

Abstract
Few mammals—cetaceans, domestic cats and select bats and rodents—can send and receive vocal signals contained within the ultrasonic domain, or pure ultrasound (greater than 20 kHz). Here, we use the auditory brainstem response (ABR) method to demonstrate that a species of nocturnal primate, the Philippine tarsier (Tarsius syrichta), has a high-frequency limit of auditory sensitivity of ca 91 kHz. We also recorded a vocalization with a dominant frequency of 70 kHz. Such values are among the highest recorded for any terrestrial mammal, and a relatively extreme example of ultrasonic communication. For Philippine tarsiers, ultrasonic vocalizations might represent a private channel of communication that subverts detection by predators, prey and competitors, enhances energetic efficiency, or improves detection against low-frequency background noise.
Keywords: ultrasound, sensory ecology, bioacoustics, auditory brainstem response, evoked potential audiogram
1. Introduction
Human hearing is relatively poor at higher frequencies; our putative high-frequency limit is 20 kHz, and frequencies above this boundary are classified as ultrasound. The hearing of most haplorhine primates is similarly constrained, although some species have high-frequency limits approaching 45 kHz [1]. For owl monkeys (Aotus), the functional significance of such enhanced hearing is uncertain; all recorded vocalizations are contained below 10 kHz [2]. Other primates—Callithrix, Cebuella, Cheirogaleus, Galago, Microcebus, Nycticebus, Prolemur—can emit and respond to calls with ultrasonic components [3–8]; however, the dominant frequencies are always well within the human audible range. Thus, the prospects for primate communication solely within the ultrasound, or pure ultrasound [9], appear limited. In fact, few mammals are reported to send and receive pure ultrasonic signals. Such mammals include cetaceans, domestic cats, and a few select bats and rodents [10–15].
Among primates, the potential for pure ultrasonic communication is perhaps greatest in the family Tarsiidae. Tarsiers are small (113–142 g) nocturnal faunivores that form simple social groups, typically consisting of male–female pairs and their offspring. These factors are seldom associated with complex vocal signals [16], yet tarsiers have relatively large vocal repertoires. For example, Tarsius spectrum can emit at least 15 distinct call types, all contained below 16 kHz, that appear to serve several functions, such as conveying alarm, deterring rivals and facilitating social interactions [17]. Other species such as T. bancanus (Bornean tarsier) and T. syrichta (Philippine tarsier) vocalize less often—they are ‘ordinarily silent’ [18]—yet their calls are described as ‘piercing’ and ‘bat-like’ [18]. As a result, Niemitz [18] suggested that tarsiers might communicate in the ultrasound. Later recordings of T. bancanus were consistent with this hypothesis; harmonic bands in the ultrasound were detected in five call types, but the dominant frequencies were audible to humans [19].
Such results are promising, but technical and practical limitations have restricted the study of tarsier sensory faculties, especially hearing. Tarsiers are rare, endangered and challenging to maintain in captivity. Thus, traditional behavioural audiograms that require months of husbandry and training are impractical and difficult to justify. Here, we capitalize on recent technical advances to safely generate audiograms from wild animals under field conditions. Our findings not only verify that tarsiers are sensitive to the ultrasound, but also that T. syrichta can send and receive vocal signals in the pure ultrasound.
2. Material and methods
(a). Audiograms
Six adult or subadult tarsiers (T. syrichta; figure 1a) were captured by hand or mist net in the vicinity of Motorpool, Surigao del Norte, Mindanao, Philippines (09°38′ N; 125°33′ E). The animals were anaesthetized for ca 1 h (5–8 mg kg−1 Telazol, supplemented with 3 mg kg−1 Telazol or 15 μg kg−1 dexmedetomidine) and positioned in a custom-built sound-attenuating chamber (electronic supplementary material, figure S1).
Figure 1.

Philippine tarsier and its auditory and acoustic capabilities. (a) Tarsius syrichta in its natural habitat, Mindanao, Philippines. (b) Representative ABR waveform series for 45 kHz stimuli. (c) Average audiogram and standard error of six individuals; the high-frequency limit was extrapolated from the thresholds at 45 and 64 kHz. (d) Spectrogram of vocalization shows the call duration (approx. 650 ms) and dominant frequency (approx. 70 kHz); signal intensity is represented by the density of the red–black scale (hear electronic supplementary material, sound file S1, available online).
