

Abstract
Social selection offers an alternative to sexual selection by reversing its logic. Social selection starts with offspring production and works back to mating, and starts with behavioural dynamics and works up to gene pool dynamics. In social selection, courtship can potentially be deduced as a negotiation, leading to an optimal allocation of tasks during offspring rearing. Ornaments facilitate this negotiation and also comprise ‘admission tickets’ to cliques. Mating pairs may form ‘teams’ based on the reciprocal sharing of pleasure. The parent–offspring relation can be managed by the parent considered as the owner of a ‘family firm’ whose product is offspring. The cooperation in reproductive social behaviour evolves as a mutual direct benefit through individual selection rather than as some form of altruism requiring kin or multi-level selection.
Keywords: cooperation, pleasure, nest production function, courtship semantics, teamwork, family firm
1. Introduction
Social selection differs from sexual selection in two respects, as illustrated in figures 1 and 2. First, the modelling of mating systems starts from the stage of offspring production and works back from there to earlier life-history stages, including courtship. Second, the modelling of behaviour starts with cooperative as well as competitive game theory and works up from there to gene pool dynamics. The logical progression in social selection is the opposite to that in sexual selection and is motivated by the modelling approaches of evolutionary and population ecology. The name ‘social selection’ as used here indicates the hypothesis that the adaptive function of choosing mates and other actions taken during reproductive social behaviour is to fashion the social infrastructure from which offspring emerge [1–3].
Figure 1.
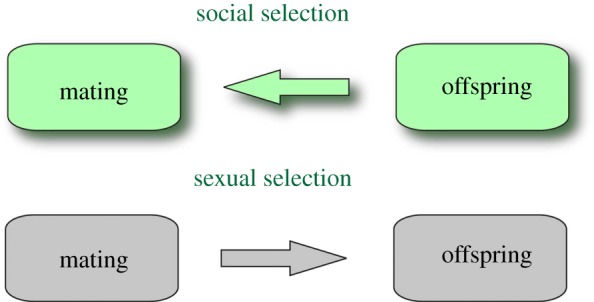
Direction of inference for mating systems theory.
Figure 2.
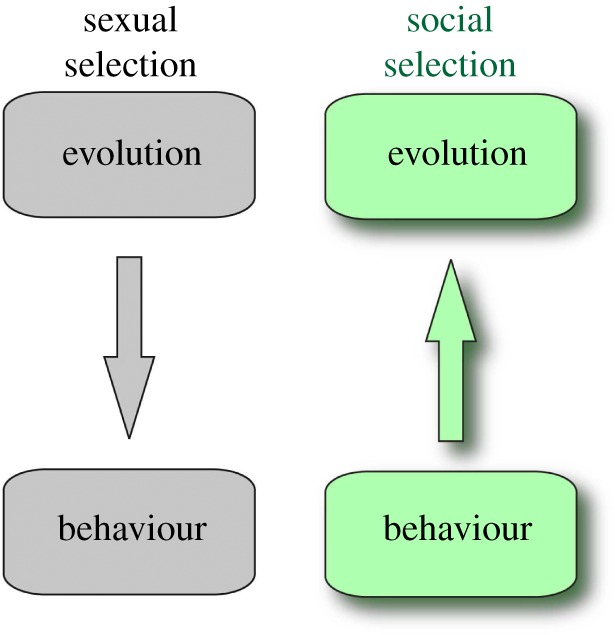
Top-down versus bottom-up modelling of social evolution.
The phrase ‘social selection’ has been previously used by West-Eberhard [4, p. 157] who writes, ‘The special characteristics of sexual selection discussed by Darwin apply as well for social competition for resources other than mates’. She describes social competition as ‘competition in which an individual must win in interactions or comparisons with conspecific rivals in order to gain access to some resource. The contested resources might include food, hibernation space, nesting material, mates or places to spend the night. Seen in this broader perspective, sexual selection refers to the subset of social competition in which the resource at stake is mates. And social selection is differential success … in social competition’. (Italics in original.) Thus, sexual selection is a subset of social selection sensu West-Eberhard.
Moore et al. [5, p. 5188] have since broadened West-Eberhard's definition by writing, ‘selection reflecting associations between fitness and social behaviour has been termed ‘social selection”. Similarly, Frank [6, p. 358] writes, ‘I continue to use the word ‘social’ in the broadest way, to cover all aspects of evolutionary change that deal with the tension between conflict and cooperation'. Still another definition of social selection has been proposed in connection to the evolution of animal communication by Tanaka [7, p. 512] who writes, ‘I define social selection as the selective force that arises when a signal influences the fitness of signalers or both the fitness of signalers and receivers’.
2. The mating system as a life-history stage
Concerning figure 1, social selection (sensu Roughgarden) views mating systems as a stage in an optimal life history. Finding optimal life histories relies on techniques from optimal control theory and is often achieved using backward induction [8,9]. To illustrate, suppose a control variable for a plant is to allocate newly fixed photosynthate into root versus shoot. Backwards induction means solving for the optimal root/shoot allocation at some terminal time, T, at the end of the growing season. Then, one moves back a step to find what the optimal root/shoot allocation is at time T−1 given that the optimal decision at time T has already been determined. Gradually, one works back to the initial decision.
Unlike the optimal life histories usually studied in ecology, with the focus on the optimal strategy of an isolated individual, two parties are involved in mating. Therefore, one begins with the outcome of a game between two potential parents at time T, then works back to the outcome at time T−1, culminating in the outcome of a game between the potential parents at the time of courtship or earlier. Biologically, this approach would first consider the local ecological situation into which the offspring are released from their parents and determine what actions by them will yield to each the largest number of its offspring at that time. Then, second, the signalling and negotiation needed during courtship to bring about those actions would be calculated. Thereafter, with these solution in hand, one could play the predictions forward and observe the optimal courtship followed by the optimal parental care.
(a). Optimal parental care
An illustration of the first step in the social selection programme pertains to how the production of young from a nest depends on the time allocations of both the male and the female each to two activities, say, bringing food to the young and patrolling for predators ([3, pp. 198–201], [10]). The formula that describes how many offspring are successfully reared depending on the fraction of the day the male and female each spend foraging versus patrolling for predators is a ‘production function’ for the nest—it is an empirical formula potentially measurable in the field. Using the nest-production function, one can solve for the best joint allocation of effort into the two activities by the male and the female.
To illustrate, suppose the female spends tf feeding and 1−tf patrolling, and the male spends tm feeding and 1−tm patrolling. Consider how the male's and female's work can be combined to yield the nest's production. One possibility is that the male and female each raise some young, which are then pooled to yield a total production. Suppose that the contribution to nestling rearing from the female is tf(1−tf) and that from the male is tm(1−tm). That is, the young that each rears for their common pool is the product of each's foraging and patrolling times—the young die if not fed or if predators attack, so the product formula captures the need for both these activities. The overall nest production is then the sum of both the male and female contributions, 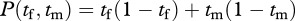 . In this case, the birds are pooling separately produced products. This sum is the nest production function. According to this formula, the number of young reared by the female is maximized when she spends 50 per cent of her time foraging and 50 per cent of her time patrolling, and similarly for the male.
