

Abstract
Adherens junctions connect the actin cytoskeleton of neighboring cells through transmembrane cadherin receptors and a network of adaptor proteins. The interactions between these adaptors and cadherin as well as the activity of actin regulators localized to adherens junctions are tightly controlled to facilitate cell junction assembly or disassembly in response to changes in external or internal forces and/or signaling. Phosphorylation of tyrosine, serine, or threonine residues acts as a switch on the majority of adherens junction proteins, turning “on” or “off” their interactions with other proteins and/or their enzymatic activity. Here, we provide an overview of the kinases and phosphatases regulating phosphorylation of adherens junction proteins and bring examples of phosphorylation events leading to the assembly or disassembly of adherens junctions, highlighting the important role of phosphorylation switches in regulating their dynamics.
1. Introduction
Adherens junctions (AJs) are cell-cell adhesion sites where calcium-dependent cadherin receptors bind with their extracellular domains to cadherins on opposing cells and with their cytoplasmic tails connect—via adaptors—to filamentous actin [1, 2]. By essentially providing a physical link between the actin cytoskeleton of neighboring cells AJs facilitate the integration of individual cells into a tissue. Additionally, AJs are instrumental in setting up and maintaining the apicobasal polarity of epithelial cells [3, 4], they function as mechanosensors [5] and serve as a nexus for signaling affecting important cell decisions, such as survival and differentiation [6].
During the development and lifetime of an organism, cells frequently change shape and position relative to their neighbors. Hence, the ability of cells to regulate their adhesive interactions plays a key role during tissue morphogenesis, repair, and renewal [3, 7, 8]. Defects in the adhesive characteristics of epithelial cells are pathological signs and loss of cell-cell adhesion can generate dedifferentiation and invasiveness of human carcinoma cells [9]. Thus, there is great interest in understanding the factors that affect assembly and disassembly of cell-cell adhesion at the molecular level.
When considering regulatory mechanisms controlling AJ proteins, we distinguish between three subsequent steps of regulation: synthesis, localization, and activation. First, a cell controls whether proteins are synthesized or not. Indeed, transcriptional regulation of E-cadherin, notably by the snail transcription factor, plays an important role in the breaking down of AJs accompanying epithelial to mesenchymal transition [10]. Once a protein is expressed the cell can determine its localization by controlling its transport. In fact, both exocytosis and endocytosis of E-cadherin are tightly controlled and the balance between the two processes has been shown to regulate AJ turnover both in vitro and in vivo [6]. Finally, a cell can control the activity and interactions of a protein at a given location by posttranslational modifications. These modifications include glycosylation, lipidation, ubiquitination, acetylation, proteolysis, and phosphorylation [11]. Phosphorylation of tyrosine (Y), serine (S), or threonine (T) residues, the topic of this review, is a rapid and reversible form of regulation affecting the majority of AJ proteins [12–16]. In some cases, posttranslational modifications have secondary effects on transcription and/or protein transport [17, 18]. However, here we will focus on the more direct mechanisms in which AJs are regulated by phosphorylation. First, we will introduce the enzymes responsible for phosphorylation and dephosphorylation at AJs and discuss how they are recruited into AJ and activated. Next, we will describe the targets of phosphorylation within AJ and by examining the consequences of specific phosphorylation events will show how phosphorylation is involved both in assembly and disassembly of AJ, essentially driving the dynamics of this highly responsive structure. In the end, we will point out open questions and suggest methods to address them.
2. Recruitment of Protein Kinases and Phosphatases into AJ
So far, twelve S/T kinases and one S/T phosphatase have been implicated in regulating phosphorylation of AJ proteins, and they are all cytoplasmic (Table 1). Prominent kinases in this group include PKC-α, cAMP-dependent protein kinase, Casein Kinase 1, Pak1, and ROCK1. Nine tyrosine kinases and twelve tyrosine phosphatases have been shown to be active in AJ, roughly half of them cytoplasmic and half part of a transmembrane receptor (Table 1). Prominent tyrosine kinases include the cytoplasmic Src, Fyn, Fer, and Abl, and the receptors of epidermal and hepatocyte growth factors. Major phosphatases involved are the cytoplasmic PTP-1B, PTP-PEST, SHP-1, SHP-2, and receptor-type tyrosine-protein phosphatases Mu, U, and Kappa.
Table 1.
Kinases and phosphatases regulating phosphorylation of AJ proteins.
| Gene symbol | Protein name | Phosphorylation type | Localization | Reference |
|---|---|---|---|---|
| Kinases | ||||
| SRC | Proto-oncogene tyrosine- | Tyr | nonreceptor | [23, 30, 31] |
| protein kinase Src | ||||
| CSK | c-src tyrosine kinase | Tyr | nonreceptor | [24, 32, 33] |
| FYN | Tyrosine-protein kinase Fyn | Tyr | nonreceptor | [34–36] |
| ABL1 | Abl1 | Tyr | nonreceptor | [37, 38] |
| SYK | Tyrosine protein kinase SYK | Tyr | nonreceptor | [39, 40] |
| PTK2B | Protein-tyrosine kinase 2-beta | Tyr | nonreceptor | [41–43] |
| FER | Tyrosine-protein kinase Fer | Tyr | nonreceptor | [44] |
| EGFR | Epidermal growth factor receptor | Tyr | Receptor | [45] |
| cMET/HGF | Hepatocyte growth factor receptor | Tyr | Receptor | [46] |
| PRKCA | Protein kinase C alpha type | Ser/Thr | nonreceptor | [47] |
| PRKACA | cAMP-dependent protein | Ser/Thr | nonreceptor | [48, 49] |
| Kinase catalytic subunit alpha | ||||
| ROCK1 | Rho-associated, coiled-coil | Ser/Thr | nonreceptor | [50] |
| containing protein kinase 1 | ||||
| PRKCD | Protein kinase C delta type | Ser/Thr | nonreceptor | [51, 52] |
| CSNK1E | Casein kinase I isoform epsilon | Ser/Thr | nonreceptor | [53] |
| CSNK2A1 | Casein kinase 2 | Ser/Thr | nonreceptor | [54] |
| PAK1 | Serine/threonine-protein kinase PAK 1 | Ser/Thr | nonreceptor | [55–57] |
| MAPK8 | JNK | Ser/Thr | nonreceptor | [58] |
| PRKD1 | Protein kinase D1 | Ser/Thr | nonreceptor | [59] |
| PRKCI | Atypical protein kinase C-lambda/iota | Ser/Thr | nonreceptor | [60] |
| PRKCZ | Protein kinase C zeta type | Ser/Thr | nonreceptor | [60] |
| MARK2 | MAP/microtubule affinity- | Ser/Thr | nonreceptor | [61] |
| Regulating kinase 2, Par-1 | ||||
|
| ||||
| Phosphatases | ||||
| PTPN1 | Tyrosine-protein phosphatase non | Tyr | nonreceptor | [19, 62–64] |
| receptor type 1, PTP1B | ||||
| PTPN6 | Tyrosine-protein phosphatase non | Tyr | nonreceptor | [65] |
| receptor type 6, SHP1 | ||||
| PTPN11 | Tyrosine-protein phosphatase non- | Tyr | nonreceptor | [66] |
| receptor type 11, SHP2 | ||||
| PTPN12 | Tyrosine-protein phosphatase non- | Tyr | nonreceptor | [67] |
| receptor type 12, PTP-PEST | ||||
| PTPN14 | Tyrosine-protein phosphatase non- | Tyr | nonreceptor | [68] |
| receptor type 14, PEZ | ||||
| ACP1 | Acid phosphatase of erythrocyte, LMW-PTP | Tyr | nonreceptor | [69, 70] |
| PTPRJ | Receptor-type tyrosine-protein | Tyr | Receptor | [71] |
| phosphatase eta (R-PTP-eta), DEP1 | ||||
| PTPRM | Receptor-type tyrosine-protein | Tyr | Receptor | [72–74] |
| phosphatase mu (RPTP mu) | ||||
| PTPRT | Receptor-type tyrosine-protein | Tyr | Receptor | [75] |
| phosphatase T (R-PTP-T) | ||||
| PTPRU | Receptor-type tyrosine-protein | Tyr | Receptor | [76, 77] |
| phosphatase U (R-PTP-U) | ||||
| PTPRK | Receptor-type tyrosine-protein | Tyr | Receptor | [78, 79] |
| phosphatase kappa | ||||
| PTPRF | Receptor-type tyrosine-protein | Tyr | Receptor | [80–82] |
| phosphatase F, LAR | ||||
| PPP2CA | Serine/threonine-protein phosphatase 2A | Ser/Thr | nonreceptor | [83–85] |
| catalytic subunit alpha isoform | ||||
Some of these kinases and phosphatases have been localized to AJ by immunofluorescence (e.g., [19–21]) and others have been shown to associate with AJ by coimmunoprecipitation (e.g., [21–23]), but the exact mechanism of recruitment into AJ of most of them is largely unknown. A few were shown to bind directly with cadherin, such as CSK with VE-cadherin and PTP-1B with E- and N-cadherin [19, 24, 25]; others bind one of the catenins (adaptor proteins linking cadherin with actin), such as MET and PTPRF with β-catenin [26, 27] and ROCK1 with p120-catenin [21]; some interact with other AJ adaptor proteins, such as PRKCA with vinculin and ROCK1 with Shroom3 [28, 29].
While it is most likely every kinase and phosphatase can recognize at least one docking site within the AJ, it is not currently known which of the kinases and phosphatases reside in AJ permanently and which are transient components, homing in to phosphorylate or dephosphorylate AJ proteins only under specific conditions. Even permanent residents may not always be active, as most kinases and phosphatases need themselves to be activated.
3. Activation of Kinases and Phosphatases in AJ
Receptor tyrosine kinases are commonly activated by an external ligand, such as a growth factor or cytokine, which induces dimerization, cis-phosphorylation or autophosphorylation and activation of the catalytic domain [124, 125]. Receptor tyrosine phosphatases may be activated by homophilic association with their counterparts on neighboring cells [126], as well as by tyrosine phosphorylation [127]. Several S/T kinases are activated by binding of Rho GTPases, for example ROCK1 is activated by RhoA and PAK1 is activated by Rac1 and Cdc42 [128, 129]. S/T kinases are also regulated by tyrosine phosphorylation and tyrosine kinases and phosphatases are regulated by S/T phosphorylation, in a complex web of feedback and feedforward loops that is poorly understood (Figure 1). For example, Src phosphorylates PRKCD, which phosphorylates PTPN6, which in turn dephosphorylates SRC (feedback) [130–132]; PRKACA phosphorylates Src and Csk, and Csk also phosphorylates Src (feedforward) [133–135].
Figure 1.
