

Abstract
Here we used an array-based differential screen to uncover the expression of the neuropeptide neuromedin B (NMB) in the trigeminal ganglia of mice. Double-labeling experiments reveal NMB is expressed in a subset of sensory neurons that colabel with calcitonin gene-related peptide and TRPV1 suggestive of a role for NMB in nociception. Indeed, administration of NMB antagonist greatly attenuates edema and nerve sensitization following stimulation of peripheral nerves with mustard oil, demonstrating that NMB contributes to neurogenic inflammation. Moreover, direct injection of NMB causes local swelling and nociceptive sensitization. Interestingly, we also find that the receptor for NMB is expressed in interneurons in the superficial layers of the dorsal horn. We used NMB-saporin to specifically eliminate NMBR-expressing neurons and determined they are required in responses to noxious heat, but not for reaction to mechanical and pruritic stimuli. Thus, NMB may be a neurotransmitter that is selectively involved in the perception of thermal stimuli.
Introduction
Neuropeptides and their receptors are important in somatosensory signaling pathways. Many different peptides have been reported to be present in the trigeminal (TG) and dorsal root ganglia (DRG), including substance P (SP), calcitonin gene-related peptide (CGRP), galanin, neuropeptide Y (NPY), and gastrin releasing peptide (GRP) (Zhang et al., 1995; Bergman et al., 1996; Sun and Chen, 2007; Xu et al., 2010). SP and CGRP are the best characterized of these neuropeptides and are involved in peripheral and central nociceptive processes. They enhance excitatory stimulation in the dorsal horn of the spinal cord and are also released in the periphery, resulting in the local heating, wheal, and flare, a reaction known as neurogenic inflammation (Brain and Cox, 2006).
Neuromedin B (NMB) and GRP are the only two members of the mammalian bombesin family of peptides. These two peptides activate structurally similar but pharmacologically distinct G-protein-coupled receptors: the NMB receptor (NMBR) and the GRP receptor (GRPR) (Corjay et al., 1991; Jensen et al., 2008). In the periphery NMB and GRP have a wide variety of actions including smooth muscle contraction and exocrine and endocrine functions (Heasley, 2001). In the CNS these peptides regulate food intake and body temperature, as well as stress behavioral responses (Battey and Wada, 1991; Ohki-Hamazaki et al., 1999; Ladenheim et al., 2002). Additionally, immunolocalization studies showed that NMB protein is present in the dorsal horn of the spinal cord (Massari et al., 1983; Namba et al., 1985) and expression was also seen in sensory neurons (Wada et al., 1990). Intrathecal injection of NMB elicits nociceptive reflex responses, but the nature of the reaction produced is unclear (O'Donohue et al., 1984; De Koninck and Henry, 1989; Cridland and Henry, 1992). In contrast, GRP has been clearly demonstrated to be involved in itch sensation (Sun and Chen, 2007; Sun et al., 2009). GRPR knock-out mice, and animals in which GRPR-expressing neurons in the dorsal horn are eliminated, exhibit attenuated responses to most pruritic agents. Moreover, loss of GRPR or ablation of the GRPR-expressing neurons has no effect on other sensory modalities (Sun and Chen, 2007; Sun et al., 2009). This has led to the proposal that GRP is an itch-specific signaling neuropeptide and GRPR neurons in the dorsal horn represent a labeled line for itch.
A direct approach to examine the central role of nociceptive neuropeptides is the targeted ablation of peptide receptors expressing neurons with saporin-conjugated peptides (Wiley and Lappi, 2003). The elimination of neurons expressing different neuropeptide receptors in the dorsal horn reveals that these cells are functionally heterogeneous; loss of spinal neurons expressing TacR1, NPY, μ-opioid, and the galanin receptor generates mice that exhibit a variety of different deficits in processing nociceptive stimuli (Nichols et al., 1999; Kline and Wiley, 2008; Wiley et al., 2009; Lemons and Wiley, 2011). The differences in behavior of mice lacking different cell types highlights the distinct roles of diverse classes of dorsal horn neurons (excitatory and inhibitory interneurons and projection neurons) (Nichols et al., 1999; Kline and Wiley, 2008; Wiley et al., 2009; Lemons and Wiley, 2011).
To begin to characterize the role of NMB in sensory neurons, we have examined its effects on neurogenic inflammation. In addition, using a cell-ablation approach we have established a central function for NMB in the spinal cord.
Materials and Methods
Animals.
Mice were 25–30 g male C57BL/6 and Tg(Nmbr-EGFP) JG22Gsat (GENSAT Project at Rockefeller University). Procedures followed the National Institutes of Health (NIH) guidelines for the use and care of laboratory animals and were approved by the National Institute of Dental and Craniofacial Research Animal Care and Use Committee.
To eliminate specific subsets of dorsal horn interneurons 10 μg of NMB-saporin or 0.13 μg of stable SP-saporin (SSP-saporin) or 1.3 μg of bombesin-saporin or 10 μg of blank-saporin (20 μl of PBS; Advanced Targeting System) were administered to the lumbar 4–5 segments (Mishra and Hoon, 2010). Behavioral testing occurred between 1 and 3 weeks after toxin treatment. Tissue was harvested for expression analysis 7 d after injection of toxin.
Behavioral assays.
For thermal responses, a radiant heat source was focused on the hindpaw and the time from the initiation until withdrawal was measured using an automated Hargreaves test apparatus (Ugo Basile). Animals were habituated for at least 30 min in individual chambers before experiments and a maximum cutoff of 20 s was used to prevent tissue damage. Using an automated von Frey apparatus (Ugo Basile), mechanical responses were assayed by determining the threshold force required to induce paw withdrawal. For itch assays, hair was removed from the lumbar back region and 100 nm of histamine trifluoromethyl toluidide (HTMT) was administered in a 10 μl subcutaneous injection. Following administration of itch-inducing agent, numbers of itching bouts were counted over 30 min.
To test for effects of neuropeptides 10 μl of NMB (50 μg) or 10 μl of NMB (50 μg) CGRP (10 μg) and SP (1 μg) were injected into the hindpaw. Thirty minutes after injection of peptides, paw edema was measured using a plethysmometer (IITC) and thermal and mechanical sensitization measured by Hargreaves and von Frey apparatus assays (Ugo Basile), respectively. We examined if pretreatment with peptide antagonists could prevent neurogenic inflammation. Ten minutes before inducing neurogenic inflammation, neuropeptide antagonists or vehicle control were administered via tail vein injection; 10 μg of NMB antagonist BIM 23042 (Tocris Bioscience) or a mixture of 10 μg BIM 23042, 10 μg CGRP 8–37, and 1 μg spantide were given as a single bolus (100 μl in PBS vehicle). BIM 23042 is a specific antagonist of the NMBR (Orbuch et al., 1993), CGRP 8–37 is an antagonist of the CGRP receptor (Deems et al., 1991; Wang et al., 1991), and spantide is an inhibitor of TacR1 (Folkers et al., 1984). Hindpaw reflex responses for thermal (Hargreaves) and mechanical stimuli (von Frey) and changes in paw volume were assessed before treatment and 30 min after induction of inflammation with mustard oil (10 μl; 1% in mineral oil). Increased paw volume to injection of neuropeptide or mustard oil was normalized to the untreated or mustard oil-treated condition, respectively.
To investigate the effects of inflammation on mechanical sensitivity, the plantar surface of the hindpaw was injected with 20 μl of complete Freund's adjuvant (CFA; Sigma). Behavioral testing was performed 24 h after injection.
In situ hybridization and immunohistochemistry.