To estimate auditory sensitivities, we used the minimally invasive auditory brainstem response (ABR) method [20] and EVREST software [21,22] operating on a PC with a data acquisition card (NI-USB-6251; National Instruments, Austin, USA). The stimuli were tone pips (sinusoidal, 2-cycle linear rise/fall, 1-cycle plateau) digitally generated within EVREST, converted to analogue (500 kHz, 16-bit), bandpass filtered from 0.02 to 200 kHz (3B series, 24 dB/octave rolloff, Butterworth; Krohn-Hite, Brockton, USA), attenuated (PA5; Tucker-Davis, Alachua, USA), and delivered at a rate of 39.1 s–1 (alternating polarity) for 2048 repetitions per frequency/level step via an electrostatic speaker (ES1; Tucker-Davis) positioned 10 cm from the left ear. Test frequencies were half octaves from 1 to 64 kHz, delivered in steps of 10 dB from ca 60 to 80 dB and decreasing until the response approached residual electroencephalographic background noise (BN) level and was undetectable; we then tested 5 dB steps around the estimated threshold. We calibrated the peak-equivalent levels (dB peSPL re 20 μPA) of the tone pip stimuli by recording 50 ms pure tones via a free-field 0.5 inch condenser microphone (MKH 800, Sennheiser, Old Lyme, USA; frequency response 0.03–50 kHz, 0°) connected to a PC running Raven Pro v. 1.3 (Cornell Laboratory of Ornithology, Ithaca, USA); calibration was periodically cross-checked with an ultrasound condenser microphone (USM 10-2, Laar, Gut Klein Goernow, Germany; frequency response 0.1–140 kHz, 0°) and a Korg MR-1 mobile recorder (Melville, USA).
The ABR was recorded with 28-guage subdermal needle electrodes (F-E3; Grass Instruments, West Warwick, USA) positioned in the skin over the cranial vertex (positive), the ipsilateral mastoid (reference) and the contralateral mastoid (ground). The signals were recorded with a biopotential amplifier (P511; Grass Instruments), amplified (×105), filtered (0.03–3 kHz bandpass, 60 Hz notch), digitized (10 kHz, 16-bit), input into EVREST (20 ms epochs, 12 μV reject level) and digitally filtered offline [20]. We used a linear regression method [20] to determine the threshold (quietest detectable level) for each frequency. Generally, responses above average BN were included in the regression. We set the criterion at 62 ηV (average BN + 40 ηV); at this level, the ABR was consistently distinct above random fluctuations in BN (figure 1b and electronic supplementary material, figure S2). For the average audiogram, we calculated two parameters that agree across methods [1,20]: the frequency of best sensitivity and the highest frequency detectable at 60 dB SPL. Given that the threshold at 64 kHz was below 60 dB, we estimated the high-frequency limit by linear extrapolation of thresholds at 45 and 64 kHz.
(b). Vocal recordings
We recorded the vocalizations of 35 wild tarsiers from the islands of Bohol and Leyte with an ultrasound microphone/recording unit (Song Meter SM2BAT; Wildlife Acoustics, Concord, USA; frequency response 0–96 kHz). During fieldwork, we occasionally observed stereotypical vocal behaviours that were inaudible to us; and, for eight individuals, we recorded a call in the pure ultrasound (Sound File S1, available online). The call was emitted in two contexts: (i) during human handling and (ii) during unrestrained activity within temporary enclosures. In Raven Pro, we defined the dominant frequency of the call as that with the most energy, and determined the lowest frequency by visual distinctiveness from spectrogram noise. This protocol was approved by the Institutional Animal Care and Use Committee of Texas A&M University (approval no. AUP2008-7).
3. Results
Among six tarsiers, we found two frequency regions of best auditory sensitivity (1.4 and 16 kHz), and an audible range that extended substantially into the ultrasound (figure 1c). The 60 dB high-frequency limit of T. syrichta is estimated at 91 kHz (figure 1c), a value that surpasses the known range of all other primates (figure 2) and is matched by few animals [10–14]. We also recorded a distinctive vocalization in the pure ultrasound (bandwidth 67–79 kHz, dominant frequency, approx. 70 kHz) from eight animals (figure 1d; electronic supplementary material, sound file S1, available online). The minimum frequency of the call (67 kHz) is the highest value for any terrestrial mammal excluding bats and some rodents [10,14]. The distinctive tone-like structure of the call partly resembles the vocalizations of other tarsier species [19]; however, none of these calls was purely ultrasonic. They were contained below 34 kHz and featured strong harmonics less than 20 kHz.