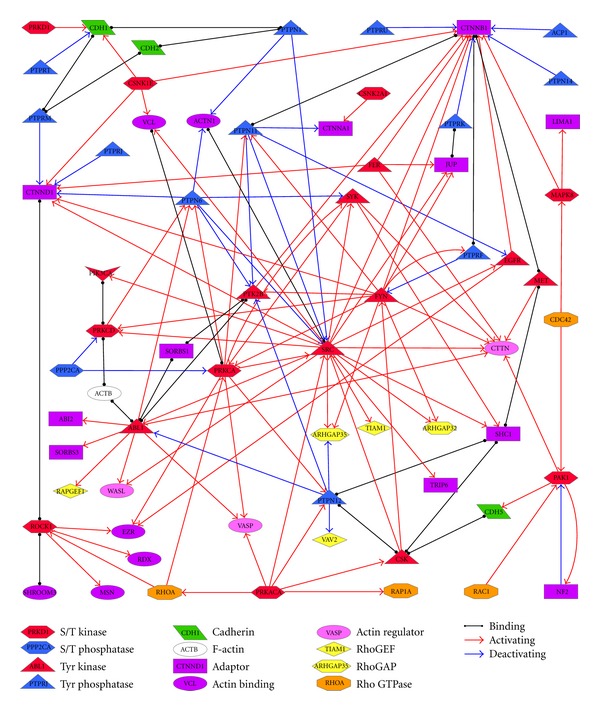
Network of phosphorylation enzymes and targets in the adherens junction.
As will be discussed further below, some of the phosphorylation events serve to activate the kinases or phosphatases and others are inhibitory. One well-understood example of kinase activation is the mechanism of activation of Src. As reviewed in [136], the family of Src tyrosine kinases can be found in a nonactive “closed” conformation or in an “open” active conformation, depending on the phosphorylation status of a tyrosine residue at the C-terminus. When this residue is phosphorylated, it interacts with an SH2 domain in the middle of Src, blocking the catalytic site. Upon dephosphorylation of this specific tyrosine, the SH2 domain is released, and the protein unfolds, allowing autophosphorylation of another tyrosine residue situated within the enzyme's activation loop, rendering the kinase fully active [137]. It is important to point out that cadherin ligation and clustering may act as an activation signal for some kinases. Most notably, Src and Fer have been shown to be recruited to the membrane upon cadherin binding [138, 139], and EGFR signaling was shown to be stimulated by AJ formation independently of EGF ligand [140]. Furthermore, cadherin clustering has been found to indirectly induce activation of Rho GTPases [141], which in turn could activate S/T kinases.
4. Phosphorylation Targets within the AJ
The AJ can conceptually be divided into four layers (Table 2). The first, in the plane of the membrane, is where cadherins and other transmembrane proteins, such as nectin and AJAP1, reside. The next layer consists of membrane-bound adaptors, such as ERM proteins and MAGI1, and adaptors that directly bind transmembrane proteins, such as p120- and β-catenin (bind cadherin) and afadin (binds nectin). The following layer is composed of adaptor proteins, such as α-catenin and vinculin, which bind to the second layer adaptors and also bind F-actin. F-actin, along with actin-binding proteins, such as α-actinin, and actin regulators, such as cortactin, would be considered the last layer. Regulatory proteins, such as GAPs, GEFs, and GTPases, can be found throughout the AJ as reviewed in [14, 142].
Table 2.
Targets of phosphorylation in AJ.
| Gene symbol | Protein name | Phosphorylated residue | Reference |
|---|---|---|---|
| Transmembrane | |||
| CDH1 | Ecadherin | S/Y | [54, 86, 87] |
| PVRL1 | Nectin | Y | [88] |
|
| |||
| Cadherin- or membrane bound | |||
| CTNNB1 | β-catenin | S/T/Y | [16, 89, 90] |
| EZR, RDX, MSN | ERM proteins (ezrin/radixin/moesin) | S/T/Y | [91–93] |
| CTNND1 | p120-catenin | S/T/Y | [94, 95] |
| JUP | Gamma-catenin | Y | [96] |
| PARD3 | Partitioning defective 3 homolog | S/T/Y | [97, 98] |
|
| |||
| Secondary adaptors | |||
| CTNNA1 | α-catenin | S/T/Y | [99, 100] |
| VCL | Vinculin | S/Y | [101, 102] |
| LIMA1 | Eplin | S | [103] |
| VASP | Vasodilator-stimulated phosphoprotein | S/T/Y | [104–106] |
| SHC1 | SHC-transforming protein 1 | Y | [107, 108] |
|
| |||
| Actin and actin regulators | |||
| ACTN1 | α-actinin | S/Y | [109, 110] |
| CTTN | Cortactin | S/T/Y | [111, 112] |
| ACTB | F-actin | S/Y | [113–115] |
|
| |||
| GTPASE regulators | |||
| PI3K | Phospho-inositide-3-kinase | Y | [116] |
| RAPGEF1 | Rap guanine nucleotide exchange factor 1 | Y | [117] |
| ARHGAP35 | rho GAP p190A | Y | [118, 119] |
| ARHGAP32 | p200RhoGAP | Y | [120, 121] |
| TIAM1 | T-lymphoma invasion and metastasis-inducing protein 1 | Y | [122] |
| VAV2 | Vav 2 guanine nucleotide exchange factor | Y | [122, 123] |
There is evidence demonstrating both Y and S/T phosphorylation of proteins in all layers of the AJ (Table 2). As illustrated in Figure 1, often the same kinase will phosphorylate proteins from different layers. For example, Abl phosphorylates actin regulators WASP and VASP [143, 144], as well as cadherin-bound adaptor δ-catenin [145] and second layer adaptors Abi2 and Vinexin (SORBS3) [146, 147].
Some AJ proteins have additional functions in the cell, and phosphorylation is also involved in regulating their non-AJ roles [148]. The most notable example is β-catenin, which plays an important role in the Wnt signaling pathway as a cotranscription factor of TCF/LEF [149]. Whether nonjunctional β-catenin will reach the nucleus or not depends on whether it is phosphorylated by GSK3 and Casein Kinse I in the “destruction complex” [150]. However, such phosphorylation events taking place outside the context of AJ are beyond the scope of this paper.
For the phosphorylation events occurring within AJ an important question is how do they affect the target proteins?
5. Consequences of Phosphorylation on AJ Proteins
A phosphorylated tyrosine, serine, or threonine residue can affect a protein in three major ways: it can increase the affinity for another protein, it can inhibit a protein-protein interaction, or it can activate enzymatic activity. In proteins with an intramolecular interaction, phosphorylation and dephosphorylation can elicit a conformational change in the protein. AJ components provide examples of each type of these outcomes, as detailed henceforth.
(1) Turn “on” Protein-Protein Interaction —
Tyrosine phosphorylation can create a docking site for an SH2 or PTB domain of a partner protein. For example, tyrosine phosphorylation of cadherin creates docking sites for the SH2 domain of the adaptor SHC1 [151] and the PTB domain of the cell polarity protein Numb [152]. As mentioned earlier, SRC family kinases are inhibited by an intramolecular interaction between a central SH2 domain and a phosphorylated tyrosine at the C-terminus [137].
(2) Turn “off” Protein-Protein Interaction —
Examples of interaction inhibition by phosphorylation are also found both between different proteins and intramolecularly: tyrosine phosphorylation of VE-cadherin at certain residues prevents the binding of p120-catenin and β-catenin [153]; phosphorylation of a threonine residue in the C-terminal actin binding domain of ERM proteins interferes with its interaction with the N-terminal FERM domain, helping to keep the protein in an active open conformation [154].
(3) Turn “on” Enzymatic Activity —
Activation of the catalytic activity of tyrosine kinases and phosphatases by tyrosine phosphorylation has already been mentioned above [127]. Another important example is the activation of the motor activity of myosin by the phosphorylation of serine and threonine residues of myosin light chain [155].
We next address the question what are the ramifications of phosphorylation of AJ proteins on AJ structure and dynamics.
6. Global versus Specific Consequences of Phosphorylation on AJ Structure and Dynamics
Numerous experiments have been carried out over the years to address the role of Y and S/T phosphorylation in regulating AJ. Early experiments used broad-spectrum chemical inhibitors of kinases or phosphatases to conclude that phosphorylation negatively impacts cadherin function. For example, inhibition of S/T phosphatases by Okadaic acid or Calyculin-A was reported to lead to complete disassembly of AJ within an hour, and this disruption was attributed to an increase in S/T phosphorylation of β-catenin [89]. However, Calyculin-A has also been shown to increase actomyosin contractility in cells [156], suggesting that the disruption of AJ in the above mentioned study may have been caused by mechanical tension at the junctions exceeding their adhesive strength. Inhibition of tyrosine phosphatases with sodium orthovanadate was reported to lead within minutes to a dramatic increase in phosphotyrosine signals at AJ, followed by the disassembly of AJ [157]. Consistent with the notion that excessive tyrosine phosphorylation in AJ causes their disassembly, cells expressing constitutively active Src kinase lost their AJ, and inhibition of tyrosine kinase activity by the drug tyrphostin was able to restore AJ in the Src-transformed cells [157]. These and similar experiments have led researchers in the late 90s of the previous century to the general conclusion that phosphorylation is a negative regulator of AJ.
However, in more recent years, there is accumulating evidence for a positive role of phosphorylation in AJ assembly, mainly coming out of loss-of-function experiments of specific kinases. For example, SRC and FYN were found to be essential for the formation of AJ in mouse keratinocytes [158]. Moreover, SRC activity was shown in vitro to be important for the recruitment of PI3K to AJ and the ability of cells to expand nascent cadherin-adhesive contacts [159]. Along the same lines, ABL1 tyrosine kinase activity was shown to be important for the maintenance of adherens junctions in epithelial cells [37], and S/T phosphorylation of E-cadherin by protein kinase D1 (PRKD1) was found to be associated with increased cellular adhesion and decreased cellular motility in prostate cancer [59].
Hence, the emerging view is that it is not possible to generalize the effect of phosphorylation on AJ. With some phosphorylation events leading to the switching “on” of a protein or interaction and other phosphorylation events, even on the same protein, serving as a switch “off”, the effect of phosphorylation on AJ dynamics has to be examined on a residue-by-residue basis. After we delineate the effect of each individual phosphorylation event, we should be able to integrate this information into a single network of interconnected switches and perhaps then we can follow the global effects of a single phosphorylation switch, starting, for example, with hepatocyte growth factor stimulation [160].
7. Consequences of Specific Phosphorylation Events on AJ, Composition and Dynamics
We close this paper by giving a few examples of cases in which the consequences of specific phosphorylation events are known. The phosphorylation events presented occur on proteins from each layer of the AJ as well as one cell polarity protein.
