

Abstract
In the present study, we examined the effect of schisandrin (SCH) of Schisandra chinensis on the amyloid-beta1–42- (Aβ 1–42-) induced memory impairment in mice and elucidated the possible antioxidative mechanism. Mice were intracerebroventricular (i.c.v.) injected with the aggregated Aβ 1–42 and then treated with SCH (4, 12, and 36 mg/kg body weight) or donepezil (DPZ), a reference drug (0.65 mg/kg) by intragastric infusion for 14 days. Noncognitive disturbances and cognitive performance were evaluated by locomotor activity test, Y-maze test, and water maze test. Antioxidative enzyme activities including superoxide dismutase (SOD) and glutathione peroxidase (GSH-px) and levels of malondialdehyde (MDA), glutathione (GSH), and oxidized glutathione (GSSG) within the cerebral cortex and hippocampus of mice were measured to elucidate the mechanism. Our results showed that SCH significantly improved Aβ 1–42-induced short-term and spatial reference memory impairments in Y-maze test and water maze test. Furthermore, in the cerebral cortex and hippocampus of mice, SOD and GSH-px activities, GSH level, and GSH/GSSG ratio were increased, and levels of MDA and GSSG were decreased by the treatment of SCH. These results suggest that SCH is a potential cognitive enhancer against Alzheimer's disease through antioxidative action.
1. Introduction
Alzheimer's disease (AD), a progressive neurodegenerative disorder with cognitive function impairment due to the presence of senile plaques and neurofibrillary tangles in the brain, is the most common form of dementia [1]. The main component of these plaques is amyloid-beta (Aβ), which plays an important role in the pathogenesis of AD [2]. Numerous studies suggest that oxidative stresses are involved in the mechanism of Aβ-induced neurotoxicity and AD pathogenesis [3, 4]. Many attempts have been made to reverse cognitive deficits by using tacrine, donepezil (DPZ), galantamine and so on; however, the use of these medicines is not always well accepted due to their severe side effects and high cost [5, 6]. Therefore, searching for safe, better tolerated, and powerful drugs is necessary.
The fruit of Schisandra chinensis (Turcz.) Baill. (Schisandrae Fructus) has been used as a tonic for kidney and brain in traditional Chinese medicine for thousands of years. It was used to improve cognitive function by recent clinical physicians, and it has been reported to reverse cycloheximide-induced amnesia in rats [7–9]. Schisandrin (SCH, molecular structure shown in Figure 1) is a main effective compound of this herb. Previous report indicated that SCH significantly reversed the scopolamine-induced impairment of the eight-arm radial maze and the passive avoidance response test in rats [10]. We observed that SCH significantly inhibited LPS-induced activation of microglia and production of NO [11]. It is widely recognized that activation of microglia by Aβ generates elevation of several markers of oxidative stress. Therefore, in the present study, we investigated the effect of SCH on the Aβ 1–42-induced memory impairment with behavioral studies. Besides, we performed an antioxidant assay in the cerebral cortex and hippocampus of mice to elucidate the mechanism of cognitive-enhancing activity.
Figure 1.
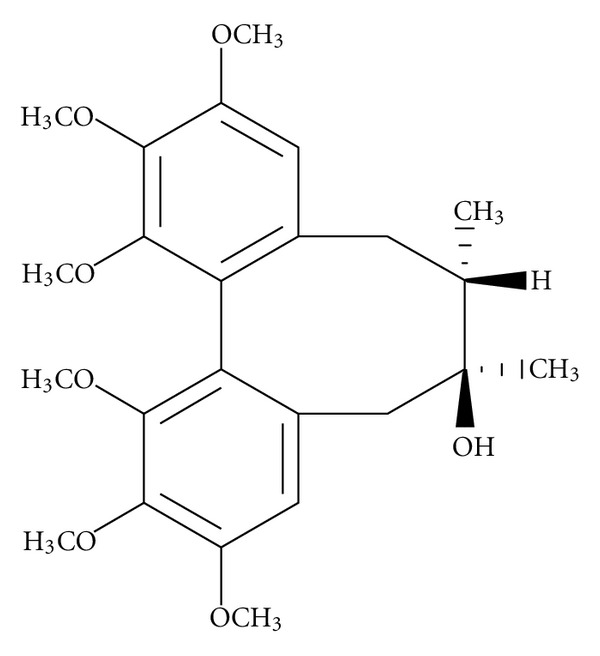
Chemical structure of SCH.
2. Materials and Methods
2.1. Plant Material
Schisandrae Fructus was collected in Liaoning province in China and identified by Professor Jun Yin (Department of Pharmacognosy, Shenyang Pharmaceutical University, Shenyang, China) according to the guidelines of the Chinese Pharmacopoeia (2010). A voucher specimen (SPU 1201) was deposited in the Herbarium of Shenyang Pharmaceutical University.
