

Throughout the last century, most neurophysiologists believed synaptic transmission in the brain was mediated solely by neurons. Glial cells, the most numerous cells in the brain, were primarily seen as providing support but not as actively participating in rapid neural signaling. Several recent findings indicate that this restrictive view of neural circuitry may be replaced in this century by a more inclusive one in which glia play an active role in synaptic signaling. Two very recent papers, one in Nature by Bergles et al. (1) and one in this issue of PNAS by Parpura and Hayden (2), show that two different types of glial cells are involved directly in rapid glutamate-mediated neural signaling. One glial cell type, oligodendrocyte precursor cells (OPCs), was shown to receive direct synaptic input from neurons in the hippocampus (1), and another glial cell type, astrocytes, was shown to release glutamate rapidly in response to physiological increases in intracellular calcium concentration (2). These two papers—together with earlier research showing that glia express neurotransmitter receptors, release neurotransmitter, and propagate intracellular calcium waves (see refs. 3 and 4 for recent reviews)—indicate that synaptic information flows through the nervous system via glial/neuronal networks.
The notion that glia solely provide support functions in brain can be traced to the naming of these cells in the 19th century: “glia“ is Greek for glue. In fact, there are several different glial cell types, and each provides different support and trophic functions. For example, astrocytes buffer extracellular potassium and neurotransmitter released by neurons, and oligodendrocytes in the central nervous system and Schwann cells in the periphery myelinate axons. In addition, precursors for these glial cells, including OPCs, are present in the adult nervous system (5, 6).
Perhaps because, unlike neurons, glia lack axons and do not generate action potentials, they have been largely ignored by neurophysiologists studying synaptic transmission. The first clues that glial cells may participate directly in synaptic transmission came from observations made in the 80s and 90s that glia express neurotransmitter receptors. In fact, glia express a surprisingly large number of different transmitter receptors including receptors for glutamate (7, 8), γ-aminobutyric acid (9, 10), serotonin, acetyl choline, dopamine, and neurepinephrine (11). At the time they were described, the neurotransmitter receptors on glial cells were hypothesized to link the activity of neural circuits with glial support functions and glial proliferation (12).
Although glia have been known to express receptors that would allow them to receive synaptic input directly, two important pieces of information were missing before glia could be considered seriously in synaptic transmission. First, could glial cells receive direct synaptic input, and second, could glia release neurotransmitter in response to physiological stimuli and stimulate neighboring glia or neurons? The first classical synaptic connection between neurons and glia was demonstrated this year by Bergles, Roberts, Somogyi, and Jahr (1). In their paper, OPCs in the hippocampus were shown to receive a direct synaptic connection from CA3 pyramidal cells (Fig. 1). Both electrophysiological and ultrastructural evidence shows that this synapse operates in the same manner as traditional neuron-to-neuron synapses. Transmitter at these synapses is released by vesicles, and the glutamate receptors activated are calcium-permeable α-amino-3-hydroxy5-methyl-4-isoxazolepropionic acid receptors. This neuro-glia synapse is, to our knowledge, the first to be both physiologically and structurally defined in the central nervous system, and it will certainly stimulate a search for other varieties of neuro-glia synapses. In addition, the identification of these synapses begs the question of what biological role they play. Are these synapses “dead ends” or signal terminators; do they serve as “synaptic sinks;” or do the OPCs release neurotransmitter on synaptic activation? Although the question remains as to their function, the presence of neuro-glia synapses dramatically illustrates that glia are connected directly to synaptic networks.
Figure 1.