(1) Cadherin —
Serine phosphorylation of residues S840, S851 and S853 in the C-terminus of human E-cadherin (likely by CSNK1E or PRKD1) increases the binding affinity towards β-catenin, whereas phosphorylation of S846 is said to inhibit the same interaction [86]. Stronger binding of β-catenin to E-cadherin is conducive to a stronger AJ structure. Tyrosine phosphorylation of VE-cadherin at two critical tyrosines, Y658 and Y731, is sufficient to prevent the binding of p120- and β-catenin, respectively [161]. Phosphorylation by Src of three tyrosines in position 753–755 on human E-cadherin creates a docking site for the E3-ligase Hakai [162]. Ubiquitination of E-cadherin by Hakai leads to internalization of E-cadherin facilitating disassembly of the AJ [162, 163].
(2) P120-Catenin —
Eight tyrosine residues in the N-terminus of p120-catenin can be phosphorylated by Src [94]. Upon phosphorylation, these sites serve as docking sites for the recruitment of interacting proteins carrying SH2 domains, such as the tyrosine phosphatase SHP-1 [164]. Under certain conditions tyrosine phosphorylation of p120-catenin was shown to increase its affinity to cadherin, while in other instances such an increase was not observed (reviewed in [95]). The affinity of p120-catenin to cadherin is significant for AJ dynamics because p120-catenin protects cadherin from being internalized [165].
(3) Zyxin —
Phosphorylation of S142 of zyxin is thought to result in the release of an intramolecular head-tail interaction [166]. Opening of the protein expose its ACTA repeats that recruit VASP, whose actin polymerization activity (see below) is important for AJ assembly and maintenance. Since zyxin-mediated recruitment of VASP has a positive effect on AJ [167, 168], it is not surprising that expression of a zyxin phosphomimetic mutant results in ultrastable AJ [166].
(4) VASP (Vasodilator Stimulated Phosphoprotein) —
As its name suggests, VASP is often found phosphorylated in cells. Three phosphorylation sites on residues S157, S239, and T274 are phosphorylated by PKA and PKG, as well as PKC [169, 170] and dephosphorylated by unknown phosphatase/s. The phosphorylation of VASP was shown to reduce its affinity towards actin [171] and essentially turn off its actin bundling and anticapping/elongation activity [171, 172]. VASP-mediated actin elongation is important for the formation of AJ and for the maintenance of actin structures associated with AJ [173, 174]. Thus, the consequence of VASP phosphorylation is to negatively regulate AJ assembly and maintenance.
(5) PARD3 —
In Drosophila epithelial cells the par-3 ortholog Bazooka is confined to AJ as a result of phosphorylation by either apical or basal polarity complexes [4]. At the apical side of cells Bazooka is phosphorylated by aPKC, resulting in its release from the cortex [175, 176]. In the basolateral membrane Par1 kinase phosphorylates Bazooka on unique sites that also lead to its cortical release [4]. Recently, it was shown that the ratio between Par-1 and aPKC determines the position of Bazooka and AJ along the lateral side and a reduction in Par-1 kinase activity leads to a basal shift of AJ followed by folding of the epithelial sheet [177].
8. Conclusions and Outlook
From the examples presented above, it is clear that phosphorylation switches play a pivotal role in regulating AJ assembly and disassembly dynamics. At the same time it is also clear that our knowledge is only scratching the surface of the phosphorylation network regulating AJ. For the majority of known phosphorylation events in AJ, we know either of a kinase or of a phosphatase involved, but rarely do we know both. Furthermore, while traditional biochemistry techniques have facilitated the characterization of a handful of phosphorylation events on AJ proteins, phosphoproteomic data indicates that the majority of AJ proteins are phosphorylated on multiple serine/threonine and tyrosine residues [178]. Phosphoproteomics, which utilizes a variety of techniques to label cells, enrich for phosphorylated peptides and identify them using mass-spectrometry (reviewed in [179–181]), not only highlights the hole in our knowledge but also offers the means to fill it.
Phospho-proteomics offers an unbiased and comprehensive snapshot of phosphorylation events, and several different approaches can be taken to elucidate phosphorylation switches in AJ: during normal assembly and maturation, following a signal for disassembly, or when a certain kinase or phosphatase is activated or missing (e.g., [39, 182–184]). The phospho-proteomic data obtained, especially if it is dynamic, can be used for a systems level analysis of phosphorylation switches in AJ [185, 186], but it seems likely to us that before the network can be modeled in a meaningful way more in depth characterization of specific phosphorylation events will be necessary, using cell biological techniques.
While for the discovery and mapping of phosphorylation events in AJ, one wants to be as comprehensive as possible, when it comes to characterizing a particular switch the more specific the tools, the better. One example of a specific tool is phosphorylation site-specific antibodies, such as those recognizing individual phosphorylation events on β-catenin and p-120-catenin [187, 188]. Another example are site-specific phospho-mimetic or nonphosphorylatable mutations, such as those successfully applied to the study of the effects of phosphorylation on cortactin, VASP, VE-cadherin, zyxin, and paxillin [153, 166, 189–191].
Facing an ever-changing landscape of forces and signaling cues, a cell must respond rapidly by adjusting the strength of its AJs according to need. For this it relies on continuous turnover and assembly of core AJ components. Phosphorylation is particularly suitable for regulating the balance between assembly and disassembly as it is rapid and affects the AJ proteins directly. Feedback loops must guarantee a combination of phosphorylated residues at AJ that matches the requirements for a given condition. Experiments have shown that when it comes to phosphorylation both “all on” and “all off” treatments are deleterious to AJ. The challenge now is to elucidate the mechanisms by which the cell maintains a “just right” level of phosphorylation in AJ. While phosphorylation is probably the most prominent regulatory switch controlling cell adhesion, other switches, such as GTPases, lipids and proteases, do exist [192]. A future challenge, therefore, will be to integrate the phosphorylation switch network with the other regulatory switches to facilitate a true understanding of how different signaling pathways and force regulate AJ dynamics.
Acknowledgment
This paper was made possible by an NRF fellowship awarded to R. Zaidel-Bar by the Singapore National Research Foundation.
References
- 1.Green KJ, Getsios S, Troyanovsky S, Godsel LM. Intercellular junction assembly, dynamics, and homeostasis. Cold Spring Harbor Perspectives in Biology. 2010;2(2):p. a000125. doi: 10.1101/cshperspect.a000125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Takeichi M. Cadherin cell adhesion receptors as a morphogenetic regulator. Science. 1991;251(5000):1451–1455. doi: 10.1126/science.2006419. [DOI] [PubMed] [Google Scholar]
- 3.Baum B, Georgiou M. Dynamics of adherens junctions in epithelial establishment, maintenance, and remodeling. Journal of Cell Biology. 2011;192(6):907–917. doi: 10.1083/jcb.201009141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.St Johnston D, Sanson B. Epithelial polarity and morphogenesis. Current Opinion in Cell Biology. 2011;23(5):540–546. doi: 10.1016/j.ceb.2011.07.005. [DOI] [PubMed] [Google Scholar]
- 5.Yonemura S, Wada Y, Watanabe T, Nagafuchi A, Shibata M. α-Catenin as a tension transducer that induces adherens junction development. Nature Cell Biology. 2010;12(6):533–542. doi: 10.1038/ncb2055. [DOI] [PubMed] [Google Scholar]
- 6.Lecuit T. Adhesion remodeling underlying tissue morphogenesis. Trends in Cell Biology. 2005;15(1):34–42. doi: 10.1016/j.tcb.2004.11.007. [DOI] [PubMed] [Google Scholar]
- 7.Harris TJC. Adherens junction assembly and function in the Drosophila embryo. International Review of Cell and Molecular Biology. 2012;293:45–83. doi: 10.1016/B978-0-12-394304-0.00007-5. [DOI] [PubMed] [Google Scholar]
- 8.Lynch AM, Hardin J. The assembly and maintenance of epithelial junctions in C. Elegans. Frontiers in Bioscience. 2009;14(4):1414–1432. doi: 10.2741/3316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Frixen UH, Behrens J, Sachs M, et al. E-cadherin-mediated cell-cell adhesion prevents invasiveness of human carcinoma cells. Journal of Cell Biology. 1991;113(1):173–185. doi: 10.1083/jcb.113.1.173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yoshino J, Monkawa T, Tsuji M, Inukai M, Itoh H, Hayashi M. Snail1 is involved in the renal epithelial-mesenchymal transition. Biochemical and Biophysical Research Communications. 2007;362(1):63–68. doi: 10.1016/j.bbrc.2007.07.146. [DOI] [PubMed] [Google Scholar]
- 11.Clark RSB, Bayir H, Jenkins LW. Posttranslational protein modifications. Critical Care Medicine. 2005;33(12):S407–S409. doi: 10.1097/01.ccm.0000191712.96336.51. [DOI] [PubMed] [Google Scholar]
- 12.Roura S, Miravet S, Piedra J, García De Herreros A, Duñachl M. Regulation of E-cadherin/catenin association by tyrosine phosphorylation. Journal of Biological Chemistry. 1999;274(51):36734–36740. doi: 10.1074/jbc.274.51.36734. [DOI] [PubMed] [Google Scholar]
