose a mate that can help with defence. If resource defence is mediated by an ornamental trait, then that trait may be preferred because it represents improved access to space and food (resources only partly related to fecundity). This process may explain the preponderance of mutual ornaments in cases where both sexes defend territories [9,12,219–221]. Crucially, the role of social selection as a key driver of ornamental trait evolution is difficult to rule out in these cases, even when mating preferences for ornaments are demonstrated.
7. Linking social contexts and sexual patterns of ornamentation
In this section, we summarize the main sources of selection on female ornamental traits, and discuss the extent to which they are likely to reflect social selection. Synthesizing the results of our survey of empirical studies (table 1), the following contexts appear in the literature:
— Mutual sexual selection is one of the main explanations for ornamentation in both sexes [52,72], though it has rarely been demonstrated convincingly. Many studies provide only indirect evidence for mutual mate choice, and mate choice itself does not necessarily equate to sexual selection [15]. We still know relatively little about how individuals gain from choosing ornamented mates—the benefits could accrue from enhanced parental care or ability to defend territories, or to a number of other factors that fall outside the core ideas of sexual selection.
— Mutual social selection (a term we introduce here) is a broader mechanism that may more correctly explain many putative cases of mutual sexual selection. Mutual social selection applies whenever mutual mechanisms extend beyond sexual selection, for example when mates choose each other on the basis of ornamental traits linked to dominance in social competition. Mutual social selection also occurs when ornamental traits function in pair coordination, i.e. when they facilitate coordination and synchronization of breeding activities, and thereby enhance fecundity by improving breeding success [17]. The plumes and soft-part colours of grebes, herons and other waterbirds are possible examples, as these ornaments appear to serve a purpose long after mate choice is completed, making them difficult to fit into the framework of sexual selection.
— Individual social selection (the default non-sexual form of social selection) occurs when there is not necessarily anything mutual (i.e. reciprocal) about the action of selection. Thus, pairs may cooperate to defend territories, and signal as a coalition to deter rival pairs [222], without any inter-sexual preference for the signal. Weapons and other signals of social dominance (e.g. badges of status) are always shaped by individual social selection. This topic has received much attention in the study of males [42,43,159,223], but our literature review suggests that the same mechanism may be widespread in females. We also note that, unlike sexual selection and mutual social selection, individual social selection can extend beyond species boundaries via interspecific competition [174].
— Indirect social selection may apply when traits mediate competition in social groups containing relatives [131], and where fitness benefits accrue to relatives (i.e. competitors gain fitness through kin selection, or other forms of indirect selection). In many cases, it will be difficult to distinguish between direct and indirect fitness benefits, but where the competitors are reproductively sterile, as in eusocial organisms, only indirect fitness benefits are feasible. Thus, selection for a ‘green-beard’ gene in Solonopsis ants [224] provides a plausible example of indirect social selection as it involves sterile workers identifying and killing all queens lacking the relevant allele, apparently based on an odour cue.
8. Why we need a social selection framework
We have outlined a number of social and ecological contexts where sexual selection may either be a less important driver of evolution than social selection, or perhaps not involved at all. These contexts cover the types of examples that have led other authors to attack the Darwinian framework of sexual selection as a failed theory [225], or to suggest replacing it with alternatives [34]. To work around these issues, and to accommodate often conspicuously different mechanisms in males and females, some authors have suggested classifying female-mediated mechanisms under natural selection [226], whereas others have suggested expanding sexual selection to include all forms of reproductive competition [3].
Unfortunately, these approaches bring problems of their own. On the one hand, invoking natural selection obscures the social dynamics of the underlying mechanisms. On the other hand, expanding traditional sexual selection theory to include all of reproduction moves far from Darwin's original concept, and still misses some sources of selection, particularly in females (figure 1). Moreover, these solutions do not resolve the general problem that similar traits can be produced by different forms of selection (figure 3). It could be argued that we should simply abandon names for different selective forces altogether, and instead focus on identifying the components, intensity and targets of selection themselves [3,31]. This makes good sense, but we suggest that classifying selection into broad categories—even when boundaries are blurred (figure 1)—provides useful guidance when designing, explaining or interpreting empirical studies.
We favour the alternative route suggested by West-Eberhard [12], who argued that sexual selection should be retained intact, but that it is more usefully nested within the general framework of social selection. This seems to us a superior solution because it helps to clarify that similar traits arise in reproductive and non-reproductive contexts, and provides us with different labels for these contexts. These labels force us to be explicit about the components of fitness driving the evolution of traits in each sex. Thinking in terms of social selection therefore provides a more complete framework for disentangling the contrasting dynamics of aggression and competition in both sexes [75,76].
One of the main arguments against adopting a social selection framework is that it may be difficult to distinguish clearly between non-sexual and sexual selection [3]. However, this reasoning ignores the nested relationship of sexual selection within the broader framework of social selection, and the fact that traits can function in multiple ambiguous contexts (figure 1). We believe the logic should be inverted: it is necessary to adopt social selection as our fundamental framework precisely because it is so difficult to rule out selection arising from social interactions in non-sexual contexts. A major advantage of social selection theory is that it reduces the arbitrariness and confusion caused by applying the concept of sexual selection to all socially mediated processes driving the evolution of ornamental traits. In effect, social selection serves as an important catch-all term encompassing the full array of interactions within the social environment, and is particularly useful when the link between selection and reproduction is unclear [174,227].
Social selection is useful not only in terms of a conceptual framework, but because it also helps to resolve questions arising from a narrow focus on sexual selection. It can explain confusing patterns of trait expression [33,228], and it can provide the missing component of fitness in studies of evolution. For example, sexual selection struggles to explain cases in which the fitness costs of an ornament are not matched by gains in mating success [229], whereas the social selection framework points to the additional fitness accrued from social dominance in non-sexual contexts.
9. Where next?
Social mechanisms have already been incorporated into theoretical studies that define social selection as the effects on the fitness of one individual that directly result from traits of interacting individuals [30,38]. This definition is useful in that it widens the focus beyond the competitive processes highlighted by West-Eberhard [12], and incorporates other forms of interaction, such as cooperation. Quantitative genetic models designed to capture indirect genetic effects [230] provide a promising toolkit, and should continue to be developed and refined into a formal theoretical framework for social selection. However, theoretical models can tell us little about the balance of contexts underpinning selection on ornamental traits in real-world scenarios—they do not clarify how traits function across different social contexts, and how these functions contribute to fitness. For this, we need explicit tests of social selection theory, which tends to be overlooked in empirical studies.
To deepen our understanding, empiricists need to consider the potential role of alternative resources—beyond matings, and even beyond reproductive success—in driving selection via social interactions. This is particularly important in females as so much female–female competition targets ecological resources, but it is also worth revisiting studies of males because male–male competition for mates and breeding opportunities are probably less important in some social contexts than is currently believed. These questions may lend themselves to meta-analytical approaches and broad-scale comparative analyses.
Our survey of literature (table 1) also suggests that we need a broader sample of contexts and taxa. Successful paradigms such as sexual selection tend to be self-reinforcing because researchers pick systems that are ideal for studying the central questions. Evolutionary studies have tended to focus on species with extravagant male-biased ornamentation, where sexual selection seems an obvious driver of trait evolution. This bias is being corrected to some extent by a recent focus on social mammals [2,66] and mutually ornamented birds [6], but the questions still tend to be viewed from the perspective of sexual selection. Insects, fish, mammals and birds offer a wealth of systems for testing hypotheses about the role of female ornaments in both sexual and non-sexual contexts. Tropical regions in particular are home to a multitude of poorly known species with exaggerated traits in females [6].
10. Conclusions
West-Eberhard's [12] concept of social selection encompasses all traits and mechanisms involved in social interactions, and therefore focuses attention on the full spectrum of contexts that favour the evolution of ornamental traits, while leaving the original concept of sexual selection intact. Social selection thus provides a consistent framework for studying socially mediated mechanisms of selection, for understanding the separate influences of competition and cooperation, and for the language we use when talking about trait evolution in different social contexts. We argue that this theory deserves greater prominence as a potential solution to the current debate about how ornamental traits evolve in females.
Sexual selection can logically be considered a nested subset of social selection, but the boundary between these categories is somewhat arbitrary. In effect, socially mediated selection for ornamental traits falls along a continuum from purely sexual to purely non-sexual forms of social selection. Studies of phenotypic evolution in males and females are currently difficult to classify along this sexual–non-sexual selection continuum because in most systems we still have only a superficial understanding of the mechanisms that have shaped ornamental traits. Social selection provides a broader perspective that not only can help to remedy this problem, but also can reveal the underlying links between patterns of social organization and patterns of trait evolution. We need to make fuller use of this framework when designing and interpreting studies of selection on animal phenotypes.
Acknowledgements
We thank The Columbia University Seminar in Population Biology for inviting us to their Symposium on Sexual Selection, Sexual Conflict and the Female Perspective in April 2011. We are also grateful to Ken Kraaijeveld, Geoffrey While and two anonymous reviewers for constructive comments on the manuscript. J.A.T. was funded by a John Fell Fund grant; R.M. by the Natural Sciences and Engineering Research Council of Canada and Centre national de la recherche scientifique of France; and B.E.L. by the U.S. National Science Foundation. Photo credits for figure 1: A. cristatella (Ian Jones), O. aries (Julian Sawyer), E. rubecula (Graham Eaton), O. sagittarius (Sean Stankowski), Sceloporus virgatus (Stacey Weiss), Rhamphomyia longicuada (David Funk); all others B.E.L.