. In this case, the birds are pooling separately produced products. This sum is the nest production function. According to this formula, the number of young reared by the female is maximized when she spends 50 per cent of her time foraging and 50 per cent of her time patrolling, and similarly for the male.
Another possibility is to imagine that the male and female pool effort towards raising a common clutch of young, in which case the production would be 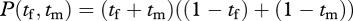 . Here, they are pooling effort to produce a common product. Because the male and female efforts are interchangeable in the formula, all that matters is that the sum of tf and tm should be equal to 50 per cent of their combined efforts. A split in which both do equal amounts of foraging and patrolling is just as good as a split in which one does all the foraging and the other does all the patrolling. As long as the total effort by both to foraging equals the total effort by both to patrolling, their common clutch size is maximized.
. Here, they are pooling effort to produce a common product. Because the male and female efforts are interchangeable in the formula, all that matters is that the sum of tf and tm should be equal to 50 per cent of their combined efforts. A split in which both do equal amounts of foraging and patrolling is just as good as a split in which one does all the foraging and the other does all the patrolling. As long as the total effort by both to foraging equals the total effort by both to patrolling, their common clutch size is maximized.
Still another possibility is that the nest production function is a mixture of these two extremes. Perhaps the male bestows special attention on some of the nestlings, and the female bestows hers on the others, but they still tend all the nestlings to some degree. If the mixture between pooling privately tended young and jointly tended young is, say, 50 : 50, then the nest production function becomes 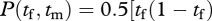
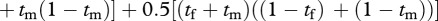 . The best production is again realized when the female and male each spend 50 per cent of their time foraging and 50 per cent patrolling.
. The best production is again realized when the female and male each spend 50 per cent of their time foraging and 50 per cent patrolling.
If the male is disadvantaged in his foraging however, or experimentally handicapped by attaching a weight to its leg, the nest production function might become 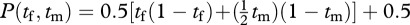
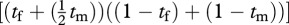 . This production function is maximized when the female spends 65 per cent of her time foraging and the male only 26 per cent of his time foraging.
. This production function is maximized when the female spends 65 per cent of her time foraging and the male only 26 per cent of his time foraging.
Thus, the local ecological situation, together with the abilities of the parents, determines the optimal time allocation of each to accomplish their joint task of raising offspring. In general, the ecological situation might feature high or low predation risk, clumped or dispersed resources, etc. Depending on ecological details and the physiological abilities of each sex, the optimal division of labour from the nest production function might be to rear together as above or, not illustrated, for one or the other sex to rear exclusively or perhaps for neither sex to provide any parental care at all.
(b). Optimal courtship
According to social selection, ornaments and armaments evolve as signalling traits to implement the communication needed to establish the social infrastructure from which offspring emerge.
If the optimal parental effort is to rear jointly, then courtship could result in an agreement between two members of a breeding pair for each to carry out the optimal allocation to tasks involved in offspring rearing. By this hypothesis, courtship represents the negotiation of a ‘pre-nuptial agreement’. (I thank Richard Prum for this term.) Examples might include the courtship dance of the Laysan albatross [11] and the negotiation of nest sites in blue-footed boobies [12].
To implement the idea of courtship as a negotiation, one assumes a system of ‘courtship semantics’ that assigns a meaning to each of the moves during a courtship dance. Suppose the two potentially cooperating mates are negotiating for an optimal allocation, tf and tm, from the nest production function for their ecological situation. In particular, suppose a male is receiving signals from a female based on her experience in the habitat about what her ‘opinion’ for tf would be. The initial offer might be taken as 50 per cent. The male might then signal that, from his experience in the habitat, in his opinion the female should lower her foraging time tf and spend more time at the nest to achieve the highest nest production. If she agrees to the suggestion, then she might signal agreement, or if not, signal otherwise. The male could then disengage or accept the female's decision by signalling agreement. Thereafter, the female could receive signals from him concerning his opinion about what his allocation should be. She might signal that she feels he should spend more time foraging and less time at the nest to achieve the highest nest production. If he agrees, then he would signal agreement. And so forth. A visual animation can illustrate hypothetical dances that culminate in an agreed tf and tm, or in other instances can illustrate the failure to agree, whereupon the parties move on to court other potential mates who share their opinion of the best task allocations. In this approach, courtship brings about a social infrastructure, however simple or complex, in which offspring grow and mature. In this situation, ornaments and associated behaviour might function as the vocabulary needed to carry out pre-nuptial negotiation. This idea is the subject of unpublished research in my laboratory.
Alternatively, the nest production function might not admit an optimal solution in which both members of a breeding pair raise the young, and instead, the rearing is carried out by only one sex. In this case, the ornaments in either male or female might serve as ‘admission tickets’ to power-holding cliques that control the opportunity for offspring production ([1, pp. 150, 178–181], [3, pp. 242–243]) Examples might include the exaggerated traits of males in lekking species like the peacock [13] and the penis in female spotted hyenas [14]. Admission tickets are demanded by members of a clique as a condition for participation in the clique. The selection pressure for such traits is strong because not possessing them is reproductively lethal. The tickets must be expensive to prevent new members from entering the clique and diluting its power of monopoly. Admission-ticket ornaments are the intra-sexual counterpart of exaggerated traits produced by inter-sexual selection according to Fisher's runaway selection scheme [15,16] that postulates an escalating coevolution of female preference and male extravagance. Although Fisher's runaway trait exaggeration is ultimately countered by natural selection, exaggeration in an admission ticket is inherently limited by the benefit conferred from membership in the clique—a trait should be no more expensive than what can be offset by the benefit of membership. Iyer [17, ch. 5]) has investigated the conditions for the evolution of an admission ticket, showing among her results that a polymorphism can form between those with ornaments on the ‘inside’ and those without ornaments on the ‘outside’. This result is suggestive of the many species such as coho salmon [18] and the plainfin midshipman [19] that have two types of males—those with ornaments that hold territories in a lek and those without ornaments who intrude into the territories to find mates.
Prum [20] has suggested that manakins are possible examples of Fisher's runaway selection. Another possibility is that exaggerated characters in manakins are admission tickets to male cliques.
Another courtship topic is the solicitation of extra-pair matings. The social selection position suggests that these matings serve social purposes such as advertising social networks and alliances when the matings are carried out in open view, as in razorbills [21,22]. The extra-pair matings also promote a cooperative distribution of eggs among adjacent nests, equalizing productivity differentials between nest pairs and spreading risk so that a bird literally does not have to place all its eggs in one basket ([1, pp. 119–121], [3,23, pp. 209–233]).
A feature of social selection is that courtship dynamics and the evolution of ornaments are logically subordinated to an extended theory of parental investment. Shuker [24, p. E5] has commented that ‘much of the context given for this alternative modeling framework sits firmly in the field of the evolution of parental care, not sexual selection at all … parental investment need have no a priori link to the pattern of sexual selection’. Shuker's comment notwithstanding, social selection's approach to courtship and mating behaviour is to focus on what is needed for offspring production in various ecological situations and to work back to whatever mating system is needed to realize that degree of offspring production.