- 13.Michalides R, Volberg T, Geiger B. Augmentation of adherens junction formation in mesenchymal cells by co-expression of N-CAM or short-term stimulation of tyrosine-phosphorylation. Cell Adhesion and Communication. 1994;2(6):481–490. doi: 10.3109/15419069409014212. [DOI] [PubMed] [Google Scholar]
- 14.Brunton VG, MacPherson IRJ, Frame MC. Cell adhesion receptors, tyrosine kinases and actin modulators: a complex three-way circuitry. Biochimica et Biophysica Acta. 2004;1692(2-3):121–144. doi: 10.1016/j.bbamcr.2004.04.010. [DOI] [PubMed] [Google Scholar]
- 15.Daniel JM, Reynolds AB. Tyrosine phosphorylation and cadherin/catenin function. BioEssays. 1997;19(10):883–891. doi: 10.1002/bies.950191008. [DOI] [PubMed] [Google Scholar]
- 16.Lilien J, Balsamo J. The regulation of cadherin-mediated adhesion by tyrosine phosphorylation/dephosphorylation of β-catenin. Current Opinion in Cell Biology. 2005;17(5):459–465. doi: 10.1016/j.ceb.2005.08.009. [DOI] [PubMed] [Google Scholar]
- 17.Harris TJC, Peifer M. Decisions, decisions: β-catenin chooses between adhesion and transcription. Trends in Cell Biology. 2005;15(5):234–237. doi: 10.1016/j.tcb.2005.03.002. [DOI] [PubMed] [Google Scholar]
- 18.Park M-H, Kim D-J, You S-T, et al. Phosphorylation of β-catenin at serine 663 regulates its transcriptional activity. Biochemical and Biophysical Research Communications. 2012;419(3):543–549. doi: 10.1016/j.bbrc.2012.02.056. [DOI] [PubMed] [Google Scholar]
- 19.Balsamo J, Leung T, Ernst H, Zanin MKB, Hoffman S, Lilien J. Regulated binding of a PTP1B-like phosphatase to N-cadherin: control of cadherin-mediated adhesion by dephosphorylation of β-catenin. Journal of Cell Biology. 1996;134(3):801–813. doi: 10.1083/jcb.134.3.801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tsukita S, Oishi K, Akiyama T, Yamanashi Y, Yamamoto T, Tsukita S. Specific proto-oncogenic tyrosine kinases of src family are enriched in cell-to-cell adherens junctions where the level of tyrosine phosphorylation is elevated. Journal of Cell Biology. 1991;113(4):867–879. doi: 10.1083/jcb.113.4.867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Smith AL, Dohn MR, Brown MV, Reynolds AB. Association of Rho-associated protein kinase 1 with E-cadherin complexes is mediated by p120-catenin. Molecular Biology of the Cell. 2012;23(1):99–110. doi: 10.1091/mbc.E11-06-0497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jiang J, Dean D, Burghardt RC, Parrish AR. Disruption of cadherin/catenin expression, localization, and interactions during HgCl2-induced nephrotoxicity. Toxicological Sciences. 2004;80(1):170–182. doi: 10.1093/toxsci/kfh143. [DOI] [PubMed] [Google Scholar]
- 23.Lee NPY, Cheng CY. Protein kinases and adherens junction dynamics in the seminiferous epithelium of the rat testis. Journal of Cellular Physiology. 2005;202(2):344–360. doi: 10.1002/jcp.20119. [DOI] [PubMed] [Google Scholar]
- 24.Baumeister U, Funke R, Ebnet K, Vorschmitt H, Koch S, Vestweber D. Association of Csk to VE-cadherin and inhibition of cell proliferation. EMBO Journal. 2005;24(9):1686–1695. doi: 10.1038/sj.emboj.7600647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sheth P, Seth A, Atkinson KJ, et al. Acetaldehyde dissociates the PTP1B-E-cadherin-β-catenin complex in Caco-2 cell monolayers by a phosphorylation-dependent mechanism. Biochemical Journal. 2007;402(2):291–300. doi: 10.1042/BJ20060665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Monga SPS, Mars WM, Pediaditakis P, et al. Hepatocyte growth factor induces Wnt-independent nuclear translocation of β-catenin after Met-β-catenin dissociation in hepatocytes. Cancer Research. 2002;62(7):2064–2071. [PubMed] [Google Scholar]
- 27.Aicher B, Lerch MM, Müller T, Schilling J, Ullrich A. Cellular redistribution of protein tyrosine phosphatases LAR and PTPσ by inducible proteolytic processing. Journal of Cell Biology. 1997;138(3):681–696. doi: 10.1083/jcb.138.3.681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ziegler WH, Tigges U, Zieseniss A, Jockusch BM. A lipid-regulated docking site on vinculin for protein kinase C. Journal of Biological Chemistry. 2002;277(9):7396–7404. doi: 10.1074/jbc.M110008200. [DOI] [PubMed] [Google Scholar]
- 29.Nishimura T, Takeichi M. Shroom3-mediated recruitment of Rho kinases to the apical cell junctions regulates epithelial and neuroepithelial planar remodeling. Development. 2008;135(8):1493–1502. doi: 10.1242/dev.019646. [DOI] [PubMed] [Google Scholar]
- 30.Shindo M, Wada H, Kaido M, et al. Dual function of Src in the maintenance of adherens junctions during tracheal epithelial morphogenesis. Development. 2008;135(7):1355–1364. doi: 10.1242/dev.015982. [DOI] [PubMed] [Google Scholar]
- 31.Owens DW, McLean GW, Wyke AW, et al. The catalytic activity of the Src family kinases is required to disrupt cadherin-dependent cell-cell contacts. Molecular Biology of the Cell. 2000;11(1):51–64. doi: 10.1091/mbc.11.1.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jin H, Garmy-Susini B, Avraamides CJ, Stoletov K, Klemke RL, Varner JA. A PKA-Csk-pp60Src signaling pathway regulates the switch between endothelial cell invasion and cell-cell adhesion during vascular sprouting. Blood. 2010;116(25):5773–5783. doi: 10.1182/blood-2010-07-296210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rengifo-Cam W, Konishi A, Morishita N, et al. Csk defines the ability of integrin-mediated cell adhesion and migration in human colon cancer cells: implication for a potential role in cancer metastasis. Oncogene. 2004;23(1):289–297. doi: 10.1038/sj.onc.1207041. [DOI] [PubMed] [Google Scholar]
- 34.Piedra J, Miravet S, Castaño J, et al. p120 catenin-associated Fer and Fyn tyrosine kinases regulate β-catenin Tyr-142 phosphorylation and β-catenin-α-catenin interaction. Molecular and Cellular Biology. 2003;23(7):2287–2297. doi: 10.1128/MCB.23.7.2287-2297.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Huang RYJ, Wang SM, Hsieh CY, Wu JC. Lysophosphatidic acid induces ovarian cancer cell dispersal by activating Fyn kinase associated with p120-catenin. International Journal of Cancer. 2008;123(4):801–809. doi: 10.1002/ijc.23579. [DOI] [PubMed] [Google Scholar]
- 36.Hsu KL, Fan HJ, Chen YC, et al. Protein kinase C-Fyn kinase cascade mediates the oleic acid-induced disassembly of neonatal rat cardiomyocyte adherens junctions. International Journal of Biochemistry and Cell Biology. 2009;41(7):1536–1546. doi: 10.1016/j.biocel.2008.12.016. [DOI] [PubMed] [Google Scholar]
- 37.Zandy NL, Playford M, Pendergast AM. Abl tyrosine kinases regulate cell- cell adhesion through Rho GTPases. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(45):17686–17691. doi: 10.1073/pnas.0703077104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Grevengoed EE, Loureiro JJ, Jesse TL, Peifer M. Abelson kinase regulates epithelial morphogenesis in Drosophila . Journal of Cell Biology. 2001;155(7):1185–1197. doi: 10.1083/jcb.200105102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Larive RM, Urbach S, Poncet J, et al. Phosphoproteomic analysis of Syk kinase signaling in human cancer cells reveals its role in cell-cell adhesion. Oncogene. 2009;28(24):2337–2347. doi: 10.1038/onc.2009.99. [DOI] [PubMed] [Google Scholar]
- 40.Zhang X, Shrikhande U, Alicie BM, Zhou Q, Geahlen RL. Role of the protein tyrosine kinase Syk in regulating cell-cell adhesion and motility in breast cancer cells. Molecular Cancer Research. 2009;7(5):634–644. doi: 10.1158/1541-7786.MCR-08-0371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van Buul JD, Anthony EC, Fernandez-Borja M, Burridge K, Hordijk PL. Proline-rich tyrosine kinase 2 (Pyk2) mediates vascular endothelial-cadherin-based cell-cell adhesion by regulating β-catenin tyrosine phosphorylation. Journal of Biological Chemistry. 2005;280(22):21129–21136. doi: 10.1074/jbc.M500898200. [DOI] [PubMed] [Google Scholar]
- 42.De Amicis F, Lanzino M, Kisslinger A, et al. Loss of proline-rich tyrosine kinase 2 function induces spreading and motility of epithelial prostate cells. Journal of Cellular Physiology. 2006;209(1):74–80. doi: 10.1002/jcp.20709. [DOI] [PubMed] [Google Scholar]
- 43.Cain RJ, Vanhaesebroeck B, Ridley AJ. The PI3K p110α isoform regulates endothelial adherens junctions via Pyk2 and Rac1. Journal of Cell Biology. 2010;188(6):863–876. doi: 10.1083/jcb.200907135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rosato R, Veltmaat JM, Groffen J, Heisterkamp N. Involvement of the tyrosine kinase Fer in cell adhesion. Molecular and Cellular Biology. 1998;18(10):5762–5770. doi: 10.1128/mcb.18.10.5762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hoschuetzky H, Aberle H, Kemler R. β-Catenin mediates the interaction of the cadherin-catenin complex with epidermal growth factor receptor. Journal of Cell Biology. 1994;127(5):1375–1380. doi: 10.1083/jcb.127.5.1375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Davies G, Jiang WG, Mason MD. HGF/SF modifies the interaction between its receptor c-Met, and the E-cadherin/catenin complex in prostate cancer cells. International Journal of Molecular Medicine. 2001;7(4):385–388. doi: 10.3892/ijmm.7.4.385. [DOI] [PubMed] [Google Scholar]
- 47.Williams CL, Noti JD. Reduced expression of Wnt-1 and E-cadherin, and diminished beta-catenin stability in MCF-7 breast cancer cells that overexpress protein kinase C-alpha. International Journal of Oncology. 2001;19(6):1227–1233. doi: 10.3892/ijo.19.6.1227. [DOI] [PubMed] [Google Scholar]
- 48.Boucher MJ, Laprise P, Rivard N. Cyclic AMP-dependent protein kinase A negatively modulates adherens junction integrity and differentiation of intestinal epithelial cells. Journal of Cellular Physiology. 2005;202(1):178–190. doi: 10.1002/jcp.20104. [DOI] [PubMed] [Google Scholar]
