

Abstract
The antennal lobe is the primary olfactory center in the insect brain and represents the anatomical and functional equivalent of the vertebrate olfactory bulb1-5. Olfactory information in the external world is transmitted to the antennal lobe by olfactory sensory neurons (OSNs), which segregate to distinct regions of neuropil called glomeruli according to the specific olfactory receptor they express. Here, OSN axons synapse with both local interneurons (LNs), whose processes can innervate many different glomeruli, and projection neurons (PNs), which convey olfactory information to higher olfactory brain regions.
Optical imaging of the activity of OSNs, LNs and PNs in the antennal lobe - traditionally using synthetic calcium indicators (e.g. calcium green, FURA-2) or voltage-sensitive dyes (e.g. RH414) - has long been an important technique to understand how olfactory stimuli are represented as spatial and temporal patterns of glomerular activity in many species of insects6-10. Development of genetically-encoded neural activity reporters, such as the fluorescent calcium indicators G-CaMP11,12 and Cameleon13, the bioluminescent calcium indicator GFP-aequorin14,15, or a reporter of synaptic transmission, synapto-pHluorin16 has made the olfactory system of the fruitfly, Drosophilamelanogaster, particularly accessible to neurophysiological imaging, complementing its comprehensively-described molecular, electrophysiological and neuroanatomical properties2,4,17. These reporters can be selectively expressed via binary transcriptional control systems (e.g. GAL4/UAS18, LexA/LexAop19,20, Q system21) in defined populations of neurons within the olfactory circuitry to dissect with high spatial and temporal resolution how odor-evoked neural activity is represented, modulated and transformed22-24.
Here we describe the preparation and analysis methods to measure odor-evoked responses in the Drosophila antennal lobe using G-CaMP25-27. The animal preparation is minimally invasive and can be adapted to imaging using wide-field fluorescence, confocal and two-photon microscopes.
Keywords: Neuroscience, Issue 61, Drosophila, calcium imaging, antennal lobe, olfaction, neuroscience
Protocol
1. Mounting Block Assembly
The plexiglass mounting block (Figure 1A) should be made to the exact specifications given in the blueprint (available from http://neuro.uni-konstanz.de/jove_mountingblock_blueprint) to allow easy mounting and dissection of the fly. Thread through the screws from the back but do not extend them beyond the front of the block; their position will be adjusted during the preparation of the animal (see 2.7).
Using a precision drill, make a dent in the upper border of the circular chamber for the fly, digging under the surface to make space for the fly's body, reducing the thickness of the block at this point.
Cut a copper grid with a scalpel perpendicular and close to the bottom of the slit to create a "collar". Make sure the two resulting flaps are level, as this will help insertion of the fly.
With a toothpick, place a small drop of superglue onto the mounting block above the circular chamber for the fly. Place the copper grid onto the glue and position so that the slit is centered on the block (Figure 1A). Press down gently and remove any excess glue. With a pair of tweezers, add tiny drops of glue under the flaps on each side and fold the grid over the front of the block so that the slit is pointing straight downwards. Make sure the top of the grid is level with the block. Allow to dry completely.
Assemble a wire holder: cut a plastic coverslip in half and remove a central piece to make a "U" shape. Use beeswax to glue a piece of wire across the top of the "U"; do not make the wire too taut.
Construct the antennal shield: make a hole in a plastic coverslip with a paper hole-punch. Trim the edges of the coverslip to 0.5 x 0.7 cm. Glue the pierced plastic coverslip to sticky tape and then place a drop of ethanol on the tape exposed through the hole to remove the adhesive.
2. Animal Preparation
Flies of the appropriate genotype - i.e. bearing the desired combination of "driver" and activity-reporter transgenes - should be 1-3 weeks old; the cuticle of younger flies is too soft and difficult to cut. When experiments require direct comparison of different genotypes, and in order to match transgene expression levels, flies should be age-matched to avoid age-related physiological differences. If gender is not important, we prefer to use female flies since they have larger heads and are easier to manipulate.
Anesthetize flies by cooling on ice.