3. Two tiers in social evolution
Turning to figure 2, social selection envisions a bottom-up logic for the evolution of behaviour. Population ecology has successfully derived equations for population dynamics from properties of individuals, an approach termed variously, ‘individual-based models’, ‘individual oriented models’, ‘agent-based models’ or ‘mechanism-based models’. Population-dynamic models that are developed bottom-up from individuals to populations offer a more relevant and testable theory than the venerable equations of population ecology such as the logistic or Lotka–Volterra equations that feature a top-down logic.
Bottom-up logic can also be used to develop evolutionary theory, and theory for social evolution in particular. The advantage to a bottom-up approach is that the full range of game theoretic analysis and solution concepts are available to model behaviour, including both cooperative as well as competitive game theory. In contrast, top-down modelling imposes gene-pool competitive dynamics, including the evolutionarily stable strategy (ESS) solution concept [25] on behaviour, whereas it is conceivable that behavioural interactions can lead to other solution concepts such as the Nash bargaining solution (NBS) [26]. Social selection implements a bottom-up logic by introducing two tiers of analysis, a lower tier within which behavioural interactions are modelled on a fast time scale, and a higher tier in which evolutionary dynamics are modelled on a slow population-genetic time scale [3,27].
Within the evolutionary tier the dynamics are unremarkable, although the genetical frequency dependence that emanates from the behavioural tier might lead to polymorphism [28]. Typically, selection simply favours those who have accumulated the highest individual fitness. Those individuals may have acquired their high individual fitness because of how well they participated in social interactions.
Within the behavioural tier, the animals are envisioned to be continually accumulating fitness according to increments measured day by day, hour by hour, event by event, play by play or with some other natural time step in behavioural time. Then, all the increments of fitness accumulated during its life are summed to obtain the full genetic fitness seen by natural selection.
Two schemes of social organization seem particularly relevant to courtship, mating, parental care and family life: a ‘team’ and a ‘firm’.
(a). Teams
A ‘team’ consists of participants who take coordinated actions to achieve a team objective. During teamwork, animals are hypothesized to experience pleasure from physical and/or vocal intimacy. The pursuit of this pleasure is hypothesized to be the proximal motivation for animals to participate in teamwork [3,29,30].
Darwin [31] reviewed the facial and bodily expressions of many animals, especially mammals, and frequently mentioned how pleasure is expressed. He writes, ‘With the lower animals we see the same principle of pleasure derived from contact in association with love. Dogs and cats manifestly take pleasure in rubbing against their masters and mistresses, and in being rubbed or patted by them. Many kinds of monkeys … delight in fondling and being fondled by each other, and by persons to whom they are attached’ [31, p. 215–216]. Darwin also recognized pleasure through song: ‘We can plainly perceive, with some of the lower animals, that the males employ their voices to please the females, and that they themselves take pleasure in their own vocal utterances’ [31, p. 87–88]. The pleasure associated with physical and vocal intimacy that Darwin describes is the key feature to social selection's concept of teamwork.
Neurobiology offers methods for testing for teamwork. The brain centres activated during pleasurable sensations are increasingly well delineated [32–35]. The teamwork should be associated with activity in known neural pleasure centres. Pleasure in teamwork might also be revealed by assays of hormones in the blood, such as oxytocin. This and other neuromodulating polypeptide hormones have been implicated in social phenomena as diverse as flocking behaviour in birds [36] and ethnocentrism in humans ([37], and references cited therein). The assumption that animals choose to participate in teamwork because of seeking pleasure is consistent with studies on humans showing that decision-making is hedonic rather than rational [38–40].
It is further hypothesized that the act of cooperation itself is pleasurable. As a human analogy, consider the difference in pleasure between making an ‘alley-oop pass’ in basketball compared with making two foul shots, both of which yield the same two points. Of course, every member of a basketball team feels some pleasure when two foul shots are successful, but the pleasure experienced is even greater if the two points are obtained with a beautiful acrobatic pass followed by a teammate's dunk at the basket.
Equations contrasting the behavioural dynamics of teamwork with that of individualism were introduced by Roughgarden et al. [29] and Roughgarden [30]. The behavioural state variable is the fraction of time spent in one of two activities during a behavioural time increment. For example, the activities of two birds might be foraging for food versus guarding the nest. How do they decide whether to change their allocations to these activities? They will alter their allocations to enhance their pleasure, or leave them unchanged if no change in allocation is more pleasurable. The birds can alter their allocations either separately as individuals or in unison as a team.
If individualism is pleasurable, the bird experiences improving its own fitness increment as pleasurable independently of how its actions affect the other bird's fitness increment. Each bird's motivation is solely ‘self-regarding’. In seeking pleasure, each bird separately climbs the gradient in its own pleasure function. If both birds continually alter their behaviour individualistically, then their time allocations to foraging and guarding after several time steps converge to an equilibrium that is a behavioural counterpart of the familiar ESS [25,41]. The behavioural-tier counterpart to the ESS is the Nash equilibrium from non-cooperative game theory, here abbreviated as the Nash competitive equilibrium (NCE). At this equilibrium, neither bird will deviate in its time allocation to foraging and guarding, given the allocation of the other bird, because it is not pleasurable to do so.
In contrast, if teamwork is pleasurable, then the pleasure of each bird depends on both its own direct fitness accumulation increment and also that of the other bird. That is, each bird has an ‘other-regarding’ motivation in addition to its self-regarding motivation. What might this dependence be? One possibility would be some type of exponentially weighted product of the fitness increments because this would capture some sense of pleasure in cooperative synergy. In ‘perfect teamwork’, the birds enjoy each other's welfare as much as their own and, furthermore, the birds also take their actions jointly. In perfect teamwork, both birds are hypothesized to possess the same pleasure function.
A motivation must be supplied for continuing to work as a team rather than reverting to individualism. Therefore, each's fitness increment is decreased by whatever fitness would be lost if the other were to threaten it, the so-called ‘threat point’. The loss of fitness from incurring a threat is painful. Each individual maximizes its own pleasure with steps taken jointly with the other even in the face of the pain sustained from receiving threats from that other individual. The joint change in their behaviours from one hour to the next is proportional to the slope of their common pleasure function. Thus, in seeking pleasure, each bird works with the other to jointly climb the gradient in their shared pleasure function. If both birds jointly alter their behaviour in this way, then their joint time allocations to foraging and guarding will tend to converge to a behavioural equilibrium over several time steps that is the NBS from cooperative game theory [26]. The NBS may be quite different from the NCE.
Several illustrations of contrasting behavioural outcomes resulting from team play versus individual play have been published: (i) a game patterned after the peacock wrasse [42] in which females can choose to lay eggs in a male's nest or to broadcast them, and the male may choose to guard a nest anticipating that females will lay eggs there or to follow after the females who are broadcasting their eggs on the ocean floor [29]; (ii) a game with three players patterned after the Eurasian oystercatcher [43] in which some reproductive groups involve two females and one male. In these trios, the females may choose to cooperate or to compete and cooperating females share extensive sexual intimacy [29]; (iii) a game wherein a female member of a pair can choose to share access to the nestlings or can control the nestlings while denying access to the male, whereas the male can choose to help at the nest or to abandon the nest [3, table 11]; and (iv) a game in which a male and a female each have to allocate between feeding and guarding a nest [30]. Conditions for the evolution of individualistic versus team play are analysed in Akçay et al. [27] and Roughgarden [30].