In situ hybridization (ISH) was performed at high stringency (washed 30 min, 0.2× SSC, 70°C) as described previously (Hoon et al., 1999). Probes were generated for genes of interest by PCR and Topo-cloned into pCRII (Invitrogen). Sequences of cloned genes were confirmed and in vitro transcription was used to generate antisense cRNA probes. ISH was used to detect select genes in the TG and DRG and TacR1 and GRPR-positive interneurons in the spinal cord. Immunohistochemistry with chicken anti-green fluorescent protein (GFP) (1:500; Abcam) was used to detect NMBR-positive neurons in Nmbr-EGFP mice. Development of immunopositive GFP signal was achieved with donkey anti-chicken HRP-conjugated secondary antibody (Jackson Immunolabs) together with NovaRed. Analysis of ablation was performed on tissue collected from ≥3 mice. For each mouse, numbers of positive neurons for each receptor were counted from ≥10 16 μm serial sections. Total numbers of neurons expressing receptor were calculated for each animal, averaged, and normalized to values from blank-saporin controls.
Double-label ISH/immunohistochemistry was developed with anti-digoxigenin antibodies coupled to alkaline phosphatase together with Fast Red. CGRP and TRPV1 were detected with anti-CGRP (1:200; Millipore Bioscience Research Reagents) and anti-TRPV1 antibodies (1:100; Millipore Bioscience Research Reagents) and developed with donkey anti-rabbit FITC (Jackson Immunolabs) (Adler et al., 2000). The ISH signal for Nmb is nuclear because the doubling-labeling technique preferentially detects unspliced nuclear transcripts. Quantization of NMB overlap with CGRP and TRPV1 was performed by counting positive neurons from more than four sections for each double staining. Images were collected using a Microphot FX microscope (Nikon) and a Zeiss LSM 510 confocal microscope and processed using Adobe Photoshop.
Gene array analysis.
Total RNA was isolated from TG, DRG, or geniculate ganglion (GG) using an RNeasy minikit (Qiagen). cDNA was synthesized from total RNA and used as a template to produce a cy5-labeled cRNA probe following protocols recommended by the manufacturer (Agilent). Gene arrays (GE4 × 44K V2; Agilent) were hybridized, washed, and scanned as recommended by the manufacturer. Data were analyzed using Gene Spring (version 11) software.
Heterologous expression of NMBR and GRPR.
Full-length NMBR and GRPR were amplified from brain cDNA and cloned into mammalian expression vector pEAK8 (Egde Biosystems). HEK-293 cells were transiently transfected with pEAK8-NMBR-ires-EGFP or pEAK8-GRPR and pEAK8-GFP using the Trans-it-293 kit in 24-well culture plates as recommended by the manufacturer (Mirus). Twenty-four hours after transfection, 20 μg/ml of NMB-saporin or blank-saporin or 2.5 μg/ml of bombesin-saporin were added to cultured media and 48 h later fields of live cells were photographed and GFP-positive cells counted. Cells were also stained with the vital stain propidium iodide to measure cell viability. Images were collected from at least four randomly chosen fields of cells from three independent experiments, and data were normalized versus untreated cells.
Results
We set out to identify novel molecules involved in detection and signaling processes of somatosensory neurons and used mouse whole genome microarrays to analyze genes expressed in the TG. To subtract genes expressed in glia and molecules involved in the general housekeeping functions, we compared gene expression profiles of TG with GG. Unlike the TG, which is composed of neurons that detect as well as transmit somatosensory input, the neurons in the GG are only involved in transmitting signals from taste buds. Table 1 lists genes we identified with TG versus GG differential expression > 5.5 (genes whose expression is highly significant with >2 SDs from the mean). As expected, many of the molecules we uncovered have been identified previously and are known to be important in somatosensation, e.g., TrpM8, TrpA1, Scn10a, Scn11a, Mrgpcr, and neurotrophic factor receptors. To identify genes likely to be involved in mediating responses to specific stimuli, we examined the expression patterns of selected molecules by ISH, looking for genes expressed in a subsets of neurons (Fig. 1). Among the molecules prominently enriched and expressed in subsets of neurons were neuropeptides (Fig. 1). One of the neuropeptides we found was NMB. As expected from its differential expression, NMB is expressed in the TG and DRG but is not detectable by ISH in the GG (Fig. 2). The level of expression of NMB estimated from normalized gene array data indicates it is expressed at a level similar to CGRP (Calca and Calcb) and SP (Tac1) (Table 2). Previously, immunological methods were used to show NMB fibers, but no cell bodies were observed in the spinal cord (Massari et al., 1983; Namba et al., 1985). We confirmed using ISH that there are no NMB-expressing cells in the spinal cord (Fig. 2j). In contrast, mRNA transcripts for tachykinin (Tac1) and GRP are expressed in neurons in the dorsal horn as recently reported (Polgár et al., 2006; Koga et al., 2011)(Fig. 2k,l). The studies characterizing immunopositive NMB fibers in the spinal cord (Massari et al., 1983; Namba et al., 1985) are consistent with this staining due to afferent projections of DRG neurons.
Table 1.
Molecules identified by comparison of trigeminal ganglia and genticulate ganglion RNA transcripts
| Change | Gene name | Acc number | Change | Gene name | Acc number |
|---|---|---|---|---|---|
| 65.9 | Npy2r | NM_008731 | 7.2 | Nmur1 | NM_010341 |
| 41.2 | Dgkk | NM_177914 | 7.2 | Kcnmb1 | NM_031169 |
| 30.1 | Oprm1 | AK079569 | 7.2 | Prl | NM_011164 |