Introduce a single fly into the mounting block. Hold the fly by the wing joint and slide it, dorsal-surface first, along the slit of the copper grid so that it is suspended by its neck (Figure 1B-C). If necessary, rotate it until it is looking down.
Place a fine cactus spine in front of the fly's head to prevent it from escaping (Figure 1B-C). Place the cactus spine in front of the proboscis to prevent it from moving. Fix the cactus spine to the block using beeswax. Make sure that the top of the head is level with the top of the block.
Fix the head of the fly to the block with a small drop of rosin (dissolved in ethanol). Place the block in a humidified box and allow the rosin to harden for about one hour.
Using forceps, pull the antennal plate forward and drop the wire on the holder (wax-side out; see 1.5) in the cuticular fold between the antenna and head. Fix the holder to the front of the block with beeswax.
Using the screws built into the block, gently push the wire holder forward to create some space between the antennal plate and the head.
Place the antennal shield (see 1.6) over the top of the fly's head with the hole positioned centrally. Fix with beeswax to the top of the mounting block.
Using a blade-splitter, cut a small hole in the tape on the shield exposing the cuticle of the fly's head behind the antenna. The hole should not extend beyond the eyes to prevent the preparation from leaking.
Seal the hole between the tape and the cuticle with two-component silicon. Remove any excess silicon from the top of the fly's head.
Place a drop of Drosophila Ringer's saline (130 mM NaCl, 5 mM KCl, 2 mM MgCl2, 2 mM CaCl2, 36 mM saccharose, 5 mM HEPES, pH 7.3) on the head. Make sure that there are no leaks: the antennae should remain completely dry. Using a sapphire blade, cut a hole in the cuticle. First, lightly score along the cuticular fold directly behind the antenna, then cut along the eyes and across the ocelli on the back. Finally, fold the piece of cuticle over the scored edge and cut from the underside to avoid severing the antennal nerves.
Carefully remove glands and trachea with fine tweezers. Rinse repeatedly with Drosophila Ringer's, leaving a large drop on the head.
3. Odor Stimulus Delivery
Odor syringes: insert a cellulose pad into a 2 ml plastic syringe with clean forceps and pipette onto it (using filter tips) 20 μl of odor, diluted as desired in an appropriate solvent, using the pipette tip to push the pad deep into the barrel. Plug and cap the syringe until required. Fresh odor dilutions should be prepared at least every 3 months (or more frequently for highly volatile and/or often-used odors), and new pads should be prepared for odor syringes just before each recording session. Do not use odor pads for more than three stimulations, since the odor concentration might decay.
Custom-made olfactometer (Figure 2): a primary air stream (1 l/min) is directed through Teflon tubing (0.4 mm internal diameter); 27 cm upstream of the exit of this tube, a secondary air stream (1 l/min) is added. Both air streams are generated by membrane pumps, and the flow rate is regulated by two independent flowmeters. The air streams can be humidified by passing through water in gas washing bottles, although care should be taken to avoid over-humidification (i.e. more than ~60%). The secondary air stream is directed either through an odor syringe or an empty syringe: syringes are connected to the airflow through a syringe needle (1.2 mm outer diameter) piercing the plunger. A three-way magnetic valve controlled via a valve controller unit (e.g. Harvard Apparatus VC6) is used to switch between the clean air and odor stimulus.
4. In vivo Imaging
We describe here imaging using an upright fluorescent microscope (e.g. upright fixed Stage Zeiss Axio Examiner D1) equipped with a 20x water objective (e.g. Zeiss W "Plan-Apochromat" 20x/1,0 M27 DIC) (Figure 2). For imaging with G-CaMP (excitation/emission: 488/509 nm), use a filter block with the following properties: 450-490 nm excitation filter, dichroic mirror (T495LP) and 500-550 nm emission filter (Chroma ET). Place the mounting block containing the fly under the microscope. Position the exit of the olfactometer approximately 0.5 cm in front of the fly's antennae.