2.2. Isolation of SCH
Schisandrae Fructus (4000 g) was macerated with EtOAc (12 L × 3) at room temperature for 24 h and then concentrated under reduced pressure to yield an extract (643 g). The extract was applied to a column of silica gel column chromatography (10 × 115 cm, 200–300 mesh, 2000 g) and eluted with a gradient of petroleum ether-EtOAc (25 : 1, 11 L; 10 : 1, 25 L; 5 : 1, 29 L; 2 : 1, 7 L) to obtain 6 fractions. Fraction 5 (56 g) was further separated by silica gel column (10 × 115 cm, 200–300 mesh, 2000 g) eluted with petroleum ether-EtOAc (7 : 1, 20 L; 5 : 1, 31 L) to afford SCH (5 g) which was verified by comparison of the NMR data (Bruker ARX-300, Bruker AV-600) with those in previous reports [12].
2.3. Animals
Male Kunming mice (18–22 g) were purchased from the animal experiment centre of Shenyang Pharmaceutical University and housed 4-5 per cage, allowed access to water and food ad libitum, and maintained under a constant temperature (23 ± 1°C) and relative humidity (50 ± 10%) under a 12 h light/dark cycle (light on 07.00–19.00 h). All the studies were carried out in accordance with the principles approved by the Animal Ethical Committee of Shenyang Pharmaceutical University (no.: 2009-0004, date: September 28, 2009).
2.4. Intracerebroventricular (i.c.v.) Injection and Drug Administration
SCH (greater than 95% purity) was suspended in a 0.5% carboxyl-methylcellulose (CMC) saline solution according to the desired concentration. DPZ (greater than 98% purity, Weicai Pharmaceuticals Co., Beijing, China) was dissolved in distilled water with the concentration (20 mL/kg for dose of 0.65 mg/kg body weigh). Aβ 1–42 peptide (0.1 mg) was dissolved in DMSO (1.2 μL), then the solution was diluted with sterile physiological saline (161 μL) in the tube, which was then sealed and incubated for 120 h at 37°C to cause the peptide to aggregate.
Mice were randomly assigned to six groups of 8–10 individuals each: sham, control, positive, and SCH (4, 12, 36 mg/kg). All the mice were anesthetized with 3.5% chloral hydrate (0.1 mL/10 g). Then, mice in the control, positive, and SCH (4, 12, 36 mg/kg) groups were injected with aggregated Aβ 1–42 peptide (410 pmol/3 μL) into the right lateral ventricle within 3 s by means of a stereotaxic apparatus (AP, −0.2 mm, ML, ±1.0 mm, DV, −2.5 mm) [13]. The needle was removed with 1 min delay to allow diffusion. Mice in the sham-operated group were injected in an identical manner with the same amount of 0.7% DMSO-physiological saline (3 μL). After surgery, animals were returned to their cages.
From the next day, mice in the SCH (4, 12, 36 mg/kg) groups were administered SCH (20 mL/kg for doses of 4, 12, and 36 mg/kg body weight) daily for 14 consecutive days by intragastric infusion (i.g.). Mice in positive group were administered with DPZ (0.65 mg/kg, i.g.), which is clinically used for symptomatic treatment of patients with AD [14, 15]. Mice in sham-operated and control groups were treated with distilled water (20 mL/kg, i.g.) in the same period.
2.5. Locomotor Activity Test
7 days after the surgical operation, locomotor activity of each animal was observed in a locomotor cage (50 × 50 × 40 cm) using video-recorded analytical system (Shanghai Jiliang Software Technology Co.Ltd., Shanghai, China). The total distance of movements was evaluated over a 10 min period.
2.6. Y-Maze Task
The Y-maze task was carried out as previously described on day 8 after surgical operation [16]. The Y-maze is a three-arm maze with all arms at equal angles (labelled A, B, and C). Each arm was 40 cm long, 13 cm high, 5 cm wide at the bottom, 10 cm wide at the top. Each mouse was placed on one end of an arm and allowed to move freely through the maze during an 8 min session. The sequence and number of arm entries were recorded manually. A spontaneous alternation behavior, which is regarded as a measure of working memory, was defined as entries into all three arms on consecutive occasions (i.e., ACB, CBC, BCA, CBA). The percentage alternation was calculated as the ratio of actual to possible alternations (defined as the total number of arm entries minus two), multiplied by 100. The number of arm entries was used as an indicator of locomotor activity.
2.7. Water Maze Task
The water maze task was carried out as previously described on day 9 after surgical operation [17–20]. The water maze apparatus consisted of a brown organic glass water tank (63 cm long, 36 cm wide, and 20 cm high), which is crisscross divided into 5 conjoint parts by brown plastic clapboards to form a circuitous waterway containing 10 cm depth of water at 23 ± 1°C. A hidden platform was set inside one corner of the tank, which was called safety area. Mice were placed in the diagonal area of the safety area, allowed to swim for a maximum of 60 s. In early experiments, they can be induced to safety area. When a mouse arrived at the platform, it can get a rest. Such training trials were repeated for 5 days (10 trials/day). The time required to reach the safety area (the escape latency time) was recorded.