References
- 1.Darwin C. 1871. The descent of man, and selection in relation to sex, vol. 2 London, UK: John Murray [Google Scholar]
- 2.Clutton-Brock T. H. 2007. Sexual selection in males and females. Science 318, 1882–1885 10.1126/science.1133311 (doi:10.1126/science.1133311) [DOI] [PubMed] [Google Scholar]
- 3.Clutton-Brock T. H. 2009. Sexual selection in females. Anim. Behav. 77, 3–11 10.1016/j.anbehav.2008.08.026 (doi:10.1016/j.anbehav.2008.08.026) [DOI] [Google Scholar]
- 4.Amundsen T. 2000. Female ornaments: genetically correlated or sexually selected? In Signalling and signal design in animal communication (eds Espmark Y., Amundsen T., Rosenqvist G.), pp. 133–154 Trondheim, Norway: Tapir Academic Press [Google Scholar]
- 5.Rosvall K. A. 2011. Intrasexual selection on females: evidence for sexual selection? Behav. Ecol. 22, 1131–1140 10.1093/beheco/arr106 (doi:10.1093/beheco/arr106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kraaijeveld K., Kraaijeveld-Smit F. J. L., Komdeur J. 2007. The evolution of mutual ornamentation. Anim. Behav. 74, 657–677 10.1016/j.anbehav.2006.12.027 (doi:10.1016/j.anbehav.2006.12.027) [DOI] [Google Scholar]
- 7.Lande R. 1980. Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 34, 292–305 10.2307/2407393 (doi:10.2307/2407393) [DOI] [PubMed] [Google Scholar]
- 8.LeBas N. R. 2006. Female finery is not for males. Trends Ecol. Evol. 21, 170–173 10.1016/j.tree.2006.01.007 (doi:10.1016/j.tree.2006.01.007) [DOI] [PubMed] [Google Scholar]
- 9.Tobias J. A., Gamarra-Toledo V., Garcia-Olaechea D., Pulgarin P. C., Seddon N. 2011. Year-round resource defence and the evolution of male and female song in suboscine birds: social armaments are mutual ornaments. J. Evol. Biol. 24, 2118–2138 10.1111/j.1420-9101.2011.02345.x (doi:10.1111/j.1420-9101.2011.02345.x) [DOI] [PubMed] [Google Scholar]
- 10.Lande R. 1987. Genetic correlations between the sexes in the evolution of sexual dimorphism and mating preferences. In Sexual selection: testing the alternatives (eds Bradbury J. W., Andersson M. B.), pp. 83–94 Chichester, UK: Wiley [Google Scholar]
- 11.Stankowich T., Caro T. 2009. The evolution of weaponry in female bovids. Proc. R. Soc. B 276, 4329–4334 10.1098/rspb.2009.1256 (doi:10.1098/rspb.2009.1256) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.West-Eberhard M. J. 1983. Sexual selection, social competition, and speciation. Q. Rev. Biol. 58, 155–183 10.1086/413215 (doi:10.1086/413215) [DOI] [Google Scholar]
- 13.West-Eberhard M. J. 1979. Sexual selection, social competition, and evolution. Proc. Am. Philos. Soc. 123, 222–234 [Google Scholar]
- 14.Darwin C. 1859. The origin of species by means of natural selection. London, UK: John Murray [Google Scholar]
- 15.Arnold S. J. 1994. Is there a unifying concept of sexual selection that applies to both plants and animals? Am. Nat. 144, S1–S12 10.1086/285650 (doi:10.1086/285650) [DOI] [Google Scholar]
- 16.Hosken D. J., House C. M. 2011. Sexual selection. Curr. Biol. 21, R62–65 10.1016/j.cub.2010.11.053 (doi:10.1016/j.cub.2010.11.053) [DOI] [PubMed] [Google Scholar]
- 17.Clutton-Brock T. H. 2010. We do not need a sexual selection 2.0: nor a theory of genial selection. Anim. Behav. 79, 7–10 10.1016/j.anbehav.2009.10.018 (doi:10.1016/j.anbehav.2009.10.018) [DOI] [Google Scholar]
- 18.Huxley J. 1914. The courtship-habits of the Great Crested Grebe (Podiceps cristatus); with an addition to the theory of sexual selection. Proc. Zool. Soc. Lond. 35, 491–562 [Google Scholar]
- 19.Berglund A., Magnhagen C., Bisazza A., Konig B., Huntingford F. 1993. Female–female competition over reproduction. Behav. Ecol. 4, 184–187 10.1093/beheco/4.2.184 (doi:10.1093/beheco/4.2.184) [DOI] [Google Scholar]
- 20.Crook J. H. 1972. Sexual selection, dimorphism and social organization in the primates. In Sexual selection and the de scent of man 1871–1971 (ed. Campbell B.), pp. 231–281 Chicago, IL: Aldine [Google Scholar]
- 21.Amundsen T. 2000. Why are female birds ornamented? Trends Ecol. Evol. 15, 149–155 10.1016/S0169-5347(99)01800-5 (doi:10.1016/S0169-5347(99)01800-5) [DOI] [PubMed] [Google Scholar]
- 22.Trivers R. L. 1972. Parental investment and sexual selection. In Sexual selection and the descent of man 1871–1971 (ed. Campbell B.), pp. 136–179 Chicago, IL: Aldine [Google Scholar]
- 23.Bateman A. J. 1948. Intra-sexual selection in Drosophila. Heredity 2, 349–368 10.1038/hdy.1948.21 (doi:10.1038/hdy.1948.21) [DOI] [PubMed] [Google Scholar]
- 24.Murphy T. G., Hernández-Muciño D., Osorio-Beristain M., Montgomerie R., Omland K. E. 2009. Carotenoid-based status signaling by females in the tropical streak-backed oriole. Behav. Ecol. 20, 1000–1006 10.1093/beheco/arp089 (doi:10.1093/beheco/arp089) [DOI] [Google Scholar]
- 25.Daunt F., Monaghan P., Wanless S., Harris M. P. 2003. Sexual ornament size and breeding performance in female and male European shags Phalacrocorax aristotelis. Ibis 145, 54–60 10.1046/j.1474-919X.2003.00120.x (doi:10.1046/j.1474-919X.2003.00120.x) [DOI] [Google Scholar]
- 26.Velando A., Lessells C. M., Márquez J. C. 2001. The function of male and female ornaments in the Inca Tern: evidence for links between ornament expression and both adult condition and reproductive performance. J. Avian Biol. 32, 311–318 10.1111/j.0908-8857.2001.320404.x (doi:10.1111/j.0908-8857.2001.320404.x) [DOI] [Google Scholar]
- 27.Cain K. E., Ketterson E. D. 2012. Competitive females are successful females; phenotype, mechanism, and selection in a common songbird. Behav. Ecol. Sociobiol. 66, 241–252 10.1007/s00265-011-1272-5 (doi:10.1007/s00265-011-1272-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Price T. 2008. Speciation in birds. Greenwood, CO: Roberts and Co [Google Scholar]
- 29.Nesse R. 2007. Runaway social selection for displays of partner value and altruism. Biol. Theory 2, 143–155 10.1162/biot.2007.2.2.143 (doi:10.1162/biot.2007.2.2.143) [DOI] [Google Scholar]
- 30.McGlothlin J. W., Moore A. J., Wolf J. B., Brodie E. D. 2010. Interacting phenotypes and the evolutionary process. III. Social evolution. Evolution 64, 2558–2574 10.1111/j.1558-5646.2010.01012.x (doi:10.1111/j.1558-5646.2010.01012.x) [DOI] [PubMed] [Google Scholar]
- 31.Cornwallis C. K., Uller T. 2010. Towards an evolutionary ecology of sexual traits. Trends Ecol. Evol. 25, 145–152 10.1016/j.tree.2009.09.008 (doi:10.1016/j.tree.2009.09.008) [DOI] [PubMed] [Google Scholar]
- 32.Santana S. E., Alfaro J. L., Alfaro M. E. 2012. Adaptive evolution of facial colour patterns in Neotropical primates. Proc. R. Soc. B 279, 2204–2211 10.1098/rspb.2011.2326 (doi:10.1098/rspb.2011.2326) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Friedman N. R., Hofmann C. M., Kondo B., Omland K. E. 2009. Correlated evolution of migration and sexual dichromatism in the New World orioles (Icterus). Evolution 63, 3269–3274 10.1111/j.1558-5646.2009.00792.x (doi:10.1111/j.1558-5646.2009.00792.x) [DOI] [PubMed] [Google Scholar]
- 34.Roughgarden J., Oishi M., Akcay E. 2006. Reproductive social behavior: cooperative games to replace sexual selection. Science 311, 965–969 10.1126/science.1110105 (doi:10.1126/science.1110105) [DOI] [PubMed] [Google Scholar]
- 35.Roughgarden J. 2012. The social selection alternative to sexual selection. Phil. Trans. R. Soc. B 367, 2294–2303 10.1098/rstb.2011.0282 (doi:10.1098/rstb.2011.0282) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wynne-Edwards V. C. 1962. Animal dispersion in relation to social behaviour. Edinburgh, UK: Oliver and Boyd [Google Scholar]
- 37.Ghiselin M. T. 1974. The economy of nature and the evolution of sex. Berkeley, CA: University of California Press [Google Scholar]
