

I consider that a man's brain originally is like a little empty attic, and you have to stock it with such furniture as you choose.
Sir Arthur Conan Doyle. Spoken by Sherlock Holmes in A Study in Scarlet, chapter 2.
It has been said that the solutions to many of the problems of physics are to be found in the attic of mathematics. The history of the interplay between chemistry and biology reflects a similar relationship. Many compounds—and some concepts—no longer at the cutting edge of chemical research have proven to be invaluable tools in cutting-edge biology and medicine. Perhaps the most dramatic example is the extraordinary success of cisplatin (Peyrone's salt), a compound known for over 100 years, as an anticancer drug. This extremely simple inorganic substance, often used as a basic example of isomerism in freshman chemistry textbooks, was found by Barnett Rosenberg et al. in 1969 to be a powerful antitumor agent (1), is now a complete cure for testicular cancer if the disease is caught early enough, and has shown effectiveness, particularly in combination therapy, against many other cancers as well. In a recent issue of PNAS, Yan et al. (2) provide a new example. They report that beryllofluoride, another simple inorganic compound, can be used to form stable analogs of the normally unstable phosphorylated form of bacterial signal transduction response regulator proteins, finally making the active state of these molecules amenable to detailed structural study.
Response regulator proteins are the switches that control the response of bacteria to external stimuli. They constitute the second half of the so-called “two-component” signal transduction systems; the other half is an autokinase (3). The kinase phosphorylates an aspartate residue on the response regulator, converting the regulator protein structure from an “off” state to an “on” state. Phosphorylated regulators may turn on or repress the transcription of virulence genes (4), switch on the flagellar motor apparatus (5), control antibiotic synthesis (6), or initiate sporulation (7), among other functions. Structure/function studies of such signal transduction pathways are well advanced (for a review, see ref. 8), but there has been one glaring deficiency: studies of the response regulators have for the most part had to be confined to the off state, the inactive, nonphosphorylated form of these proteins. The reason is a simple one. Aspartylphosphates have extremely short life-times in aqueous solution.
Overcoming this sort of problem is old hat to biochemists who study ATP- and GTPase enzymes. They use nonhydrolyzable nucleotide triphosphate analogs to generate the ground-state, ATP- or GTP-bound forms of the protein, and, to characterize the transition state for phosphate ester hydrolysis, they employ metal complexes such as vanadates and AlF4− (9). In the case of the two-component response regulators, however, it is the ground-state structure of a covalent phosphoprotein that is needed, and analogs that could generate this state have proven elusive. Enter Yan et al. (2) and beryllofluoride (Fig. 1).
Figure 1.

AlF4− and BeF3−⋅H2O (or BeF42−) have been used as analogs of the tetrahedral phosphate group.
They did not set out to find a simple compound that could trap the regulator in its on state. Rather, they were looking at a downstream effector whose activity is controlled by a particular regulator, an effector that happens to be a transcriptional activator with ATPase activity. Often, such effectors are actually part of the regulator protein; they constitute a biological output domain that is fused to the regulatory domain. NtrC, the particular effector they were looking at, is of this type. Yan et al. (2) hoped to use ATP analogs to inhibit the ATPase activity of the effector domain, but they found that neither vanadates nor aluminofluorides were able to block this reaction. One of the compounds they tested, however, stimulated the ATPase activity, and further analysis showed that this stimulation was caused by the compound forming an acylphosphate analog in the regulatory domain, forcing it to adopt the on conformation. The compound was beryllofluoride (2).