2.8. Antioxidant Assay
On day 14 after surgical operation, mice were anesthetized by pentobarbital (50 mg/kg) and then decapitated. The skulls were opened, and the brains were quickly harvested and placed on ice. The cerebral cortexes were excised, then the hippocampus below them were also carefully dissected, and the tissues were stored at −80°C until analysis. Samples were homogenized in a glass Teflon homogenizer containing 9-fold volumes of precooling physiological saline and then centrifuged at 3000 × g for 15 min at 4°C. The supernatant obtained was used to measure antioxidative enzyme activities including superoxide dismutase (SOD) and glutathione peroxidase (GSH-px), and contents of protein, malondialdehyde (MDA), glutathione (GSH), and oxidized glutathione (GGSG) by means of the assay kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
2.9. Statistical Analysis
All statistical analyses were performed using the SPSS software, version 13.0. Data of escape latency values in water maze test were analyzed by a two-way analysis of variance (ANOVA) with the day as one variable and the treatment as a second, followed by the Bonferroni post hoc test. The other data were analyzed using one-way ANOVA followed by LSD test in the condition of variance homogeneity or Tamhane test in variance heterogeneity to detect intergroup differences. Data were expressed as mean ± S.E.M. Statistical significance was set at P < 0.05.
3. Results
3.1. Locomotor Activity
Locomotor activity was evaluated to exclude the possibility that the improved learning and memory effects were caused by the increased locomotor activity of the animals. As summarized in Table 1, there was no significant difference in the locomotor activity among the sham, control, DPZ (0.65 mg/kg), and SCH (4, 12, 36 mg/kg) groups (F (5, 51) = 2.036, P > 0.05).
Table 1.
Effects of SCH on locomotor activity of Aβ 1–42-induced memory impairment in mice (n = 9-10).
| Group | Dose (mg/kg) | Locomotor activity (cm/10 min, mean ± S.E.M.) |
|---|---|---|
| Sham | — | 4923.37 ± 399.171 |
| Control | — | 5205.88 ± 213.25 |
| DPZ | 0.65 | 5293.63 ± 457.79 |
| SCH | 4 | 4685.69 ± 406.90 |
| SCH | 12 | 5035.32 ± 757.24 |
| SCH | 36 | 5082.48 ± 281.53 |
Mice in sham-operated and control groups were treated with distilled water orally (20 mL/kg). Values were mean ± S.E.M. and analyzed by one-way ANOVA followed by LSD test for multiple comparisons.
3.2. Y-Maze Task
In the Y-maze test, the spontaneous alternation behavior in the control group was significantly less than that in the sham group (22.30% decrease in alternation behavior, P < 0.001). However, this decreased spontaneous alternation behaviors induced by Aβ 1–42 that were significantly improved by DPZ (0.65 mg/kg, P = 0.037, Figure 2(a)) and SCH (12, 36 mg/kg, P = 0.007 and P = 0.011, resp., Figure 2(a)). In the SCH (4, 12, 36 mg/kg) and DPZ (0.65 mg/kg) groups, the spontaneous alternation behaviors were increased to 82.26, 94.12, 92.38, and 89.82% of the sham group, respectively. The total number of arm entries was not significantly different among all groups (F (5, 57) = 0.421, P > 0.05, Figure 2(b)), suggesting that the changes in spontaneous alternation behavior were not due to locomotor deficits.
Figure 2.
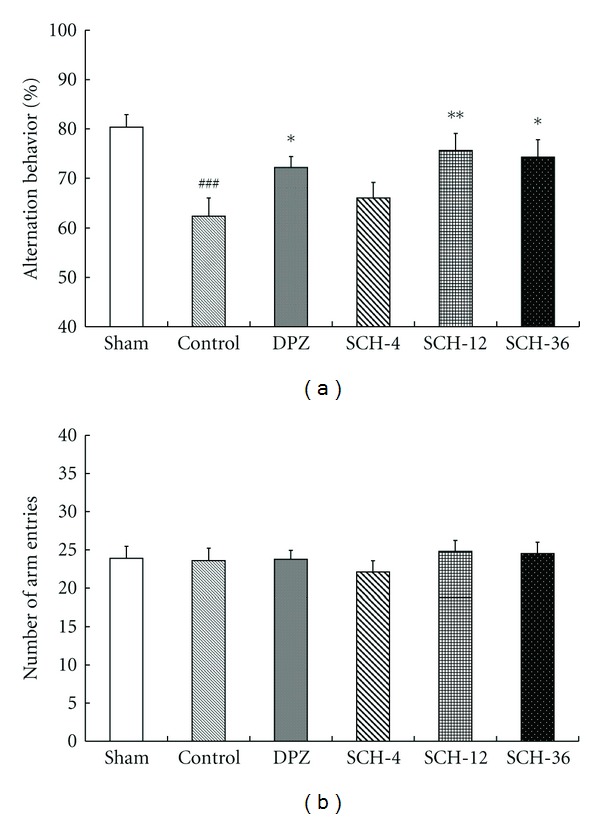
Effects of SCH against Aβ 1–42-induced memory impairment in Y-maze test. Spontaneous alternation behavior (a) and total number of arm entries (b) were measured during an 8-min session. Sham-operated and control groups: mice were given distilled water orally (20 mL/kg). Positive control: mice were given DPZ 0.65 mg/kg, i.g. SCH-4, SCH-12, and SCH-36: mice were given 4, 12, and 36 mg/kg of SCH, i.g., respectively. Data in the bar graphs were represented as mean ± S.E.M. and analyzed by one-way ANOVA followed by LSD test for multiple comparisons (n = 8–10). ### P < 0.001 compared with the sham-operated group; *P < 0.05, **P < 0.01 compared with the control group.