- 38.Wolf J. B., Brodie E. D., Moore A. J. 1999. Interacting phenotypes and the evolutionary process. II. Selection resulting from social interactions. Am. Nat. 153, 254–266 10.1086/303168 (doi:10.1086/303168) [DOI] [PubMed] [Google Scholar]
- 39.Tanaka Y. 1996. Social selection and the evolution of animal signals. Evolution 50, 512–523 10.2307/2410827 (doi:10.2307/2410827) [DOI] [PubMed] [Google Scholar]
- 40.Moore A. J., Brodie E. D., Wolf J. B. 1997. Interacting phenotypes and the evolutionary process. I. Direct and indirect genetic effects of social interactions. Evolution 51, 1352–1362 10.2307/2411187 (doi:10.2307/2411187) [DOI] [PubMed] [Google Scholar]
- 41.Lyon B. E., Montgomerie R. 2012. Sexual selection is a form of social selection. Phil. Trans. R. Soc B 367, 2266–2273 10.1098/rstb.2012.0012 (doi:10.1098/rstb.2012.0012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rohwer S. 1985. Dyed birds achieve higher social status than controls in Harris Sparrows. Anim. Behav. 33, 1325–1331 10.1016/S0003-3472(85)80193-7 (doi:10.1016/S0003-3472(85)80193-7) [DOI] [Google Scholar]
- 43.Senar J. C. 2006. Bird coloration as an intrasexual signal of aggression and dominance. In Bird coloration, vol. 2: function and evolution (eds Hill G. E., McGraw K. J.), pp. 125–193 Cambridge, MA: Harvard University Press [Google Scholar]
- 44.Santos E. S. A., Scheck D., Nakagawa S. 2011. Dominance and plumage traits: meta-analysis and metaregression analysis. Anim. Behav. 82, 3–19 10.1016/j.anbehav.2011.03.022 (doi:10.1016/j.anbehav.2011.03.022) [DOI] [Google Scholar]
- 45.Krebs E. A., Putland D. A. 2004. Chic chicks: the evolution of chick ornamentation in rails. Behav. Ecol. 6, 946–951 10.1093/beheco/arh078 (doi:10.1093/beheco/arh078) [DOI] [Google Scholar]
- 46.Lyon B. E., Eadie J. M., Hamilton L. D. 1994. Parental choice selects for ornamental plumage in American coot chicks. Nature 371, 240–243 10.1038/371240a0 (doi:10.1038/371240a0) [DOI] [Google Scholar]
- 47.Kilner R. M. 2006. Function and evolution of color in young birds. In Bird coloration, vol. 2 (eds Hill G., McGraw K.), pp. 201–232 London, UK: Harvard University Press [Google Scholar]
- 48.Darwin C. 1871. The descent of man, and selection in relation to sex. London, UK: J. Murray [Google Scholar]
- 49.Ryan M. J. 2011. Replication in field biology: the case of the frog-eating bat. Science 334, 1229–1230 10.1126/science.1214532 (doi:10.1126/science.1214532) [DOI] [PubMed] [Google Scholar]
- 50.Wallace A. R. 1889. Darwinism: an exposition of the theory of natural selection with some of its applications. London, UK: Macmillan [Google Scholar]
- 51.Kirkpatrick M., Price T., Arnold S. J. 1990. The Darwin–Fisher theory of sexual selection in monogamous birds. Evolution 44, 180–193 10.2307/2409533 (doi:10.2307/2409533) [DOI] [PubMed] [Google Scholar]
- 52.Jones I. L., Hunter F. M. 1993. Mutual sexual selection in a monogamous seabird. Nature 362, 238–239 10.1038/362238a0 (doi:10.1038/362238a0) [DOI] [Google Scholar]
- 53.Parker G. A. 1970. Sperm competition and its evolutionary consequences in insects. Biol. Rev. Cambridge Philos. Soc. 45, 525–567 10.1111/j.1469-185X.1970.tb01176.x (doi:10.1111/j.1469-185X.1970.tb01176.x) [DOI] [Google Scholar]
- 54.Andersson M. 1994. Sexual selection. Princeton, NJ: Princeton University Press [Google Scholar]
- 55.Arnqvist G., Rowe L. 2005. Sexual conflict. Princeton, NJ: Princeton University Press [Google Scholar]
- 56.Kokko H., Jennions M. D., Brooks R. 2006. Unifying and testing models of sexual selection. Annu. Rev. Ecol. Evol. Syst. 37, 43–66 10.1146/annurev.ecolsys.37.091305.110259 (doi:10.1146/annurev.ecolsys.37.091305.110259) [DOI] [Google Scholar]
- 57.Berglund A., Rosenqvist G. 2001. Male pipefish prefer ornamented females. Anim. Behav. 61, 345–350 10.1006/anbe.2000.1599 (doi:10.1006/anbe.2000.1599) [DOI] [Google Scholar]
- 58.Amundsen T., Forsgren E. 2001. Male mate choice selects for female coloration in a fish. Proc. Natl Acad. Sci. USA 98, 13 155–13 160 10.1073/pnas.211439298 (doi:10.1073/pnas.211439298) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Forsgren E., Amundsen T., Borg A. A., Bjelvenmark J. 2004. Unusually dynamic sex roles in a fish. Nature 429, 551–554 10.1038/nature02562 (doi:10.1038/nature02562) [DOI] [PubMed] [Google Scholar]
- 60.Muma K. E., Weatherhead P. J. 1989. Male traits expressed in females: direct or indirect sexual selection. Behav. Ecol. Sociobiol. 25, 25–31 [Google Scholar]
- 61.Cuervo J. J., de Lope F., Møller A. P. 1996. The function of long tails in female barn swallows (Hirundo rustica): an experimental study. Behav. Ecol. 7, 132–136 10.1093/beheco/7.2.132 (doi:10.1093/beheco/7.2.132) [DOI] [Google Scholar]
- 62.Tella J. L., Forero M. G., Donazar J. A., Hiraldo F. 1997. Is the expression of male traits in female lesser kestrels related to sexual selection? Ethology 103, 72–81 10.1111/j.1439-0310.1997.tb00008.x (doi:10.1111/j.1439-0310.1997.tb00008.x) [DOI] [Google Scholar]
- 63.Chenoweth S. F., Blows M. W. 2005. Contrasting mutual sexual selection on homologous signal traits in Drosophila serrata. Am. Nat. 165, 281–289 10.1086/427271 (doi:10.1086/427271) [DOI] [PubMed] [Google Scholar]
- 64.Bonduriansky R. 2001. The evolution of male mate choice in insects: a synthesis of ideas and evidence. Biol. Rev. 76, 305–339 10.1017/S1464793101005693 (doi:10.1017/S1464793101005693) [DOI] [PubMed] [Google Scholar]
- 65.Hauber M. E., Lacey E. A. 2005. Bateman's principle in cooperatively breeding vertebrates: the effects of non-breeding alloparents on variability in female and male reproductive success. Integr. Comp. Biol. 45, 903–914 10.1093/icb/45.5.903 (doi:10.1093/icb/45.5.903) [DOI] [PubMed] [Google Scholar]
- 66.Clutton-Brock T. H., Hodge S. J., Spong G., Russell A. F., Jordan N. R., Bennett N. C., Sharpe L. L., Manser M. B. 2006. Intrasexual competition and sexual selection in cooperative mammals. Nature 444, 1065–1068 10.1038/nature05386 (doi:10.1038/nature05386) [DOI] [PubMed] [Google Scholar]
- 67.Langmore N. E., Davies N. B., Hatchwell B. J., Hartley I. R. 1996. Female song attracts males in the alpine accentor Prunella collaris. Proc. R. Soc. Lond. B 263, 141–146 10.1098/rspb.1996.0022 (doi:10.1098/rspb.1996.0022) [DOI] [Google Scholar]
- 68.Rubenstein D. R., Lovette I. J. 2009. Reproductive skew and selection on female ornamentation. Nature 462, 786–789 10.1038/nature08614 (doi:10.1038/nature08614) [DOI] [PubMed] [Google Scholar]
- 69.Hooper P. L., Miller G. F. 2008. Mutual mate choice can drive costly signaling even under perfect monogamy. Adaptive Behav. 16, 53–70 10.1177/1059712307087283 (doi:10.1177/1059712307087283) [DOI] [Google Scholar]
- 70.Johnstone R. A., Reynolds J. D., Deutsch J. C. 1996. Mutual mate choice and sex differences in choosiness. Evolution 50, 1382–1391 10.2307/2410876 (doi:10.2307/2410876) [DOI] [PubMed] [Google Scholar]
- 71.Kozak G. M., Reisland M., Boughman J. W. 2008. Sex differences in mate recognition and conspecific preference in species with mutual mate choice. Evolution 63, 353–365 10.1111/j.1558-5646.2008.00564.x (doi:10.1111/j.1558-5646.2008.00564.x) [DOI] [PubMed] [Google Scholar]
- 72.Kraaijeveld K. 2003. Degree of mutual ornamentation is related to divorce rate in birds. Proc. R. Soc. Lond. B 270, 1785–1791 10.1098/rspb.2003.2450 (doi:10.1098/rspb.2003.2450) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Amundsen T., Forsgren E., Hansen L. T. T. 1997. On the function of female ornaments: male bluethroats prefer colourful females. Proc. R. Soc. Lond. B 264, 1579–1586 10.1098/rspb.1997.0220 (doi:10.1098/rspb.1997.0220) [DOI] [Google Scholar]