(b). Firms
A firm is a collection of animals working together because one member is in a position to control incentives that align the self-interests of others to cooperate. For example, parents and offspring form a hierarchy in which parents control the food. Parents can dispense incentives that lead the offspring to cooperate with one another and with the parents themselves [44,45]. When the optimal incentives have been put in place, cooperation occurs because of a coincidence of individual interests. Each member's objectives are purely self-regarding. Because a family may be viewed as a ‘firm’ whose product is offspring it is appropriate to turn to management science and the economic theory of the firm to see how a family might be organized to maximize its offspring production. Groves [46] considers the problem of optimal incentives in a kind of firm called a conglomerate. The profit for the conglomerate accrues from the earnings of its divisions, and similarly, the profit in fitness earned by a family accrues from the fitness of the offspring it produces.
The family-firm approach confirms predictions, based on an evolutionary-strategy analysis using the ESS solution concept, that offspring should signal honestly to their parent [47–49]. However, the family-firm approach offers a different derivation and shows that if the parent implements ‘optimal incentive policies’, the parent and chicks work honestly together as a team to maximize the fitness produced by the nest. Unlike an evolutionary-strategy analysis, the family-firm approach in the behavioural tier can be extended to predict the time of weaning or fledging—this amounts to the time for ‘spinning off’ divisions from the parent corporation.
Like Grafen, Godfray and Johnstone [47–49], the family-firm approach agrees with Alexander's [50] perspective that emphasizes parental control of the parent–offspring interaction. In contrast to Alexander however, who visualized continuing parent offspring conflict dominated by the parent, according to the family-firm theory the parent resolves conflict by how it allocates resources to the offspring. This theory fundamentally disagrees with the Trivers/Parker/Mcnair perspective [51–55], which features unresolved and continuing parent–offspring conflict as well as dishonesty in parent–offspring signalling, whereby offspring continually psychologically manipulate their parents, especially at the time of weaning or fledging.
Our ‘biological conglomerate’ consists of a parent as ‘manager’, and one, or potentially two or more chicks, as the ‘divisions’ within the conglomerate, literally, a ‘parent corporation with offspring divisions’. The following analysis, taken from Roughgarden & Song [44], concentrates on a family with one chick as illustrated in figure 3.
Figure 3.
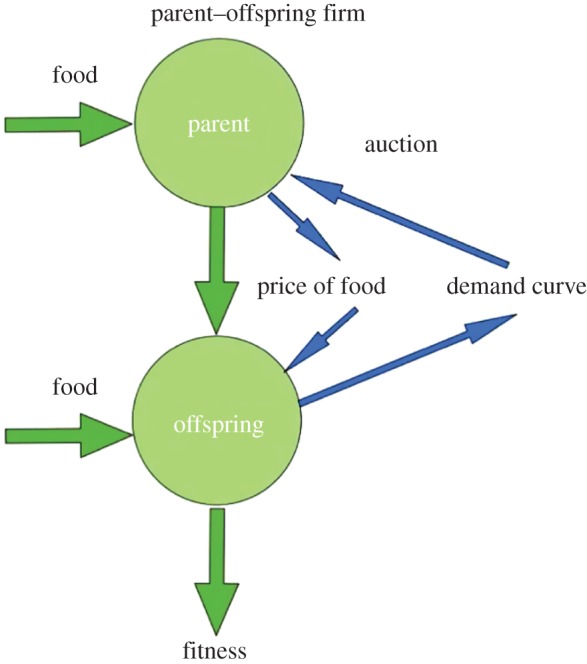
Diagram of family firm.
The chick is assumed to possess private knowledge about its state that must somehow be communicated to the parent—it must tell the parent how much food it wants as a function of how much effort it must spend in begging or other activities to obtain that food. This information is contained in its ‘demand function’ for food. For a given size and physiological state, if the chick can obtain a unit of food with but a mere peep, then it would accept lots of food, whereas if the chick can obtain a unit of food only from a long bout of loud begging, then it will only bother to request less food.
The demand function will change as the chick grows and also surely depends on various random environmental fluctuations such as temperature. Low temperature days, for example, would require the chick to have more food on that day to sustain a higher metabolic rate to maintain its body temperature than a hot temperature day would. In any case, the parent is assumed not to know the chick's demand function for food at any given time, and needs to have the chick honestly communicate its state so that it may act accordingly at that time.
During the growing season, the parent feeds the chick each day, and the chick grows. The fitness a chick earns is assumed to depend on the size it attains when fledged. The fitness of a parent equals the fitness of its chick, discounted by the hazard it has encountered while foraging to provide for the chick. The task is for the parent to structure its payout of food to the chick such that its own fitness is maximized, as is the fitness of its chick as well.
Groves [46] discusses incentive mechanisms whereby the conglomerate's manager communicates with the divisions and vice versa. Among the incentive structures Groves describes is a scheme that amounts to setting up an internal market for the goods supplied by the manager to the divisions. He shows that if the manager requires the divisions to participate in a kind of auction for the resources they need, then the resulting distribution of resources from the manager to the divisions is optimal in the sense that the production from the corporation is maximized, as is the production from each of the divisions, and the information being conveyed by the divisions to the manager is accurate. This ‘internal-market’ incentive mechanism motivates thinking of food-begging by chicks as an auction whereby the parent distributes food to the chicks optimally.
The parent is interested in maximizing its own fitness production rate (i.e. fitness accumulation per time step). It must determine how much food to give the chick to maximize its own fitness production rate. To make this determination, the parent can set up a mechanism whereby it charges for the food it dispenses to the chicks.
The parent can view each unit of time spent begging as purchasing a certain amount of food from it. The parent can set the amount of food it will deliver per unit of begging. This is the per-unit begging ‘price’ it will charge. For any given price, there is a certain amount of food the chick will want to buy, its ‘optimal demand’ for the given price, that the chick itself determines to maximize its own fitness at that price. For a given price, if the chick buys less than the optimal it does not grow much, whereas if it buys more than optimal it has wasted too much time and energy in begging. So, the chick has an optimal demand for each price. If the parent knows the chick's optimal demand for each price, it can set the price so that the chick will want to purchase the exactly the amount of food it, the parent, wants to give.
This market can be thought of as a form of auction, albeit with just one bidder because there is only one chick. A ‘Walrasian’ auction (Léon Walras, cf. [56]) is where each agent calculates its demand for the good at every possible price and submits this to an auctioneer. The price is then set so that the total demand across all agents equals the total amount of the good. Thus, a Walrasian auction perfectly matches the supply with the demand. In this case, because the parent can set the price, the chick will want to buy exactly what the parent has to give it. This perfect match of demand with supply leads to ‘market clearing’, whereby no food is left on the table, so to speak. Using this auction to dispense resources from the manager to the divisions is shown by Groves [46] to be an optimal incentive system in the sense that both parties, the parent and chick find the outcome to be optimal. The Walrasian auction is not the only type of auction consistent with an optimal incentive system. Akçay [45] explores another auction setup called the Vickrey–Clark–Groves mechanism [57] in which each successful bidder winds up bidding enough to compensate the collective for its impact on the collective's welfare, as though each chick bids enough to compensate the remaining chicks for the food it consumes.