| 29.8 | Trpm8 | NM_134252 | 7.2 | Grid1 | AK039103 |
| 28.4 | Klhl5 | AK090246 | 7.1 | Gh | NM_008117 |
| 28.0 | Krt27 | NM_010666 | 7.1 | Pfkfb3 | NM_133232 |
| 28.0 | Scn11a | NM_011887 | 7.1 | Dgkh | AK083897 |
| 27.5 | BC023719 | BC023719 | 7.0 | Dopey2 | AK009785 |
| 27.2 | Calca | NM_001033954 | 7.0 | Rab27b | NM_030554 |
| 27.0 | Tshr | NM_011648 | 7.0 | Cpn1 | NM_030703 |
| 26.6 | Fgg | NM_133862 | 7.0 | Myh7 | NM_080728 |
| 26.0 | Prokr2 | NM_144944 | 6.9 | Rnf112 | NM_009548 |
| 25.8 | 9130204L05Rik | NM_001101461 | 6.9 | Epha10 | NM_177671 |
| 25.1 | Gfra3 | NM_010280 | 6.9 | Shisa9 | NM_028277 |
| 22.2 | Ntrk1 | NM_001033124 | 6.9 | Myt1l | AK012660 |
| 22.1 | Mrgprb5 | NM_207538 | 6.9 | Plau | NM_008873 |
| 21.9 | Tmem45b | NM_144936 | 6.8 | Chrna7 | NM_007390 |
| 20.4 | Giyd2 | NM_029420 | 6.8 | Gpr35 | NM_022320 |
| 19.5 | Calcb | NM_054084 | 6.8 | Lrrn4 | NM_177303 |
| 19.3 | Tshb | NM_009432 | 6.8 | LOC791382 | AK048476 |
| 18.5 | Scn10a | NM_009134 | 6.7 | LOC545785 | AK037393 |
| 17.7 | Prdm8 | NM_029947 | 6.7 | Zdhhc14 | AK033799 |
| 15.9 | LOC100504399 | AK081497 | 6.6 | Mas1 | NM_008552 |
| 15.7 | Dyrk4 | NM_207210 | 6.6 | 5930438M14 | AK031263 |
| 14.9 | Trpc6 | NM_013838 | 6.6 | Celf6 | NM_175235 |
| 14.7 | IgG2a like | XR_033948 | 6.6 | Arg2 | NM_009705 |
| 14.4 | Chrna7 | NM_007390 | 6.6 | E430024P14Rik | AK083703 |
| 13.6 | Npy5r | NM_016708 | 6.5 | Ms4a3 | NM_133246 |
| 13.3 | GlyT2 like | XM_001004722 | 6.5 | Dnajc22 | NM_176835 |
| 13.2 | Gna14 | NM_008137 | 6.5 | Serpine3 | XM_893895 |
| 13.2 | 7530428D23Rik | XM_887943 | 6.5 | Osta | AK147155 |
| 12.9 | Kcnf1 | NM_201531 | 6.4 | Myo15 | NM_010862 |
| 12.1 | Vstm2b | NM_021387 | 6.4 | Opalin | NM_153520 |
| 12.1 | Pkib | NM_001039050 | 6.4 | Myh3 | NM_001099635 |
| 12.1 | 1700030C10Rik | NR_015521 | 6.4 | Wnt3a | NM_009522 |
| 11.8 | Kcng2 | NM_001190373 | 6.4 | Kif17 | NM_010623 |
| 11.3 | 1300015D01Rik | AK005014 | 6.4 | Opn5 | NM_181753 |
| 11.2 | Oxtr | D86599 | 6.4 | Dlk1 | NM_010052 |
| 11.1 | LOC242594 | NM_001162980 | 6.4 | Gys2 | NM_145572 |
| 11.0 | Ankrd58 | NM_173779 | 6.4 | Cdkn3 | NM_028222 |
| 11.0 | Ly75 | AK157921 | 6.3 | Inadl | NM_001005787 |
| 11.0 | Slc4a11 | NM_001081162 | 6.3 | Kcng3 | NM_153512 |
| 10.9 | Kcng4 | NM_025734 | 6.3 | Sst | NM_009215 |
| 10.8 | Grik1 | NM_010348 | 6.3 | Kcna6 | AK031890 |
| 10.8 | Mrgprb3 | NM_207537 | 6.3 | Hoxd8 | NM_008276 |
| 10.7 | Col11a1 | NM_007729 | 6.2 | Aldh1l2 | NM_153543 |
| 10.7 | Acpp | NM_019807 | 6.2 | Agxt2l1 | NM_027907 |
| 10.6 | Bad | AK029400 | 6.2 | Cryba2 | NM_021541 |
| 10.1 | Mas1 | NM_008552 | 6.1 | Mrgpra3 | NM_153067 |
| 9.7 | Scn10a | Y09108 | 6.1 | Arhgap36 | NM_001081123 |
| 9.6 | Ptgir | NM_008967 | 6.1 | Ccdc68 | NM_201362 |
| 9.5 | Nppb | NM_008726 | 6.1 | Bcl11b | NM_001079883 |
| 9.5 | Nmb | NM_026523 | 6.0 | B230311B06Rik | XM_001001884 |
| 9.3 | Pitx2 | NM_001042502 | 6.0 | Tmem114 | NM_029070 |
| 9.1 | Tep1 | AK033525 | 6.0 | Dgkh | NM_001081336 |
| 9.1 | AceCS | AK035497 | 6.0 | Dgkh | AK031069 |
| 9.0 | L3mbtl4 | NM_177278 | 5.9 | Grpr | NM_008177 |
| 8.8 | Tmem233 | AK047066 | 5.9 | Pit-1 | D12885 |
| 8.5 | Lgals7 | NM_008496 | 5.9 | Apc | NM_007462 |
| 8.4 | Pmp2 | NM_001030305 | 5.9 | Mrgpra2b | NM_153101 |
| 8.3 | Ret | NM_009050 | 5.9 | Fam19a1 | NM_182808 |
| 8.3 | Trpa1 | NM_177781 | 5.9 | Slc7a10 | NM_017394 |
| 8.3 | Fam19a1 | NM_182808 | 5.9 | Dmbx1 | NM_130865 |
| 8.2 | Mpped1 | NM_172610 | 5.9 | Gpr83 | NM_010287 |
| 8.1 | Dnase1l3 | NM_007870 | 5.8 | Cgnl1 | NM_026599 |
| 8.1 | Prkcq | NM_008859 | 5.8 | Fam188b2 | AK030561 |
| 8.0 | Capn9 | NM_023709 | 5.8 | Fam19a1 | NM_182808 |
| 7.9 | Gpr35 | NM_022320 | 5.7 | Syt16 | NM_172804 |
| 7.8 | Ropn1 | NM_030744 | 5.7 | Lgi1 | NM_020278 |
| 7.8 | Htr1a | NM_008308 | 5.6 | Rnf112 | NM_009548 |
| 7.8 | Lpar3 | NM_022983 | 5.6 | Myh7 | NM_080728 |
| 7.5 | Gsg1l | NM_001101488 | 5.6 | Tmem26 | NM_177794 |
| 7.5 | Krt79 | NM_146063 | 5.6 | Robo3 | NM_001164767 |
| 7.5 | Kiaa1239 | NM_177006 | 5.5 | Slc34a2 | NM_011402 |
| 7.4 | Mal2 | NM_178920 | 5.5 | Bcl2l14 | NM_025778 |
| 7.4 | Syt5 | NM_016908 | 5.5 | Casc1 | AK034148 |
| 7.4 | Slc5a1 | NM_019810 |
List of genes differentially expressed in trigeminal ganglia versus genticulate ganglion with fold -change >2 SDs from the mean; n = 4. ISH data are presented for genes highlighted in bold (see Fig. 1). Fold differences in expression (Change) and GenBank accession numbers (Acc number) are indicated.
Figure 1.

ISH reveals the expression pattern of genes identified in the differential screen of TG versus GG. Sections of TG were hybridized with probes to select molecules as indicated.
Figure 2.
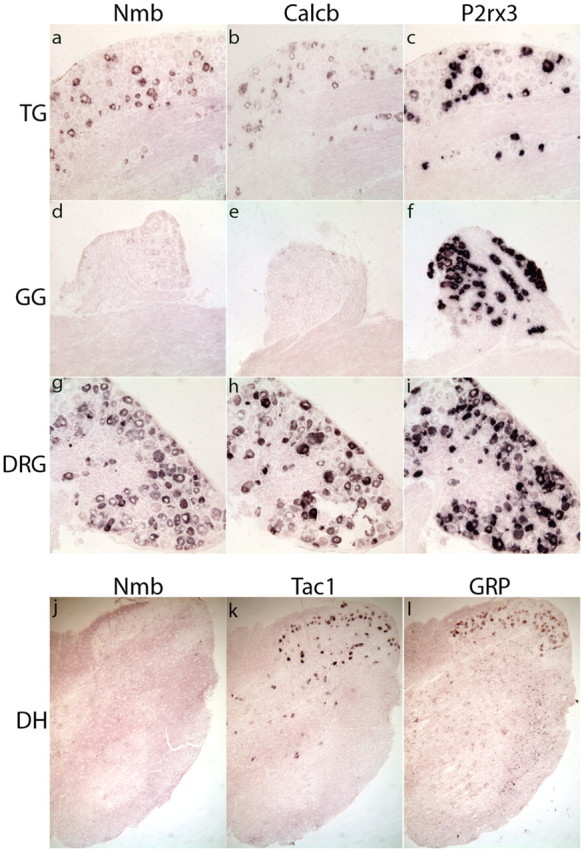
NMB is specifically expressed in a subset of TG and DRG neurons and is undetectable in the GG. ISH reveals that Nmb (a, d, g) and Calcb (b, e, h) are expressed in the TG (a–c) and DRG (g–i), but are undetectable in the GG (d–f). In contrast, the ionotrophic ion-channel P2Rx3 is expressed in all ganglia (c, f, i). ISH of sections through the spinal cord (j–l) establishes that Nmb (j) is not expressed, while Tac1 (k) and GRP (l) are found in a small number of interneurons in the dorsal horn.
Table 2.
Relative expression levels of neuropeptides in dorsal root ganglia
| Mean expression | SD | Gene name | Neuropeptide |
|---|---|---|---|
| 861.6 | 229 | Calca | CGRP |
| 215.0 | 34 | Nmb | NMB |
| 129.9 | 19 | Tac1 | SP |
| 22.9 | 3.8 | Gal | Galanin |
| 18.4 | 3.0 | Calcb | CGRP |
Expression was determined in arbitrary units; data were normalized to median microarray values. Data represent means ± SD; n = 4 arrays. CGRP, calcitonin gene-related peptide; NMB, neuromedin B; SP, substance P.
