

Abstract
The principal function of vitamin D in calcium homeostasis is to increase calcium absorption from the intestine. Calcium is absorbed by both an active transcellular pathway, which is energy dependent, and by a passive paracellular pathway through tight junctions. 1,25Dihydroxyvitamin D3 (1,25(OH)2D3) the hormonally active form of vitamin D, through its genomic actions, is the major stimulator of active intestinal calcium absorption which involves calcium influx, translocation of calcium through the interior of the enterocyte and basolateral extrusion of calcium by the intestinal plasma membrane pump. This article reviews recent studies that have challenged the traditional model of vitamin D mediated transcellular calcium absorption and the crucial role of specific calcium transport proteins in intestinal calcium absorption. There is also increasing evidence that 1,25(OH)2D3 can enhance paracellular calcium diffusion. The influence of estrogen, prolactin, glucocorticoids and aging on intestinal calcium absorption and the role of the distal intestine in vitamin D mediated intestinal calcium absorption are also discussed.
Introduction
Studies in vitamin D receptor (VDR) null mice have indicated that the principal function of vitamin D in mineral homeostasis is to increase calcium absorption from the intestine (Li et al., 1997, Yoshizawa et al., 1997). This conclusion was made based on the findings that rickets and osteomalacia are prevented and serum calcium and parathyroid hormone (PTH) are normalized when VDR null mice are fed a rescue diet high in calcium and lactose. In the event of decreased calcium levels in the serum due to low dietary intake or increased demand of calcium due to growth, pregnancy or lactation, the synthesis of the hormonally active form of vitamin D, 1,25-dihydroxyvitamin D3 (1,25(OH)2D3) is increased leading to increased intestinal calcium absorption. If normal serum calcium is unable to be maintained by intestinal absorption, then 1,25(OH)2D3 together with PTH will mobilize bone calcium and increase reabsorption of calcium from the renal distal tubule.
1,25(OH)2D3 and Transcellular Calcium Absorption
In the intestine 1,25(OH)2D3 affects the process of transcellular calcium transport which has been proposed to involve entry of calcium via the apical calcium channel, transient receptor potential vanilloid type 6 (TRPV6), translocation of calcium through the interior of the enterocyte (it has been suggested that the calcium binding protein calbindin-D9k acts to facilitate calcium diffusion through the cell) and basolateral extrusion of calcium by the intestinal plasma membrane pump PMCA1b (Wasserman 2005) (Fig. 1). Previous studies provided indirect evidence for a role of TRPV6 and calbindin in intestinal calcium absorption. Both are co-localized in the intestine, they are both similarly regulated [induced at weaning (the time of onset of active intestinal calcium transport), under conditions of low dietary calcium and after 1,25(OH)2D3 injection] (Song et al., 2003). TRPV6 and calbindin-D9k are both induced prior to the peak of intestinal calcium absorption (Song et al., 2003). In addition, in vitamin D receptor (VDR) KO mice, where the major defect that results in rickets is the decrease in intestinal calcium absorption, there is a 50% reduction in intestinal calbindin-D9k mRNA and a more marked decrease in TRPV6 mRNA (less than 5 – 10% of the levels in wild type (WT) mice) (Li et al., 1998, Van Cromphant et al., 2001). However, recent studies using calbindin-D9k knock out (KO) mice and TRPV6 KO mice have challenged the traditional model of vitamin D mediated transcellular calcium absorption in the intestine (Kutuzova et al., 2006, Akhter et al., 2007, Benn et al., 2008, Kutuzova et al., 2008, Bianco et al., 2008). Calbindin-D9k KO