3.3. Water Maze Task
In water maze task, the changes in escape latency to find the safety area after training trials in each group were shown in Figure 3. In all groups, escape latencies decreased with training (F (4, 288) = 160.97, P < 0.001) and escape latencies were significantly different among these groups (F (5, 288) = 4.237, P = 0.001). On day 1, the escape latencies of the sham-operated and control group were 41.71 s and 42.87 s. On day 2, the escape latencies of the sham, control, DPZ (0.65 mg/kg), and SCH (4, 12, 36 mg/kg) groups were 29.67 s, 35.73 s, 30.54 s, 34.16 s, 32.03 s, and 30.56 s, respectively. On day 3, they were 22.93 s, 27.71 s, 22.04 s, 25.88 s, 24.82 s, and 23.02 s, respectively. The sham-operated group rapidly lessened the escape latency time to find the location of platform from day 2 and reached stable escape latency time from day 3 to day 5. In contrast, the escape latencies of the control group were significantly delayed compared to that of the sham group on day 2 and day 3, DPZ (0.65 mg/kg) and SCH (36 mg/kg) significantly reduced the escape latencies prolonged by Aβ 1–42 on day 2 and day 3.
Figure 3.
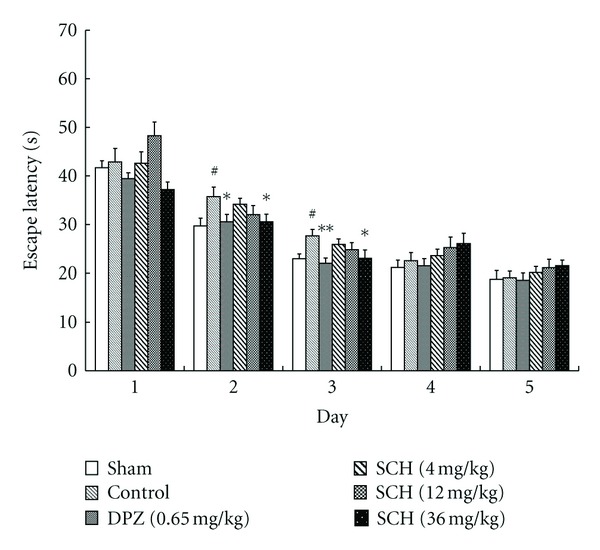
Effects of SCH against Aβ 1–42-induced memory impairment on average escape latency time of the water maze test. Sham-operated and control groups: mice were given distilled water orally (20 mL/kg). Positive control: mice were given DPZ 0.65 mg/kg, i.g. SCH-4, SCH-12, and SCH-36: mice were given 4, 12, and 36 mg/kg of SCH, i.g., respectively. Values indicated mean ± S.E.M. and were analyzed by a two-way analysis of variance (ANOVA) followed by the Bonferroni post hoc test (n = 8–10). # P < 0.05 compared with the sham-operated group; *P < 0.05, **P < 0.01 compared with the control group.
3.4. Antioxidant Assay
In order to elucidate the biochemical mechanism of antidementia effects of SCH in brain tissue, the antioxidative enzyme activities including SOD and GSH-px and levels of MDA, GSH, and GSSG were determined. When compared with the sham group, the Aβ 1–42 injection generated a dramatic decrease in SOD (−34.33%) and GSH-px (−31.22%) activities as well as a significant increase in MDA (49.49%) production in the cerebral cortex or hippocampus of mice, while DPZ (0.65 mg/kg) preserved the activities of SOD (88.09%) and GSH-px (89.72%) caused by Aβ 1–42, and SCH (36 mg/kg) preserved 82.18% SOD and 87.02% GSH-px activities (Table 2), respectively. The increase in MDA production was also significantly attenuated by the treatment with DPZ (0.65 mg/kg) and SCH (36 mg/kg), and the increase was 9.09% and 14.29%, respectively, by comparison with the sham group.
Table 2.
Effects of SCH on SOD, GSH-px activities; MDA, GSH, GSSG levels and GSH/GSSG ratio in the cerebral cortex or hippocampus of Aβ 1–42-induced memory impairment in mice.