The analysis for this model appears in Roughgarden & Song [44]. Specifically, figure 4 illustrates the optimal demand curve for three sizes of chick. This is computed by determining the demand that maximizes the chick's growth for each price. Each curve is a decreasing function of price, indicating that as the price increases, the optimal demand decreases. Furthermore, as the chick ages, its demand curve shifts on the graph to the left, indicating that for any given price, the optimal demand for a small chick is higher than for a large chick, i.e. a small chick optimally ‘wants’ more food for a given price than a large chick.
Figure 4.
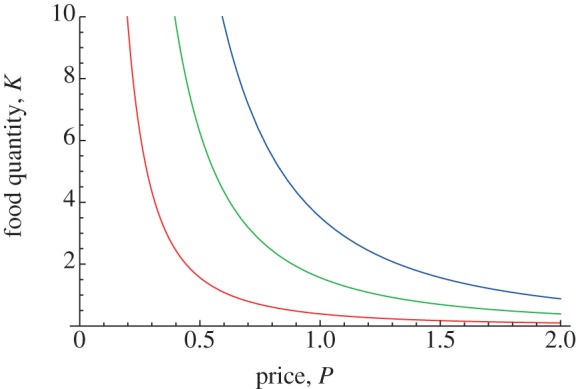
Optimal demand curves for chick. Each curve shows that the amount of food a chick begs for, K, decreases as the price charged by the parent, P, increases. The curves, from right to left, pertain to increasingly bigger chicks. The curve farthest from the origin, for a small chick, shows a higher demand at all prices than the curve, nearest the origin, for a large chick. Adapted from Roughgarden & Song [44].
The chick then must somehow communicate this curve to the parent. Groves [46] describes an iterative process (called tâtonnement) in which the auctioneer displays a trial price and the bidders indicate what they will buy at that price. Then, the auctioneer adjusts the price up or down until a price is found at which all the material for sale would be allocated. There is continuing research in economics and operations research about how a monopolist, i.e. the parent, can learn the demand function of its customers, i.e. the chick [58–60], which might be adapted to give insight into how a parent can interpret a chick's begging as revealing its food demand function.
Next, consider the parent's perspective, and determine the best price for it to charge. The fitness the parent accumulates during the time interval equals the fitness earned through investing in the chick's growth discounted by a function of the probability of surviving through the time interval during which the food has been gathered. And for a given degree of inherent danger in the environment (few versus many predators, exposure to abiotic hazards, etc.), the more food the parent must collect, the lower the probability of survival.
At this point, the parent must choose the price of food to maximize its fitness production. Once the chick has communicated its optimal demand curve to the parent, the parent takes this information into account and forms its own fitness production function by substituting the chick's demand curve into its own fitness calculations, as illustrated in figure 5. The curves have a mode. If the parent sets the price too low, then the chick demands a lot of food, and the parent is suffering from the hazard of gathering a large quantity of food. If the parent sets the price too high, then the chick does not demand much food, grows slowly, and parent does not earn much fitness accumulation. So between these extremes, an optimal price balances between the parent's foraging hazard and the chick's growth rate.
Figure 5.
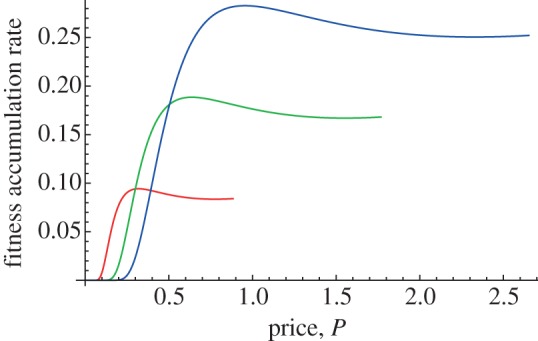
Parental fitness production function with low chick self-feeding. Fitness accumulated per time interval by the parent as a result of the chick's growth during the period, as a function of how much the parent is charging the chick for food, and taking into account its own hazard while foraging and a limited amount of self-feeding by the chick. The self-feeding here yields little food and the optimal price, located at the local maximum, is low enough to induce the chick to solicit parental food. The top curve is for a small chick, the middle curve for a medium sized chick, and the bottom curve for a large chick. Adapted fom Roughgarden & Song [44].
Figure 5 shows the parental fitness accumulation function for three sizes of chick, assuming the chick cannot forage much for itself. Maximizing this function with respect to price, given the chick's size, yields the optimal price the parent should charge to a chick of that size. On this basis of price, the chick bids to buy its optimal demand and the parent should forage for this amount of food to satisfy the bid. At this price, the best interests of both parties, the parent and chick, are simultaneously satisfied.
Now, we add to the model the ability of a chick to collect some food by itself. This ability improves as the chick ages and grows in size, until a time is reached at which it is advantageous for both parent and chick to discontinue the parental feeding. This time is when ‘weaning’ or ‘fledging’ occurs, and is analogous to the ‘spin-off’ of a division by a parent company.
Here is how the time of weaning can be determined according to the family-firm approach. The chick's fitness production function is now modified to include self-feeding. Because the demand for parental food is decremented by what the chick can gather for itself, there is a price for food at which the demand for parental food is zero.
Figures 5–7 illustrate the parental fitness production functions for three levels of self-feeding by the chick. The sequence of figures shows the parental fitness production function for an increasing degree of self-feeding. The curves terminate at the maximum price the parent can charge and still have the chick solicit food.
Figure 7.
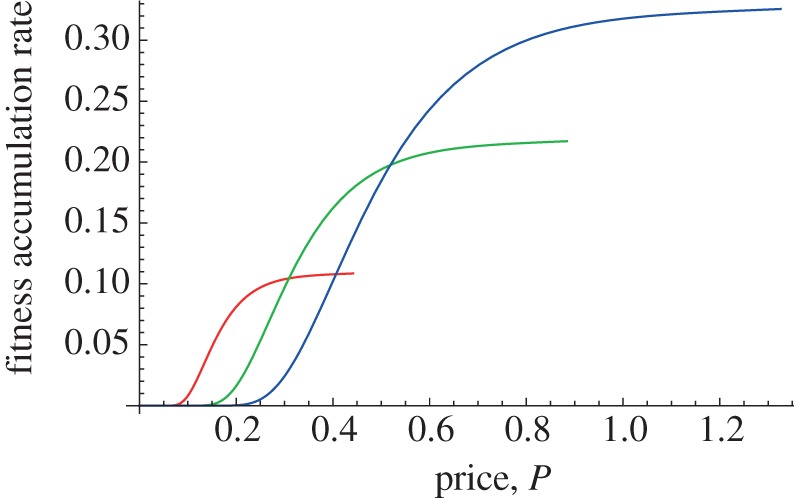
Parental fitness production function with high chick self-feeding. The self-feeding here yields ample food so that the optimal price, which is located where each curve terminates at the right end of the curves, is so high that the chick will not solicit parental food. Adapted from Roughgarden & Song [44].
For a low degree of self-feeding (figure 5), the curves reveal a local maximum that indicates an optimal price that involves some degree of parental support for the chick. In contrast, when the self-feeding yields a lot of food, the parent's highest fitness is attained by pricing the food so high that no food is solicited by the chick—in figure 7, the curves are monotonically increasing with the price and the peak fitness is towards the right where the curves terminate.