- 74.Torres R., Velando A. 2005. Male preference for female foot colour in the socially monogamous blue-footed booby, Sula nebouxii. Anim. Behav. 69, 59–65 10.1016/j.anbehav.2004.03.008 (doi:10.1016/j.anbehav.2004.03.008) [DOI] [Google Scholar]
- 75.Elias D. O., Botero C. A., Andrade M. C. B., Mason A. C., Kasumovic M. M. 2010. High resource valuation fuels ‘‘desperado’’ fighting tactics in female jumping spiders. Behav. Ecol. 21, 868–875 10.1093/beheco/arq073 (doi:10.1093/beheco/arq073) [DOI] [Google Scholar]
- 76.Nilsen S. P., Chan Y. B., Huber R., Kravitz E. A. 2004. Gender-selective patterns of aggressive behavior in Drosophila melanogaster. Proc. Natl Acad. Sci. USA 101, 12 342–12 347 10.1073/pnas.0404693101 (doi:10.1073/pnas.0404693101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Liker A., Székely T. 1997. Aggression among female Lapwings Vanellus vanellus. Anim. Behav. 54, 797–802 10.1006/anbe.1996.0491 (doi:10.1006/anbe.1996.0491) [DOI] [PubMed] [Google Scholar]
- 78.Kempenaers B. 1994. Polygyny in the blue tit: unbalanced sex ratio and female aggression restrict mate choice. Anim. Behav. 47, 943–957 10.1006/anbe.1994.1126 (doi:10.1006/anbe.1994.1126) [DOI] [Google Scholar]
- 79.Yasukawa K., Searcy W. A. 1982. Aggression in female red-winged blackbirds—a strategy to ensure male parental investment. Behav. Ecol. Sociobiol. 11, 13–17 10.1007/BF00297660 (doi:10.1007/BF00297660) [DOI] [Google Scholar]
- 80.Summers K. 1989. Sexual selection and intra-female competition in the green poison-dart frog, Dendrobates auratus. Anim. Behav. 37, 797–805 10.1016/0003-3472(89)90064-X (doi:10.1016/0003-3472(89)90064-X) [DOI] [Google Scholar]
- 81.Slagsvold T. 1993. Female–female aggression and monogamy in great tits Parus major. Ornis Scand. 24, 155–158 10.2307/3676366 (doi:10.2307/3676366) [DOI] [Google Scholar]
- 82.Norris K. J. 1990. Female choice and the evolution of the conspicuous plumage coloration of monogamous male great tits. Behav. Ecol. Sociobiol. 26, 129–138 [Google Scholar]
- 83.Seddon N., Tobias J. A. 2006. Duets defend mates in a suboscine passerine, the warbling antbird (Hypocnemis cantator). Behav. Ecol. 17, 73–83 10.1093/beheco/ari096 (doi:10.1093/beheco/ari096) [DOI] [Google Scholar]
- 84.Tobias J. A., Seddon N. 2009. Signal jamming mediates sexual conflict in a duetting bird. Curr. Biol. 19, 577–582 10.1016/j.cub.2009.02.036 (doi:10.1016/j.cub.2009.02.036) [DOI] [PubMed] [Google Scholar]
- 85.Watson N. L., Simmons L. W. 2010. Reproductive competition promotes the evolution of female weaponry. Proc. R. Soc. B 277, 2035–2040 10.1098/rspb.2009.2335 (doi:10.1098/rspb.2009.2335) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Watson N. L., Simmons L. W. 2010. Mate choice in the dung beetle Onthophagus sagittarius: are female horns ornaments? Behav. Ecol. 21, 424–430 10.1093/beheco/arp207 (doi:10.1093/beheco/arp207) [DOI] [Google Scholar]
- 87.Tibbetts E. A., Sheehan M. J. 2011. Facial patterns are a conventional signal of agonistic ability in Polistes exclamans paper wasps. Ethology 117, 1138–1146 10.1111/j.1439-0310.2011.01967.x (doi:10.1111/j.1439-0310.2011.01967.x) [DOI] [Google Scholar]
- 88.Lin N. 1972. Territorial behavior among males of the social wasp Polistes exclamans Viereck. Proc. Entomol. Soc. Wash. 74, 148–155 [Google Scholar]
- 89.Thornhill R., Alcock J. 1983. The evolution of insect mating systems. Cambridge, MA: Harvard University Press [Google Scholar]
- 90.Espmark Y. 1964. Studies in dominance–subordination relationship in a group of semi-domestic reindeer (Rangifer tarandus L.). Anim. Behav. 12, 420–426 10.1016/0003-3472(64)90061-2 (doi:10.1016/0003-3472(64)90061-2) [DOI] [Google Scholar]
- 91.Robinson M. R., Kruuk L. E. B. 2007. Function of weaponry in females: the use of horns in intrasexual competition for resources in female Soay sheep. Biol. Lett. 3, 651–654 10.1098/rsbl.2007.0278 (doi:10.1098/rsbl.2007.0278) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wolff J. O., Peterson J. A. 1998. An offspring-defense hypothesis for territoriality in female mammals. Ethol. Ecol. Evol. 10, 227–239 10.1080/08927014.1998.9522854 (doi:10.1080/08927014.1998.9522854) [DOI] [Google Scholar]
- 93.Madden J. R. 1974. Female territoriality in a Suffolk County, Long Island, population of Glaucomys volans. J. Mammal 55, 647–652 10.2307/1379554 (doi:10.2307/1379554) [DOI] [Google Scholar]
- 94.Christenson T. E., Leboeuf B. J. 1978. Aggression in female northern elephant seal, Mirounga angustirostris. Behaviour 64, 158–172 10.1163/156853978X00495 (doi:10.1163/156853978X00495) [DOI] [PubMed] [Google Scholar]
- 95.Kahlenberg S. M., Thompson M. E., Wrangham R. W. 2008. Female competition over core areas in Pan troglodytes schweinfurthii, Kibale National Park, Uganda. Int. J. Primatol. 29, 931–947 10.1007/s10764-008-9276-3 (doi:10.1007/s10764-008-9276-3) [DOI] [Google Scholar]
- 96.Murray C. M., Mane S. V., Pusey A. E. 2007. Dominance rank influences female space use in wild chimpanzees, Pan troglodytes: towards an ideal despotic distribution. Anim. Behav. 74, 1795–1804 10.1016/j.anbehav.2007.03.024 (doi:10.1016/j.anbehav.2007.03.024) [DOI] [Google Scholar]
- 97.Ron T., Henzi S. P., Motro U. 1996. Do female chacma baboons compete for a safe spatial position in a southern woodland habitat? Behaviour 133, 475–490 10.1163/156853996X00549 (doi:10.1163/156853996X00549) [DOI] [Google Scholar]
- 98.Huchard E., Cowlishaw G. 2011. Female–female aggression around mating: an extra cost of sociality in a multimale primate society. Behav. Ecol. 22, 1003–1011 10.1093/beheco/arr083 (doi:10.1093/beheco/arr083) [DOI] [Google Scholar]
- 99.Wrangham R. W. 1980. An ecological model of female-bonded primate groups. Behaviour 75, 262–300 10.1163/156853980X00447 (doi:10.1163/156853980X00447) [DOI] [Google Scholar]
- 100.Stockley P., Bro-Jørgensen J. 2011. Female competition and its evolutionary consequences in mammals. Biol. Rev. 86, 341–366 10.1111/j.1469-185X.2010.00149.x (doi:10.1111/j.1469-185X.2010.00149.x) [DOI] [PubMed] [Google Scholar]
- 101.Ralls K. 1976. Mammals in which females are larger than males. Q. Rev. Biol. 51, 245–276 10.1086/409310 (doi:10.1086/409310) [DOI] [PubMed] [Google Scholar]
- 102.Bennett A. T. D., Cuthill I. C., Partridge J. C., Lunau K. 1997. Ultraviolet plumage colors predict mate preferences in starlings. Proc. Natl Acad. Sci. USA 94, 8618–8621 10.1073/pnas.94.16.8618 (doi:10.1073/pnas.94.16.8618) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Swaddle J. P., Witter M. S. 1995. Chest plumage, dominance and fluctuating asymmetry in female starlings. Proc. R. Soc. Lond. B 260, 219–223 10.1098/rspb.1995.0083 (doi:10.1098/rspb.1995.0083) [DOI] [Google Scholar]
- 104.Johnson K., Rosetta D., Burley N. 1993. Preferences of female American goldfinches (Carduelis tristis) for natural and artificial male traits. Behav. Ecol. 4, 138–143 10.1093/beheco/4.2.138 (doi:10.1093/beheco/4.2.138) [DOI] [Google Scholar]
- 105.Murphy T. G., Rosenthal M. F., Montgomerie R., Tarvin K. A. 2009. Female American goldfinches use carotenoid-based bill coloration to signal status. Behav. Ecol. 20, 1348–1355 10.1093/beheco/arp140 (doi:10.1093/beheco/arp140) [DOI] [Google Scholar]
- 106.Beletsky L. D. 1983. Aggressive and pair-bond maintenance songs of female red-winged blackbirds. Z. Tierpsychol. 62, 47–54 [Google Scholar]