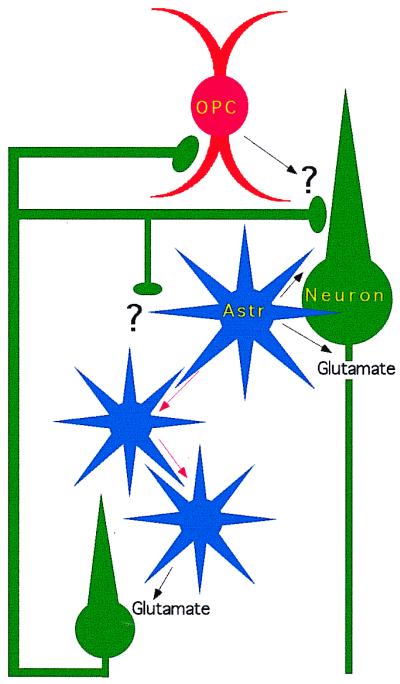
Synaptic networks of neurons and glia. OPCs in hippocampus receive direct synaptic connections from CA3 pyramidal neurons. Astrocytes (Astr) propagate intracellular calcium waves (red arrows) and release glutamate, which activates neurotransmitter receptors on neurons.
Because glia do not generate action potentials and do not have axons, they were initially thought incapable of propagating neural impulses. Calcium-imaging studies, however, have shown that astrocytes are capable of propagating cellular signals as waves of intracellular calcium changes that spread from astrocyte to astrocyte (13). Furthermore, calcium waves through astrocyte networks have been shown to influence the activity of adjacent neurons and modulate synaptic transmission (14–16). After an astrocyte calcium wave passes a neuron, the neuron responds with its own elevation in intracellular calcium, and this neuronal response is blocked by blocking α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors. This result suggests that, similar to presynaptic buttons of neurons, glia release glutamate when intracellular calcium concentrations are elevated. The exact mechanism of this transmitter release from astrocytes is still unclear, although it may be vesicular as opposed to the reversal of neurotransmitter uptake, because it is not affected by blockers of glutamate uptake and is reduced by toxins that interfere with vesicle trafficking (17).
Typically, calcium waves in astrocytes are initiated experimentally by mechanical stimulation, high-frequency electrical stimulation, or application of exogenous neurotransmitter (3). All of these stimuli can induce large calcium transients and waves in astrocytes, but it was argued that they may be more related to pathophysiological states than to physiological signaling. In the experiments described by Parpura and Hayden (2) in this issue of PNAS, the authors used flash photolysis of caged calcium in astrocytes and simultaneous recording of neighboring neurons to determine the magnitude of intracellular calcium concentration necessary to stimulate glutamate release from astrocytes. The authors show that surprisingly small increases in intracellular calcium concentration (80–100 nM) are sufficient to induce large glutamate-activated currents in neighboring neurons. These results support the hypothesis that calcium elevation in astrocytes and subsequent release of glutamate can operate during physiological states and may not depend on large nonphysiological or traumatic stimuli.
What properties of synaptic transmission are regulated by glial synaptic signaling? At individual synapses in hippocampus, the stimulation of a glial calcium signal has been shown to depress the response at neuronal synapses (17, 18). This finding suggests that one role of glia may be to modulate the activity at neuron-to-neuron synapses. In retina, the induction of a glial calcium signal has been shown to modulate the response of retinal ganglion cells to light (19), and in visual cortex, measurements with certain slow voltage-activated fluorescent dyes, which primarily record glial cell depolarization, give an accurate map of neocortical receptive fields (20). One might speculate that glial signals not only may be consequences of neuronal signaling but may in fact shape neuronal responses to stimuli.
Future studies must address whether OPCs release transmitter like astrocytes after they are synaptically stimulated and whether and under what conditions astrocytes are stimulated by neurotransmitter released from neurons or other glia. In fact, different glial cells may form types of chemical synapses with each other. But perhaps the most challenging and important questions to be addressed in the future relate to specifying exactly what aspects of neural processing are determined and shaped by glia. For example, do they participate in synaptic plasticity, or do they shape receptive fields? Ultimately, experiments will have to be designed in which glia are selectively removed from or silenced in central nervous system circuits. Answers to these and other questions will likely reshape our view of neural circuits in the next century from exclusively neuronal to glial/neuronal circuits.
Footnotes
See companion article on page 8629.