Beryllium is a very interesting metal to chemists. Although it is a Group 2A (alkaline earth) metal, its chemistry is somewhat different from that of magnesium, calcium, strontium or barium. Because of its small size, the beryllium atom has a high ionization energy and thus does not tend to form the Be2+ ion. Molten BeF2 is a poor electrical conductor, suggesting that it is essentially a covalent compound. The other striking characteristic of beryllium is its tendency to form compounds in which it has an incomplete octet of valence electrons, a tendency shared only by aluminum and boron. Because the octet rule can be violated, a number of different compounds of beryllium with, for example, fluorine, can be made. One of them, BeF42−, satisfies the octet rule, but BeF2 does not, and neither does BeF3−. These latter compounds have an electron-deficient beryllium atom, which can accept pairs of electrons to complete an octet if suitable coordinate covalent bond donors are available. Water is such a donor, so the species BeF2⋅OH−⋅H2O and BeF3−⋅H2O predominate in aqueous solution, along with BeF42−; their relative amounts depend, among other things, on the ratio of F to Be. Another such donor would be the lone pair of electrons on the oxygen atom of either a phosphate group or an aspartate carboxylate group. Note that all of these beryllofluorides would be tetrahedral, with Be-F bond lengths of 1.55 Å, making them strictly isomorphous to a phosphate group.
That beryllofluoride—usually written BeFx to indicate that a mixture of species can be formed and the exact one responsible for any given activity may not be known—can be a useful analog of a phosphate ion for biological studies has been known for over 10 years. Chabre and colleagues demonstrated in 1987 that not just AlF4− but also BeFx could form reversibly bound analogs of the γ phosphate group of GTP in the nucleotide binding site of transducin and, in the presence of GDP, could activate that protein (10). Activation could be totally reversed by elution of fluoride. Combeau and Carlier showed that BeF2⋅OH−⋅H2O and BeF3−⋅H2O could bind tightly to F-actin and microtubules (11), mimicking the XDP-Pi transient state of polymerization as depicted in Fig. 2.
Figure 2.

Proposed binding of BeF3−⋅H2O to magnesium–nucleotide diphosphates to form a stable analog of the nucleotide triphosphate species. Compare with the actual structure in Fig. 3.
Since these pioneering studies, aluminum and beryllium fluorides have been widely employed in the fields of muscle contraction and eukaryotic signal transduction (12). Recent examples include the use of BeFx-ADP by Eady and colleagues to study the mechanism of nitrogenase (13) and of MgADP⋅BeFx and MgADP⋅AlF4− by Rayment and associates in crystallographic studies of the motor domain of myosin (14). The myosin study is particularly interesting, because the structural data suggests that MgADP⋅AlF4− is an analog of the transition state for ATP hydrolysis, just as in the case of the GTPases whereas MgADP⋅BeFx appears to mimic the ATP-bound state (see Fig. 3). This result is consistent with the obligatory tetrahedral geometry of BeFx species in aqueous solution, implying that the bound species is either MgADP⋅BeF3 or MgADP⋅BeF2(OH), with an oxygen atom from the β phosphate group of ADP forming a coordinate covalent bond to the beryllium atom, thereby completing the Be octet (15).
Figure 3.
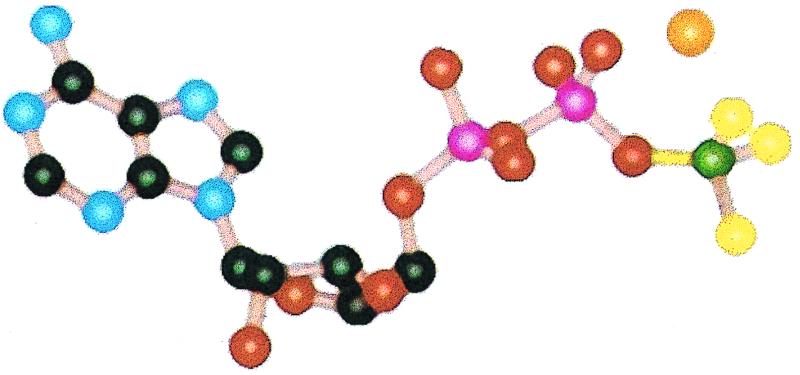
The structure of MgADP⋅BeFx in the ATP-binding site of myosin. Be, C, F, Mg2+, N, O, and P are depicted in green, black, yellow, orange, blue, red, and pink respectively. Adapted from ref. 14. Compare with Fig. 2.