Lower the objective until it touches the Ringer's solution on the fly's head. Looking through the viewer under bright-field illumination, position the preparation so that the hole in the head capsule is centered and its borders are in focus. Switch to fluorescent light and adjust the focus until you can see the labeled neurons (apparent from basal green fluorescence of G-CaMP).
Fine focus on the glomeruli of interest by acquiring images with a Charge-coupled device (CCD) camera (e.g. CoolSNAP-HQ2 Digital Camera System) mounted on the microscope using appropriate acquisition software (e.g. Metafluor) (Figure 3A). Close the diaphragm in the illumination light path to limit excitation light to the imaged area.
For recording odor-evoked activity, set the acquisition rate to 4 Hz and adjust the exposure time to obtain fluorescence values within the dynamic range of your CCD camera, but not lower than 1000 (arbitrary units), with at least 12-bit dynamic resolution. The exposure time should not exceed 100 ms, if possible, to reduce G-CaMP photobleaching. If the spatial resolution of your camera allows, you can increase the binning (number of wells on the chip that will be binned into one digital pixel) to reduce the exposure time. We obtain images of 350x225 pixels (with a binning of 2x2), corresponding to 210x135 μm at the preparation. These settings are optimal to capture glomerular odor-induced responses of G-CaMP with good spatial and temporal resolution while limiting reporter bleaching.
Set the measurement parameters. We typically record for 10 seconds (i.e. 40 frames), with odor stimulation typically starting 4 seconds after the beginning of the acquisition period (frame 15) and lasting one second (until frame 19). A fly preparation can normally be tested with up to 25-30 different odor stimuli, often limited by an irreversible decline in the fluorescent signal of the reporter due to photobleaching. The inter-stimulus interval will depend on the strength of response observed and the rate of photobleaching. The appropriate length of inter-stimulus interval to avoid sensory adaptation can be determined approximately by repeated presentation of a positive control odor (if known) in trial experiments. Inclusion of this odor every ~10 stimuli in a given series can allow verification of the continued viability and consistent responsiveness of the preparation.
5. Data Processing and Analysis
Data is processed using ImageJ (with Mac biophotonics plug-in, www.macbiophotonics.ca) and custom written programs in Matlab and R as follows:
Correct for sample movement artifacts: align all the images corresponding to one animal preparation (e.g. 30 measurements of 40 frames each) using the "rigid body" mode of StackReg/TurboReg (bigwww.epfl.ch/thevenaz/stackreg) in ImageJ. This step is required to remove shifts in the position of the labeled neurons with respect to the acquisition frame within and between measurements. However, it is only possible to remove shifts in the xy plane. Data should be discarded if strong changes in focus occur.
Segment the stack into individual 10-second (40-frame) measurements.
Correct for bleaching artifacts: exposure to fluorescent light will cause fluorescence to decay during a single measurement10. In some cases, it is important to correct for this decay before analyzing the data. For each measurement calculate a background frame. This should be the mean of several frames before stimulus onset (e.g. frames 6-14). Divide each frame by this background frame to obtain the relative fluorescence changes. Calculate the mean relative fluorescence value for each frame by averaging all pixels within the area labeled by G-CaMP. Fit a double exponential to the mean fluorescence curve, giving stronger weight to the frames before the stimulus and zero weight to the time of the odor response (20 frames after stimulus onset in our case). Subtract the fitted curve from the relative fluorescence curve of each pixel. Multiply the corrected frames by the background frame to re-obtain raw data. This kind of correction may lead to a modification in the response dynamics if the signal does not return to the baseline by the end of the measurement (e.g. if inhibitory or very strong excitatory responses are evoked) (Figure 4). While correction of bleaching artifacts is useful in many cases, interpretation of temporal parameters should be made with caution if a bleach correction is applied.
Calculate the relative change in fluorescence (ΔF/F) for each frame of each measurement as (Fi-F0)/F0*100, where F0 is the background frame (see 5.3), and Fi the fluorescence value for the ith frame of the measurement.
Calculate the mean ΔF/F within a circular region of interest in a glomerulus (Figure 3B) with a diameter of 10 pixels to obtain activity traces (Figure 5A). If desired, transform these traces into heatmaps using ImageJ (Figure 5A).