- 49.Leve F, De Souza W, Morgado-Díaz JA. A cross-link between protein kinase A and Rho-family GTPases signaling mediates cell-cell adhesion and actin cytoskeleton organization in epithelial cancer cells. Journal of Pharmacology and Experimental Therapeutics. 2008;327(3):777–788. doi: 10.1124/jpet.108.140798. [DOI] [PubMed] [Google Scholar]
- 50.Taguchi K, Ishiuchi T, Takeichi M. Mechanosensitive EPLIN-dependent remodeling of adherens junctions regulates epithelial reshaping. Journal of Cell Biology. 2011;194(4):643–656. doi: 10.1083/jcb.201104124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Chen CL, Chen HC. Functional suppression of E-cadherin by protein kinase Cδ . Journal of Cell Science. 2009;122(4):513–523. doi: 10.1242/jcs.035469. [DOI] [PubMed] [Google Scholar]
- 52.Singh R, Lei P, Andreadis ST. PKC-δ binds to E-cadherin and mediates EGF-induced cell scattering. Experimental Cell Research. 2009;315(17):2899–2913. doi: 10.1016/j.yexcr.2009.07.002. [DOI] [PubMed] [Google Scholar]
- 53.Casagolda D, Del Valle-Pérez B, Valls G, et al. A p120-catenin-CK1ε complex regulates Wnt signaling. Journal of Cell Science. 2010;123(15):2621–2631. doi: 10.1242/jcs.067512. [DOI] [PubMed] [Google Scholar]
- 54.Lickert H, Bauer A, Kemler R, Stappert J. Casein kinase II phosphorylation of E-cadherin increases E-cadherin/β- catenin interaction and strengthens cell-cell adhesion. Journal of Biological Chemistry. 2000;275(7):5090–5095. doi: 10.1074/jbc.275.7.5090. [DOI] [PubMed] [Google Scholar]
- 55.Pirraglia C, Walters J, Myat MM. Pak1 control of E-cadherin endocytosis regulates salivary gland lumen size and shape. Development. 2010;137(24):4177–4189. doi: 10.1242/dev.048827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Elloul S, Vaksman O, Stavnes HT, Trope CG, Davidson B, Reich R. Mesenchymal-to-epithelial transition determinants as characteristics of ovarian carcinoma effusions. Clinical and Experimental Metastasis. 2010;27(3):161–172. doi: 10.1007/s10585-010-9315-2. [DOI] [PubMed] [Google Scholar]
- 57.He H, Shulkes A, Baldwin GS. PAK1 interacts with β-catenin and is required for the regulation of the β-catenin signalling pathway by gastrins. Biochimica et Biophysica Acta. 2008;1783(10):1943–1954. doi: 10.1016/j.bbamcr.2008.04.016. [DOI] [PubMed] [Google Scholar]
- 58.Lee MH, Koria P, Qu J, Andreadis ST. JNK phosphorylates β-catenin and regulates adherens junctions. FASEB Journal. 2009;23(11):3874–3883. doi: 10.1096/fj.08-117804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jaggi M, Rao PS, Smith DJ, et al. E-cadherin phosphorylation by protein kinase D1/protein kinase Cμ is associated with altered cellular aggregation and motility in prostate cancer. Cancer Research. 2005;65(2):483–492. [PubMed] [Google Scholar]
- 60.Suzuki A, Yamanaka T, Hirose T, et al. Atypical protein kinase C is involved in the evolutionarily conserved PAR protein complex and plays a critical role in establishing epithelia-specific junctional structures. Journal of Cell Biology. 2001;152(6):1183–1196. doi: 10.1083/jcb.152.6.1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Elbert M, Cohen D, Müsch A. PAR1b promotes cell-cell adhesion and inhibits dishevelled-mediated transformation of Madin-Darby canine kidney cells. Molecular Biology of the Cell. 2006;17(8):3345–3355. doi: 10.1091/mbc.E06-03-0193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Balsamo J, Arregui C, Leung T, Lilien J. The nonreceptor protein tyrosine phosphatase PTP1B binds to the cytoplasmic domain of N-cadherin and regulates the cadherin-actin linkage. Journal of Cell Biology. 1998;143(2):523–532. doi: 10.1083/jcb.143.2.523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Pathre P, Arregui C, Wampler T, et al. PTP1B regulates neurite extension mediated by cell-cell and cell-matrix adhesion molecules. Journal of Neuroscience Research. 2001;63(2):143–150. doi: 10.1002/1097-4547(20010115)63:2<143::AID-JNR1006>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 64.Hernández MV, Wehrendt DP, Arregui CO. The protein tyrosine phosphatase PTP1B is required for efficient delivery of N-cadherin to the cell surface. Molecular Biology of the Cell. 2010;21(8):1387–1397. doi: 10.1091/mbc.E09-10-0880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Schnekenburger J, Mayerle J, Krüger B, et al. Protein tyrosine phosphatase κ and SHP-1 are involved in the regulation of cell-cell contacts at adherens junctions in the exocrine pancreas. Gut. 2005;54(10):1445–1455. doi: 10.1136/gut.2004.063164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ukropec JA, Hollinger MK, Salva SM, Woolkalis MJ. SHP2 association with VE-cadherin complexes in human endothelial cells is regulated by thrombin. Journal of Biological Chemistry. 2000;275(8):5983–5986. doi: 10.1074/jbc.275.8.5983. [DOI] [PubMed] [Google Scholar]
- 67.Espejo R, Rengifo-Cam W, Schaller MD, Evers BM, Sastry SK. PTP-PEST controls motility, adherens junction assembly, and Rho GTPase activity in colon cancer cells. American Journal of Physiology. 2010;299(2):C454–C463. doi: 10.1152/ajpcell.00148.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wadham C, Gamble JR, Vadas MA, Khew-Goodall Y. The protein tyrosine phosphatase Pez is a major phosphatase of adherens junctions and dephosphorylates β-catenin. Molecular Biology of the Cell. 2003;14(6):2520–2529. doi: 10.1091/mbc.E02-09-0577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Taddei ML, Chiarugi P, Cirri P, et al. β-catenin interacts with low-molecular-weight protein tyrosine phosphatase leading to cadherin-mediated cell-cell adhesion increase. Cancer Research. 2002;62(22):6489–6499. [PubMed] [Google Scholar]
- 70.Fang WB, Ireton RC, Zhuang G, Takahashi T, Reynolds A, Chen J. Overexpression of EPHA2 receptor destabilizes adherens junctions via a RhoA-dependent mechanism. Journal of Cell Science. 2008;121(3):358–368. doi: 10.1242/jcs.017145. [DOI] [PubMed] [Google Scholar]
- 71.Holsinger LJ, Ward K, Duffield B, Zachwieja J, Jallal B. The transmembrane receptor protein tyrosine phosphatase DEP1 interacts with p120ctn. Oncogene. 2002;21(46):7067–7076. doi: 10.1038/sj.onc.1205858. [DOI] [PubMed] [Google Scholar]
- 72.Zondag GCM, Reynolds AB, Moolenaar WH. Receptor protein-tyrosine phosphatase RPTPμ binds to and dephosphorylates the catenin p120(ctn) Journal of Biological Chemistry. 2000;275(15):11264–11269. doi: 10.1074/jbc.275.15.11264. [DOI] [PubMed] [Google Scholar]
- 73.Brady-Kalnay SM, Mourton T, Nixon JP, et al. Dynamic interactions of PTPμ with multiple cadherins in vivo. Journal of Cell Biology. 1998;141(1):287–296. doi: 10.1083/jcb.141.1.287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hiscox S, Jiang WG. Association of PTPμ with catenins in cancer cells: a possible role for E-cadherin. International Journal of Oncology. 1998;13(5):1077–1080. doi: 10.3892/ijo.13.5.1077. [DOI] [PubMed] [Google Scholar]
- 75.Besco JA, Hooft van Huijsduijnen R, Frostholm A, Rotter A. Intracellular substrates of brain-enriched receptor protein tyrosine phosphatase rho (RPTPρ/PTPRT) Brain Research. 2006;1116(1):50–57. doi: 10.1016/j.brainres.2006.07.122. [DOI] [PubMed] [Google Scholar]
- 76.Yan HX, He YQ, Dong H, et al. Physical and functional interaction between receptor-like protein tyrosine phosphatase PCP-2 and β-catenin. Biochemistry. 2002;41(52):15854–15860. doi: 10.1021/bi026095u. [DOI] [PubMed] [Google Scholar]
- 77.Yan HX, Yang W, Zhang R, et al. Protein-tyrosine phosphatase PCP-2 inhibits β-catenin signaling and increases E-cadherin-dependent cell adhesion. Journal of Biological Chemistry. 2006;281(22):15423–15433. doi: 10.1074/jbc.M602607200. [DOI] [PubMed] [Google Scholar]
- 78.Novellino L, De Filippo A, Deho P, et al. PTPRK negatively regulates transcriptional activity of wild type and mutated oncogenic β-catenin and affects membrane distribution of β-catenin/E-cadherin complexes in cancer cells. Cellular Signalling. 2008;20(5):872–883. doi: 10.1016/j.cellsig.2007.12.024. [DOI] [PubMed] [Google Scholar]
- 79.Wang C, Li Z, Yang Z, et al. The effect of receptor protein tyrosine phosphatase kappa on the change of cell adhesion and proliferation induced by N-acetylglucosaminyltransferase V. Journal of Cellular Biochemistry. 2010;109(1):113–123. doi: 10.1002/jcb.22387. [DOI] [PubMed] [Google Scholar]
- 80.Kypta RM, Su H, Reichardt LF. Association between a transmembrane protein tyrosine phosphatase and the cadherin-catenin complex. Journal of Cell Biology. 1996;134(6):1519–1529. doi: 10.1083/jcb.134.6.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Symons JR, Levea CM, Mooney RA. Expression of the leucocyte common antigen-related (LAR) tyrosine phosphatase is regulated by cell density through functional E-cadherin complexes. Biochemical Journal. 2002;365(2):513–519. doi: 10.1042/BJ20020381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Müller T, Choidas A, Reichmann E, Ullrich A. Phosphorylation and free pool of β-catenin are regulated by tyrosine kinases and tyrosine phosphatases during epithelial cell migration. Journal of Biological Chemistry. 1999;274(15):10173–10183. doi: 10.1074/jbc.274.15.10173. [DOI] [PubMed] [Google Scholar]
- 83.Nita-Lazar M, Rebustini I, Walker J, Kukuruzinska MA. Hypoglycosylated E-cadherin promotes the assembly of tight junctions through the recruitment of PP2A to adherens junctions. Experimental Cell Research. 2010;316(11):1871–1884. doi: 10.1016/j.yexcr.2010.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Suzuki K, Takahashi K. Induction of E-cadherin endocytosis by loss of protein phosphatase 2A expression in human breast cancers. Biochemical and Biophysical Research Communications. 2006;349(1):255–260. doi: 10.1016/j.bbrc.2006.08.037. [DOI] [PubMed] [Google Scholar]