The function of NMB in somatosensation is still not fully understood, therefore, we focused our studies on this neuropeptide. Initially, we examined the expression pattern of NMB relative to two markers of nociceptive cells: CGRP and TRPV1. Double-label ISH/immunochemistry was used to determine the overlap in expression. Figure 3 shows an overlap of NMB and CGRP expression, ∼25% of NMB-expressing neurons coexpress CGRP (mean 26% ±2.7), similar to the partial superimposition of CGRP and SP (data not shown). In addition, we found NMB expression overlaps with TRPV1-positive cells (24% ± 2.6 NMB-cells coexpress TRPV1; Fig. 3b). Thus, NMB is present in a subset of nociceptive sensory neurons.
Figure 3.
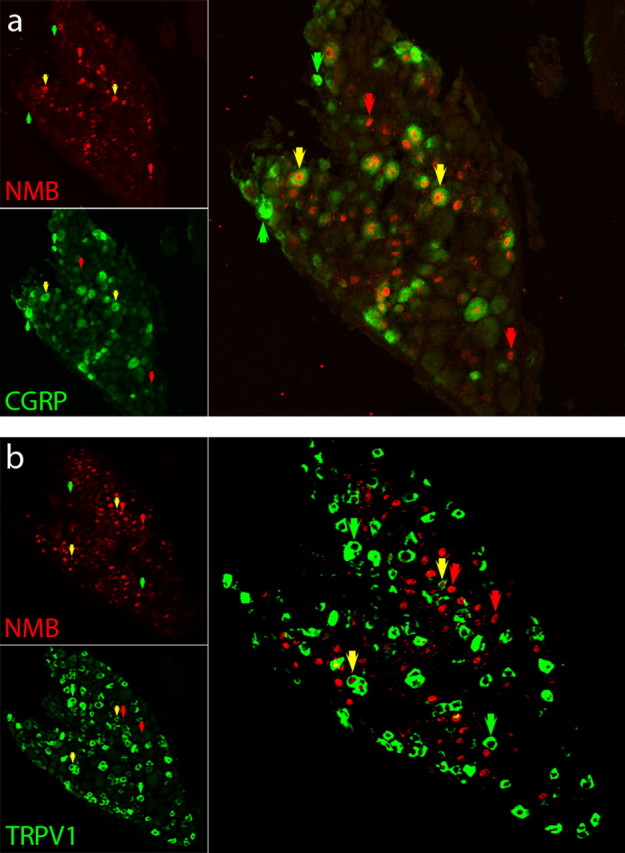
Nmb is coexpressed with markers of nociceptive neurons CGRP and TRPV1. Double-label ISH/immunochemistry of sections through TG establish that Nmb (a, b; red) is coexpressed with CGRP (a; green) and TRPV1 (b; green). Nmb was detected using ISH and cells positive for CGRP and TRPV1 staining were identified using immunohistochemistry. The ISH signal for Nmb is nuclear (see Materials and Methods), while CGRP and TRPV1 antibodies recognize antigen in the cell body. Yellow arrows indicate cells that coexpress both Nmb and either CGRP (a) or TRPV1 (b), red arrows indicate neurons that are positive for Nmb alone, and green arrows highlight cells that contain only CGRP (a) or only TRPV1 (b).
Peripheral role for NMB
Release of CGRP and SP from sensory neurons elicits peripheral vasodilation and extravasation via stimulation of endothelial cells and also induces local hyperalgesia (Brain and Cox, 2006). To examine if NMB can cause swelling and sensitization, we performed injections of NMB into the plantar surface of the hindpaw of mice. Injection of NMB produced a significant enlargement of the foot and swelling was increased further by coadministration with SP and CGRP (Fig. 4a). Withdrawal latencies to thermal and mechanical stimulation were also decreased upon injection of NMB (Fig. 4b,c). In addition, coinjection of NMB, SP, and CGRP elicited greater hyperalgesia than NMB alone (Fig. 4b,c). These results indicate NMB can cause local swelling and induce sensitization of peripheral neurons.
Figure 4.

Injection of NMB induces edema and hyperalgesia and administration of NMB antagonist attenuates mustard oil-induced neurogenic inflammation. To determine the effects of NMB and BIM 23042, paw volume was measured by plethysmometry (a, d), thermal sensitization was assayed using the Hargreaves test (b, e), and mechanical hyperalgesia was quantified using von Frey microfilament measurements (c, f). Hindpaws were injected with NMB or NMB, SP, and CGRP as indicated (a–c). Pretreatment of mice with NMB antagonist BIM 23042, or BIM 23042, CGRP8–37, and spantide dramatically reduces mustard oil-induced swelling and hyperalgesia compared with vehicle-treated animals (d–f). Data represent means ± SEM; n ≥ 6 animals; significant differences between treatments (Student's t test) are indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
To determine whether release of NMB is involved in peripheral sensitization, we examined if neurogenic inflammatory events can be prevented by pretreatment with the specific NMB-antagonist BIM 23042 (Orbuch et al., 1993). We activated peripheral nerves in the foot by injection of mustard oil and measured the effectiveness of antagonist to block swelling and hyperalgesia. Figure 4 shows that BIM 23042 attenuates neurogenic swelling and thermal and mechanical sensitization, demonstrating NMB contributes to neurogenic inflammation upon activation of peripheral nerve fibers. Combined pretreatment with antagonists for NMB, SP, and CGRP produced the greatest reduction of neurogenic inflammation with an ∼70% reduction in swelling and 40% attenuation of sensitization.
Central role for NMB
Many neuropeptides act centrally by activating receptors expressed in the postsynaptic membranes of interneurons in the dorsal horn (Hökfelt, 1991; De Felipe et al., 1998; Salio et al., 2006). Therefore, we examined if the receptor for NMB is expressed in cells in the dorsal horn of the spinal cord. To visualize the NMBR, we made use of Nmbr-GFP transgenic mice (Gong et al., 2003). Revealingly, we found that NMBR is expressed in a small number of neurons in the superficial layers of the dorsal horn (Fig. 5a). The NPY and galanin peptide receptors have also been shown to be expressed in sensory neurons (Zhang et al., 1997, 1998; Landry et al., 2000). Therefore, we also investigated if NMBR is expressed in sensory neurons and found a subset of NMBR-positive cells in DRG (Fig. 5b).
Figure 5.