Calculate the peak amplitude of the response by subtracting the mean ΔF/F of four frames before stimulation from the mean ΔF/F of four frames during the peak of the response. The same procedure is used both to illustrate spatial response patterns (see Figure 3B) and to quantify the response amplitude in specific regions of interest across flies (see Figure 5B). The peak response can also be quantified by integrating the area below the curve during the time of the stimulus, or averaging the frames around the maximum of the response.
6. Representative Results
We used the protocols above to record and analyze propionic acid-evoked responses of flies expressing G-CaMP1.628 under the control of the promoter for the olfactory co-receptor IR8a, which is active in several different populations of OSNs29,30 (Figures 3-5). Stimulation with 1% propionic acid elicits increases in G-CaMP fluorescence - indicating increases in intracellular calcium - in OSNs innervating two glomeruli, DC4 and DP1l (Figure 3B). We illustrate different ways of representing data from multiple flies in Figure 5: line activity traces, heatmaps and a bar plot of peak responses. Propionic acid-evoked responses have distinct peak amplitudes and temporal dynamics in DC4 and DP1l. Moreover, although responses are generally robust, variability between individual animals can also be observed in both glomeruli (see heatmaps in Figure 5A). Quantification of the peak responses (Figure 5B) allows a simple statistical comparison between different glomeruli and different odorants across animals. However, odor-evoked calcium responses can have non-uniform temporal dynamics, and quantifying their magnitude with a single value will disregard this complexity.
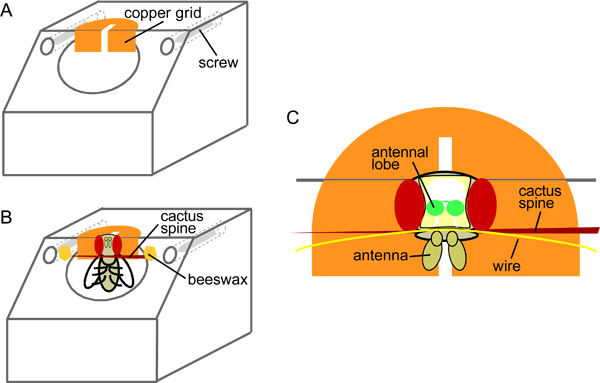 Figure 1. Schematic of the mounting block (A) before and (B) after introduction of the fly, and (C) a top view in close-up (adapted from31).
Figure 1. Schematic of the mounting block (A) before and (B) after introduction of the fly, and (C) a top view in close-up (adapted from31).
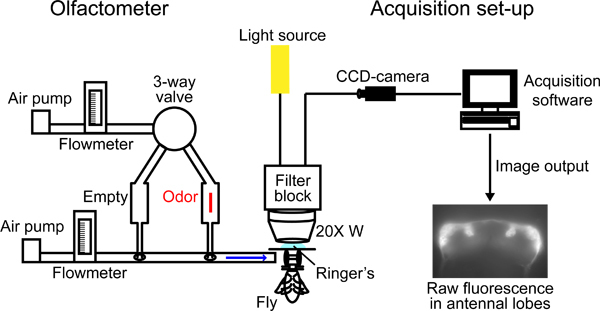 Figure 2. Schematic of the experimental set-up.
Figure 2. Schematic of the experimental set-up.
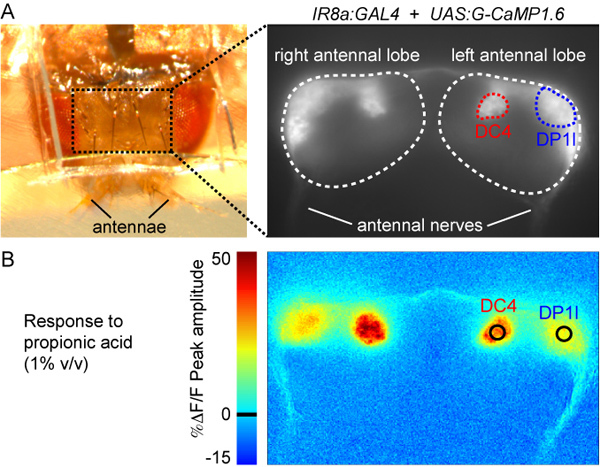 Figure 3. (A) Top view of the fly preparation after step 2.8 (left panel) and raw fluorescence image of the antennal lobes labeled with G-CaMP1.6 (Genotype: IR8a promoter:GAL4; UAS:G-CaMP1.6). DC4 and DP1l glomeruli can be distinguished by their morphology, size and position and are marked in red and blue, respectively.