- 85.Götz J, Probst A, Mistl C, Nitsch RM, Ehler E. Distinct role of protein phosphatase 2A subunit Cα in the regulation of E-cadherin and β-catenin during development. Mechanisms of Development. 2000;93(1-2):83–93. doi: 10.1016/s0925-4773(00)00267-7. [DOI] [PubMed] [Google Scholar]
- 86.Dupre-Crochet S, Figueroa A, Hogan C, et al. Casein kinase 1 is a novel negative regulator of E-cadherin-based cell-cell contacts. Molecular and Cellular Biology. 2007;27(10):3804–3816. doi: 10.1128/MCB.01590-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Qi J, Wang J, Romanyuk O, Siu CH. Involvement of Src family kinases in N-cadherin phosphorylation and β-catenin dissociation during transendothelial migration of melanoma cells. Molecular Biology of the Cell. 2006;17(3):1261–1272. doi: 10.1091/mbc.E05-10-0927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kikyo M, Matozaki T, Kodama A, Kawabe H, Nakanishi H, Takai Y. Cell-cell adhesion-mediated tyrosine phosphorylation of nectin-2δ, an immunoglobulin-like cell adhesion molecule at adherens junctions. Oncogene. 2000;19(35):4022–4028. doi: 10.1038/sj.onc.1203744. [DOI] [PubMed] [Google Scholar]
- 89.Serres M, Grangeasse C, Haftek M, Durocher Y, Duclos B, Schmitt D. Hyperphosphorylation of β-catenin on serine-threonine residues and loss of cell-cell contacts induced by calyculin A and okadaic acid in human epidermal cells. Experimental Cell Research. 1997;231(1):163–172. doi: 10.1006/excr.1996.3443. [DOI] [PubMed] [Google Scholar]
- 90.Moon HS, Choi EA, Park HY, et al. Expression and tyrosine phosphorylation of E-cadherin, β- and γ-catenin, and epidermal growth factor receptor in cervical cancer cells. Gynecologic Oncology. 2001;81(3):355–359. doi: 10.1006/gyno.2001.6163. [DOI] [PubMed] [Google Scholar]
- 91.Heiska L, Carpén O. Src phosphorylates ezrin at tyrosine 477 and induces a phosphospecific association between ezrin and a kelch-repeat protein family member. Journal of Biological Chemistry. 2005;280(11):10244–10252. doi: 10.1074/jbc.M411353200. [DOI] [PubMed] [Google Scholar]
- 92.Zhou R, Cao X, Watson C, et al. Characterization of protein kinase A-mediated phosphorylation of ezrin in gastric parietal cell activation. Journal of Biological Chemistry. 2003;278(37):35651–35659. doi: 10.1074/jbc.M303416200. [DOI] [PubMed] [Google Scholar]
- 93.Chen J, Cohn JA, Mandel LJ. Dephosphorylation of ezrin as an early event in renal microvillar breakdown and anoxic injury. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(16):7495–7499. doi: 10.1073/pnas.92.16.7495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Mariner DJ, Anastasiadis P, Keilhack H, Böhmer FD, Wang J, Reynolds AB. Identification of Src phosphorylation sites in the catenin p120 ctn. Journal of Biological Chemistry. 2001;276(30):28006–28013. doi: 10.1074/jbc.M102443200. [DOI] [PubMed] [Google Scholar]
- 95.Alemà S, Salvatore AM. p120 catenin and phosphorylation: mechanisms and traits of an unresolved issue. Biochimica et Biophysica Acta. 2007;1773(1):47–58. doi: 10.1016/j.bbamcr.2006.06.001. [DOI] [PubMed] [Google Scholar]
- 96.Miravet S, Piedra J, Castaño J, et al. Tyrosine phosphorylation of plakoglobin causes contrary effects on its association with desmosomes and adherens junction components and modulates β-catenin-mediated transcription. Molecular and Cellular Biology. 2003;23(20):7391–7402. doi: 10.1128/MCB.23.20.7391-7402.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Wang Y, Du D, Fang L, et al. Tyrosine phosphorylated Par3 regulates epithelial tight junction assembly promoted by EGFR signaling. EMBO Journal. 2006;25(21):5058–5070. doi: 10.1038/sj.emboj.7601384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Traweger A, Wiggin G, Taylor L, Tate SA, Metalnikov P, Pawson T. Protein phosphatase 1 regulates the phosphorylation state of the polarity scaffold Par-3. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(30):10402–10407. doi: 10.1073/pnas.0804102105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Burks J, Agazie YM. Modulation of α-catenin Tyr phosphorylation by SHP2 positively effects cell transformation induced by the constitutively active FGFR3. Oncogene. 2006;25(54):7166–7179. doi: 10.1038/sj.onc.1209728. [DOI] [PubMed] [Google Scholar]
- 100.Ji H, Wang J, Nika H, et al. EGF-Induced ERK activation promotes CK2-mediated disassociation of α-catenin from β-catenin and transactivation of β-catenin. Molecular Cell. 2009;36(4):547–559. doi: 10.1016/j.molcel.2009.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Nakajo S, Nakaya K, Nakamura Y. Phosphorylation of actin-binding proteins by casein kinases 1 and 2. Biochemistry International. 1987;15(2):321–327. [PubMed] [Google Scholar]
- 102.Zhang Z, Izaguirre G, Lin SY, Lee HY, Schaefer E, Haimovich B. The phosphorylation of vinculin on tyrosine residues 100 and 1065, mediated by Src kinases, affects cell spreading. Molecular Biology of the Cell. 2004;15(9):4234–4247. doi: 10.1091/mbc.E04-03-0264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Han MY, Kosako H, Watanabe T, Hattori S. Extracellular signal-regulated kinase/mitogen-activated protein kinase regulates actin organization and cell motility by phosphorylating the actin cross-linking protein EPLIN. Molecular and Cellular Biology. 2007;27(23):8190–8204. doi: 10.1128/MCB.00661-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Maruoka M, Sato M, Yuan Y, et al. Abi-1-bridged tyrosine phosphorylation of VASP by Abelson kinase impairs association of VASP to focal adhesions and regulates leukaemic cell adhesion. Biochemical Journal. 2012;441(3):889–899. doi: 10.1042/BJ20110951. [DOI] [PubMed] [Google Scholar]
- 105.Thomson DM, Ascione MPA, Grange J, Nelson C, Hansen MDH. Phosphorylation of VASP by AMPK alters actin binding and occurs at a novel site. Biochemical and Biophysical Research Communications. 2011;414(1):215–219. doi: 10.1016/j.bbrc.2011.09.059. [DOI] [PubMed] [Google Scholar]
- 106.Deguchi A, Soh JW, Li H, Pamukcu R, Thompson WJ, Weinstein IB. Vasodilator-stimulated phosphoprotein (VASP) phosphorylation provides a biomarker for the action of exisulind and related agents that activate protein kinase G. Molecular cancer therapeutics. 2002;1(10):803–809. [PubMed] [Google Scholar]
- 107.Walk SF, March ME, Ravichandran KS. Roles of Lck, Syk and ZAP-70 tyrosine kinases in TCR-mediated phosphorylation of the adapter protein Shc. European Journal of Immunology. 1998;28(8):2265–2275. doi: 10.1002/(SICI)1521-4141(199808)28:08<2265::AID-IMMU2265>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- 108.Van Der Geer P, Wiley S, Gish GD, Pawson T. The Shc adaptor protein is highly phosphorylated at conserved, twin tyrosine residues (Y239/240) that mediate protein-protein interactions. Current Biology. 1996;6(11):1435–1444. doi: 10.1016/s0960-9822(96)00748-8. [DOI] [PubMed] [Google Scholar]
- 109.Grønborg M, Kristiansen TZ, Stensballe A, et al. A mass spectrometry-based proteomic approach for identification of serine/threonine-phosphorylated proteins by enrichment with phospho-specific antibodies: identification of a novel protein, Frigg, as a protein kinase A substrate. Molecular & Cellular Proteomics. 2002;1(7):517–527. doi: 10.1074/mcp.m200010-mcp200. [DOI] [PubMed] [Google Scholar]
- 110.Shao H, Wu C, Wells A. Phosphorylation of α-actinin 4 upon epidermal growth factor exposure regulates its interaction with actin. Journal of Biological Chemistry. 2010;285(4):2591–2600. doi: 10.1074/jbc.M109.035790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Sangrar W, Gao Y, Scott M, Truesdell P, Greer PA. Fer-mediated cortactin phosphorylation is associated with efficient fibroblast migration and is dependent on reactive oxygen species generation during integrin-mediated cell adhesion. Molecular and Cellular Biology. 2007;27(17):6140–6152. doi: 10.1128/MCB.01744-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Grassart A, Meas-Yedid V, Dufour A, Olivo-Marin JC, Dautry-Varsat A, Sauvonnet N. Pak1 phosphorylation enhances cortactin-N-WASP interaction in clathrin-caveolin-independent endocytosis. Traffic. 2010;11(8):1079–1091. doi: 10.1111/j.1600-0854.2010.01075.x. [DOI] [PubMed] [Google Scholar]
- 113.Van Delft S, Verkleij AJ, Boonstra J. Epidermal growth factor induces serine phosphorylation of actin. FEBS Letters. 1995;357(3):251–254. doi: 10.1016/0014-5793(94)01359-9. [DOI] [PubMed] [Google Scholar]
- 114.Papakonstanti EA, Stournaras C. Association of PI-3 kinase with PAK1 leads to actin phosphorylation and cytoskeletal reorganization. Molecular Biology of the Cell. 2002;13(8):2946–2962. doi: 10.1091/mbc.02-01-0599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Liu X, Shu S, Hong MSS, Levine RL, Korn ED. Phosphorylation of actin Tyr-53 inhibits filament nucleation and elongation and destabilizes filaments. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(37):13694–13699. doi: 10.1073/pnas.0606321103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Arcaro A, Aubert M, Espinosa del Hierro ME, et al. Critical role for lipid raft-associated Src kinases in activation of PI3K-Akt signalling. Cellular Signalling. 2007;19(5):1081–1092. doi: 10.1016/j.cellsig.2006.12.003. [DOI] [PubMed] [Google Scholar]
- 117.Mitra A, Radha V. F-actin-binding domain of c-Abl regulates localized phosphorylation of C3G: role of C3G in c-Abl-mediated cell death. Oncogene. 2010;29(32):4528–4542. doi: 10.1038/onc.2010.113. [DOI] [PubMed] [Google Scholar]