Figure 6 illustrates the transition degree of self-feeding that marks the break between parental feeding and no parental feeding. For the parameters in the illustration, the transition from parental feeding to no feeding happens to occur when the chick obtains 0.857864 units of food per time interval from its own foraging efforts. Self-feeding that yields more food than this threshold implies that the parent does not find it advantageous to continue feeding the chick, and self-feeding that yields less than this threshold implies that the parent finds it optimal to continue feeding the chick.
Figure 6.
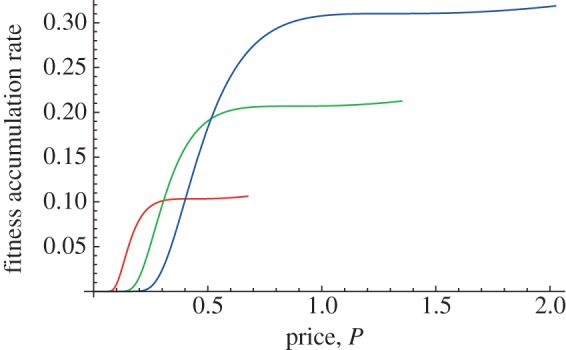
Parental fitness production function with moderate chick self-feeding. The self-feeding here yields exactly the amount that marks the transition from parental feeding to no parental feeding. This threshold degree self-feeding yield is the point of weaning. Adapted from Roughgarden & Song [44].
The criterion for weaning is defined to be the self-feeding level at which the parental fitness production function changes from having a local maximum to becoming monotonic. In practice, as the chick grows its foraging capacity grows too, and when the chick reaches an age/size at which it passes the weaning level of self-feeding, the parent should price the food it delivers so high that the chick no longer solicits parental food. At this point, the parent ‘spins off’ the chick to forage on its own. The parent continues to accrue fitness gains as the chick grows under its own power and without further parental investment.
The optimal quantity of food delivered by the parent to the chick declines, but remains positive, as the amount of self-feeding increases up until the weaning threshold is passed. Once the weaning threshold is passed however, the optimal quantity drops abruptly to zero. Thus, weaning is not a gradual loss of the chick's dependence on the parent, but a sudden onset of full independence. Yet, this situation is not one of conflict because it is optimal to both parties given the parental incentive structure.
A biological firm is not necessarily harmonious as the above illustration might suggest. In some ecological circumstances, as illustrated earlier, the offspring might know their value to the parent and be able communicate it to the parent. (The parent might value the size of the offspring and the offspring might know its own size and thus be able to communicate it.) If so, a parent's best interest would be served by setting incentives to ensure honest communication and to minimize the inefficiency of lying and conflict. In other circumstances, offspring might not know their value to the parent and thus not be able to communicate it to their parent even if they were attempting an honest report. (The parent might value the prospective resource capturing ability of an offspring, which would be unknown to the offspring itself until it learned how to forage effectively.) In this case, a parent's best interest would be served by allowing conflict between itself and the offspring and competition between the sibs with one another to reveal their capabilities. The inefficiency of conflict would be the cost to the parent of obtaining information about the offsprings' value to it that would not otherwise be knowable. This contrast between whether the information about their value to the parent is communicable or not is perhaps a better way to view the distinction between ‘offspring control’ and ‘parent control’ [52–55] because in either case, the parent does control the resources made available to the offspring. However, if the offspring know what the parent values about them then the parent's interest is served by setting incentives for honest communication, whereas if the offspring are ignorant of their value to the parent, then the parent's interest is served by witnessing the outcome of offspring squabbling.
4. Conclusion
Milam [61] has reviewed disagreements about Darwin's [62] sexual selection that began with Wallace [63] and continued through the mid-1900s. The 1970s witnessed renewed interest in sexual selection [51,64,65]. Yet sexual selection remains open to debate [1–3,29,66–69]. Sexual selection can be considered a specific hypothesis to explain the evolution of ornaments and armaments, as well as a general approach to understanding the evolution of mating and reproductive behaviour overall. Social selection as presented here offers an alternative to sexual selection both as an explanation for the evolution of ornaments and as a general approach to mating behaviour and parental investment. Social selection emphasizes the role of cooperation in reproductive activities, although competition is acknowledged too.
Pleasure-based teamwork and incentive-structured firms offer mechanisms by which cooperation can evolve as a direct benefit. That is, cooperation realized through teamwork or working in a firm is not altruism, and its evolution is consistent with, but does not require, kin/multilevel selection or other evolutionary processes that cause the evolution of traits that benefit the receiver but disadvantage the donor. Literature on how cooperation evolves has been slanted towards kin, group and multi-level selection because of the prominence of social insects as paradigmatic examples of altruism in the seminal work of Hamilton [70], although attention to mutual direct benefits is increasing [71]. Nonetheless, mutual direct benefits, including pleasure-based teamwork and working in incentive-structured firms, may be more important overall in explaining cooperative behaviour than altruism-based explanations.
Acknowledgments
I thank Richard Prum and four anonymous reviewers for their help and consideration of this manuscript, and Dustin Rubenstein and Michael Levandowsky for organizing the Columbia University conference in April 2011 at which the issues this paper raises were discussed.