(B) Color-coded image of the response to propionic acid (1% v/v). The image corresponds to the ΔF/F of the peak of the response calculated as explained in step 5.6 (see also Figure 5A). The colorscale on the left indicates the %ΔF/F value. The black circles indicate the position and size of the region of interest used to quantify the responses (see Figure 5).
Figure 3. (A) Top view of the fly preparation after step 2.8 (left panel) and raw fluorescence image of the antennal lobes labeled with G-CaMP1.6 (Genotype: IR8a promoter:GAL4; UAS:G-CaMP1.6). DC4 and DP1l glomeruli can be distinguished by their morphology, size and position and are marked in red and blue, respectively.
(B) Color-coded image of the response to propionic acid (1% v/v). The image corresponds to the ΔF/F of the peak of the response calculated as explained in step 5.6 (see also Figure 5A). The colorscale on the left indicates the %ΔF/F value. The black circles indicate the position and size of the region of interest used to quantify the responses (see Figure 5).
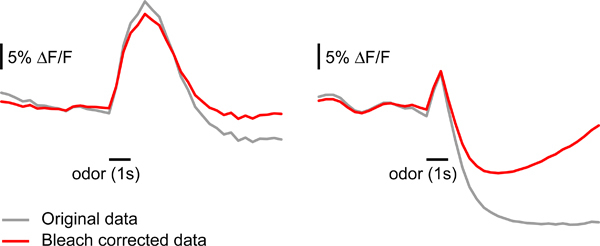 Figure 4. Correcting bleaching of the fluorescent reporter signal. Examples of the effect of bleach correction as described in step 5.3. Sample data on the left panel can be corrected without major changes in the shape of the response. Sample data on the right panel is severely affected by bleach correction, likely because the signal drops and is maintained below baseline after odor stimulus termination.
Figure 4. Correcting bleaching of the fluorescent reporter signal. Examples of the effect of bleach correction as described in step 5.3. Sample data on the left panel can be corrected without major changes in the shape of the response. Sample data on the right panel is severely affected by bleach correction, likely because the signal drops and is maintained below baseline after odor stimulus termination.
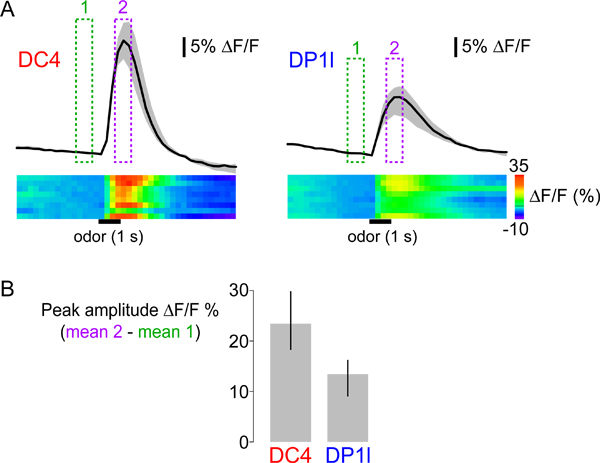 Figure 5. (A) Top: temporal dynamics of calcium responses of glomeruli DC4 (left) and DP1l (right) to propionic acid (1% v/v). The black traces show the median of the mean ΔF/F within the region of interest (see Figure 3B) across eight animals. The grey surface indicates the range between the first and third quartiles of the distribution. Presentation of median and quartile values provides more information about the variability and distribution of observed responses than mean and standard error. The green and magenta boxes indicate the frames averaged to calculate the peak responses shown in Figure 3B, and quantified in Figure 5B. Bottom: the heatmaps show the response data for all individual animals in separate rows, with ΔF/F values represented by the colorscale (far right). The horizontal black bars indicate the time and duration of the stimulus. (B) Median peak responses to propionic acid across eight animals for DC4 and DP1l glomeruli. The error bars indicate the range between first and third quartiles of the distribution.