mice show no difference in phenotype compared to WT mice and they maintain normal serum calcium (Kutuzova et al., 2006, Akhter et al., 2007, Benn et al., 2008). Active intestinal calcium transport is similarly induced in both calbindin-D9k KO mice and WT mice in response to a low calcium diet or 1,25(OH)2D3, suggesting that calbindin-D9k may be compensated by another calcium binding protein (Benn et al., 2008). Calbindin may have another role in the intestine. Calbindin-D9k may be a modulator of the activity of TRPV6 and/or calbindin may act as a cytosolic buffer to prevent toxic levels of calcium from accumulating in the intestinal cell when there is an increase in apical calcium entry. TRPV6 KO mice also maintain normal serum calcium levels (Benn et al., 2008, Kutuzova et al., 2008). In response to 1,25(OH)2D3 intestinal calcium transport is similar in WT and TRPV6 KO mice (Benn et al., 2008, Kutuzova et al., 2008). However, under conditions of low dietary calcium, compared to WT mice, intestinal calcium transport is less efficient in the TRPV6 null mice (Benn et al., 2008). Also, in response to restriction of dietary calcium, although impaired bone mass accrual is comparable in WT and TRPV6 null mice, there is an increase in osteoblast activity and osteoid abundance is more pronounced in the TRPV6 null mice, suggesting a role for TRPV6 under low calcium conditions in intestinal calcium absorption to maintain proper bone mineralization (Lieben et al., 2010). When dietary supply of calcium is normal, lack of TRPV6 did not affect bone mass, remodeling parameters or growth plate morphology in young adult or aging mice. Thus, under normal calcium intake, TRPV6 is redundant for intestinal calcium transport, suggesting compensation by another channel or protein. Although the phenotype is mild, studies in the TRPV6 null mutant mice suggest that TRPV6 does contribute to the calcium absorptive process during calcium restriction. Recent studies showed that TRPV6 can interact with other proteins including calmodulin, which facilitates rapid inactivation of TRPV6, S100A10-annexin 2 protein complex which maybe involved in constitutive trafficking of TPRV6 to the plasma membrane and Rab11a which plays a role in recycling of TPRV6 to the plasma membrane (Derler et al., 2006, Van de Graaf et 2003, Van de Graaf et al, 2006). These TRPV6 associated proteins may represent novel components of the regulation by 1,25(OH)2D3 of calcium entry into the intestinal cell. Recent preliminary results show that transgenic mice overexpressing TRPV6 throughout the intestine (from duodenum to distal colon) of WT mice develop hypercalcemia, hypercalciuria and soft tissue calcification (Cui and Fleet, 2010). Although TRPV6 is not critical for 1,25(OH)2D3 mediated intestinal calcium absorption and maybe compensated by another channel yet to be identified, these studies indicate that TRPV6 does have a role in the process of intestinal calcium absorption.
Figure 1.
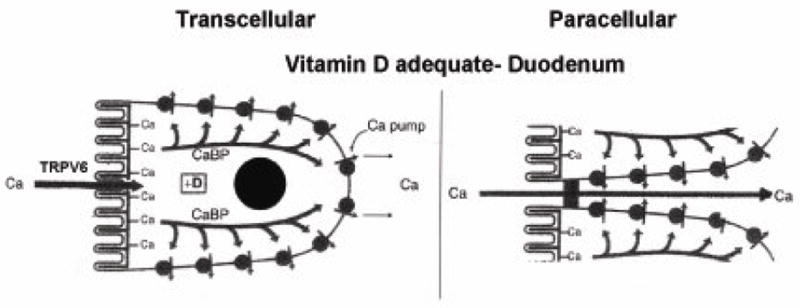
Models of vitamin D mediated intestinal calcium absorption. Left panel: Transcellular intestinal calcium absorption. 1,25(OH)2D3, through its genomic actions, stimulates active intestinal calcium absorption. The traditional model of transcellular calcium transport involves calcium influx through TRPV6, intracellular calcium transfer by calbindin (CaBP) and calcium extrusion by the plasma membrane calcium ATPase (Ca pump). Recent studies using KO mice have suggested that TRPV6 and calbindin are not critical for 1,25(OH)2D3 calcium absorption and maybe compensated by another channel or protein. Right panel: Paracellular pathway. There is increasing evidence that 1,25(OH)2D3 can enhance paracellular calcium diffusion by regulating tight junction proteins. (Reproduced with permission from Wasserman RH, 2005).