References
- 1.Roughgarden J. 2004. Evolution's rainbow: diversity, gender and sexuality in nature and people. Berkeley CA: University of California Press [Google Scholar]
- 2.Roughgarden J. 2007. Challenging Darwin's theory of sexual selection. Daedalus 136, 1–14 10.1162/daed.2007.136.2.23 (doi:10.1162/daed.2007.136.2.23) [DOI] [Google Scholar]
- 3.Roughgarden J. 2009. The genial gene: deconstructing Darwinian selfishness. Berkeley, CA: University of California Press [Google Scholar]
- 4.West-Eberhard M. J. 1983. Sexual selection, social competition and speciation. Q. Rev. Biol. 58, 155–183 10.1086/413215 (doi:10.1086/413215) [DOI] [Google Scholar]
- 5.Moore A., Haynes K., Preziosi R., Moore P. 2002. The evolution of interacting phenotypes: genetics and evolution of social dominance. Am. Nat. 160, S186–S197 10.1086/342899 (doi:10.1086/342899) [DOI] [PubMed] [Google Scholar]
- 6.Frank S. 2006. Social selection. In Evolutionary genetics: concepts and case studies (eds Fox C. W., Wolf J. B.), pp. 350–363 Oxford, UK: Oxford University Press [Google Scholar]
- 7.Tanaka Y. 1996. Social selection and the evolution of animal signals. Evolution 50, 512–523 10.2307/2410827 (doi:10.2307/2410827) [DOI] [PubMed] [Google Scholar]
- 8.Mangel M., Clark C. 1989. Dynamic modeling in behavioral ecology. Princeton, NJ: Princeton University Press [Google Scholar]
- 9.Iwasa Y. 2000. Dynamic optimization of plant growth. Evol. Ecol. Res. 2, 437–455 [Google Scholar]
- 10.Akçay E., Roughgarden J. 2009. The perfect family: decision making in biparental care. PLoS ONE 4, 1–10 10.1371/journal.pone.0007345 (doi:10.1371/journal.pone.0007345) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Meseth E. 1975. The dance of the Laysan Albatross, Diomedea immutabilis. Behaviour 54, 217–257 10.1163/156853975X00263 (doi:10.1163/156853975X00263) [DOI] [Google Scholar]
- 12.Stamps J., Alderón-De Anda C. M., Perez C., Drummond H. 2002. Collaborative tactics for nestsite selection by pairs of blue footed boobies. Behaviour 139, 1383–1412 10.1163/15685390260514672 (doi:10.1163/15685390260514672) [DOI] [Google Scholar]
- 13.Takahashi M., Arita H., Hiraiwa-Hasegawa M., Hasegawa T. 2008. Peahens do not prefer peacocks with more elaborate trains. Anim. Behav. 75, 209–1219 10.1016/j.anbehav.2007.10.004 (doi:10.1016/j.anbehav.2007.10.004) [DOI] [Google Scholar]
- 14.Frank L. 1997. Evolution of genital masculinization: why do female hyaenas have such a large ‘penis’? Trends Ecol. Evol. 12, 58–62 10.1016/S0169-5347(96)10063-X (doi:10.1016/S0169-5347(96)10063-X) [DOI] [PubMed] [Google Scholar]
- 15.Fisher R. A. 1915. The evolution of sexual preference. Eugenics Rev. 7, 184–191 [PMC free article] [PubMed] [Google Scholar]
- 16.Fisher R. A. 1930. The genetical theory of natural selection. Oxford, UK: Clarendon Press [Google Scholar]
- 17.Iyer P. 2009. Evolution of sexual dimorphism from gametes to ornaments. Thesis, Department of Biological Sciences, Stanford University. (UMI Microform 3382941, ProQuest LLC, Ann Arbor, MI.) [Google Scholar]
- 18.Gross M. 1985. Disruptive selection for alternative life histories in salmon. Nature 313, 47–48 10.1038/313047a0 (doi:10.1038/313047a0) [DOI] [Google Scholar]
- 19.Goodson J., Bass A. 2000. Forebrain peptides modulate sexually polymorphic vocal circuity. Nature 403, 769. 10.1038/35001581 (doi:10.1038/35001581) [DOI] [PubMed] [Google Scholar]
- 20.Prum R. O. 2010. The Lande–Kirkpatrick mechanism is the null model of evolution by intersexual selection: implications for meaning, honesty, and design in intersexual signals. Evolution 64, 3085–3100 10.1111/j.1558-5646.2010.01054.x (doi:10.1111/j.1558-5646.2010.01054.x) [DOI] [PubMed] [Google Scholar]
- 21.Wagner R. 1992. The pursuit of extra-pair copulations by monogamous razorbills: how do females benefit? Behav. Ecol. Sociobiol. 29, 455–464 10.1007/BF00170176 (doi:10.1007/BF00170176) [DOI] [Google Scholar]
- 22.Wagner R. 1996. Male–male mountings by a sexually monomorphic bird: mistaken identity or fighting tactic? J. Avian Biol. 27, 209–214 10.2307/3677224 (doi:10.2307/3677224) [DOI] [Google Scholar]
- 23.Akçay E., Roughgarden J. 2007. Extra-pair parentage: a new theory based on transactions in a cooperative game. Evol. Ecol. Res. 9, 1223–1243 [Google Scholar]
- 24.Shuker D. 2010. Sexual selection: endless forms or tangled bank? Anim. Behav. 79, E11–E17 10.1016/j.anbehav.2009.06.006 (doi:10.1016/j.anbehav.2009.06.006) [DOI] [Google Scholar]
- 25.Maynard S. J. 1982. Evolution and the theory of games. Cambridge, UK: Cambridge University Press [Google Scholar]
- 26.Nash J. 1950. The bargaining problem. Econometrica 18, 155–162 10.2307/1907266 (doi:10.2307/1907266) [DOI] [Google Scholar]
- 27.Akçay E., Van Cleve J., Feldman M. W., Roughgarden J. 2009. A theory for the evolution of other-regard integrating proximate and ultimate perspectives. Proc. Natl Acad. Sci. USA 106, 19 061–19 066 10.1073/pnas.0904357106 (doi:10.1073/pnas.0904357106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Akçay E., Roughgarden J. 2011. The evolution of payoff matrices: providing incentives to cooperate. Proc. R. Soc. B 278, 2198–2206 10.1098/rspb.2010.2105 (doi:10.1098/rspb.2010.2105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Roughgarden J., Oishi M., Akçay E. 2006. Reproductive social behavior: cooperative games to replace sexual selection. Science 311, 965–969 10.1126/science.1110105 (doi:10.1126/science.1110105) [DOI] [PubMed] [Google Scholar]
- 30.Roughgarden J. In press Teamwork, pleasure and bargaining in animal social behaviour. J. Evol. Biol. 25, 1454–1462 10.1111/j.1420-9101.2012.02505.x (doi:10.1111/j.1420-9101.2012.02505.x) [DOI] [PubMed] [Google Scholar]
- 31.Darwin C. 1872. The expression of the emotions in man and animals. London, UK: John Murray [Google Scholar]
- 32.Berridge K., Kringelbach M. 2008. Affective neuroscience of pleasure: reward in humans and animals. Psychopharmacology 199, 457–480 10.1007/s00213-008-1099-6 (doi:10.1007/s00213-008-1099-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kringlebach M., Berridge K. 2010. The functional neuroanatomy of pleasure and happiness. Discov. Med. 9, 579–587 [PMC free article] [PubMed] [Google Scholar]
- 34.Kringlebach M., Berridge K. 2010. The neuroscience of happiness and pleasure. Soc. Res. (NY) 77, 659–678 [PMC free article] [PubMed] [Google Scholar]
- 35.Kringlebach M., Berridge K. 2010. (eds) Pleasures of the brain. Oxford, UK: Oxford University Press [Google Scholar]