- 107.Cooney R., Cockburn A. 1995. Territorial defence is the major function of female song in the superb fairy-wren, Malurus cyaneus. Anim. Behav. 49, 1635–1647 10.1016/0003-3472(95)90086-1 (doi:10.1016/0003-3472(95)90086-1) [DOI] [Google Scholar]
- 108.Price J. J., Yunes-Jiménez L., Osorio-Beristain M., Omland K. E., Murphy T. G. 2008. Sex-role reversal in song? Females sing more frequently than males in the Streak-backed Oriole. Condor 110, 387–392 10.1525/cond.2008.8430 (doi:10.1525/cond.2008.8430) [DOI] [Google Scholar]
- 109.Illes A. E., Yunes-Jimenez L. 2009. A female songbird out-sings male conspecifics during simulated territorial intrusions. Proc. R. Soc. B 276, 981–986 10.1098/rspb.2008.1445 (doi:10.1098/rspb.2008.1445) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Rosvall K. A. 2008. Sexual selection on aggressiveness in females: evidence from an experimental test with tree swallows. Anim. Behav. 75, 1603–1610 10.1016/j.anbehav.2007.09.038 (doi:10.1016/j.anbehav.2007.09.038) [DOI] [Google Scholar]
- 111.Heinsohn R., Legge S., Endler J. A. 2005. Extreme reversed sexual dichromatism in a bird without sex role reversal. Science 309, 617–619 10.1126/science.1112774 (doi:10.1126/science.1112774) [DOI] [PubMed] [Google Scholar]
- 112.Rosvall K. A. 2011. Maintenance of variation in sexually selected traits in females: a case study using intrasexual aggression in tree swallows Tachycineta bicolor. J. Avian Biol. 42, 454–462 10.1111/j.1600-048X.2011.05322.x (doi:10.1111/j.1600-048X.2011.05322.x) [DOI] [Google Scholar]
- 113.Fedy B. C., Stutchbury B. J. M. 2005. Territory defence in tropical birds: are females as aggressive as males? Behav. Ecol. Sociobiol. 58, 414–422 10.1007/s00265-005-0928-4 (doi:10.1007/s00265-005-0928-4) [DOI] [Google Scholar]
- 114.Hau M., Stoddard S. T., Soma K. K. 2004. Territorial aggression and hormones during the non-breeding season in a tropical bird. Horm. Behav. 45, 40–49 10.1016/j.yhbeh.2003.08.002 (doi:10.1016/j.yhbeh.2003.08.002) [DOI] [PubMed] [Google Scholar]
- 115.George T. L. 1987. Behavior of territorial male and female Townsend's solitaires (Myadestes townsendi) in winter. Am. Midland Nat. 118, 121–127 10.2307/2425634 (doi:10.2307/2425634) [DOI] [Google Scholar]
- 116.Tobias J. A. 1997. Food availability as a determinant of pairing behaviour in the European robin. J. Anim. Ecol. 66, 629–639 10.2307/5916 (doi:10.2307/5916) [DOI] [Google Scholar]
- 117.Pryke S. R. 2007. Fiery red heads: female dominance among head color morphs in the Gouldian finch. Behav. Ecol. 18, 621–627 10.1093/beheco/arm020 (doi:10.1093/beheco/arm020) [DOI] [Google Scholar]
- 118.Crowhurst C. J., Zanollo V., Griggio M., Robertson J., Kleindorfer S. 2012. White flank spots signal feeding dominance in female diamond firetails, Stagonopleura guttata. Ethology 118, 63–75 10.1111/j.1439-0310.2011.01986.x (doi:10.1111/j.1439-0310.2011.01986.x) [DOI] [Google Scholar]
- 119.Griggio M., Zanollo V., Hoi H. 2010. Female ornamentation, parental quality, and competitive ability in the rock sparrow. J. Ethol. 28, 455–462 10.1007/s10164-010-0205-5 (doi:10.1007/s10164-010-0205-5) [DOI] [Google Scholar]
- 120.Pryke S. R., Griffith S. C. 2007. The relative role of male versus female mate choice in maintaining assortative pairing among discrete colour morphs. J. Evol. Biol. 20, 1512–1521 10.1111/j.1420-9101.2007.01332.x (doi:10.1111/j.1420-9101.2007.01332.x) [DOI] [PubMed] [Google Scholar]
- 121.Griggio M., Valera F., Casas A., Pilastro A. 2005. Males prefer ornamented females: a field experiment of male choice in the rock sparrow. Anim. Behav. 69, 1243–1250 10.1016/j.anbehav.2004.10.004 (doi:10.1016/j.anbehav.2004.10.004) [DOI] [Google Scholar]
- 122.Griggio M., Devigili A., Hoi H., Pilastro A. 2009. Female ornamentation and directional male mate preference in the rock sparrow. Behav. Ecol. 20, 1072–1078 10.1093/beheco/arp099 (doi:10.1093/beheco/arp099) [DOI] [Google Scholar]
- 123.Vergara P., Martínez-Padilla J., Redpath S. M., Mougeot F. 2011. The ornament–condition relationship varies with parasite abundance at population level in a female bird. Naturwissenschaften 98, 897–902 10.1007/s00114-011-0833-9 (doi:10.1007/s00114-011-0833-9) [DOI] [PubMed] [Google Scholar]
- 124.Martínez-Padilla J., Vergara P., Pérez-Rodríguez L., Mougeot F., Casas F., Ludwig S. C., Haines J. A., Zeineddine M., Redpath S. M. 2011. Condition- and parasite-dependent expression of a male-like trait in a female bird. Biol. Lett. 7, 364–367 10.1098/rsbl.2010.0991 (doi:10.1098/rsbl.2010.0991) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Weiss S. L. 2006. Female-specific color is a signal of quality in the striped plateau lizard (Sceloporus virgatus). Behav. Ecol. 17, 726–732 10.1093/beheco/arl001 (doi:10.1093/beheco/arl001) [DOI] [Google Scholar]
- 126.Kriner E., Schwabl H. 1991. Control of winter song and territorial aggression of female robins (Erithacus rubecula) by testosterone. Ethology 87, 37–44 10.1111/j.1439-0310.1991.tb01186.x (doi:10.1111/j.1439-0310.1991.tb01186.x) [DOI] [PubMed] [Google Scholar]
- 127.Eens M., Van Duyse E., Berghman L., Pinxten R. 2000. Shield characteristics are testosterone-dependent in both male and female moorhens. Horm. Behav. 37, 126–134 10.1006/hbeh.1999.1569 (doi:10.1006/hbeh.1999.1569) [DOI] [PubMed] [Google Scholar]
- 128.Johnson K. 1988. Sexual selection in pinyon jays. II. Male choice and female–female competition . Anim. Behav. 36, 1048–1053 10.1016/S0003-3472(88)80064-2 (doi:10.1016/S0003-3472(88)80064-2) [DOI] [Google Scholar]
- 129.Midamegbe A., Grégoire A., Perret P., Doutrelant C. 2011. Female–female aggressiveness is influenced by female coloration in blue tits. Anim. Behav. 21, 226–233 10.1016/j.anbehav.2011.04.020 (doi:10.1016/j.anbehav.2011.04.020) [DOI] [Google Scholar]
- 130.Crowley C. E., Magrath R. D. 2004. Shields of offence: signalling competitive ability in the dusky moorhen, Gallinula tenebrosa. Aust. J. Zool. 52, 463–474 10.1071/ZO04013 (doi:10.1071/ZO04013) [DOI] [Google Scholar]
- 131.Rubenstein D. R. 2012. Family feuds: social competition and sexual conflict in complex societies. Phil. Trans. R. Soc. B 367, 2304–2313 10.1098/rstb.2011.0283 (doi:10.1098/rstb.2011.0283) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.McComb K., Packer C., Pusey A. 1994. Roaring and numerical assessment in contests between groups of female lions Panthera leo. Anim. Behav. 47, 379–387 10.1006/anbe.1994.1052 (doi:10.1006/anbe.1994.1052) [DOI] [Google Scholar]
- 133.Seddon N., Tobias J. A. 2003. Communal singing in the cooperatively breeding subdesert mesite Monias benschi: evidence of numerical assessment? J. Avian Biol. 34, 72–80 10.1034/j.1600-048X.2003.03067.x (doi:10.1034/j.1600-048X.2003.03067.x) [DOI] [Google Scholar]
- 134.Seddon N., Tobias J. A., Alvarez A. 2002. Vocal communication in the pale winged trumpeter (Psophia leucoptera): repertoire, context and functional reference. Behaviour 139, 1331–1359 10.1163/156853902321104190 (doi:10.1163/156853902321104190) [DOI] [Google Scholar]
- 135.Hoelzer G. A. 1989. The good parent process of sexual selection. Anim. Behav. 38, 1067–1078 10.1016/S0003-3472(89)80146-0 (doi:10.1016/S0003-3472(89)80146-0) [DOI] [Google Scholar]
- 136.Nisbet I. C. T. 1973. Courtship-feeding, egg size and breeding success in common terns. Nature 241, 141–142 10.1038/241141a0 (doi:10.1038/241141a0) [DOI] [Google Scholar]
- 137.Griffith S. C., Pryke S. R. 2006. Benefits to female birds of assessing color displays. In Bird coloration, volume 2: function and evolution (eds Hill G. E., McGraw K. J.), pp. 233–279 Cambridge, MA: Harvard University Press [Google Scholar]