- 118.Wolf RM, Wilkes JJ, Chao MV, Resh MD. Tyrosine phosphorylation of p190 RhoGAP by Fyn regulates oligodendrocyte differentiation. Journal of Neurobiology. 2001;49(1):62–78. doi: 10.1002/neu.1066. [DOI] [PubMed] [Google Scholar]
- 119.Roof RW, Haskell MD, Dukes BD, Sherman N, Kinter M, Parsons SJ. Phosphotyrosine (p-Tyr)-dependent and -independent mechanisms of p190 RhoGAP-p120 RasGAP interaction: Tyr 1105 of p190, a substrate for c-Src, is the sole p-Tyr mediator of complex formation. Molecular and Cellular Biology. 1998;18(12):7052–7063. doi: 10.1128/mcb.18.12.7052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Taniguchi S, Liu H, Nakazawa T, Yokoyama K, Tezuka T, Yamamoto T. p250GAP, a neural RhoGAP protein, is associated with and phosphorylated by Fyn. Biochemical and Biophysical Research Communications. 2003;306(1):151–155. doi: 10.1016/s0006-291x(03)00923-9. [DOI] [PubMed] [Google Scholar]
- 121.Moon SY, Zang H, Zheng Y. Characterization of a brain-specific Rho GTPase-activating protein, p200RhoGAP. Journal of Biological Chemistry. 2003;278(6):4151–4159. doi: 10.1074/jbc.M207789200. [DOI] [PubMed] [Google Scholar]
- 122.Servitja JM, Marinissen MJ, Sodhi A, Bustelo XR, Gutkind JS. Rac1 function is required for Src-induced transformation: evidence of a role for Tiam1 and Vav2 in Rac activation by Src. Journal of Biological Chemistry. 2003;278(36):34339–34346. doi: 10.1074/jbc.M302960200. [DOI] [PubMed] [Google Scholar]
- 123.Sastry SK, Rajfur Z, Liu BP, Cote JF, Tremblay ML, Burridge K. PTP-PEST couples membrane protrusion and tail retraction via VAV2 and p190RhoGAP. Journal of Biological Chemistry. 2006;281(17):11627–11636. doi: 10.1074/jbc.M600897200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Hubbard SR, Till JH. Protein tyrosine kinase structure and function. Annual Review of Biochemistry. 2000;69:373–398. doi: 10.1146/annurev.biochem.69.1.373. [DOI] [PubMed] [Google Scholar]
- 125.Stoker AW. Protein tyrosine phosphatases and signalling. Journal of Endocrinology. 2005;185(1):19–33. doi: 10.1677/joe.1.06069. [DOI] [PubMed] [Google Scholar]
- 126.Zondag GCM, Moolenaar WH. Receptor protein tyrosine phosphatases: involvement in cell-cell interaction and signaling. Biochimie. 1997;79(8):477–483. doi: 10.1016/s0300-9084(97)82739-3. [DOI] [PubMed] [Google Scholar]
- 127.Dadke S, Kusari A, Kusari J. Phosphorylation and activation of protein tyrosine phosphatase (PTP) 1B by insulin receptor. Molecular and Cellular Biochemistry. 2001;221(1-2):147–154. doi: 10.1023/a:1010909031310. [DOI] [PubMed] [Google Scholar]
- 128.Fujisawa K, Fujita A, Ishizaki T, Saito Y, Narumiya S. Identification of the Rho-binding domain of p160(ROCK), a Rho-associated coiled-coil containing protein kinase. Journal of Biological Chemistry. 1996;271(38):23022–23028. doi: 10.1074/jbc.271.38.23022. [DOI] [PubMed] [Google Scholar]
- 129.Lu W, Mayer BJ. Mechanism of activation of Pak1 kinase by membrane localization. Oncogene. 1999;18(3):797–806. doi: 10.1038/sj.onc.1202361. [DOI] [PubMed] [Google Scholar]
- 130.Rosenzweig T, Aga-Mizrachi S, Bak A, Sampson SR. Src tyrosine kinase regulates insulin-induced activation of protein kinase C (PKC) δ in skeletal muscle. Cellular Signalling. 2004;16(11):1299–1308. doi: 10.1016/j.cellsig.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 131.Yoshida K, Kufe D. Negative regulation of the SHPTP1 protein tyrosine phosphatase by protein kinase C δ in response to DNA damage. Molecular Pharmacology. 2001;60(6):1431–1438. doi: 10.1124/mol.60.6.1431. [DOI] [PubMed] [Google Scholar]
- 132.Somani AK, Bignon JS, Mills GB, Siminovitch KA, Branch DR. Src kinase activity is regulated by the SHP-1 protein-tyrosine phosphatase. Journal of Biological Chemistry. 1997;272(34):21113–21119. doi: 10.1074/jbc.272.34.21113. [DOI] [PubMed] [Google Scholar]
- 133.Obara Y, Labudda K, Dillon TJ, Stork PJS. PKA phosphorylation of Src mediates Rap1 activation in NGF and cAMP signaling in PC12 cells. Journal of Cell Science. 2004;117(25):6085–6094. doi: 10.1242/jcs.01527. [DOI] [PubMed] [Google Scholar]
- 134.Vang T, Torgersen KM, Sundvold V, et al. Activation of the COOH-terminal Src kinase (Csk) by cAMP-dependent protein kinase inhibits signaling through the T cell receptor. Journal of Experimental Medicine. 2001;193(4):497–507. doi: 10.1084/jem.193.4.497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Superti-Furga G, Fumagalli S, Koegl M, Courtneidge SA, Draetta G. Csk inhibition of c-Src activity requires both the SH2 and SH3 domains of Src. EMBO Journal. 1993;12(7):2625–2634. doi: 10.1002/j.1460-2075.1993.tb05923.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.McLachlan RW, Yap AS. Not so simple: the complexity of phosphotyrosine signaling at cadherin adhesive contacts. Journal of Molecular Medicine. 2007;85(6):545–554. doi: 10.1007/s00109-007-0198-x. [DOI] [PubMed] [Google Scholar]
- 137.Nagar B, Hantschel O, Young MA, et al. Structural basis for the autoinhibition of c-Abl tyrosine kinase. Cell. 2003;112(6):859–871. doi: 10.1016/s0092-8674(03)00194-6. [DOI] [PubMed] [Google Scholar]
- 138.El Sayegh TY, Arora PD, Fan L, et al. Phosphorylation of N-cadherin-associated cortactin by Fer kinase regulates N-cadherin mobility and intercellular adhesion strength. Molecular Biology of the Cell. 2005;16(12):5514–5527. doi: 10.1091/mbc.E05-05-0410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Zandy NL, Pendergast AM. Abl tyrosine kinases modulate cadherin-dependent adhesion upstream and downstream of Rho family GTPases. Cell Cycle. 2008;7(4):444–448. doi: 10.4161/cc.7.4.5452. [DOI] [PubMed] [Google Scholar]
- 140.Joo CK, Kim HS, Park JY, Seomun Y, Son MJ, Kim JT. Ligand release-independent transactivation of epidermal growth factor receptor by transforming growth factor-β involves multiple signaling pathways. Oncogene. 2008;27(5):614–628. doi: 10.1038/sj.onc.1210649. [DOI] [PubMed] [Google Scholar]
- 141.Watanabe T, Sato K, Kaibuchi K. Cadherin-mediated intercellular adhesion and signaling cascades involving small GTPases. Cold Spring Harbor Perspectives in Biology. 2009;1(3):p. a003020. doi: 10.1101/cshperspect.a003020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Braga VMM, Yap AS. The challenges of abundance: epithelial junctions and small GTPase signalling. Current Opinion in Cell Biology. 2005;17(5):466–474. doi: 10.1016/j.ceb.2005.08.012. [DOI] [PubMed] [Google Scholar]
- 143.Burton EA, Oliver TN, Pendergast AM. Abl kinases regulate actin comet tail elongation via an N-WASP-dependent pathway. Molecular and Cellular Biology. 2005;25(20):8834–8843. doi: 10.1128/MCB.25.20.8834-8843.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Martin M, Ahern-Djamali SM, Hoffmann FM, Saxton WM. Abl tyrosine kinase and its substrate Ena/VASP have functional interactions with kinesin-1. Molecular Biology of the Cell. 2005;16(9):4225–4230. doi: 10.1091/mbc.E05-02-0116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Lu Q, Mukhopadhyay NK, Griffin JD, Paredes M, Medina M, Kosik KS. Brain armadillo protein δ-catenin interacts with Abl tyrosine kinase and modulates cellular morphogenesis in response to growth factors. Journal of Neuroscience Research. 2002;67(5):618–624. doi: 10.1002/jnr.10151. [DOI] [PubMed] [Google Scholar]
- 146.Dai Z, Pendergast AM. Abi-2, a novel SH3-containing protein interacts with the c-Abl tyrosine kinase and modulates c-Abl transforming activity. Genes and Development. 1995;9(21):2569–2582. doi: 10.1101/gad.9.21.2569. [DOI] [PubMed] [Google Scholar]
- 147.Mitsushima M, Takahashi H, Shishido T, Ueda K, Kioka N. Abl kinase interacts with and phosphorylates vinexin. FEBS Letters. 2006;580(17):4288–4295. doi: 10.1016/j.febslet.2006.06.072. [DOI] [PubMed] [Google Scholar]
- 148.McCrea PD, Gu D, Balda MS. Junctional music that the nucleus hears: cell-cell contact signaling and the modulation of gene activity. Cold Spring Harbor Perspectives in Biology. 2009;1(4):p. a002923. doi: 10.1101/cshperspect.a002923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Brembeck FH, Rosário M, Birchmeier W. Balancing cell adhesion and Wnt signaling, the key role of β-catenin. Current Opinion in Genetics and Development. 2006;16(1):51–59. doi: 10.1016/j.gde.2005.12.007. [DOI] [PubMed] [Google Scholar]
- 150.Nelson WJ, Nusse R. Convergence of Wnt, β-Catenin, and Cadherin pathways. Science. 2004;303(5663):1483–1487. doi: 10.1126/science.1094291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Xu Y, Carpenter G. Identification of cadherin tyrosine residues that are phosphorylated and mediate Shc association. Journal of Cellular Biochemistry. 1999;75(2):264–271. doi: 10.1002/(sici)1097-4644(19991101)75:2<264::aid-jcb9>3.3.co;2-2. [DOI] [PubMed] [Google Scholar]
- 152.Wang Z, Sandiford S, Wu C, Li SSC. Numb regulates cell-cell adhesion and polarity in response to tyrosine kinase signalling. EMBO Journal. 2009;28(16):2360–2373. doi: 10.1038/emboj.2009.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Potter MD, Barbero S, Cheresh DA. Tyrosine phosphorylation of VE-cadherin prevents binding of p120- and β-catenin and maintains the cellular mesenchymal state. Journal of Biological Chemistry. 2005;280(36):31906–31912. doi: 10.1074/jbc.M505568200. [DOI] [PubMed] [Google Scholar]
- 154.Yonemura S, Matsui T, Tsukita S, Tsukita S. Rho-dependent and -indenpendent activation mechanisms of ezrin/radixin/moesin proteins: an essential role for polyphosphoinositides in vivo. Journal of Cell Science. 2002;115(12):2569–2580. doi: 10.1242/jcs.115.12.2569. [DOI] [PubMed] [Google Scholar]
- 155.Amano M, Ito M, Kimura K, et al. Phosphorylation and activation of myosin by Rho-associated kinase (Rho- kinase) Journal of Biological Chemistry. 1996;271(34):20246–20249. doi: 10.1074/jbc.271.34.20246. [DOI] [PubMed] [Google Scholar]