Figure 5. (A) Top: temporal dynamics of calcium responses of glomeruli DC4 (left) and DP1l (right) to propionic acid (1% v/v). The black traces show the median of the mean ΔF/F within the region of interest (see Figure 3B) across eight animals. The grey surface indicates the range between the first and third quartiles of the distribution. Presentation of median and quartile values provides more information about the variability and distribution of observed responses than mean and standard error. The green and magenta boxes indicate the frames averaged to calculate the peak responses shown in Figure 3B, and quantified in Figure 5B. Bottom: the heatmaps show the response data for all individual animals in separate rows, with ΔF/F values represented by the colorscale (far right). The horizontal black bars indicate the time and duration of the stimulus. (B) Median peak responses to propionic acid across eight animals for DC4 and DP1l glomeruli. The error bars indicate the range between first and third quartiles of the distribution.
Discussion
The preparation and analysis methods described here can be used to analyze odor-evoked responses in any population of neurons in the antennal lobe (OSNs, LN and PNs) for which a corresponding driver transgene is available. While we describe its application using a fluorescence microscope, optical imaging of this preparation can equally be performed using confocal or two-photon microscopy22,32. Indeed, these latter instruments alleviate the light-scattering problem of wide-field microscopy, which can reduce the resolution of images, especially when large numbers of neurons express the reporter. This preparation usually permits recordings of odor-evoked responses for at least half an hour, but this time can be significantly longer or shorter depending on the number and length of measurements recorded, the exposure time used for each frame and the inter-stimulus interval. Furthermore, two-photon excitation can reduce photobleaching of the fluorescent reporter, as well as cellular phototoxicity, to permit measurement over longer time periods22,32.
Regardless of the choice of microscope, movement of the fly brain within the head capsule is the most recurrent problem when imaging in intact animals. While movement correction can correct minor issues (step 5.1), it is usually better only to record from highly stable preparations. Strategies to reduce movement artifacts include: (i) properly constraining the proboscis of the fly with the cactus spine; (ii) immobilizing the fly's legs by fixing them to the stage with eicosane (prior to step 2.6) (eicosane can be melted at ~37 °C; use a silver wire to extend and refine the tip of the soldering iron for this purpose); (iii) nicking the oesophagus (visible between the antennal nerves) with fine tweezers.
In addition to G-CaMP1.6 used here, a number of optimized versions of G-CaMP and other calcium indicators are available, which differ in their signal-to-noise ratio, baseline fluorescence and temporal dynamics12,28,33. The precise choice of sensor should take into account all of these properties, the type of neurons to be analyzed and the biological question being addressed. Current sensors report calcium changes that correlate fairly well with action potentials12,34, and their further improvement, coupled with increasing numbers of specific genetic driver lines35 will continue to enhance the potential of optical imaging of neural activity in the intact fly.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We acknowledge Silke Sachse and Beate Eisermann in the development of this methodology, Pablo Traversa for drawing the mounting block blueprints and Daniela Pelz for use of the schematic in Figure 1. We are grateful to Silke Sachse, Pavan Ramdya and Raphael Rytz for comments on the manuscript. R. Bell is supported by a Boehringer Ingelheim Foundation PhD Fellowship. Research in R. Benton’s laboratory is funded by a European Research Council Starting Independent Researcher Grant and the Swiss National Science Foundation.