- 138.Møller A. P., Jennions M. D. 2001. How important are direct fitness benefits of sexual selection? Naturwissenschaften 88, 401–415 10.1007/s001140100255 (doi:10.1007/s001140100255) [DOI] [PubMed] [Google Scholar]
- 139.Smiseth P. T., Amundsen T. 2000. Does female plumage coloration signal parental quality? A male removal experiment with the bluethroat (Luscinia s. svecica). Behav. Ecol. Sociobiol. 47, 205–212 10.1007/s002650050657 (doi:10.1007/s002650050657) [DOI] [Google Scholar]
- 140.Massaro M., Davis L. S., Darby J. T. 2003. Carotenoid-derived ornaments reflect parental quality in male and female yellow-eyed penguins (Megadyptes antipodes). Behav. Ecol. Sociobiol. 55, 169–175 10.1007/s00265-003-0683-3 (doi:10.1007/s00265-003-0683-3) [DOI] [Google Scholar]
- 141.Hager S. B. 2001. The role of nuptial coloration in female Holbrookia maculata: evidence for a dual signaling system. J. Herpetol. 35, 624–632 10.2307/1565901 (doi:10.2307/1565901) [DOI] [Google Scholar]
- 142.Chan R., Stuart-Fox D., Jessop T. 2009. Why are females ornamented? A test of the courtship stimulation and courtship rejection hypotheses. Behav. Ecol. 20, 1334–1342 10.1093/beheco/arp136 (doi:10.1093/beheco/arp136) [DOI] [Google Scholar]
- 143.LeBas N. R., Marshall N. J. 2000. The role of colour in signalling and mate choice in the agamid lizard Ctenophorus ornatus. Proc. R. Soc. Lond. B 267, 445–452 10.1098/rspb.2000.1020 (doi:10.1098/rspb.2000.1020) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Van Gossum H., Stoks R., De Bruyn L. 2001. Male mate choice for female colour morphs: frequency and method dependence. Anim. Behav. 61, F31–F34 10.1006/anbe.2000.1555 (doi:10.1006/anbe.2000.1555) [DOI] [PubMed] [Google Scholar]
- 145.Robertson A. M. 1985. Female dimorphism and mating behaviour in a damselfly Ischnura ramburi: females mimicking males. Anim. Behav. 33, 805–809 10.1016/S0003-3472(85)80013-0 (doi:10.1016/S0003-3472(85)80013-0) [DOI] [Google Scholar]
- 146.Arnqvist G., Rowe L. 2005. Sexual conflict. Princeton, NJ: Princeton University Press [Google Scholar]
- 147.Daly J., Myers C. 1967. Toxicity of Panamanian poison frogs (Dendrobates): some biological and chemical aspects. Science 156, 970–973 10.1126/science.156.3777.970 (doi:10.1126/science.156.3777.970) [DOI] [PubMed] [Google Scholar]
- 148.Summers K., Symula R., Clough M., Cronin T. 1999. Visual mate choice in poison frogs. Proc. R. Soc. Lond. B 266, 2141–2145 10.1098/rspb.1999.0900 (doi:10.1098/rspb.1999.0900) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Murphy T. G. 2006. Predator-elicited visual signal: why the turquoise-browed motmot wag-displays its racketed tail. Behav. Ecol. 17, 547–553 10.1093/beheco/arj064 (doi:10.1093/beheco/arj064) [DOI] [Google Scholar]
- 150.Murphy T. G. 2007. Racketed tail of the male and female turquoise-browed motmot: male but not female tail length correlates with pairing success, performance, and reproductive success. Behav. Ecol. Sociobiol. 61, 911–918 10.1007/s00265-006-0320-z (doi:10.1007/s00265-006-0320-z) [DOI] [Google Scholar]
- 151.Jones I. L., Hunter F. M. 1999. Experimental evidence for mutual inter- and intrasexual selection favouring a crested auklet ornament. Anim. Behav. 57, 521–528 10.1006/anbe.1998.1012 (doi:10.1006/anbe.1998.1012) [DOI] [PubMed] [Google Scholar]
- 152.Jones I. L. 1990. Plumage variability functions for status signaling in least auklets. Anim. Behav. 39, 967–975 10.1016/s0003-3472(05)80962-5 (doi:10.1016/s0003-3472(05)80962-5). [DOI] [Google Scholar]
- 153.Johnsen T. S., Hengeveld J. D., Blank J. L., Yasukawa K., Nolan V. 1996. Epaulet brightness and condition in female red-winged blackbirds. Auk 113, 356–362 [Google Scholar]
- 154.Dobson F. S., Couchoux C., Jouventin P. 2011. Sexual selection on a coloured ornament in king penguins. Ethology 117, 872–879 10.1111/j.1439-0310.2011.01940.x (doi:10.1111/j.1439-0310.2011.01940.x) [DOI] [Google Scholar]
- 155.Bretagnolle V., Lequette B. 1990. Structural variation in the call of the Cory's shearwater, Calonectris diomedea. Ethology 85, 313–323 10.1111/j.1439-0310.1990.tb00410.x (doi:10.1111/j.1439-0310.1990.tb00410.x) [DOI] [Google Scholar]
- 156.Bretagnolle V. 1996. Acoustic communication in a group of nonpasserine birds, the Petrels. In Ecology and Evolution of Acoustic Communication in Birds (eds Kroodsma D. E., Miller E. H.), pp. 160–177. Ithaca, NY: Cornell University Press. [Google Scholar]
- 157.Silva N., Aviles J. M., Danchin E., Parejo D. 2008. Informative content of multiple plumage-coloured traits in female and male European Rollers. Behav. Ecol. Sociobiol. 62, 1969–1979 10.1007/s00265-008-0628-y (doi:10.1007/s00265-008-0628-y). [DOI] [Google Scholar]
- 158.Hunt S., Cuthill I. C., Bennett A. T. D., Griffiths R. 1999. Preferences for ultraviolet partners in the blue tit. Anim. Behav. 58, 809–815 10.1006/anbe.1999.1214 (doi:10.1006/anbe.1999.1214) [DOI] [PubMed] [Google Scholar]
- 159.Rémy A., Grégoire A., Perret P., Doutrelant C. 2010. Mediating male–male interactions: the role of the UV blue crest coloration in blue tits. Behav. Ecol. Sociobiol. 64, 1839–1847 10.1007/s00265-010-0995-z (doi:10.1007/s00265-010-0995-z) [DOI] [Google Scholar]
- 160.Vedder O., Schut E., Magrath M. J. L., Komdeur J. 2010. Ultraviolet crown colouration affects contest outcomes among male blue tits, but only in the absence of prior encounters. Funct. Ecol. 24, 417–425 10.1111/j.1365-2435.2009.01660.x (doi:10.1111/j.1365-2435.2009.01660.x) [DOI] [Google Scholar]
- 161.Kraaijeveld K., Gregurke J., Hall C., Komdeur J., Mulder R. A. 2004. Mutual ornamentation, sexual selection, and social dominance in the black swan. Behav. Ecol. 15, 380–389 10.1093/beheco/arh023 (doi:10.1093/beheco/arh023) [DOI] [Google Scholar]
- 162.Pryke S. R., Griffith S. C. 2006. Red dominates black: agonistic signalling among head morphs in the colour polymorphic Gouldian finch. Proc. R. Soc. B 273, 949–957 10.1098/rspb.2005.3362 (doi:10.1098/rspb.2005.3362) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Garamszegi L. Z., Rosivall B., Hegyi G., Szöllősi E., Török J., Eens M. 2006. Determinants of male territorial behaviour in a Hungarian collared flycatcher population: plumage traits of residents and challengers. Behav. Ecol. Sociobiol. 60, 663–671 10.1007/s00265-006-0210-4 (doi:10.1007/s00265-006-0210-4) [DOI] [Google Scholar]
- 164.Hegyi G., Garamszegi L. Z., Eens M., Török J. 2008. Female ornamentation and territorial conflicts in collared flycatchers (Ficedula albicollis). Naturwissenschaften 95, 993–996 10.1007/s00114-008-0408-6 (doi:10.1007/s00114-008-0408-6) [DOI] [PubMed] [Google Scholar]
- 165.de Heij M. E., Gustafsson L., Brommer J. E. 2011. Experimental manipulation shows that the white wing patch in collared flycatchers is a male sexual ornament. Ecol. Evol. 1, 546–555 10.1002/ece3.48 (doi:10.1002/ece3.48) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Lyon B. E. 2003. Ecological and social constraints on conspecific brood parasitism by nesting female American coots (Fulica americana). J. Anim. Ecol. 72, 47–60 10.1046/j.1365-2656.2003.00674.x (doi:10.1046/j.1365-2656.2003.00674.x). [DOI] [Google Scholar]
- 167.Sæther S. A. 2002. Female calls in lek-mating birds: indirect mate choice, female competition for mates, or direct mate choice? Behav. Ecol. 13, 344–352 10.1093/beheco/13.3.344 (doi:10.1093/beheco/13.3.344) [DOI] [Google Scholar]
- 168.Sæther S. A., Fiske P., Kalas J. A. 2001. Male mate choice, sexual conflict and strategic allocation of copulations in a lekking bird. Proc. R. Soc. Lond. B 268, 2097–2102 10.1098/rspb.2001.1745 (doi:10.1098/rspb.2001.1745) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Petrie M. 1983. Female moorhens compete for small fat males. Science 220, 413–415 10.1126/science.220.4595.413 (doi:10.1126/science.220.4595.413). [DOI] [PubMed] [Google Scholar]