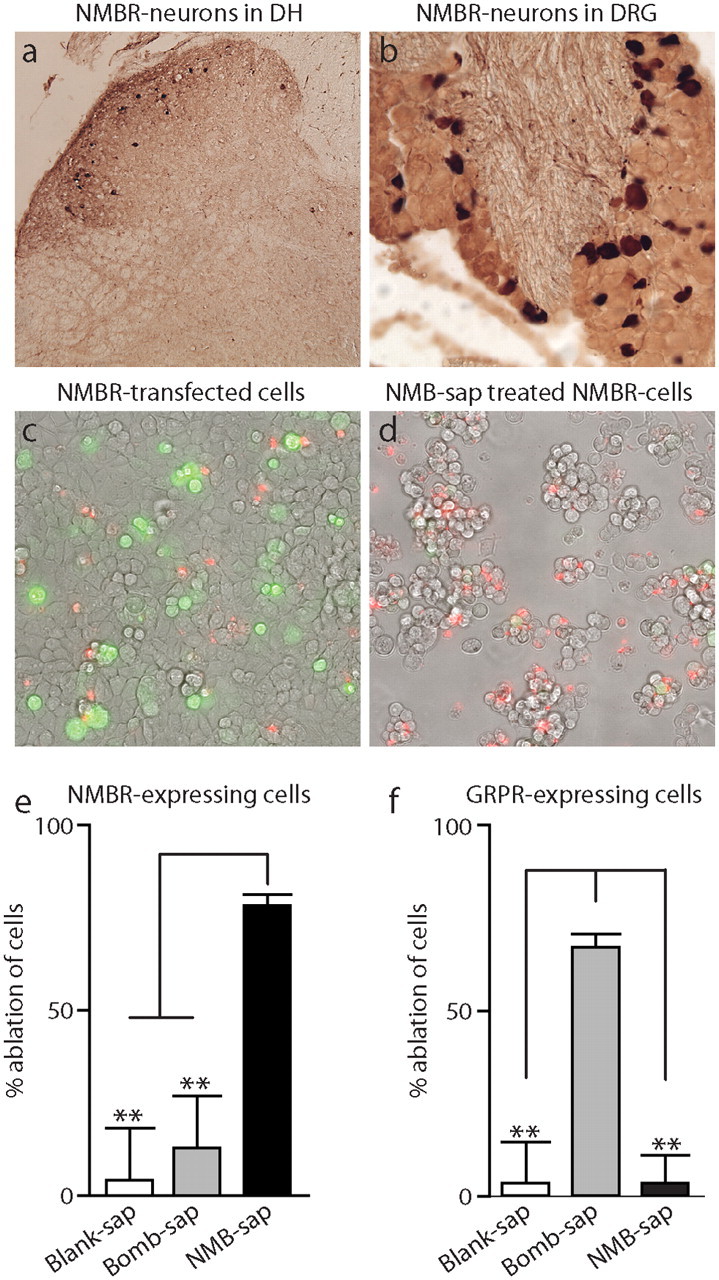
NMBR is expressed in the dorsal horn of the spinal cord and DRG; NMB-saporin selectively ablates NMBR-expressing cells. Immunostaining of sections through dorsal horn (DH) of the spinal cord (a) and DRG (b) from Tg(Nmbr-EGFP) mice detects cells expressing NMBR. Brightfield images were captured for fields of HEK-293 cells expressing NMBR-ires-GFP (c, d). Untreated (c) and NMB-saporin (NMB-sap)-treated cells (d) were counterstained with vital stain propidium iodide (red) to illustrate the increase in nonviable cells following administration of toxin. Quantification of the ablation of cells expressing NMBR (e) or GRPR (f) was measured by the loss of GFP fluorescent cells normalized to untreated controls. NMB-sap results in efficient ablation while bombesin-saporin (Bomb-sap) or blank-saporin (blank-sap) have no effect on NMBR-expressing cells. Cells transfected with GRPR were successfully eliminated with bombesin-saporin, whereas NMB-saporin and blank-saporin treatment did not alter numbers of GRPR-expressing cells. Data represent means ± SEM; n ≥ 3 independent experiments; significant differences between treatments (Student's t test) are indicated by **p < 0.001.
A useful approach to examine the function of neurons that express peptide receptors (Wiley and Lappi, 2003) is to use specific toxins to ablate these cells. Therefore, we synthesized a targeted toxin against NMBR by conjugating carboxy-amidated NMB with the ribosome-inactivating protein saporin. To determine the specificity of this reagent, we tested toxin on cells that heterologously express NMBR or GRPR. Expression of receptors was performed in HEK-293 cells, and the effectiveness of toxin treatment was determined by measuring loss of GFP fluorescent cells (GFP was cotranscribed with receptor). Figure 5 shows that HEK-293 cells expressing NMBR were efficiently ablated following application of NMB-saporin. In contrast, GRPR-transfected cells were not killed by NMB-saporin (Fig. 5e). As expected, bombesin-saporin-conjugated toxin eliminated GRPR-transfected cells, but did not reduce numbers of NMBR-expressing cells (Fig. 5f). Therefore, NMB-saporin is a selective ablation agent for NMBR-expressing cells.
Ablation of TacR1 and GRPR-expressing neurons in the dorsal horn leads to the selective loss of thermal and mechanical nociceptive responses or itch responses, respectively (Nichols et al., 1999; Sun et al., 2009). Since NMB and GRP have similar signaling roles in the CNS (Battey and Wada, 1991; Ohki-Hamazaki et al., 1999; Ladenheim et al., 2002), we were interested if there is overlap in the function of NMBR and GRPR neurons in somatosensory signaling. We determined that a dose of 10 μg NMB-saporin was effective at ablating ∼80% of NMBR-expressing neurons. Importantly, toxin treatment did not reduce numbers of DRG neurons expressing NMBR. After administration of toxin, animals were injected subcutaneously at the lumbar back region with histamine agonist HTMT and the number of scratching bouts determined. Itch responses of animals given NMB-saporin were not significantly reduced compared with control animals, whereas bombesin-saporin treatment led to a dramatic reduction of responses (Fig. 6a). Thus, NMBR neurons are not required for responses to pruritic substances. Since NMB is expressed in many TRPV1 sensory neurons, we reasoned that the loss of NMBR neurons might have an effect on responses to heat. Thus, we tested mice using the Hargreaves test (Fig. 6b). NMB-saporin-treated mice exhibited considerably less sensitivity to heat stimulus with response latencies approximately twice as long as blank saporin-treated controls (Fig. 6b). In addition, it has been shown that SSP-saporin treatment produces deficits in reaction to mechanical stimulation (Nichols et al., 1999). Therefore, to measure responses to noxious mechanical stimuli, we induced inflammation in the plantar surface of the foot with CFA and used von Frey microfilaments to measure mechanical sensitivity. As expected, SSP-saporin-injected animals showed reduced reaction to noxious mechanical force (Nichols et al., 1999)(Fig. 6c). In contrast, NMB-saporin-treated mice exhibited reflex responses similar to control animals (Fig. 6c). In summary, eliminating NMBR-expressing neurons produces a reduction of responses to heat, but no change in reaction to itch or noxious mechanical stimulation.
Figure 6.

Ablation of NMBR-expressing dorsal horn neurons produces a selective deficit in noxious heat sensation. Itch sensation was assayed by quantitating bouts of itching following subcutaneous injection of the histamine agonist HTMT (a). Paw withdrawal latency was used to monitor thermal sensitivity (b) and mechanical sensitivity was measured using a von Frey device after sensitization of the paw with CFA (c). Intrathecal injection of bombesin-saporin (Bomb-sap) attenuates pruritic reaction, but NMB-saporin (NMB-sap) and blank-saporin (Blank-sap) do not alter itch responses compared with untreated controls. Administration of NMB-saporin resulted in an increased latency in response to thermal stimulation but did not change mechanical sensitivity. In contrast, SSP-saporin (SSP-sap) treatment led to attenuation of responses to both mechanical and noxious heat stimuli. Data represent means ± SEM; n ≥ 6 animals; significant differences between untreated controls and treatments (Student's t test) are indicated by **p < 0.01.
Given the distinct phenotype of mice with reduced numbers of neurons expressing NMBR, GRPR, and TacR1, we were interested in determining the effectiveness of toxins in eliminating cells expressing these receptors in the dorsal horn of the spinal cord. Examination of sections of untreated animals showed the numbers of NMBR- and GRPR-expressing neurons was approximately the same (Fig. 7a,c; mean of 14.2 ± 0.7 cells/16 μm sections in the lumbar spinal cord for NMBR and 9.0 ± 0.6 cells for GRPR). While NMBR and GRPR neurons are present predominantly in the superficial layers of the spinal cord, TacR1 is found in approximately four times as many cells throughout this region and also in deeper areas. For this reason we chose to score TacR1-positive neurons in laminar layer I through IV (mean of 43.8 ± 1.9 cells/16 μm sections in the lumbar spinal cord for TacR1). Injection of blank-saporin did not reduce numbers of NMBR-, GRPR-, or TacR1-positive neurons. Quantization of neurons expressing NMBR revealed that NMB-saporin treatment leads to loss of most NMBR-expressing neurons (Fig. 7d,g). As expected from our behavioral results, bombesin-saporin treatment does not significantly reduce numbers of NMBR-positive neurons but ablates GRPR-expressing neurons (Fig. 7e,h). SSP-saporin was less effective at eliminating TacR1-expressing cells, but, interestingly, numbers of NMBR-expressing neurons were reduced ∼65% after injection with this toxin (Fig. 7g). In addition, NMB toxin results in the elimination of significant numbers of TacR1-positive cells (including most lamina I neurons; Fig. 7g). These results indicate that there is a partial overlap in the expression of NMBR and TacR1 and suggests there are two separate classes of interneurons: cells that express NMBR and TacR1 and neurons that express GRPR but not NMBR or TacR1.