| Group | Dose (mg/kg) | SOD | MDA | GSH-px | GSH | GSSG | GSH/GSSG |
|---|---|---|---|---|---|---|---|
| (U/mg protein) | (nmol/mg protein) | (U/mg protein) | (μmol/L) | (μmol/L) | |||
| Cerebral cortex | Cerebral cortex | Hippocampus | Hippocampus | Hippocampus | Hippocampus | ||
| Sham | — | 9.99 ± 0.71 | 6.93 ± 0.77 | 20.34 ± 1.37 | 260.63 ± 19.29 | 94.88 ± 5.37 | 2.79 ± 0.23 |
| Control | — | 6.56 ± 0.55### | 10.36 ± 0.8## | 13.99 ± 1.46### | 161.04 ± 9.45# | 122.36 ± 11.55 | 1.38 ± 0.14### |
| DPZ | 0.65 | 8.80 ± 0.32∗∗ | 7.56 ± 0.62∗ | 18.25 ± 0.95∗ | 220.63 ± 16.74 | 95.76 ± 2.40 | 2.31 ± 0.18∗∗∗ |
| SCH | 4 | 6.93 ± 0.36 | 10.81 ± 0.83 | 13.94 ± 1.36 | 136.25 ± 14.14 | 114.08 ± 6.84 | 1.21 ± 0.14 |
| SCH | 12 | 7.65 ± 0.47 | 9.66 ± 1.18 | 13.61 ± 1.00 | 167.3 ± 11.557 | 104.54 ± 4.08 | 1.64 ± 0.16 |
| SCH | 36 | 8.21 ± 0.48∗ | 7.92 ± 0.57∗ | 17.70 ± 0.49∗ | 202.71 ± 10.79 | 98.52 ± 1.35 | 2.06 ± 0.12∗∗ |
Mice in sham-operated and control groups were treated with distilled water orally (20 mL/kg). Values represented mean ± S.E.M. and analyzed by one-way ANOVA followed by LSD test or Tamhane test for multiple comparisons (n = 8–10). # P < 0.05, ## P < 0.01, ### P < 0.001 compared with the sham-operated group; ∗ P < 0.05, ∗∗ P < 0.01, ∗∗∗ P < 0.01 compared with the control group.
In the sham group, the levels of GSH and GSSG and GSH/GSSG ratio were 260.63 ± 54.56 μmol/L, 94.88 ± 15.19 μmol/L, and 2.79 ± 0.65, respectively. In the Aβ 1–42-injected control group, the GSH level (161.04 ± 26.74 μmol/L) and the GSH/GSSG ratio (1.38 ± 0.40) were significantly decreased with a concomitant increase of GSSG level (122.36 ± 32.67 μmol/L) which was not significant changed compared with the sham-operated group. In DPZ (0.65 mg/kg) and SCH (4, 12, 36 mg/kg) groups, the levels of GSH and GSSG (μmol/L) and GSH/GSSG ratio were 220.63 ± 47.36 μmol/L, 136.25 ± 40.00 μmol/L, 167.3 ± 32.67 μmol/L, and 202.71 ± 30.52 μmol/L; 95.76 ± 6.79 μmol/L, 114.08 ± 19.36 μmol/L, 104.54 ± 11.55 μmol/L, and 98.52 ± 3.81 μmol/L; 2.31 ± 0.5, 1.21 ± 0.38, 1.64 ± 0.46, and 2.06 ± 0.34, respectively.
4. Discussion
At present, there is no animal model which could mimic all the cognitive, behavioral, biochemical, and histopathological abnormalities observed in patients with AD. However, partial reproductions of AD neuropathology and cognitive deficits have been achieved by pharmacological and genetic approaches [21]. Iβ hinders memory and cognitive function in both animals and humans, specially short-term and spatial reference memory. As such, continuous infusion or acute injection of Aβ into the brain has been used to an experimental model for AD [22–24]. The anti-inflammatory effect of SCH was observed in vitro in our previous work [11], but its role in the dementia model is unclear. For these reasons, we used Aβ 1–42-injection mice model in this research to clarify the role of SCH.
In this research, a single i.c.v. injection of Aβ 1–42-induced significant impairment of working memory in the Y-maze and a deficit of short-term spatial learning and memory in the water maze on day 2-3 (Figures 2(a) and 3), which partially simulated the earlier stage characteristics of learning and memory obstacles of AD.
The blood-brain barrier (BBB) is a major obstacle to the effective delivery of drugs to the brain. It is generally accepted that only small molecules with low molecular mass (<450 Da) and high lipid solubility permeate the healthy BBB by a passive transcellular process [25]. The molecular mass of SCH is 432 Da, and it is liposoluble and can distribute widely in various areas of the rat's brain [26]. Schisandrae Fructus has been used as a tonic in traditional Chinese medicine and a health food product in China for thousands of years and currently is being used as a dietary supplement in the United States. To determine whether SCH can be a potential candidate for preventing and/or delaying progression of the learning and memory impairments of AD, it was tested in vivo experiments by oral administration due to its liposoluble property. The administration doses of SCH were chosen according to the preliminary study (data were not shown here), which were much lower than the most tolerant dosage 1000 mg/kg measured in the acute toxicity experiment [27]. DPZ which has been approved for treatment of AD was used to evaluate whether the model was successfully established and the treatment effect of SCH on memory impairment [14, 15]. Dose of DPZ was calculated based on the clinical dose (5 mg per day) [28] according to the formula recorded in the pharmacology experimental methodology edited by Xu et al. [29].
Our results demonstrated total distance of movements of locomotor activity test, and the total number of arm entries in Y-maze test was not significantly different among all groups (Table 1, Figure 2(b)), suggesting that there were no potential drug effects on nonspecific processes, such as arousal, attention, or sensory motor functions [30].