- 170.Petrie M. 1984. Territory size in the moorhen (Gallinula chloropus)—an outcome of RHP asymmetry between neighbors. Anim. Behav. 32, 861–870 10.1016/s0003-3472(84)80163-3 (doi:10.1016/s0003-3472(84)80163-3). [DOI] [Google Scholar]
- 171.Johnson K. 1988. Sexual selection in pinyon jays I: female choice and male-male competition. Anim. Behav. 36, 1038–1047 10.1016/S0003-3472(88)80063-0 (doi:10.1016/S0003-3472(88)80063-0) [DOI] [Google Scholar]
- 172.Møller A. P. 1993. Sexual selection in the barn swallow Hirundo rustica III. Female tail ornaments. Evolution 47, 417–431 10.2307/2410061 (doi:10.2307/2410061) [DOI] [PubMed] [Google Scholar]
- 173.Møller A. P., Barbosa A., Cuervo J. J., de Lope F., Merino S., Saino N. 1998. Sexual selection and tail streamers in the barn swallow. Proc. R. Soc. Lond. B 265, 409–414 10.1098/rspb.1998.0309 (doi:10.1098/rspb.1998.0309) [DOI] [Google Scholar]
- 174.Tobias J. A., Seddon N. 2009. Signal design and perception in Hypocnemis antbirds: evidence for convergent evolution via social selection. Evolution 63, 3168–3189 10.1111/j.1558-5646.2009.00795.x (doi:10.1111/j.1558-5646.2009.00795.x) [DOI] [PubMed] [Google Scholar]
- 175.Mays H. L., Hopper K. R. 2004. Differential responses of yellow-breasted chats, Icteria virens, to male and female conspecific model presentations. Anim. Behav. 67, 21–26 10.1016/j.anbehav.2003.01.003 (doi:10.1016/j.anbehav.2003.01.003). [DOI] [Google Scholar]
- 176.Johnsen A., Lifjeld J. T., Andersson S., Ornborg J., Amundsen T. 2001. Male characteristics and fertilisation success in bluethroats. Behaviour 138, 1371–1390 10.1163/156853901317367645 (doi:10.1163/156853901317367645) [DOI] [Google Scholar]
- 177.Arcese P., Smith J. N. M. 1985. Phenotypic correlates and ecological consequences of dominance in song sparrows. J. Anim. Ecol. 54, 817–830 10.2307/4380 (doi:10.2307/4380). [DOI] [Google Scholar]
- 178.Bretagnolle V., Genevois F., Mougeot F. 1998. Intra and intersexual function in the call of a non-passerine bird. Behaviour 135, 1161–1184 10.1163/156853998792913555 (doi:10.1163/156853998792913555) [DOI] [Google Scholar]
- 179.Järvi W. T., Bakken M. A. 1984. The function of the variation in the breast-stripe of the great tit (Parus major). Anim. Behav. 32, 590–596 10.1016/S0003-3472(84)80296-1 (doi:10.1016/S0003-3472(84)80296-1) [DOI] [Google Scholar]
- 180.Griggio M., Serra L., Licheri D., Monti A., Pilastro A. 2007. Armaments and ornaments in the rock sparrow: a possible dual utility of a carotenoid based feather signal. Behav. Ecol. Sociobiol. 61, 423–433 10.1007/s00265-006-0270-5 (doi:10.1007/s00265-006-0270-5) [DOI] [Google Scholar]
- 181.Gowaty P. A. 1981. Aggression of breeding eastern bluebirds (Sialia sialis) toward their mates and models of intraspecific and interspecific intruders. Anim. Behav. 29, 1013–1027 10.1016/s0003-3472(81)80055-3 (doi:10.1016/s0003-3472(81)80055-3). [DOI] [Google Scholar]
- 182.Siefferman L., Hill G. E. 2005. Evidence for sexual selection on structural plumage coloration in female eastern bluebirds (Sialia sialis). Evolution 59, 1819–1828 10.1111/j.0014-3820.2005.tb01828.x (doi:10.1111/j.0014-3820.2005.tb01828.x) [DOI] [PubMed] [Google Scholar]
- 183.Rowe K. M. C., Weatherhead P. J. 2011. Assortative mating in relation to plumage traits shared by male and female American robins. Condor 113, 881–889 10.1525/cond.2011.100207 (doi:10.1525/cond.2011.100207) [DOI] [Google Scholar]
- 184.Roulin A. 1999. Nonrandom pairing by male barn owls (Tyto alba) with respect to a female plumage trait. Behav. Ecol. 10, 688–695 10.1093/beheco/10.6.688 (doi:10.1093/beheco/10.6.688). [DOI] [Google Scholar]
- 185.Roulin A., Riols C., Dijkstra C., Ducrest A. L. 2001. Female plumage spottiness signals parasite resistance in the barn owl (Tyto alba). Behav. Ecol. 12, 103–110 10.1093/oxfordjournals.beheco.a000371 (doi:10.1093/oxfordjournals.beheco.a000371) [DOI] [Google Scholar]
- 186.Chaine A. S., Tjernell K. A., Shizuka D., Lyon B. E. 2011. Sparrows use multiple status signals in winter social flocks. Anim. Behav. 81, 447–453 10.1016/j.anbehav.2010.11.016 (doi:10.1016/j.anbehav.2010.11.016). [DOI] [Google Scholar]
- 187.Watt D. J. 1986. A comparative-study of status signaling in sparrows (genus Zonotrichia). Anim. Behav. 34, 1–15 10.1016/0003-3472(86)90001-1 (doi:10.1016/0003-3472(86)90001-1) [DOI] [Google Scholar]
- 188.Roberts S. C., Dunbar R. I. M. 2000. Female territoriality and the function of scent-marking in a monogamous antelope (Oreotragus oreotragus). Behav. Ecol. Sociobiol. 47, 417–423 10.1007/s002650050685 (doi:10.1007/s002650050685). [DOI] [Google Scholar]
- 189.Pizzolon M., Rasotto M. B., Mazzoldi C. 2008. Male lagoon gobies, Knipowitschia panizzae, prefer more ornamented to larger females. Behav. Ecol. Sociobiol. 62, 521–528 10.1007/s00265-007-0476-1 (doi:10.1007/s00265-007-0476-1). [DOI] [Google Scholar]
- 190.Brandtmann G., Scandura M., Trillmich F. 1999. Female–female conflict in the harem of a snail cichlid (Lamprologus ocellatus): behavioural interactions and fitness consequences. Behaviour 136, 1123–1144 10.1163/156853999501793 (doi:10.1163/156853999501793). [DOI] [Google Scholar]
- 191.Wong M. Y. L., Munday P. L., Buston P. M., Jones G. P. 2008. Monogamy when there is potential for polygyny: tests of multiple hypotheses in a group-living fish. Behav. Ecol. 19, 353–361 10.1093/beheco/arm141 (doi:10.1093/beheco/arm141). [DOI] [Google Scholar]
- 192.Baldauf S. A., Bakker T. C. M., Kullmann H., Thunken T. 2011. Female nuptial coloration and its adaptive significance in a mutual mate choice system. Behav. Ecol. 22, 478–485 10.1093/beheco/arq226 (doi:10.1093/beheco/arq226) [DOI] [Google Scholar]
- 193.Cuadrado M. 2000. Body colors indicate the reproductive status of female common chameleons: experimental evidence for the intersex communication function. Ethology 106, 79–91 10.1046/j.1439-0310.2000.00493.x (doi:10.1046/j.1439-0310.2000.00493.x). [DOI] [Google Scholar]
- 194.Baird T. A., Sloan C. L. 2003. Interpopulation variation in the social organization of female collared lizards, Crotaphytus collaris. Ethology 109, 879–894 10.1046/j.0179-1613.2003.00925.x (doi:10.1046/j.0179-1613.2003.00925.x). [DOI] [Google Scholar]
- 195.Baird T. A. 2004. Reproductive coloration in female collared lizards, Crotophytus collaris, stimulates courtship by males. Herpetologica 60, 337–348 10.1655/03-17 (doi:10.1655/03-17). [DOI] [Google Scholar]
- 196.Lappin A. K., Husak J. F. 2005. Weapon performance, not size, determines mating success and potential reproductive output in the collared lizard (Crotaphytus collaris). Am. Nat. 166, 426–436 10.1086/432564 (doi:10.1086/432564). [DOI] [PubMed] [Google Scholar]
- 197.Cooper W. E., Crews D. 1988. Sexual coloration, plasma-concentrations of sex steroid-hormones, and responses to courtship in the female keeled earless lizard (Holbrookia propinqua). Horm. Behav. 22, 12–25 10.1016/0018-506x(88)90027-x (doi:10.1016/0018-506x(88)90027-x). [DOI] [PubMed] [Google Scholar]
- 198.Watkins G. G. 1997. Inter-sexual signalling and the functions of female coloration in the tropidurid lizard Microlophus occipitalis. Anim. Behav. 53, 843–852 10.1006/anbe.1996.0350 (doi:10.1006/anbe.1996.0350). [DOI] [Google Scholar]
- 199.Weiss S. L. 2002. Reproductive signals of female lizards: pattern of trait expression and male response. Ethology 108, 793–813 10.1046/j.1439-0310.2002.00819.x (doi:10.1046/j.1439-0310.2002.00819.x) [DOI] [Google Scholar]