- 36.Goodson J., Schrock S., Klatt J., Kabelik D., Kingsbury M. 2009. Mesotocin and nonapeptide receptors promote Estrildid flocking behavior. Science 325, 862–866 10.1126/science.1174929 (doi:10.1126/science.1174929) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.De Dreu C., Greer L., Van Kleef G., Shalvi S., Handgraaf M. 2011. Oxytocin promotes human ethnocentrism. Proc. Natl Acad. Sci. USA 108, 1262–1266 10.1073/pnas.1015316108 (doi:10.1073/pnas.1015316108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cabanac M. 1992. Pleasure: the common currency. J. Theor. Biol. 155, 173–200 10.1016/S0022-5193(05)80594-6 (doi:10.1016/S0022-5193(05)80594-6) [DOI] [PubMed] [Google Scholar]
- 39.Cabanac M., Guillaume J., Balasko M., Fleury A. 2002. Pleasure in decision-making situations. BMC Psychiatry 2, 7. 10.1186/1471-244X-2-7 (doi:10.1186/1471-244X-2-7) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cabanac M., Bonniot-Cabanac M. 2007. Decision making: rational or hedonic? Behav. Brain Funct. 3, 45. 10.1186/1744-9081-3-45 (doi:10.1186/1744-9081-3-45) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nash J. 1951. Non-cooperative games. Ann. Math. 54, 286–295 10.2307/1969529 (doi:10.2307/1969529) [DOI] [Google Scholar]
- 42.Warner R., Lejeune F., van den Berghe E. 1994. Dynamics of female choice for parental care in a fish species where care is facultative. Behav. Ecol. 6, 73–81 10.1093/beheco/6.1.73 (doi:10.1093/beheco/6.1.73) [DOI] [Google Scholar]
- 43.Heg D., van Treuren R. 1998. Female–female cooperation in polygynous oystercatchers. Nature 391, 687–691 10.1038/35612 (doi:10.1038/35612) [DOI] [Google Scholar]
- 44.Roughgarden J., Song Z. In press. Incentives in the family I: the family firm, an evolutionary/economic theory for parent-offspring relations. In Ancestral landscapes in human evolution: culture, childrearing and social wellbeing (eds Narvaez D., Valentino K., Gray P., Fuentes A., McKenna J.), New York: Oxford University Press [Google Scholar]
- 45.Akçay E. 2012. Incentives in the family II: behavioral dynamics and the evolution of non-costly signaling. J. Theor. Biol. 294, 9–18 10.1016/j.jtbi.2011.10.031 (doi:10.1016/j.jtbi.2011.10.031) [DOI] [PubMed] [Google Scholar]
- 46.Groves T. 1973. Incentives in teams. Econometrica 41, 617–631 10.2307/1914085 (doi:10.2307/1914085) [DOI] [Google Scholar]
- 47.Grafen A. 1990. Biological signals as handicaps. J. Theor. Biol. 144, 517–546 10.1016/S0022-5193(05)80088-8 (doi:10.1016/S0022-5193(05)80088-8) [DOI] [PubMed] [Google Scholar]
- 48.Godfray H. C. J. 1991. Signaling of need by offspring to their parents. Nature 352, 328–330 10.1038/352328a0 (doi:10.1038/352328a0) [DOI] [Google Scholar]
- 49.Godfray H., Johnstone R. 2000. Begging and bleating: the evolution of parent-offspring signalling. Phil. Trans. R. Soc. Lond. B. 355, 1581–1591 10.1098/rstb.2000.0719 (doi:10.1098/rstb.2000.0719) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Alexander R. D. 1974. The evolution of social behaviour. Annu. Rev. Ecol. Syst. 5, 325–383 10.1146/annurev.es.05.110174.001545 (doi:10.1146/annurev.es.05.110174.001545) [DOI] [Google Scholar]
- 51.Trivers R. L. 1972. Parental investment and sexual selection. In Sexual selection and the descent of man (ed. Campbell B.), pp. 136–179 Chicago, IL: Aldine Publishing [Google Scholar]
- 52.Parker G., Macnair M. 1978. Models of parent-offspring conflict. I. Monogamy. Anim. Behav. 26, 97–110 10.1016/0003-3472(78)90009-X (doi:10.1016/0003-3472(78)90009-X) [DOI] [PubMed] [Google Scholar]
- 53.Parker G., Macnair M. 1979. Models of parent-offspring conflict. IV. Suppression: evolutionary retaliation by the parent. Anim. Behav. 27, 1210–1235 10.1016/0003-3472(79)90068-X (doi:10.1016/0003-3472(79)90068-X) [DOI] [Google Scholar]
- 54.Macnair M. R., Parker G. A. 1978. Models of parent-offspring conflict. II. Promiscuity. Anim. Behav. 26, 111–122 10.1016/0003-3472(78)90010-6 (doi:10.1016/0003-3472(78)90010-6) [DOI] [PubMed] [Google Scholar]
- 55.Macnair M. R., Parker G. A. 1979. Models of parent-offspring conflict. III. Intra-brood conflict. Anim. Behav. 27, 1202–1209 10.1016/0003-3472(79)90067-8 (doi:10.1016/0003-3472(79)90067-8) [DOI] [Google Scholar]
- 56.Jaffé W., Walker D. (eds) 1983. Essays on Walras. Cambridge, UK: Cambridge University Press [Google Scholar]
- 57.Nisan N., Roughgarden T., Tardos E., Vazirani V. (eds) 2007. Algorithmic game theory. Cambridge, UK: Cambridge University Press [Google Scholar]
- 58.Balvers R., Cosimano T. 1990. Actively learning about demand and the dynamics of price adjustment. Econ. J. 100, 882–898 10.2307/2233664 (doi:10.2307/2233664) [DOI] [Google Scholar]
- 59.Aghion P., Bolton P., Harris C., Jullien B. 1991. Optimal learning by experimentation. Rev. Econ. Stud. 58, 621–654 10.2307/2297825 (doi:10.2307/2297825) [DOI] [Google Scholar]
- 60.Lin K. 2006. Dynamic pricing with real-time demand learning. Eur. J. Oper. Res. 174, 522–538 10.1016/j.ejor.2005.01.041 (doi:10.1016/j.ejor.2005.01.041) [DOI] [Google Scholar]
- 61.Milam E. 2010. Looking for a few good males: female choice in evolutionary biology. Baltimore, MD: Johns Hopkins University Press [Google Scholar]
- 62.Darwin C. 1871. The descent of man and selection in relation to sex. London, UK: John Murray [Google Scholar]
- 63.Wallace A. R. 1871. Darwin's ‘The descent of man and selection in relation to sex’. The Academy 15, 177 [Google Scholar]
- 64.Parker G., Baker R., Smith V. 1972. The origin and evolution of gamete dimorphism and the male–female phenomenon. J. Theor. Biol. 36, 529–553 10.1016/0022-5193(72)90007-0 (doi:10.1016/0022-5193(72)90007-0) [DOI] [PubMed] [Google Scholar]
- 65.Dawkins R. 1976. The selfish gene. Oxford, UK: Oxford University Press [Google Scholar]
- 66.Roughgarden J. 2005. The myth of sexual selection. California Wild 53, 18–23 [Google Scholar]
- 67.Clutton-Brock T. 2009. Sexual selection in females. Anim. Behav. 77, 3–11 10.1016/j.anbehav.2008.08.026 (doi:10.1016/j.anbehav.2008.08.026) [DOI] [Google Scholar]
- 68.Roughgarden J., Akçay E. 2010. Do we need a sexual selection 2.0? Anim Behav. 79, E1–E4 10.1016/j.anbehav.2009.06.006 (doi:10.1016/j.anbehav.2009.06.006) [DOI] [Google Scholar]
- 69.Roswell K. 2011. Intrasexual competition in females: evidence for sexual selection? Behav. Ecol. 22, 1131–1140 10.1093/beheco/arr106 (doi:10.1093/beheco/arr106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hamilton W. D. 1964. The genetical evolution of social behaviour, I, II. J. Theor. Biol. 7, 1–52 10.1016/0022-5193(64)90038-4 (doi:10.1016/0022-5193(64)90038-4) [DOI] [PubMed] [Google Scholar]
- 71.Leimar O., Hammerstein P. 2010. Cooperation for direct fitness benefits. Phil. Trans. R. Soc. B 365, 2619–2626 10.1098/rstb.2010.0116 (doi:10.1098/rstb.2010.0116) [DOI] [PMC free article] [PubMed] [Google Scholar]