Figure 7.
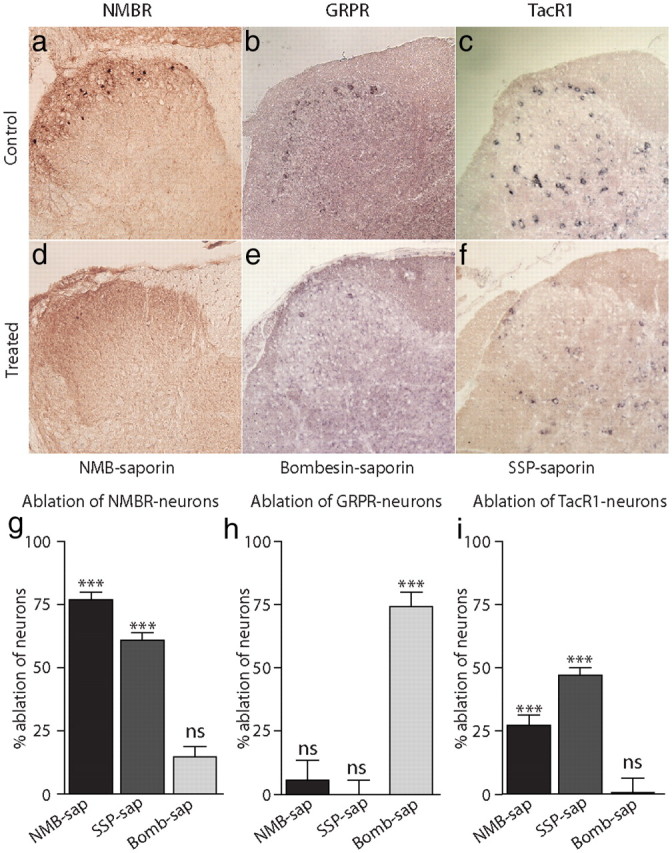
NMBR is expressed in a specific class of dorsal horn interneurons separated from GRPR-expressing neurons. Sections through the dorsal horn of spinal cord were examined for expression of NMBR by immunostaining for GFP in Tg(Nmbr-EGFP) transgenic mice (a, d, g). ISH was used to determine cells positive for GRPR (b, e, h) and TacR1 (c, f, i). NMBR, GRPR, and TacR1 are found in the dorsal horn (a–c) and treatment with specific conjugated peptide saporin (as indicated) led to ablation of most of their respective cell types (d–f). Quantification of NMBR-positive neurons normalized to blank-saporin controls reveals that treatment with NMB-saporin (NMB-sap) and SSP-saporin (SSP-sap) greatly reduces their numbers, but bombesin-saporin (Bomb-sap) has little effect (g). Ablation of GRPR-expressing cells only occurs following bombesin-saporin treatment (h). Administration of NMB-saporin and SSP-saporin eliminates ∼25 and 50% of TacR1-staining cells, respectively, and bombesin-saporin has no effect (i). Data represent means ± SEM of the levels of ablation relative to untreated control samples; n ≥ 3 animals; significant differences between treatments and untreated animals (Student's t test) are indicated by ***p < 0.001.
Discussion
We set out to discover new somatosensory signaling molecules in the TG. By comparing genes transcribed in two functionally distinct ganglia, we identified a small number of novel genes and validated their expression by ISH. These genes are presented in Table 1, and offer a resource of TG-specific candidate signaling molecules for the somatosensory research community. Some of the genes uncovered were expressed in subsets of neurons, suggestive of a modality-specific role (Fig. 1). The significance of these molecules is emphasized by their relative expression levels compared with known signaling molecules like TRPA1 and TRPM8 (McKemy et al., 2002; Peier et al., 2002; Story et al., 2003; Jordt et al., 2004). A number of molecules we identified could well be components of signaling pathways that have been previously partially characterized. For other molecules, further work will be needed to assign definitive functions. Our screen also revealed neuropeptides highly expressed in TG, including NMB. Although the function of NMB in the CNS has been studied (Battey and Wada, 1991; Ohki-Hamazaki et al., 1999; Ladenheim et al., 2002), its role in the peripheral nervous system is not clear (O'Donohue et al., 1984; De Koninck and Henry, 1989; Cridland and Henry, 1992), therefore, we performed experiments to better understand its function.
Peripheral sensory neurons are classified histologically and functionally as “peptidergic” or “nonpeptidergic” (Snider and McMahon, 1998; Patapoutian and Reichardt, 2001). We found that NMB is expressed with CGRP and TRPV1 (Fig. 3); therefore, NMB must be coexpressed also with SP in many neurons. A possible explanation for the coexpression of peptides is that they have partially redundant functions, and they combine to produce their effects. This concept is supported by the partial nociceptive phenotypes of mice lacking CGRP and SP (Cao et al., 1998; Salmon et al., 2001, 2004). Our studies also suggest that these neuropeptides combine to induce hyperalgesia and neurogenic swelling at a site of injury (Fig. 4). An additional potential role for NMB could be in feedback processes. Indeed, we find the receptor for NMB is expressed in sensory neurons; therefore, these cells could be activated by NMB (Fig. 5).
In addition to its role in the periphery, we discovered that NMB, akin to its bombesin family counterpart GRP, is probably also involved as a central neurotransmitter. We found that the receptor for NMB is present in neurons in the superficial lamina of the spinal cord (Fig. 5). When NMBR-expressing neurons are eliminated, there was no change in itch responses (Fig. 6). Therefore, NMB is not involved in pruritic signaling. Revealingly, NMB-saporin-treated mice exhibited specific attenuation of responses to heat (Fig. 6), consistent with the expression of NMB in TRPV1 neurons. The multiple neuropeptide expressed in the same neurons, the moderate blocking effects of individual antagonists (Lecci et al., 1991; Birch et al., 1992), and the partial phenotypes of neuropeptide knock-out mice (Cao et al., 1998; Salmon et al., 2001, 2004) all point to functional redundancy between nociceptive neuropeptides. Thus, for NMBR antagonists to be effective at ameliorating pain they will probably need to be combined with antagonists to other peptides or other therapeutic targets.
NMB-saporin administration attenuates responses to heat, but has no effect on noxious mechanical responses, demonstrating that NMBR-expressing neurons are selective for responses to thermal stimuli (Fig. 6). Moreover, we found indirect evidence that there is partial coexpression of NMBR with TacR1 (Fig. 7). TacR1-expressing neurons have previously been shown to be required for processing noxious thermal and mechanical pain (Nichols et al., 1999). This suggests that NMBR-TacR1-expressing cells could represent a specific class of interneuron involved in processing thermal heat reception. Sensory neurons send axons centrally, forming synapses with neurons in the dorsal horn of the spinal cord, with most nociceptive input occurring in the most superficial lamina. Recent molecular genetic studies demonstrate functionally defined subsets of sensory neuron target-specific lamina in the dorsal horn (Zylka et al., 2005; Liu et al., 2007; Seal et al., 2009; Li et al., 2011). These studies suggest that sensory afferents are topologically organized, with input of specific sensory modalities forming circuits with specific classes of interneurons found in particular zones of the spinal cord. Thus, our studies extend the concept that input into the dorsal horn is defined by selective connections and lays the foundation for future studies exploring the specificity of dorsal horn circuits.
In summary, we have identified NMB as both an agent that contributes to neurogenic inflammation and a potential neurotransmitter used by thermal nociceptive receptors.