References
- Galizia CG, Rossler W. Parallel olfactory systems in insects: anatomy and function. Annu. Rev. Entomol. 2010;55:399–420. doi: 10.1146/annurev-ento-112408-085442. [DOI] [PubMed] [Google Scholar]
- Vosshall LB, Stocker RF. Molecular Architecture of Smell and Taste in Drosophila. Annu. Rev. Neurosci. 2007;30:505–533. doi: 10.1146/annurev.neuro.30.051606.094306. [DOI] [PubMed] [Google Scholar]
- Wilson RI, Mainen ZF. Early events in olfactory processing. Annu. Rev. Neurosci. 2006;29:163–201. doi: 10.1146/annurev.neuro.29.051605.112950. [DOI] [PubMed] [Google Scholar]
- Masse NY, Turner GC, Jefferis GS. Olfactory information processing in Drosophila. Curr. Biol. 2009;19:700–713. doi: 10.1016/j.cub.2009.06.026. [DOI] [PubMed] [Google Scholar]
- Hansson BS, Anton S. Function and morphology of the antennal lobe: new developments. Annu. Rev. Entomol. 2000;45:203–231. doi: 10.1146/annurev.ento.45.1.203. [DOI] [PubMed] [Google Scholar]
- Galizia CG, Joerges J, Kuttner A, Faber T, Menzel R. A semi-in-vivo preparation for optical recording of the insect brain. J. Neurosci. Methods. 1997;76:61–69. doi: 10.1016/s0165-0270(97)00080-0. [DOI] [PubMed] [Google Scholar]
- Galizia CG, Menzel R, Holldobler B. Optical imaging of odor-evoked glomerular activity patterns in the antennal lobes of the ant Camponotus rufipes. Naturwissenschaften. 1999;86:533–537. doi: 10.1007/s001140050669. [DOI] [PubMed] [Google Scholar]
- Okada K, Kanzaki R, Kawachi K. High-speed voltage-sensitive dye imaging of an in vivo insect brain. Neurosci. Lett. 1996;209:197–200. doi: 10.1016/0304-3940(96)12646-x. [DOI] [PubMed] [Google Scholar]
- Carlsson MA, Knusel P, Verschure PF, Hansson BS. Spatio-temporal Ca2+ dynamics of moth olfactory projection neurones. The European journal of neuroscience. 2005;22:647–657. doi: 10.1111/j.1460-9568.2005.04239.x. [DOI] [PubMed] [Google Scholar]
- Galizia CG, Vetter RS. In: Methods in Insect Sensory Neuroscience (Frontiers in Neuroscience) Christensen TA, editor. Vol. 13. CRC Press; 2005. pp. 349–392. [Google Scholar]
- Nakai J, Ohkura M, Imoto K. A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein. Nat. Biotechnol. 2001;19:137–141. doi: 10.1038/84397. [DOI] [PubMed] [Google Scholar]
- Tian L. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat. Methods. 2009;6:875–881. doi: 10.1038/nmeth.1398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyawaki A, Griesbeck O, Heim R, Tsien RY. Dynamic and quantitative Ca2+ measurements using improved cameleons. Proc. Natl. Acad. Sci. U.S.A. 1999;96:2135–2140. doi: 10.1073/pnas.96.5.2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin JR, Rogers KL, Chagneau C, Brulet P. In vivo bioluminescence imaging of Ca signalling in the brain of Drosophila. PLoS One. 2007;2:275–275. doi: 10.1371/journal.pone.0000275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murmu MS, Stinnakre J, Martin JR. Presynaptic Ca2+ stores contribute to odor-induced responses in Drosophila olfactory receptor neurons. The Journal of Experimental Biology. 2010;213:4163–4173. doi: 10.1242/jeb.046474. [DOI] [PubMed] [Google Scholar]
- Yuste R, Miller RB, Holthoff K, Zhang S, Miesenbock G. Synapto-pHluorins: chimeras between pH-sensitive mutants of green fluorescent protein and synaptic vesicle membrane proteins as reporters of neurotransmitter release. Methods Enzymol. 2000;327:522–546. doi: 10.1016/s0076-6879(00)27300-x. [DOI] [PubMed] [Google Scholar]
- Benton R. Sensitivity and specificity in Drosophila pheromone perception. Trends Neurosci. 2007;30:512–519. doi: 10.1016/j.tins.2007.07.004. [DOI] [PubMed] [Google Scholar]