Figure 2.
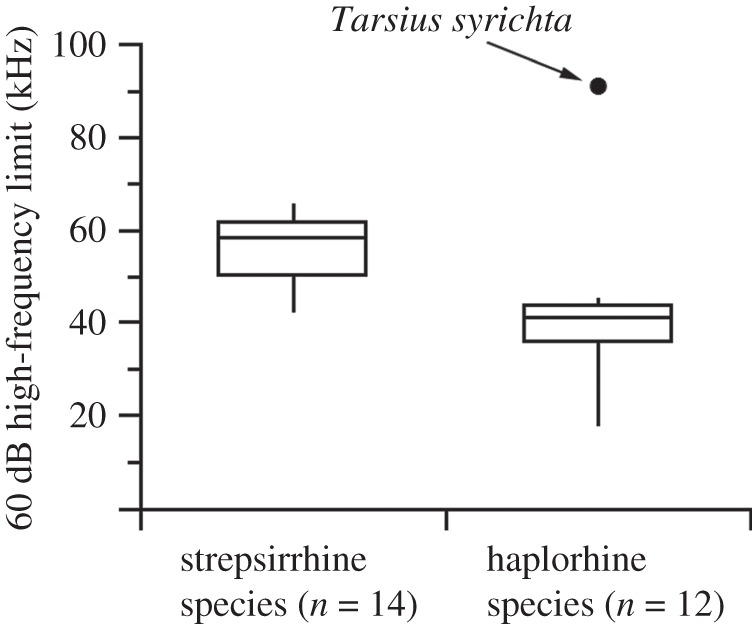
The 60 dB high-frequency limits of strepsirrhine and haplorhine primates. Boxes represent the interquartile range between the first and third quartiles and the line inside represents the median. Whiskers denote the lowest and highest values, excepting Tarsius syrichta. At ca 91 kHz, T. syrichta is substantially higher than other primate species [1,23,24].
4. Discussion
The Philippine tarsier's estimated high-frequency limit of auditory sensitivity (ca 91 kHz) and call with a dominant frequency of 70 kHz are among the highest values recorded for terrestrial mammals, and an extreme example of acoustic communication. Vocalizations in the pure ultrasound might confer several selective advantages. For example, although ultrasonic calls are expected to attenuate quickly, they are also expedient private channels of communication with the potential to subvert detection by predators, prey and competitors, enhance energetic efficiency, and improve signal-to-noise ratios in habitats inundated with low-frequency biological noise [15]. Our observation that the tarsiers emitted the call in the proximity of humans suggests a context of alarm. Ultrasonic alarm calls can be advantageous to both the signaller and receiver as they are potentially difficult for predators to detect and localize [13,15].
Yet, tarsiers are themselves specialized predators. They lack conventional visual adaptations to nocturnality, such as a reflective tapetum lucidum in the retina, and possess instead the largest eyes relative to body size of any mammal [25]. Visual predation of arthropod prey is a central element of tarsier foraging behaviour, but it is constrained by the availability of downwelling light in the forest understory. Under the darkest nocturnal conditions—when cloud cover occludes starlight—it is plausible that the exceptional hearing of tarsiers contributes to improved foraging efficiency through acoustic eavesdropping [18]. For instance, tarsiers might attend to ultrasonic signals between prey species (e.g. katydids and moths [26,27]) or broadband cues signifying the presence and location of potential prey, such as rustling leaves [28].
Our findings demonstrate that Philippine tarsiers can send and receive signals in the pure ultrasound. Although the advantages of this specialized sensory adaptation are yet uncertain, additional studies comparing the evolutionary, behavioural and physiological foundations of ultrasonic communication in tarsiers are likely to provide insights into the fundamental properties promoting high-frequency hearing in all vertebrates.
Acknowledgements
All animals were captured, examined and released unharmed under protocols approved by the Institutional Animal Care and Use Committee of Dartmouth College (approval no. 10-11-02).