- 200.Fox S. F., Heger N. A., Delay L. S. 1990. Social cost of tail loss in Uta stansburiana: lizard tails as status-signalling badges. Anim. Behav. 39, 549–554 10.1016/S0003-3472(05)80421-X (doi:10.1016/S0003-3472(05)80421-X) [DOI] [Google Scholar]
- 201.Panhuis T. M., Wilkinson G. S. 1999. Exaggerated eyespan influences male contest outcome in stalk-eyed flies. Behav. Ecol. Sociobiol. 46, 221–227 10.1007/s002650050613 (doi:10.1007/s002650050613) [DOI] [Google Scholar]
- 202.Al-khairulla H., Warburton D., Knell R. J. 2003. Do the eyestalks of female diopsid flies have a function in intrasexual aggressive encounters? J. Insect Behav. 16, 679–686 10.1023/B:JOIR.0000007703.84691.ad (doi:10.1023/B:JOIR.0000007703.84691.ad) [DOI] [Google Scholar]
- 203.Vrontou E., Nilsen S., Demir E., Kravitz E. A., Dickson B. J. 2006. fruitless regulates aggression and dominance in Drosophila. Nat. Neurosci. 9, 1469–1471 10.1038/nn1809 (doi:10.1038/nn1809) [DOI] [PubMed] [Google Scholar]
- 204.Rillich J., Buhl E., Schildberger K., Stevenson P. A. 2009. Female crickets are driven to fight by the male courting and calling songs. Anim. Behav. 77, 737–742 10.1016/j.anbehav.2008.12.009 (doi:10.1016/j.anbehav.2008.12.009). [DOI] [Google Scholar]
- 205.Bussiere L. F., Gwynne D. T., Brooks R. 2008. Contrasting sexual selection on males and females in a role-reversed swarming dance fly, Rhamphomyia longicauda Loew (Diptera: Empididae). J. Evol. Biol. 21, 1683–1691 10.1111/j.1420-9101.2008.01580.x (doi:10.1111/j.1420-9101.2008.01580.x). [DOI] [PubMed] [Google Scholar]
- 206.South S. H., Arnqvist G. 2011. Male, but not female, preference for an ornament expressed in both sexes of the polygynous mosquito Sabethes cyaneus. Anim. Behav. 81, 645–651 10.1016/j.anbehav.2010.12.014 (doi:10.1016/j.anbehav.2010.12.014) [DOI] [Google Scholar]
- 207.Vasconcelos R. O., Simões J. M., Almada V. C., Fonseca P. J., Amorim C. P. 2010. Vocal behavior during territorial intrusions in the Lusitanian toadfish: boatwhistles also function as territorial ‘keep-out’ signals. Ethology 116, 155–165 10.1111/j.1439-0310.2009.01722.x (doi:10.1111/j.1439-0310.2009.01722.x) [DOI] [Google Scholar]
- 208.Evans M. R., Hatchwell B. J. 1992. An experimental study of male adornment in the scarlet-tufted malachite sunbird. 1. The role of pectoral tufts in territorial defense. Behav. Ecol. Sociobiol. 29, 413–419 10.1007/BF00170171 (doi:10.1007/BF00170171) [DOI] [Google Scholar]
- 209.Ewald P. W., Rohwer S. 1980. Age, coloration and dominance in nonbreeding hummingbirds: a test of the asymmetry hypothesis. Behav. Ecol. Sociobiol. 7, 273–279 10.1007/BF00300667 (doi:10.1007/BF00300667) [DOI] [Google Scholar]
- 210.Kelsey M. G. 1989. A comparison of the song and territorial behaviour of a long-distance migrant, the Marsh Warbler Acrocephalus palustris, in summer and winter. Ibis 131, 403–414 10.1111/j.1474-919X.1989.tb02788.x (doi:10.1111/j.1474-919X.1989.tb02788.x) [DOI] [Google Scholar]
- 211.Mace T. R. 2000. Time budget and pair-bond dynamics in the Comb-crested Jacana Irediparra gallinacea: a test of hypotheses. Emu 100, 31–41 10.1071/MU9844 (doi:10.1071/MU9844) [DOI] [Google Scholar]
- 212.Gwynne D. T. 1991. Sexual competition among females: what causes courtship-role reversal? Trends Ecol. Evol. 6, 118–121 10.1016/0169-5347(91)90089-G (doi:10.1016/0169-5347(91)90089-G) [DOI] [PubMed] [Google Scholar]
- 213.Regosin J. V., Pruett-Jones S. 2001. Sexual selection and tail-length dimorphism in scissor-tailed flycatchers. Auk 118, 167–175 10.1642/0004-8038(2001)118[0167:SSATLD]2.0.CO;2 (doi:10.1642/0004-8038(2001)118[0167:SSATLD]2.0.CO;2) [DOI] [Google Scholar]
- 214.Ord T. J., Stuart-Fox D. 2006. Ornament evolution in dragon lizards: multiple gains and widespread losses reveal a complex history of evolutionary change. J. Evol. Biol. 19, 797–808 10.1111/j.1420-9101.2005.01050.x (doi:10.1111/j.1420-9101.2005.01050.x) [DOI] [PubMed] [Google Scholar]
- 215.Kapusta J., Marchlewska-Koj A. 1998. Interfemale aggression in adult bank voles (Clethrionomys glareolus). Aggress. Behav. 24, 53–61 (doi:10.1002/(SICI)1098-2337(1998)24:1<53::AID-AB5>3.0.CO;2-T) [DOI] [Google Scholar]
- 216.Holberton R. L., Able K. P., Wingfield J. C. 1989. Status signalling in dark-eyed juncos, Junco hyemalis: plumage manipulations and hormonal correlates of dominance. Anim. Behav. 37, 681–689 10.1016/0003-3472(89)90047-X (doi:10.1016/0003-3472(89)90047-X) [DOI] [Google Scholar]
- 217.Baker R. H., Wilkinson G. S. 2001. Phylogenetic analysis of sexual dimorphism and eyespan allometry in stalk-eyed flies (Diopsidae). Evolution 55, 1373–1385 [DOI] [PubMed] [Google Scholar]
- 218.Edwards A. C., Rollmann S. M., Morgan T. J., Mackay T. F. 2006. Quantitative genomics of aggressive behavior in Drosophila melanogaster. PLoS Genet. 2, e154. 10.1371/journal.pgen.0020154 (doi:10.1371/journal.pgen.0020154) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Irwin R. E. 1994. The evolution of plumage dichromatism in the New World blackbirds. Am. Nat. 144, 890–907 10.1086/285717 (doi:10.1086/285717) [DOI] [Google Scholar]
- 220.Price J. J. 2009. Evolution and life-history correlates of female song in the New World blackbirds. Behav. Ecol. 20, 967–977 10.1093/beheco/arp085 (doi:10.1093/beheco/arp085) [DOI] [Google Scholar]
- 221.Bleiweiss R. 1985. Iridescent polychromatism in a female hummingbird: is it related to feeding strategies? Auk 102, 701–713 [Google Scholar]
- 222.Hall M. L., Magrath R. D. 2007. Temporal coordination signals coalition quality. Curr. Biol. 17, R406–R407 10.1016/j.cub.2007.04.022 (doi:10.1016/j.cub.2007.04.022) [DOI] [PubMed] [Google Scholar]
- 223.Tibbetts E. A., Safran R. J. 2009. Co-evolution of plumage characteristics and winter sociality in New and Old World sparrows. J. Evol. Biol. 22, 2376–2386 10.1111/j.1420-9101.2009.01861.x (doi:10.1111/j.1420-9101.2009.01861.x) [DOI] [PubMed] [Google Scholar]
- 224.Keller L., Ross K. G. 1998. Selfish genes: a green beard in the red fire ant. Nature 394, 573–575 10.1038/29064 (doi:10.1038/29064) [DOI] [Google Scholar]
- 225.Huxley J. S. 1938. Darwin's theory of sexual selection and the data subsumed by it, in the light of recent research. Am. Nat. 72, 416–433 10.1086/280795 (doi:10.1086/280795) [DOI] [Google Scholar]
- 226.Langmore N. E. 1998. Functions of duet and solo songs of female birds. Trends Ecol. Evol. 13, 136–140 10.1016/S0169-5347(97)01241-X (doi:10.1016/S0169-5347(97)01241-X) [DOI] [PubMed] [Google Scholar]
- 227.Filardi C. E., Smith C. E. 2008. Social selection and geographic variation in two monarch flycatchers from the Solomon Islands. Condor 110, 24–34 10.1525/cond.2008.110.1.24 (doi:10.1525/cond.2008.110.1.24) [DOI] [Google Scholar]
- 228.Price J. J., Lanyon S. M., Omland K. E. 2009. Losses of female song with changes from tropical to temperate breeding in the New World blackbirds. Proc. R. Soc. B 276, 1971–1980 10.1098/rspb.2008.1626 (doi:10.1098/rspb.2008.1626) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Long T. A. F., Pischedda A., Stewart A. D., Rice W. R. 2009. A cost of sexual attractiveness to high-fitness females. PLoS Biol. 7, e1000254. 10.1371/journal.pbio.1000254 (doi:10.1371/journal.pbio.1000254) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Wolf J. B., Brodie E. D., Cheverud J. M., Moore A. J., Wade M. J. 1998. Evolutionary consequences of indirect genetic effects. Trends Ecol. Evol. 13, 64–69 10.1016/S0169-5347(97)01233-0 (doi:10.1016/S0169-5347(97)01233-0) [DOI] [PubMed] [Google Scholar]