Paracellular Calcium Transport
Calcium transverses the intestine by both the active transcellular pathway and the passive paracellular pathway (Hoenderop et al., 2005) (Fig. 1). The paracellular pathway functions throughout the entire length of the intestine and predominates in the more distal regions. This pathway is driven by the luminal electrochemical gradient and the integrity of the intercellular tight junctions (Tuskita et al., 2001). Recent evidence suggests that transjunctional transport of calcium by the paracellular pathway occurs in a regulated fashion, and may be coupled to active transcellular movement of calcium in a coordinated manner (Fujita et al., 2008). Tight junctions are specialized membrane domains located between the apical and basolateral membranes of the enterocyte, which form a barrier to the movement of ions, proteins, and other macromolecules across the intestine by maintaining a charge and size selectivity (Tuskita et al., 2001). Claudins are the major transmembrane components of tight junctions (Tuskita et al., 2001, Furuse et al., 1998) and it has been suggested that 1,25(OH)2D3 can promote paracellular calcium diffusion by increasing junction ion permeability (Fujita et al., 2008, Kutuzova and DeLuca 2004). VDR knockout mice, known to exhibit reduced intestinal calcium absorption, have decreased levels of claudin-2 and claudin-12 mRNA and protein (Fujita et al., 2008). In addition, 1,25(OH)2D3 has been shown to induce the expression of claudin-2 and claudin-12 in vitro in an intestinal epithelial cell line resulting in facilitated paracellular calcium conductance (Fujita et al., 2008). Gene array studies have shown that in addition to the known target genes, TRPV6 and calbindin-D9K, 1,25(OH)2D3 suppresses a number of intra- and intercellular matrix proteins including cadherin-17 (a cell adhesion protein) and aquaporin-8 (a tight junction channel), suggesting that vitamin D regulates intestinal calcium absorption via the paracellular pathway in addition to the transcellular pathway (Kutuzova and DeLuca 2004). It is possible that these other factors may compensate for the lack of TRPV6 in the TRPV6 KO mouse and thus may explain, in part, the mild phenotype of the TPRV6 KO mouse and the TRPV6/calbindin-D9k double KO mouse. Future studies examining different regions of the intestine and the identification of novel 1,25(OH)2D3 regulated proteins involved in transcellular and paracellular calcium absorption are needed.
Pregnancy, Lactation, Estrogen and Prolactin and Intestinal Calcium Absorption
Although 1,25(OH)2D3 is the principal hormone regulating active intestinal calcium absorption, other hormones have been shown to influence the process as well. Increased intestinal calcium transport has been observed in vitamin D deficient pregnant and lactating rats (Halloron and Deluca 1980, Boass et al., 1981, Brommage et al 1990). In addition, estradiol replacement in ovariectomized rats has been reported to result in an increase in intestinal calcium absorption without a stimulation of circulating 1,25(OH)2D3 levels (O’Loughlin and Morris, 1998). Studies by Van Cromphaut et al (Van Cromphant et al., 2003) in VDR KO mice showed that estrogen treatment after ovariectomy as well as pregnancy and lactation result in an induction of TRPV6 in the duodenum. In addition, in estrogen receptor α (ERα) KO mice, TRPV6 is reduced (Van Cromphant et al., 2003). These findings suggest that estrogens, independent of vitamin D, may be important regulators of calcium influx into the enterocyte and that these effects are mediated by ERα. In addition to estrogen, prolactin, a lacotgenic polypeptide hormone that is elevated during pregnancy and lactation, has been shown to have calcium regulatory effects. Prolactin has been reported to stimulate active intestinal calcium transport in vitamin D deficient rats (Pahuja and Deluca 1981). In addition, a direct effect of prolactin on active duodenal calcium transport was shown in studies using the Ussing chamber technique and prolactin applied to the incubation solution (Charoenphandhu et al., 2001). Studies in our laboratory indicate that prolactin can regulate TRPV6 in the duodenum independent of vitamin D, and also has cooperative effects with 1,25(OH)2D3 in regulation of both the intestinal calcium transport proteins TRPV6 and calbindin-D9K (Ajibade et al., 2010). Because both 1,25(OH)2D3 and prolactin levels are elevated during lactation (Halloron and Deluca 1980, Halloron et al. 1979, Pike et al., 1979, Meites et al 1972), prolactin may act together with 1,25(OH)2D3 to increase active intestinal calcium absorption. In addition, we found that prolactin has a direct effect on the transcription of the 1α(OH)ase gene, thus enhancing 1α(OH)ase protein expression and increasing levels of 1,25(OH)2D3 during lactation when there is an increased calcium requirement for the neonate (Ajibade et al., 2010). It has been suggested that prolactin also has an effect on the paracellular pathway of intestinal calcium absorption through upregulation of claudin-15 (Charoenphandhu et al., 2009).