Research permission was received from the barangay of Motorpool, the Office of the Punong Barangay, the Office of the Municipal Mayor, Congressman G.A. Romarate, the National Commission on Indigenous Peoples and the Protected Areas and Wildlife Bureau of the Department of Environment and Natural Resources (permit R13-2010-003). We thank the Mamanwa, E. M. Bade, E. S. Buiser, R. F. Burkard, C. Cuevas, L. D. Dagsaan, M. R. Duya, M. L. E. Ceso, L. M. Gales, L. R. Heaney, J.-J. Hublin, A. U. Luczon, B. E. B. Magsaganay, S. F. Matugas, T. M. S. Lim, M. G. T. Pascua, M. Pedregosa-Hospodarsky, C. Piazzas, J. D. Quiñonez, C. Salibay, F. Spoor, the Max Planck Society and De La Salle University-Dasmarinas. Funding was received from the David and Lucile Packard Foundation (NJD) and the Fulbright Foundation, Primate Conservation Inc., Conservation International and Texas A&M University (S.L.G.).
References
- 1.Coleman M. N. 2009. What to primates really hear? A meta-analysis of all known primate behavioral audiograms. Int. J. Primatol. 30, 55–91 10.1007/s10764-008-9330-1 (doi:10.1007/s10764-008-9330-1) [DOI] [Google Scholar]
- 2.Kantha S. S., Koda H., Suzuki J. 2009. Owl monkey vocalizations at the Primate Research Institute, Inuyama. Neotrop. Primates 16, 43–46 10.1896/044.016.0110 (doi:10.1896/044.016.0110) [DOI] [Google Scholar]
- 3.Pola Y. V., Snowdon C. T. 1975. The vocalizations of pygmy marmosets (Cebuella pygmaea). Anim. Behav. 23, 825–842 10.1016/0003-3472(75)90108-6 (doi:10.1016/0003-3472(75)90108-6) [DOI] [PubMed] [Google Scholar]
- 4.Zimmerman E. 1981. First record of ultrasound in two prosimian species. Naturwissenschaften 68, 531–532 10.1007/BF00365388 (doi:10.1007/BF00365388) [DOI] [Google Scholar]
- 5.Cherry J. A., Izard M. K., Simons E. 1987. Description of ultrasonic vocalizations of the mouse lemur (Microcebus murinus) and the fat-tailed dwarf lemur (Cheirogaleus medius). Am. J. Primatol. 13, 181–185 10.1002/ajp.1350130208 (doi:10.1002/ajp.1350130208) [DOI] [PubMed] [Google Scholar]
- 6.Braune P., Schmidt S., Zimmermann E. 2005. Spacing and group coordination in a nocturnal primate, the golden brown mouse lemur (Microcebus ravelobensis): the role of olfactory and acoustic signals. Behav. Ecol. Sociobiol. 58, 587–596 10.1007/s00265-005-0944-4 (doi:10.1007/s00265-005-0944-4) [DOI] [Google Scholar]
- 7.Bergey C., Patel E. R. 2008. A preliminary vocal repertoire of the greater bamboo lemur (Prolemur simus). Nexus 1, 69–84 [Google Scholar]
- 8.Bezerra B. M., Souto S. 2008. Structure and usage of the vocal repertoire of Callithrix jacchus. Int. J. Primatol. 29, 671–701 10.1007/s10764-008-9250-0 (doi:10.1007/s10764-008-9250-0) [DOI] [Google Scholar]
- 9.Arch V. S., Grafe T. U., Gridi-Papp M., Narins P. M. 2009. Pure ultrasonic communication in an endemic Bornean frog. PLoS One 4 e5413. 10.1371/journal.pone.0005413 (doi:10.1371/journal.pone.0005413) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sales G., Pye D. 1974. Ultrasonic communication by animals. London, UK: Chapman and Hall [Google Scholar]
- 11.Ehret G. 1980. Development of sounds communication in animals. In Advances in the study of animal behavior, vol. 11 (eds Rosenblatt J. S., Hinde R. A., Beer C., Busnel M. C.), pp. 179–225 New York, NY: Academic Press [Google Scholar]
- 12.Bohn K. M., Boughman J. W., Wilkinson G. S., Moss C. F. 2004. Auditory sensitivity and frequency selectivity in greater spear-nosed bats suggest specializations for acoustic communication. J. Comp. Physiol. 190A, 185–192 10.1007/s00359-003-0485-0 (doi:10.1007/s00359-003-0485-0) [DOI] [PubMed] [Google Scholar]