Previous report has demonstrated that SCH significantly reversed the scopolamine-induced impairment in rats using the passive avoidance response test which evaluates long-term memory [10]. In the present study, we measured spontaneous alternation behaviors in the Y-maze test to appraise working memory, which is one of the short-term memories impaired at an early stage of AD. The spontaneous alternation behaviors were significantly improved by SCH (12, 36 mg/kg), and the effects of them on improving working memory were better than DPZ (0.65 mg/kg) (Figure 2(a)).
Spatial memory was assessed using water maze task. In all groups, training had a significant effect on spatial learning performance. This effect was more evident than the treatment effect. These results showed that, in spite of the Aβ 1–42-induced memory impairment in mice, all the animals were able to learn to find the platform when guided by extra-maze cues on day 4 and 5. Therefore, escape latencies were not significantly different on day 4 and 5. SCH (36 mg/kg) could significantly improve spatial learning and memory impairment in mice on day 2 and day 3 (Figure 3). As a single i.c.v. injection of Aβ 1–42 only induced short-term impairment of spatial learning and memory on day 2-3, other long-term impairment models of AD should be investigated to find if they can be rescued by SCH in further study.
The etiology of AD has not been revealed clearly. Nevertheless, lots of clinical studies have reported that oxidative stress is involved in cognitive and functional decline in AD [31–33]. Lignans of Schisandrae Fructus have antioxidative effects for example, eliminating reactive oxygen species by increasing SOD and catalase activities and inhibiting lipid peroxidation in cell membranes [34–37]. So we performed an antioxidant assay in the cerebral cortex and hippocampus of mice to elucidate the mechanism of cognitive-enhancing activity.
Lipid peroxidation is one of the main outcomes of free radical-mediated tissue injury that directly damages membranes and, in turn, generates a number of secondary products, such as MDA [38]. In the present research, it was showed that Aβ 1–42-peptide generated significant increased MDA in mouse cerebral cortex and SCH (36 mg/kg) could significantly decrease the lipid peroxidation caused by Aβ 1–42 (Table 2). SOD and GSH-px are important antioxidant enzymes involved in cellular protection against damage caused by oxygen-derived free radicals, by means of removing harmful peroxide metabolites and blocking lipid peroxidation chain reaction. SCH (36 mg/kg) significantly increased the activities of SOD and GSH-px enzyme caused by Aβ 1–42 (Table 2).
GSH, which is a water-soluble endogenous antioxidant in cells, can transform to an oxidation form of GSSG by catalysis of GSH-px and removing oxygen free radicals. So it plays important roles in protection from oxidative stress in the brain. Our results showed that SCH prevented the reduction of GSH and reduced GSSG level, but this improvement was not notable, while GSH/GSSG ratios were significantly increased in SCH (36 mg/kg) group by comparison with the control group (Table 2). The reason may be that there are other mechanisms associated with the inhibiting oxidative stress and the subsequent amelioration in cognitive function in SCH-treated mice. Previous report has suggested that SCH reverses scopolamine-induced memory impairment by enhancing cholinergic function [10].
It was reported that absolute bioavailability of SCH was 14.8% after i.v. administration (1 mg/kg) and i.g. administration (10 mg/kg) in rat. Orally administered SCH can be quickly absorbed into the blood and may then express its pharmacological effects within 1 h [39]. It was shown that the major metabolite of SCH, 7, 8-dihydroxy-schisandrin had the almost same sedative effect with SCH [40]. Therefore, in future, it is necessary to investigate whether the metabolites of SCH are active compounds to improve memory impairment.
In conclusion, the present study revealed that SCH could ameliorate Aβ 1–42-induced memory impairment in mice at least in part by enhancing the activity of the antioxidative defense system and free radical-scavenging activity. Further study is required to figure out the detailed mechanism of SCH in the treatment of AD, such as measurement of neurodegeneration, oxidative stress in Aβ 1–42-induced mice, and other models in brain tissue sections with immunohistochemistry.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81102507). The authors thank Professor Libo Zou for useful technical and discussion assistance.
Abbreviations
- Aβ:
Amyloid-beta
- AD:
Alzheimer's disease
- BBB:
Blood-brain barrier
- DPZ:
Donepezil
- GGSG:
Oxidized glutathione
- GSH:
Glutathione
- GSH-px:
Glutathione peroxidase
- i.c.v.:
Intracerebroventricular
- i.g.:
Intragastric
- MDA:
Malondialdehyde
- SCH:
Schisandrin
- SOD:
Superoxide dismutase.