Footnotes
This research was supported by the intramural research program of the National Institutes of Health—National Institute of Dental and Craniofacial Health (M.A.H.) The mouse strain used for this research project, STOCK Tg(Nmbr-EGFP), was obtained from the Mutant Mouse Regional Resource Center (MMRRC), a National Center for Research Resources– NIH-funded strain repository, and was donated to the MMRRC by the National Institute of Neurological Disorders and Stroke funded GENSAT BAC transgenic project. We are grateful to Nick Ryba for encouragement and helpful advice and Dr. Siraganian and the Hoon lab for valuable suggestions.
References
- Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJ, Zuker CS. A novel family of mammalian taste receptors. Cell. 2000;100:693–702. doi: 10.1016/s0092-8674(00)80705-9. [DOI] [PubMed] [Google Scholar]
- Battey J, Wada E. Two distinct receptor subtypes for mammalian bombesin-like peptides. Trends Neurosci. 1991;14:524–528. doi: 10.1016/0166-2236(91)90005-f. [DOI] [PubMed] [Google Scholar]
- Bergman E, Johnson H, Zhang X, Hökfelt T, Ulfhake B. Neuropeptides and neurotrophin receptor mRNAs in primary sensory neurons of aged rats. J Comp Neurol. 1996;375:303–319. doi: 10.1002/(SICI)1096-9861(19961111)375:2<303::AID-CNE9>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- Birch PJ, Harrison SM, Hayes AG, Rogers H, Tyers MB. The non-peptide NK1 receptor antagonist, (+/-)-CP-96,345, produces antinociceptive and anti-oedema effects in the rat. Br J Pharmacol. 1992;105:508–510. doi: 10.1111/j.1476-5381.1992.tb09008.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brain SD, Cox HM. Neuropeptides and their receptors: innovative science providing novel therapeutic targets. Br J Pharmacol 147 [Suppl] 2006;1:S202–S211. doi: 10.1038/sj.bjp.0706461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao YQ, Mantyh PW, Carlson EJ, Gillespie AM, Epstein CJ, Basbaum AI. Primary afferent tachykinins are required to experience moderate to intense pain. Nature. 1998;392:390–394. doi: 10.1038/32897. [DOI] [PubMed] [Google Scholar]
- Corjay MH, Dobrzanski DJ, Way JM, Viallet J, Shapira H, Worland P, Sausville EA, Battey JF. Two distinct bombesin receptor subtypes are expressed and functional in human lung carcinoma cells. J Biol Chem. 1991;266:18771–18779. [PubMed] [Google Scholar]
- Cridland RA, Henry JL. Bombesin, neuromedin C and neuromedin B given intrathecally facilitate the tail flick reflex in the rat. Brain Res. 1992;584:163–168. doi: 10.1016/0006-8993(92)90890-l. [DOI] [PubMed] [Google Scholar]
- Deems RO, Cardinaux F, Deacon RW, Young DA. Amylin or CGRP (8–37) fragments reverse amylin-induced inhibition of 14C-glycogen accumulation. Biochem Biophys Res Commun. 1991;181:116–120. doi: 10.1016/s0006-291x(05)81389-0. [DOI] [PubMed] [Google Scholar]
- De Felipe C, Herrero JF, O'Brien JA, Palmer JA, Doyle CA, Smith AJ, Laird JM, Belmonte C, Cervero F, Hunt SP. Altered nociception, analgesia and aggression in mice lacking the receptor for substance P. Nature. 1998;392:394–397. doi: 10.1038/32904. [DOI] [PubMed] [Google Scholar]
- De Koninck Y, Henry JL. Bombesin, neuromedin B and neuromedin C selectively depress superficial dorsal horn neurones in the cat spinal cord. Brain Res. 1989;498:105–117. doi: 10.1016/0006-8993(89)90404-6. [DOI] [PubMed] [Google Scholar]
- Folkers K, Håkanson R, Hörig J, Xu JC, Leander S. Biological evaluation of substance P antagonists. Br J Pharmacol. 1984;83:449–456. doi: 10.1111/j.1476-5381.1984.tb16506.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Schambra UB, Nowak NJ, Joyner A, Leblanc G, Hatten ME, Heintz N. A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature. 2003;425:917–925. doi: 10.1038/nature02033. [DOI] [PubMed] [Google Scholar]
- Heasley LE. Autocrine and paracrine signaling through neuropeptide receptors in human cancer. Oncogene. 2001;20:1563–1569. doi: 10.1038/sj.onc.1204183. [DOI] [PubMed] [Google Scholar]
- Hökfelt T. Neuropeptides in perspective: the last ten years. Neuron. 1991;7:867–879. doi: 10.1016/0896-6273(91)90333-u. [DOI] [PubMed] [Google Scholar]
- Hoon MA, Adler E, Lindemeier J, Battey JF, Ryba NJ, Zuker CS. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell. 1999;96:541–551. doi: 10.1016/s0092-8674(00)80658-3. [DOI] [PubMed] [Google Scholar]
- Jensen RT, Battey JF, Spindel ER, Benya RV. International Union of Pharmacology. LXVIII. Mammalian bombesin receptors: nomenclature, distribution, pharmacology, signaling, and functions in normal and disease states. Pharmacol Rev. 2008;60:1–42. doi: 10.1124/pr.107.07108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordt SE, Bautista DM, Chuang HH, McKemy DD, Zygmunt PM, Högestätt ED, Meng ID, Julius D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature. 2004;427:260–265. doi: 10.1038/nature02282. [DOI] [PubMed] [Google Scholar]
- Kline RH, 4th, Wiley RG. Spinal mu-opioid receptor-expressing dorsal horn neurons: role in nociception and morphine antinociception. J Neurosci. 2008;28:904–913. doi: 10.1523/JNEUROSCI.4452-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koga K, Chen T, Li XY, Descalzi G, Ling J, Gu J, Zhuo M. Glutamate acts as a neurotransmitter for gastrin releasing peptide-sensitive and insensitive itch-related synaptic transmission in mammalian spinal cord. Mol Pain. 2011;7:47. doi: 10.1186/1744-8069-7-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladenheim EE, Hampton LL, Whitney AC, White WO, Battey JF, Moran TH. Disruptions in feeding and body weight control in gastrin-releasing peptide receptor deficient mice. J Endocrinol. 2002;174:273–281. doi: 10.1677/joe.0.1740273. [DOI] [PubMed] [Google Scholar]
- Landry M, Holmberg K, Zhang X, Hökfelt T. Effect of axotomy on expression of NPY, galanin, and NPY Y1 and Y2 receptors in dorsal root ganglia and the superior cervical ganglion studied with double-labeling in situ hybridization and immunohistochemistry. Exp Neurol. 2000;162:361–384. doi: 10.1006/exnr.1999.7329. [DOI] [PubMed] [Google Scholar]
- Lecci A, Giuliani S, Patacchini R, Viti G, Maggi CA. Role of NK1 tachykinin receptors in thermonociception: effect of (+/-)-CP 96,345, a non-peptide substance P antagonist, on the hot plate test in mice. Neurosci Lett. 1991;129:299–302. doi: 10.1016/0304-3940(91)90485-c. [DOI] [PubMed] [Google Scholar]
- Lemons LL, Wiley RG. Galanin receptor-expressing dorsal horn neurons: Role in nociception. Neuropeptides. 2011;45:377–383. doi: 10.1016/j.npep.2011.08.002. [DOI] [PubMed] [Google Scholar]
- Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S, Jankowski MP, Luo W, Heintz N, Koerber HR, Woodbury CJ, Ginty DD. The functional organization of cutaneous low-threshold mechanosensory neurons. Cell. 2011;147:1615–1627. doi: 10.1016/j.cell.2011.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Molecular genetic visualization of a rare subset of unmyelinated sensory neurons that may detect gentle touch. Nat Neurosci. 2007;10:946–948. doi: 10.1038/nn1937. [DOI] [PubMed] [Google Scholar]