References
- 1.Bergles D E, Roberts J D B, Somogyi P, Jahr C E. Nature (London) 2000;405:187–191. doi: 10.1038/35012083. [DOI] [PubMed] [Google Scholar]
- 2.Parpura V, Haydon P G. Proc Natl Acad Sci USA. 2000;97:8629–8634. doi: 10.1073/pnas.97.15.8629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Araque A, Parpura V, Sanzgiri R P, Haydon P G. Trends Neurosci. 1999;22:208–215. doi: 10.1016/s0166-2236(98)01349-6. [DOI] [PubMed] [Google Scholar]
- 4.Cotrina M L, Nedergaard M. The Neuroscientist. 2000;6:120–124. [Google Scholar]
- 5.Miller R H. Trends Neurosci. 1996;19:92–96. doi: 10.1016/s0166-2236(96)80036-1. [DOI] [PubMed] [Google Scholar]
- 6.Raff M C, Miller R H, Noble M. Nature (London) 1983;303:390–396. doi: 10.1038/303390a0. [DOI] [PubMed] [Google Scholar]
- 7.Steinhauser C, Gallo V. Trends Neurosci. 1996;19:339–345. doi: 10.1016/0166-2236(96)10043-6. [DOI] [PubMed] [Google Scholar]
- 8.Condorelli D F, Conti F, Gallo V, Kirchhoff F, Seifert G, Steinhauser C, Verkhratsky A, Yuan X. Adv Exp Med Biol. 1999;468:49–67. doi: 10.1007/978-1-4615-4685-6_5. [DOI] [PubMed] [Google Scholar]
- 9.Fraser D D, Mudrick-Donnon L A, MacVicar B A. Glia. 1994;11:83–93. doi: 10.1002/glia.440110203. [DOI] [PubMed] [Google Scholar]
- 10.Rosewater K, Sontheimer H. Brain Res. 1994;636:73–80. doi: 10.1016/0006-8993(94)90177-5. [DOI] [PubMed] [Google Scholar]
- 11.Hertz L, Schousboe I, Hertz L, Schousboe A. Prog Neuropsychopharmacol Biol Psychiatry. 1984;8:521–527. doi: 10.1016/0278-5846(84)90010-1. [DOI] [PubMed] [Google Scholar]
- 12.Gallo V, Zhou J M, McBain C J, Wright P, Knutson P L, Armstrong R C J. J Neurosci. 1996;16:2659–2670. doi: 10.1523/JNEUROSCI.16-08-02659.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cornell-Bell A H, Finkbeiner S M, Cooper M S, Smith S J. Science. 1990;247:470–473. doi: 10.1126/science.1967852. [DOI] [PubMed] [Google Scholar]
- 14.Parpura V, Basarsky T A, Liu F, Jeftinija K, Jeftinija S, Haydon P G. Nature (London) 1994;369:744–747. doi: 10.1038/369744a0. [DOI] [PubMed] [Google Scholar]
- 15.Pasti L, Volterra A, Pozzan T, Carmignoto G. J Neurosci. 1997;17:7817–7830. doi: 10.1523/JNEUROSCI.17-20-07817.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kang J, Jiang L, Goldman S A, Nedergaard M. Nat Neurosci. 1998;1:683–692. doi: 10.1038/3684. [DOI] [PubMed] [Google Scholar]
- 17.Bezzi P, Carmignoto G, Pasti L, Vesce S, Rossi D, Rizzini B L, Pozzan T, Volterra A. Nature (London) 1998;391:281–285. doi: 10.1038/34651. [DOI] [PubMed] [Google Scholar]
- 18.Past L. J Neurosci. 1997;17:7817–7830. doi: 10.1523/JNEUROSCI.17-20-07817.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Newman E A, Zahns K R. J Neurosci. 1998;18:4022–4028. doi: 10.1523/JNEUROSCI.18-11-04022.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Blasdel G G. Annu Rev Physiol. 1989;51:561–581. doi: 10.1146/annurev.ph.51.030189.003021. [DOI] [PubMed] [Google Scholar]