- 13.Wilson D. R., Hare J. F. 2004. Ground squirrel uses ultrasonic alarms. Nature 430, 523. 10.1038/430523a (doi:10.1038/430523a) [DOI] [PubMed] [Google Scholar]
- 14.Liu R. C., Miller K. D., Merzenich M. M., Schreiner C. E. 2003. Acoustic variability and distinguishability among mouse ultrasound vocalizations. J. Acoust. Soc. Am. 114, 3412–3422 [DOI] [PubMed] [Google Scholar]
- 15.Arch V. S., Narins P. M. 2008. ‘Silent’ signals: Selective forces acting on ultrasonic communication systems in terrestrial vertebrates. Anim. Behav. 76, 1423–1428 10.1016/j.anbehav.2008.05.012 (doi:10.1016/j.anbehav.2008.05.012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McComb K., Semple S. 2005. Coevolution of vocal communication and sociality in primates. Biol. Lett. 1, 381–385 10.1098/rsbl.2005.0366 (doi:10.1098/rsbl.2005.0366) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nietsch A. 2003. Outline of the vocal behavior of Tarsius spectrum: Call features, associated behaviors, and biological functions. In Tarsiers: past, present, and future (eds Wright P. C., Simons E. L., Gursky S.), pp. 196–220 New Brunswick, NJ: Rutgers University Press [Google Scholar]
- 18.Niemitz C. 1979. Outline of the behavior of Tarsius bancanus. In The study of prosimian behaviour (eds Doyle G. A., Martin R. D.), pp. 631–660 New York, NY: Academic Press [Google Scholar]
- 19.Niemitz C. 1984. Vocal communication of two tarsier species (Tarsius bancanus and Tarsius spectrum). In Biology of tarsiers (ed. Niemitz C.), pp. 129–141 Stuttgart, Germany: Gustav Fischer Verlag [Google Scholar]
- 20.Ramsier M. A., Dominy N. J. 2010. Comparison of auditory brainstem responses and behavioral estimates of hearing sensitivity in Lemur catta and Nycticebus coucang. Am. J. Primatol. 72, 217–233 10.1002/ajp.20780 (doi:10.1002/ajp.20780) [DOI] [PubMed] [Google Scholar]
- 21.Finneran J. J. 2008. Evoked response study tool (EVREST) user's guide. San Diego, CA: SSC; (Tech. Doc. SSC San Diego 3226) [Google Scholar]
- 22.Finneran J. J. 2009. Evoked response study tool: a portable, rugged system for single and multiple auditory evoked potential measurements. J. Acoust. Soc. Am. 126, 491–500 10.1121/1.3148214 (doi:10.1121/1.3148214) [DOI] [PubMed] [Google Scholar]
- 23.Osmanski M. S., Wang X. 2011. Measurement of absolute auditory thresholds in the common marmoset (Callithrix jacchus). Hearing Res. 227, 127–133 10.1016/j.heares.2011.02.001 (doi:10.1016/j.heares.2011.02.001) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ramsier M. A., Cunningham A. J., Finneran J. J., Dominy N. J. In press Social drive and the evolution of primate hearing. Phil. Trans. R. Soc. B [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ross C. F. 2000. Into the light: The origin of Anthropoidea. Annu. Rev. Anthropol. 29, 147–194 [Google Scholar]
- 26.Montealegre Z. F., Morris G. K., Mason A. C. 2006. Generation of extreme ultrasonics in rainforest katydids. J. Exp. Biol. 209, 4923–4937 10.1242/jeb.02608 (doi:10.1242/jeb.02608) [DOI] [PubMed] [Google Scholar]
- 27.Nakano R., Takanashi T., Fujii T., Skals N., Surlykke A., Ishikawa Y. 2009. Moths are not silent, but whisper ultrasonic courtship songs. J. Exp. Biol. 212, 4072–4078 10.1242/jeb.032466 (doi:10.1242/jeb.032466) [DOI] [PubMed] [Google Scholar]
- 28.Siemers B. M., Goerlitz H. R., Robsomanitrandrasana E., Piep M., Ramanamanjato J.-B., Rakotondravony D., Ramilijaona O., Ganzhorn J. U. 2007. Sensory basis of food detection in wild Microcebus murinus. Int. J. Primatol. 28, 291–304 10.1007/s10764-007-9135-7 (doi:10.1007/s10764-007-9135-7) [DOI] [Google Scholar]