Glucocortiocoids and Intestinal Calcium Absorption
Because of their potent anti-inflammatory and immunosuppressive properties, glucocorticoids are effectively used to treat inflammatory conditions such as asthma and rheumatoid arthritis. However, long term treatment with glucocorticoids reduces bone mineral density leading to osteoporosis (Reid 1997). In addition to direct effects on bone, glucocorticoids can also induce bone loss through diminished intestinal calcium absorption (Reid 1997, Huybers et al., 2007). In mice it has been reported that treatment with pharmacological doses of glucocorticoids results in decreased intestinal calcium absorption which is associated with a decrease in TRPV6 and calbindin-D9K (Huybers et al., 2007, Lee et al., 2006). Studies in cortisol treated chicks also noted an inhibition of intestinal calcium absorption which was associated with a decrease in calbindin (Feher and Wasserman, 1979). It has been suggested that the effect of glucocorticoids on calcium transport proteins can be independent of vitamin D (Huybers et al., 2007, Feher and Wasserman, 1979).
Effect of Aging
In aging, intestinal calcium absorption declines resulting in increased PTH which correlates to an age-related increase in bone turnover (Bullamore et al., 1970, Ledger et al., 1995). It has been proposed that the defect in intestinal calcium absorption is related both to low circulating levels of 1,25(OH)2D3 and to intestinal resistance to the action of 1,25(OH)2D3 (Wood et al., 1998). Either no change or a small decrease in intestinal VDR number has been reported with aging (Wood et al., 1998, Halloran and Portale, 2005). The expression of TRPV6 and calbindin-D9k declines with age and this decline is correlated to the decrease in intestinal calcium absorption and serum 1,25(OH)2D3 (Brown et al., 2005). In mice deficient in klotho (a multifunctional protein involved in phosphate and calcium homeostasis) a premature aging phenotype has been described (including short lifespan, infertility, atherosclerosis, skin atrophy and osteoporosis) (Kuro-o et al., 1997). Klotho functions as an essential cofactor for FGF23 which has been shown to regulate phosphate homeostasis and vitamin D biosynthesis (Kurosu et al). FGF23 and klotho, the FGF cofactor, suppress the expression of 1α(OH)ase and induce 24(OH)ase in kidney (Tsujikawa et al., 2003). Compared to WT mice, klotho KO mice have increased serum calcium and phosphate and decreased serum PTH (Kuro-o et al., 1997). Klotho KO mice also display increased serum levels of 1,25(OH)2D3 and increased expression of 1α(OH)ase (Tsujikawa et al., 2003, Yoshida et al., 2002). In the intestine klotho KO mice show increased expression of TRPV6 and calbindin-D9k and increased intestinal calcium absorption (Alexander et al., 2009) (in contrast to the decline observed in aging mice and rats; Brown et al., 2005). Thus the “aging” phenotype of the klotho KO mouse may reflect, at least in part, the effect of overproduction of 1,25(OH)2D3 rather than an effect on intestinal calcium absorption which contributes to bone loss in aging.
Calcium absorption, vitamin D and the distal intestine
In the calcium absorptive process the duodenum has been the major focus of research due to its highly active transport system. However, it is the distal intestine where 70- 80 % of the ingested calcium is absorbed (mostly in the ileum) (Wasserman 2005). Thus, it is important to understand the process by which the distal segment transports calcium. The vitamin D dependence of calcium absorption in the ileum and the colon has been shown (Petith et al., 1979, Lee et al., 1981, Favus et al., 1980, Favus, 1985, Vergne-Marini et al., 1976). VDR is expressed in all segments of the small and large intestine (highest levels have been reported in the cecum and colon) and in patients with extensive resection of the small intestine, calcium absorption has been reported to be significantly higher when the colon is preserved (Hirst and Feldman 1981, Stumpf et al., 1979, Xue and Fleet 2009, Hylander et al., 1990). 1,25(OH)2D3 has been shown to convert regions of net secretion of calcium in ileum and colon to net absorption (Favus 1985). The cecum has also been reported to be involved in 1,25(OH)2D3 mediated active transport (Favus and Angeid-Backman 1985). TRPV6 protein and calbindin-D9k protein are present in all segments of the mouse and rat intestine (immunocytochemical studies have indicated that the strongest expression of TRPV6 is in the cecum and colon) (Teerapornpuntakit et al., 2009, Zhang et al., 2010). Together these findings indicate that the distal segments of the intestine, in addition to the duodenum, play an important role in 1,25(OH)2D3 mediated calcium homeostasis. Our studies in the TRPV6/calbindin-D9k double KO mice indicate that we are missing important information related to the mechanisms involved in the regulation of intestinal calcium transport by 1,25(OH)2D3. Identification of multiple mechanisms by which 1,25(OH)2D3 acts to increase calcium absorption in different segments of the intestine is needed in order to identify new approaches to sustain calcium balance.