- Massari VJ, Tizabi Y, Park CH, Moody TW, Helke CJ, O'Donohue TL. Distribution and origin of bombesin, substance P and somatostatin in cat spinal cord. Peptides. 1983;4:673–681. doi: 10.1016/0196-9781(83)90017-7. [DOI] [PubMed] [Google Scholar]
- McKemy DD, Neuhausser WM, Julius D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature. 2002;416:52–58. doi: 10.1038/nature719. [DOI] [PubMed] [Google Scholar]
- Mishra SK, Hoon MA. Ablation of TrpV1 neurons reveals their selective role in thermal pain sensation. Mol Cell Neurosci. 2010;43:157–163. doi: 10.1016/j.mcn.2009.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namba M, Ghatei MA, Gibson SJ, Polak JM, Bloom SR. Distribution and localization of neuromedin B-like immunoreactivity in pig, cat and rat spinal cord. Neuroscience. 1985;15:1217–1226. doi: 10.1016/0306-4522(85)90264-7. [DOI] [PubMed] [Google Scholar]
- Nichols ML, Allen BJ, Rogers SD, Ghilardi JR, Honore P, Luger NM, Finke MP, Li J, Lappi DA, Simone DA, Mantyh PW. Transmission of chronic nociception by spinal neurons expressing the substance P receptor. Science. 1999;286:1558–1561. doi: 10.1126/science.286.5444.1558. [DOI] [PubMed] [Google Scholar]
- O'Donohue TL, Massari VJ, Pazoles CJ, Chronwall BM, Shults CW, Quirion R, Chase TN, Moody TW. A role for bombesin in sensory processing in the spinal cord. J Neurosci. 1984;4:2956–2962. doi: 10.1523/JNEUROSCI.04-12-02956.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohki-Hamazaki H, Sakai Y, Kamata K, Ogura H, Okuyama S, Watase K, Yamada K, Wada K. Functional properties of two bombesin-like peptide receptors revealed by the analysis of mice lacking neuromedin B receptor. J Neurosci. 1999;19:948–954. doi: 10.1523/JNEUROSCI.19-03-00948.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orbuch M, Taylor JE, Coy DH, Mrozinski JE, Jr, Mantey SA, Battey JF, Moreau JP, Jensen RT. Discovery of a novel class of neuromedin B receptor antagonists, substituted somatostatin analogues. Mol Pharmacol. 1993;44:841–850. [PubMed] [Google Scholar]
- Patapoutian A, Reichardt LF. Trk receptors: mediators of neurotrophin action. Curr Opin Neurobiol. 2001;11:272–280. doi: 10.1016/s0959-4388(00)00208-7. [DOI] [PubMed] [Google Scholar]
- Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM, Earley TJ, Dragoni I, McIntyre P, Bevan S, Patapoutian A. A TRP channel that senses cold stimuli and menthol. Cell. 2002;108:705–715. doi: 10.1016/s0092-8674(02)00652-9. [DOI] [PubMed] [Google Scholar]
- Polgár E, Furuta T, Kaneko T, Todd A. Characterization of neurons that express preprotachykinin B in the dorsal horn of the rat spinal cord. Neuroscience. 2006;139:687–697. doi: 10.1016/j.neuroscience.2005.12.021. [DOI] [PubMed] [Google Scholar]
- Salio C, Lossi L, Ferrini F, Merighi A. Neuropeptides as synaptic transmitters. Cell Tissue Res. 2006;326:583–598. doi: 10.1007/s00441-006-0268-3. [DOI] [PubMed] [Google Scholar]
- Salmon AM, Damaj MI, Marubio LM, Epping-Jordan MP, Merlo-Pich E, Changeux JP. Altered neuroadaptation in opiate dependence and neurogenic inflammatory nociception in alpha CGRP-deficient mice. Nat Neurosci. 2001;4:357–358. doi: 10.1038/86001. [DOI] [PubMed] [Google Scholar]
- Salmon AM, Evrard A, Damaj I, Changeux JP. Reduction of withdrawal signs after chronic nicotine exposure of alpha-calcitonin gene-related peptide knock-out mice. Neurosci Lett. 2004;360:73–76. doi: 10.1016/j.neulet.2004.02.031. [DOI] [PubMed] [Google Scholar]
- Seal RP, Wang X, Guan Y, Raja SN, Woodbury CJ, Basbaum AI, Edwards RH. Injury-induced mechanical hypersensitivity requires C-low threshold mechanoreceptors. Nature. 2009;462:651–655. doi: 10.1038/nature08505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snider WD, McMahon SB. Tackling pain at the source: new ideas about nociceptors. Neuron. 1998;20:629–632. doi: 10.1016/s0896-6273(00)81003-x. [DOI] [PubMed] [Google Scholar]
- Story GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, Hricik TR, Earley TJ, Hergarden AC, Andersson DA, Hwang SW, McIntyre P, Jegla T, Bevan S, Patapoutian A. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell. 2003;112:819–829. doi: 10.1016/s0092-8674(03)00158-2. [DOI] [PubMed] [Google Scholar]
- Sun YG, Chen ZF. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature. 2007;448:700–703. doi: 10.1038/nature06029. [DOI] [PubMed] [Google Scholar]
- Sun YG, Zhao ZQ, Meng XL, Yin J, Liu XY, Chen ZF. Cellular basis of itch sensation. Science. 2009;325:1531–1534. doi: 10.1126/science.1174868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada E, Way J, Lebacq-Verheyden AM, Battey JF. Neuromedin B and gastrin-releasing peptide mRNAs are differentially distributed in the rat nervous system. J Neurosci. 1990;10:2917–2930. doi: 10.1523/JNEUROSCI.10-09-02917.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang MW, Young AA, Rink TJ, Cooper GJ. 8–37h-CGRP antagonizes actions of amylin on carbohydrate metabolism in vitro and in vivo. FEBS Lett. 1991;291:195–198. doi: 10.1016/0014-5793(91)81282-d. [DOI] [PubMed] [Google Scholar]
- Wiley RG, Lappi DA. Targeted toxins in pain. Adv Drug Deliv Rev. 2003;55:1043–1054. doi: 10.1016/s0169-409x(03)00102-9. [DOI] [PubMed] [Google Scholar]
- Wiley RG, Lemons LL, Kline RH., 4th Neuropeptide Y receptor-expressing dorsal horn neurons: role in nocifensive reflex responses to heat and formalin. Neuroscience. 2009;161:139–147. doi: 10.1016/j.neuroscience.2008.12.017. [DOI] [PubMed] [Google Scholar]
- Xu XJ, Hökfelt T, Wiesenfeld-Hallin Z. Galanin and spinal pain mechanisms: past, present, and future. EXS. 2010;102:39–50. doi: 10.1007/978-3-0346-0228-0_4. [DOI] [PubMed] [Google Scholar]
- Zhang X, Xu ZQ, Bao L, Dagerlind A, Hökfelt T. Complementary distribution of receptors for neurotensin and NPY in small neurons in rat lumbar DRGs and regulation of the receptors and peptides after peripheral axotomy. J Neurosci. 1995;15:2733–2747. doi: 10.1523/JNEUROSCI.15-04-02733.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Shi T, Holmberg K, Landry M, Huang W, Xiao H, Ju G, Hökfelt T. Expression and regulation of the neuropeptide Y Y2 receptor in sensory and autonomic ganglia. Proc Natl Acad Sci U S A. 1997;94:729–734. doi: 10.1073/pnas.94.2.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Xu ZO, Shi TJ, Landry M, Holmberg K, Ju G, Tong YG, Bao L, Cheng XP, Wiesenfeld-Hallin Z, Lozano A, Dostrovsky J, Hökfelt T. Regulation of expression of galanin and galanin receptors in dorsal root ganglia and spinal cord after axotomy and inflammation. Ann N Y Acad Sci. 1998;863:402–413. doi: 10.1111/j.1749-6632.1998.tb10710.x. [DOI] [PubMed] [Google Scholar]
- Zylka MJ, Rice FL, Anderson DJ. Topographically distinct epidermal nociceptive circuits revealed by axonal tracers targeted to Mrgprd. Neuron. 2005;45:17–25. doi: 10.1016/j.neuron.2004.12.015. [DOI] [PubMed] [Google Scholar]