References
- 1.Francis PT, Palmer AM, Snape M, Wilcock GK. The cholinergic hypothesis of Alzheimer’s disease: a review of progress. Journal of Neurology Neurosurgery and Psychiatry. 1999;66(2):137–147. doi: 10.1136/jnnp.66.2.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Selkoe DJ. The molecular pathology of Alzheimer’s disease. Neuron. 1991;6(4):487–498. doi: 10.1016/0896-6273(91)90052-2. [DOI] [PubMed] [Google Scholar]
- 3.Schubert D, Behl C, Lesley R, et al. Amyloid peptides are toxic via a common oxidative mechanism. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(6):1989–1993. doi: 10.1073/pnas.92.6.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yankner BA. Mechanisms of neuronal degeneration in Alzheimer’s disease. Neuron. 1996;16(5):921–932. doi: 10.1016/s0896-6273(00)80115-4. [DOI] [PubMed] [Google Scholar]
- 5.Inglis F. The tolerability and safety of cholinesterase inhibitors in the treatment of dementia. International Journal of Clinical Practice, Supplement. 2002;(127):45–63. [PubMed] [Google Scholar]
- 6.Doody RS. Clinical profile of donepezil in the treatment of Alzheimer’s disease. Gerontology. 1999;45(1):23–32. doi: 10.1159/000052761. [DOI] [PubMed] [Google Scholar]
- 7.Chiu L, Yang SH. Fructus Schisandrae: possesses anti-cancer and anti-aging activities. Gansu Medical Journal. 1992;11:p. 288. [Google Scholar]
- 8.Hancke JL, Burgos RA, Ahumada F. Schisandra chinensis (Turcz.) Baill. Fitoterapia. 1999;70(5):451–471. [Google Scholar]
- 9.Hsieh MT, Wu CR, Wang WH, Lin LW. The ameliorating effect of the water layer of Fructus schisandrae on cycloheximide-induced amnesia in rats: interaction with drugs acting at neurotransmitter receptors. Pharmacological Research. 2001;43(1):17–22. doi: 10.1006/phrs.2000.0756. [DOI] [PubMed] [Google Scholar]
- 10.Egashira N, Kurauchi K, Iwasaki K, et al. Schizandrin reverses memory impairment in rats. Phytotherapy Research. 2008;22(1):49–52. doi: 10.1002/ptr.2258. [DOI] [PubMed] [Google Scholar]
- 11.Zhang N, Hu D, Yang JY, Cai XD, Yin J, Wu CF. Studies on chemical constituents of leaf of Schisandra chinensis and inhibitory effect of schizandrin on activation of microglia induced by LPS. Chinese Journal of Medicinal Chemistry. 2010;20(2):110–115. [Google Scholar]
- 12.Ikeya Y, Taguchi H, Sasaki H, Nakajima K, Yoshioka I. 13C NMR magnetic resonance spectroscopy of dibenzocyclooctadiene lignans. Chemical & Pharmaceutical Bulletin. 1980;27:1383–1390. [Google Scholar]
- 13.Kim DH, Jung WY, Park SJ, et al. Anti-amnesic effect of ESP-102 on Aβ 1−42-induced memory impairment in mice. Pharmacology Biochemistry and Behavior. 2010;97(2):239–248. doi: 10.1016/j.pbb.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 14.Giacobini E. Cholinesterase inhibitors stabilize Alzheimer disease. Neurochemical Research. 2000;25(9-10):1185–1190. doi: 10.1023/a:1007679709322. [DOI] [PubMed] [Google Scholar]
- 15.Nordberg A, Svensson AL. Cholinesterase inhibitors in the treatment of Alzheimer’s disease. A comparison of tolerability and pharmacology. Drug Safety. 1998;19(6):465–480. doi: 10.2165/00002018-199819060-00004. [DOI] [PubMed] [Google Scholar]
- 16.Maurice T, Hiramatsu M, Kameyama T, Hasegawa T, Nabeshima T. Cholecystokinin-related peptides, after systemic or central administration, prevent carbon monoxide-induced amnesia in mice. Journal of Pharmacology and Experimental Therapeutics. 1994;269(2):665–673. [PubMed] [Google Scholar]
- 17.Ji XF, Liu XX, Wu Z, Yang BZ, Wang LH, Zou LB. Improving effects of extract of the husk of Xanthoceras sorbifolia Bunge on amyloid beta protein-induced learning and memory impairmant in animal models. ShenYang Pharmaceutical University. 2007;24:232–237. [Google Scholar]
- 18.Chi TY, Wang LH, Ji XF, et al. Protective effects of Xanthoceraside on learning and memory impairmant induced by Aβ 1−42 in mice. Journal of China Medical University. 2009;38:734–736. [Google Scholar]
- 19.Dong AM. Anti-vascular dementia effects of Puerariae Isoflavone. Shenyang Pharmaceutical University. 2003:p. 18. [Google Scholar]
- 20.Qu S. Effects of Xanthoceraside on learning and memory impairment in mice induced by Aβ 25−35 . Shenyang Pharmaceutical University. 2007:p. 15. [Google Scholar]
- 21.Yamada K, Nabeshima T. Animal models of Alzheimer’s disease and evaluation of anti-dementia drugs. Pharmacology and Therapeutics. 2000;88(2):93–113. doi: 10.1016/s0163-7258(00)00081-4. [DOI] [PubMed] [Google Scholar]