Although Yan et al. (2) are unable to exclude BeF42− as the species that activates NtrC, these considerations would appear to favor either BeF3−⋅H2O as the activator via phosphate mimicry, or, more interestingly, aspartyl⋅BeF3− as a direct analog of aspartylphosphate. Strong evidence for the latter is provided by their demonstration that mutants in which the carboxylate is lost are unable to be activated by beryllofluoride. The ability of BeFx to activate two-component regulators appears to be a general one: Yan et al. (2) have been able to activate regulatory proteins in chemotaxis, sporulation, and σ-dependent transcriptional regulation as well (2). Preliminary NMR data suggest that conformational changes induced by phosphorylation of aspartate residues in all of these proteins are reproduced on treatment with Mg2+ and BeFx, although quantitative conversion required some fiddling with the concentrations of these species. Because the preparation of BeFx is extremely simple [one just mixes BeCl2 with a large excess of NaF (15)], it seems likely that the work of Yan et al. (2) will open a floodgate of structural studies on microbial two-component regulator proteins, by NMR and x-ray crystallography. Who knows what other useful tools for the structural biologist and biochemist are lying around in the attic of chemistry?
Footnotes
See companion article on page 14789 in issue 26 of volume 96.
References
- 1.Rosenberg B, VanCamp L, Trosko J E, Mansour V H. Nature (London) 1969;222:385–386. doi: 10.1038/222385a0. [DOI] [PubMed] [Google Scholar]
- 2.Yan D, Cho H S, Hastings C A, Igo M M, Lee S-Y, Pelton J G, Stewart V, Wemmer D E, Kustu S. Proc Natl Acad Sci USA. 1999;96:14789–14794. doi: 10.1073/pnas.96.26.14789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Goudreau P N, Stock A M. Curr Opin Microbiol. 1998;1:160–169. doi: 10.1016/s1369-5274(98)80006-4. [DOI] [PubMed] [Google Scholar]
- 4.Heath A, DiRita V J, Barg N L, Engleberg N C. Infect Immun. 1999;67:5298–5305. doi: 10.1128/iai.67.10.5298-5305.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Falke J J, Bass R B, Butler S L, Chervitz S A, Danielson M A. Annu Rev Cell Dev Biol. 1997;13:457–512. doi: 10.1146/annurev.cellbio.13.1.457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sahl H G. Ciba Found Symp. 1994;186:27–42. doi: 10.1002/9780470514658.ch3. [DOI] [PubMed] [Google Scholar]
- 7.Fisher S H. Mol Microbiol. 1999;32:223–232. doi: 10.1046/j.1365-2958.1999.01333.x. [DOI] [PubMed] [Google Scholar]
- 8.Djordjevic S, Stock A M. J Struct Biol. 1998;124:189–200. doi: 10.1006/jsbi.1998.4034. [DOI] [PubMed] [Google Scholar]
- 9.Kleuss C, Raw A S, Lee E, Sprang S R, Gilman A G. Proc Natl Acad Sci USA. 1994;91:9828–9831. doi: 10.1073/pnas.91.21.9828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bigay J, Deterre P, Pfister C, Chabre M. EMBO J. 1987;6:2907–2913. doi: 10.1002/j.1460-2075.1987.tb02594.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Combeau C, Carlier M F. J Biol Chem. 1989;264:19017–19021. [PubMed] [Google Scholar]
- 12.Chabre M. Trends Biochem Sci. 1990;15:6–10. doi: 10.1016/0968-0004(90)90117-t. [DOI] [PubMed] [Google Scholar]
- 13.Clarke T A, Yousafzai F K, Eady R R. Biochemistry. 1999;38:9906–9913. doi: 10.1021/bi9904353. [DOI] [PubMed] [Google Scholar]
- 14.Fisher A J, Smith C A, Thoden J B, Smith R, Sutoh K, Holden H M, Rayment I. Biochemistry. 1995;34:8960–8972. doi: 10.1021/bi00028a004. [DOI] [PubMed] [Google Scholar]
- 15.Lunardi J, Dupuis A, Garin J, Issartel J P, Michel L, Chabre M, Vignais P V. Proc Natl Acad Sci USA. 1988;85:8958–8962. doi: 10.1073/pnas.85.23.8958. [DOI] [PMC free article] [PubMed] [Google Scholar]