- 156.Henson JH, Kolnik SE, Fried CA, et al. Actin-based centripetal flow: phosphatase inhibition by calyculin-A alters flow pattern, actin organization, and actomyosin distribution. Cell Motility and the Cytoskeleton. 2003;56(4):252–266. doi: 10.1002/cm.10149. [DOI] [PubMed] [Google Scholar]
- 157.Volberg T, Zick Y, Dror R, et al. The effect of tyrosine-specific protein phosphorylation on the assembly of adherens-type junctions. EMBO Journal. 1992;11(5):1733–1742. doi: 10.1002/j.1460-2075.1992.tb05225.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Calautti E, Cabodi S, Stein PL, Hatzfeld M, Kedersha N, Dotto GP. Tyrosine phosphorylation and Src family kinases control keratinocyte cell-cell adhesion. Journal of Cell Biology. 1998;141(6):1449–1465. doi: 10.1083/jcb.141.6.1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Pang JH, Kraemer A, Stehbens SJ, Frame MC, Yap AS. Recruitment of phosphoinositide 3-kinase defines a positive contribution of tyrosine kinase signaling to E-cadherin function. Journal of Biological Chemistry. 2005;280(4):3043–3050. doi: 10.1074/jbc.M412148200. [DOI] [PubMed] [Google Scholar]
- 160.Shibamoto S, Hayakawa M, Takeuchi K, et al. Tyrosine phosphorylation of β-catenin and plakoglobin enhanced by hepatocyte growth factor and epidermal growth factor in human carcinoma cells. Cell Adhesion and Communication. 1994;1(4):295–305. doi: 10.3109/15419069409097261. [DOI] [PubMed] [Google Scholar]
- 161.Hatanaka K, Simons M, Murakami M. Phosphorylation of VE-cadherin controls endothelial phenotypes via p120-catenin coupling and Rac1 activation. American Journal of Physiology. 2011;300(1):H162–H172. doi: 10.1152/ajpheart.00650.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Pece S, Gutkind JS. E-cadherin and Hakai: signalling, remodeling or destruction? Nature Cell Biology. 2002;4(4):E72–E74. doi: 10.1038/ncb0402-e72. [DOI] [PubMed] [Google Scholar]
- 163.Fujita Y, Krause G, Scheffner M, et al. Hakai, a c-Cbl-like protein, ubiquitinates and induces endocytosis of the E-cadherin complex. Nature Cell Biology. 2002;4(3):222–231. doi: 10.1038/ncb758. [DOI] [PubMed] [Google Scholar]
- 164.Keilhack H, Hellman U, Van Hengel J, Van Roy F, Godovac-Zimmermann J, Böhmer FD. The protein-tyrosine phosphatase SHP-1 binds to and dephosphorylates p120 catenin. Journal of Biological Chemistry. 2000;275(34):26376–26384. doi: 10.1074/jbc.M001315200. [DOI] [PubMed] [Google Scholar]
- 165.Fukumoto Y, Shintani Y, Reynolds AB, Johnson KR, Wheelock MJ. The regulatory or phosphorylation domain of p120 catenin controls E-cadherin dynamics at the plasma membrane. Experimental Cell Research. 2008;314(1):52–67. doi: 10.1016/j.yexcr.2007.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Call GS, Chung JY, Davis JA, et al. Zyxin phosphorylation at serine 142 modulates the zyxin head-tail interaction to alter cell-cell adhesion. Biochemical and Biophysical Research Communications. 2011;404(3):780–784. doi: 10.1016/j.bbrc.2010.12.058. [DOI] [PubMed] [Google Scholar]
- 167.Nguyen TN, Uemura A, Shih W, Yamada S. Zyxin-mediated actin assembly is required for efficient wound closure. Journal of Biological Chemistry. 2010;285(46):35439–35445. doi: 10.1074/jbc.M110.119487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Hansen MDH, Beckerle MC. Opposing roles of Zyxin/LPP ACTA repeats and the LIM domain region in cell-cell adhesion. Journal of Biological Chemistry. 2006;281(23):16178–16188. doi: 10.1074/jbc.M512771200. [DOI] [PubMed] [Google Scholar]
- 169.Butt E, Abel K, Krieger M, et al. cAMP- and cGMP-dependent protein kinase phosphorylation sites of the focal adhesion vasodilator-stimulated phosphoprotein (VASP) in vitro and in intact human platelets. Journal of Biological Chemistry. 1994;269(20):14509–14517. [PubMed] [Google Scholar]
- 170.Chitaley K, Chen L, Galler A, Walter U, Daum G, Clowes AW. Vasodilator-stimulated phosphoprotein is a substrate for protein kinase C. FEBS Letters. 2004;556(1-3):211–215. doi: 10.1016/s0014-5793(03)01435-2. [DOI] [PubMed] [Google Scholar]
- 171.Harbeck B, Hüttelmaier S, Schlüter K, Jockusch BM, Illenberger S. Phosphorylation of the vasodilator-stimulated phosphoprotein regulates its interaction witn actin. Journal of Biological Chemistry. 2000;275(40):30817–30825. doi: 10.1074/jbc.M005066200. [DOI] [PubMed] [Google Scholar]
- 172.Barzik M, Kotova TI, Higgs HN, et al. Ena/VASP proteins enhance actin polymerization in the presence of barbed end capping proteins. Journal of Biological Chemistry. 2005;280(31):28653–28662. doi: 10.1074/jbc.M503957200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Vasioukhin V, Bauer C, Yin M, Fuchs E. Directed actin polymerization is the driving force for epithelial cell- cell adhesion. Cell. 2000;100(2):209–219. doi: 10.1016/s0092-8674(00)81559-7. [DOI] [PubMed] [Google Scholar]
- 174.Scott JA, Shewan AM, Den Elzen NR, Loureiro JJ, Gertler FB, Yap AS. Ena/VASP proteins can regulate distinct modes of actin organization at cadherin-adhesive contacts. Molecular Biology of the Cell. 2006;17(3):1085–1095. doi: 10.1091/mbc.E05-07-0644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Morais-de-Sá E, Mirouse V, St Johnston D. aPKC phosphorylation of bazooka defines the apical/lateral border in Drosophila epithelial cells. Cell. 2010;141(3):509–523. doi: 10.1016/j.cell.2010.02.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Walther RF, Pichaud F. Crumbs/DaPKC-dependent apical exclusion of bazooka promotes photoreceptor polarity remodeling. Current Biology. 2010;20(12):1065–1074. doi: 10.1016/j.cub.2010.04.049. [DOI] [PubMed] [Google Scholar]
- 177.Wang Y-C, Khan Z, Kaschube M, Wieschaus EF. Differential positioning of adherens junctions is associated with initiation of epithelial folding. Nature. 2012;484(7394):390–393. doi: 10.1038/nature10938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Hornbeck PV, Chabra I, Kornhauser JM, Skrzypek E, Zhang B. PhosphoSite: a bioinformatics resource dedicated to physiological protein phosphorylation. Proteomics. 2004;4(6):1551–1561. doi: 10.1002/pmic.200300772. [DOI] [PubMed] [Google Scholar]
- 179.Jones AME, Nühse TS. Phosphoproteomics using iTRAQ. Methods in Molecular Biology. 2011;779:287–302. doi: 10.1007/978-1-61779-264-9_17. [DOI] [PubMed] [Google Scholar]
- 180.Eyrich B, Sickmann A, Zahedi RP. Catch me if you can: mass spectrometry-based phosphoproteomics and quantification strategies. Proteomics. 2011;11(4):554–570. doi: 10.1002/pmic.201000489. [DOI] [PubMed] [Google Scholar]
- 181.Imamura H, Yachie N, Saito R, Ishihama Y, Tomita M. Towards the systematic discovery of signal transduction networks using phosphorylation dynamics data. BMC Bioinformatics. 2010;11, article 232 doi: 10.1186/1471-2105-11-232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Martinez-Ferrando I, Chaerkady R, Zhong J, et al. Identification of targets of c-Src tyrosine kinase by chemical complementation and phosphoproteomics. doi: 10.1074/mcp.M111.015750. Molecular and Cellular Proteomics. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Hilger M, Bonaldi T, Gnad F, Mann M. Systems-wide analysis of a phosphatase knock-down by quantitative proteomics and phosphoproteomics. Molecular and Cellular Proteomics. 2009;8(8):1908–1920. doi: 10.1074/mcp.M800559-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Huang PH, Mukasa A, Bonavia R, et al. Quantitative analysis of EGFRvIII cellular signaling networks reveals a combinatorial therapeutic strategy for glioblastoma. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(31):12867–12872. doi: 10.1073/pnas.0705158104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Kozuka-Hata H, Tasaki S, Oyama M. Phosphoproteomics-based systems analysis of signal transduction networks. Frontiers in Physiology. 2011;2(113) doi: 10.3389/fphys.2011.00113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Derouiche A, Cousin C, Mijakovic I. Protein phosphorylation from the perspective of systems biology. doi: 10.1016/j.copbio.2011.11.008. Current Opinion in Biotechnology. In press. [DOI] [PubMed] [Google Scholar]
- 187.Sadot E, Conacci-Sorrell M, Zhurinsky J, et al. Regulation of S33/S37 phosphorylated β-catenin in normal and transformed cells. Journal of Cell Science. 2002;115(13):2771–2780. doi: 10.1242/jcs.115.13.2771. [DOI] [PubMed] [Google Scholar]
- 188.Brown MV, Burnett PE, Denning MF, Reynolds AB. PDGF receptor activation induces p120-catenin phosphorylation at serine 879 via a PKCα-dependent pathway. Experimental Cell Research. 2009;315(1):39–49. doi: 10.1016/j.yexcr.2008.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Ren G, Helwani FM, Verma S, McLachlan RW, Weed SA, Yap AS. Cortactin is a functional target of E-cadherin-activated Src family kinases in MCF7 epithelial monolayers. Journal of Biological Chemistry. 2009;284(28):18913–18922. doi: 10.1074/jbc.M109.000307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Benz PM, Blume C, Seifert S, et al. Differential VASP phosphorylation controls remodeling of the actin cytoskeleton. Journal of Cell Science. 2009;122(21):3954–3965. doi: 10.1242/jcs.044537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Zaidel-Bar R, Milo R, Kam Z, Geiger B. A paxillin tyrosine phosphorylation switch regulates the assembly and form of cell-matrix adhesions. Journal of Cell Science. 2007;120(1):137–148. doi: 10.1242/jcs.03314. [DOI] [PubMed] [Google Scholar]
- 192.Zaidel-Bar R, Geiger B. The switchable integrin adhesome. Journal of Cell Science. 2010;123(9):1385–1388. doi: 10.1242/jcs.066183. [DOI] [PMC free article] [PubMed] [Google Scholar]