- 22.Harkany T, Mulder J, Sasvári M, et al. N-methyl-D-aspartate receptor antagonist MK-801 and radical scavengers protect cholinergic nucleus basalis neurons against β-amyloid neurotoxicity. Neurobiology of Disease. 1999;6(2):109–121. doi: 10.1006/nbdi.1998.0230. [DOI] [PubMed] [Google Scholar]
- 23.Jhoo JH, Kim HC, Nabeshima T, et al. β-Amyloid (1−42)-induced learning and memory deficits in mice: involvement of oxidative burdens in the hippocampus and cerebral cortex. Behavioural Brain Research. 2004;155(2):185–196. doi: 10.1016/j.bbr.2004.04.012. [DOI] [PubMed] [Google Scholar]
- 24.Pepeu G, Giovannelli L, Casamenti F, Scali C, Bartolini L. Amyloid β-peptides injection into the cholinergic nuclei: morphological, neurochemical and behavioral effects. Progress in Brain Research. 1996;109:273–282. doi: 10.1016/s0079-6123(08)62111-1. [DOI] [PubMed] [Google Scholar]
- 25.Hitchcock SA. Blood-brain barrier permeability considerations for CNS-targeted compound library design. Current Opinion in Chemical Biology. 2008;12(3):318–323. doi: 10.1016/j.cbpa.2008.03.019. [DOI] [PubMed] [Google Scholar]
- 26.Niu XY, Bian ZJ, Ren ZH. The metabolic fate of schizandrol A and its distribution in the rat brain determined by thin layer chromatography. Acta Pharmaceutica Sinica. 1983;18(7):491–495. [PubMed] [Google Scholar]
- 27.Bao TT, Tu GF, Liu GT, Sun RH, Song ZY. A comparison of the pharmacological actions of seven constituents isolated from fructus schizadrae (author’s transl) Acta Pharmaceutica Sinica. 1979;14(1):1–7. [PubMed] [Google Scholar]
- 28.Dooley M, Lamb HM. Donepezil. A review of its use in Alzheimer’s disease. Drugs and Aging. 2000;16(3):199–226. doi: 10.2165/00002512-200016030-00005. [DOI] [PubMed] [Google Scholar]
- 29.Xu SY, Bian RL, Chen X, et al. Pharmacology Experimental Methodology. 3rd edition. Beijing, China: 2005. [Google Scholar]
- 30.Hunter B, Zornetzer SF, Jarvik ME, McGaugh JL. Modulation of learning and memory: effects of drugs influencing neurotransmitters. In: Iversen LL, Iversen SD, Snyder SH, et al., editors. Handbook of Psychopharmacology. Vol. 19. New York, NY, USA: 1988. pp. 531–577. [Google Scholar]
- 31.Squitti R, Rossini PM, Cassetta E, et al. D-penicillamine reduces serum oxidative stress in Alzheimer’s disease patients. European Journal of Clinical Investigation. 2002;32(1):51–59. doi: 10.1046/j.1365-2362.2002.00933.x. [DOI] [PubMed] [Google Scholar]
- 32.Reddy PH. Amyloid precursor protein-mediated free radicals and oxidative damage: implications for the development and progression of Alzheimer’s disease. Journal of Neurochemistry. 2006;96(1):1–13. doi: 10.1111/j.1471-4159.2005.03530.x. [DOI] [PubMed] [Google Scholar]
- 33.Dumont M, Beal MF. Neuroprotective strategies involving ROS in Alzheimer disease. Free Radical Biology and Medicine. 2010;51:1014–1026. doi: 10.1016/j.freeradbiomed.2010.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li XJ, Zhao BL, Liu GT, Xin WJ. Scavenging effects on active oxygen radicals by schizandrins with different structures and configurations. Free Radical Biology and Medicine. 1990;9(2):99–104. doi: 10.1016/0891-5849(90)90111-u. [DOI] [PubMed] [Google Scholar]
- 35.Liu W, Yu R, Wu JH, Luo HM. γ-schisandrin inhibits production of amyloid β-protein 42 in M146L cells. Yaoxue Xuebao. 2006;41(12):1136–1140. [PubMed] [Google Scholar]
- 36.Opletal L, Sovová H, Bártlová M. Dibenzo[a, c]cyclooctadiene lignans of the genus Schisandra: importance, isolation and determination. Journal of Chromatography B. 2004;812(1-2):357–371. doi: 10.1016/j.jchromb.2004.07.040. [DOI] [PubMed] [Google Scholar]
- 37.Wang JP, Raung SL, Hsu MF, Chen CC. Inhibition by gomisin C (a lignan from Schizandra chinensis) of the respiratory burst of rat neutrophils. British Journal of Pharmacology. 1994;113(3):945–953. doi: 10.1111/j.1476-5381.1994.tb17084.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Slater TF. Overview of methods used for detecting lipid peroxidation. Methods in Enzymology. 1984;105:283–293. doi: 10.1016/s0076-6879(84)05036-9. [DOI] [PubMed] [Google Scholar]
- 39.Xu MJ, Wang GJ, Xie HT, et al. Determination of schizandrin in rat plasma by high-performance liquid chromatography-mass spectrometry and its application in rat pharmacokinetic studies. Journal of Chromatography B. 2005;828(1-2):55–61. doi: 10.1016/j.jchromb.2005.09.029. [DOI] [PubMed] [Google Scholar]
- 40.Cui YY, Wang MZ. Metabolic transformation of schizandrin. Acta Pharmaceutica Sinica. 1992;27(1):57–63. [PubMed] [Google Scholar]