- Elliott DA, Brand AH. The GAL4 system : a versatile system for the expression of genes. Methods Mol. Biol. 2008;420:79–95. doi: 10.1007/978-1-59745-583-1_5. [DOI] [PubMed] [Google Scholar]
- Lai SL, Lee T. Genetic mosaic with dual binary transcriptional systems in Drosophila. Nat. Neurosci. 2006;9:703–709. doi: 10.1038/nn1681. [DOI] [PubMed] [Google Scholar]
- Yagi R, Mayer F, Basler K. Refined LexA transactivators and their use in combination with the Drosophila Gal4 system. Proc. Natl. Acad. Sci. U.S.A. 2010;107:16166–16171. doi: 10.1073/pnas.1005957107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potter CJ, Tasic B, Russler EV, Liang L, Luo L. The Q System: A Repressible Binary System for Transgene Expression, Lineage Tracing, and Mosaic Analysis. Cell. 2010;141:536–548. doi: 10.1016/j.cell.2010.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang JW, Wong AM, Flores J, Vosshall LB, Axel R. Two-photon calcium imaging reveals an odor-evoked map of activity in the fly brain. Cell. 2003;112:271–282. doi: 10.1016/s0092-8674(03)00004-7. [DOI] [PubMed] [Google Scholar]
- Fiala A. Genetically expressed cameleon in Drosophila melanogaster is used to visualize olfactory information in projection neurons. Curr. Biol. 2002;12:1877–1884. doi: 10.1016/s0960-9822(02)01239-3. [DOI] [PubMed] [Google Scholar]
- Ng M. Transmission of olfactory information between three populations of neurons in the antennal lobe of the fly. Neuron. 2002;36:463–474. doi: 10.1016/s0896-6273(02)00975-3. [DOI] [PubMed] [Google Scholar]
- Sachse S. Activity-dependent plasticity in an olfactory circuit. Neuron. 2007;56:838–850. doi: 10.1016/j.neuron.2007.10.035. [DOI] [PubMed] [Google Scholar]
- Pelz D, Roeske T, Syed Z, de Bruyne M, Galizia CG. The molecular receptive range of an olfactory receptor in vivo (Drosophila melanogaster Or22a) J. Neurobiol. 2006;66:1544–1563. doi: 10.1002/neu.20333. [DOI] [PubMed] [Google Scholar]
- Silbering AF, Galizia CG. Processing of odor mixtures in the Drosophila antennal lobe reveals both global inhibition and glomerulus-specific interactions. J. Neurosci. 2007;27:11966–11977. doi: 10.1523/JNEUROSCI.3099-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiff DF. In vivo performance of genetically encoded indicators of neural activity in flies. J. Neurosci. 2005;25:4766–4778. doi: 10.1523/JNEUROSCI.4900-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abuin L. Functional architecture of olfactory ionotropic glutamate receptors. Neuron. 2011;69:44–60. doi: 10.1016/j.neuron.2010.11.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benton R, Vannice KS, Gomez-Diaz C, Vosshall LB. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell. 2009;136:149–162. doi: 10.1016/j.cell.2008.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelz D. Functional characterization of Drosophila melanogaster Olfactory Receptor Neurons. Freien Universitat Berlin; 2005. [Google Scholar]
- Ai M. Acid sensing by the Drosophila olfactory system. Nature. 2010;468:691–695. doi: 10.1038/nature09537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin JR. In Vivo Brain Imaging: Fluorescence or Bioluminescence, Which to Choose? Journal of Neurogenetics. 2008. pp. 1–23. [DOI] [PubMed]
- Jayaraman V, Laurent G. Evaluating a genetically encoded optical sensor of neural activity using electrophysiology in intact adult fruit flies. Front. Neural Circuits. 2007;1:3–3. doi: 10.3389/neuro.04.003.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer BD. Tools for neuroanatomy and neurogenetics in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 2008;105:9715–9720. doi: 10.1073/pnas.0803697105. [DOI] [PMC free article] [PubMed] [Google Scholar]