Acknowledgments
This work was supported by National Institutes of Health Grant DK38961-22 to SC
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ajibade DV, Dhawan P, Fechner AJ, Meyer MB, Pike JW, Christakos S. Evidence for a role of prolactin in calcium homeostasis: regulation of intestinal transient receptor potential vanilloid type 6, intestinal calcium absorption, and the 25-hydroxyvitamin D(3) 1alpha hydroxylase gene by prolactin. Endocrinology. 2010;151:2974–2984. doi: 10.1210/en.2010-0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akhter S, Kutuzova GD, Christakos S, DeLuca HF. Calbindin D9k is not required for 1,25-dihydroxyvitamin D3-mediated Ca2+ absorption in small intestine. Arch Biochem Biophys. 2007;460:227–232. doi: 10.1016/j.abb.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Alexander RT, Woudenberg-Vrenken TE, Buurman J, Dijkman H, van der Eerden BC, van Leeuwen JP, Bindels RJ, Hoenderop JG. Klotho prevents renal calcium loss. J Am Soc Nephrol. 2009;20:2371–2379. doi: 10.1681/ASN.2008121273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amling M, Priemel M, Holzmann T, Chapin K, Rueger JM, Baron R, Demay MB. Rescue of the skeletal phenotype of vitamin D receptor-ablated mice in the setting of normal mineral ion homeostasis: formal histomorphometric and biomechanical analyses. Endocrinology. 1999;140:4982–4987. doi: 10.1210/endo.140.11.7110. [DOI] [PubMed] [Google Scholar]
- Benn BS, Ajibade D, Porta A, Dhawan P, Hediger M, Peng JB, Jiang Y, Oh GT, Jeung EB, Lieben L, Bouillon R, Carmeliet G, Christakos S. Active intestinal calcium transport in the absence of transient receptor potential vanilloid type 6 and calbindin-D9k. Endocrinology. 2008;149:3196–3205. doi: 10.1210/en.2007-1655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianco SD, Peng JB, Takanaga H, Suzuki Y, Crescenzi A, Kos CH, Zhuang L, Freeman MR, Gouveia CH, Wu J, Luo H, Mauro T, Brown EM, Hediger MA. Marked disturbance of calcium homeostasis in mice with targeted disruption of the Trpv6 calcium channel gene. J Bone Miner Res. 2007;22:274–285. doi: 10.1359/jbmr.061110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boass A, Toverud SU, Pike JW, Haussler MR. Calcium metabolism during lactation: enhanced intestinal calcium absorption in vitamin D-deprived, hypocalcemic rats. Endocrinology. 1981;109:900–907. doi: 10.1210/endo-109-3-900. [DOI] [PubMed] [Google Scholar]
- Brommage R, Baxter DC, Gierke LW. Vitamin D-independent intestinal calcium and phosphorus absorption during reproduction. Am J Physiol. 1990;259:G631–638. doi: 10.1152/ajpgi.1990.259.4.G631. [DOI] [PubMed] [Google Scholar]
- Brown AJ, Krits I, Armbrecht HJ. Effect of age, vitamin D, and calcium on the regulation of rat intestinal epithelial calcium channels. Arch Biochem Biophys. 2005;437:51–58. doi: 10.1016/j.abb.2005.02.007. [DOI] [PubMed] [Google Scholar]
- Bullamore JR, Wilkinson R, Gallagher JC, Nordin BE, Marshall DH. Effect of age on calcium absorption. Lancet. 1970;2:535–537. doi: 10.1016/s0140-6736(70)91344-9. [DOI] [PubMed] [Google Scholar]
- Charoenphandhu N, Limlomwongse L, Krishnamra N. Prolactin directly stimulates transcellular active calcium transport in the duodenum of female rats. Can J Physiol Pharmacol. 2001;79:430–438. [PubMed] [Google Scholar]
- Charoenphandhu N, Nakkrasae LI, Kraidith K, Teerapornpuntakit J, Thongchote K, Thongon N, Krishnamra N. Two-step stimulation of intestinal Ca(2+) absorption during lactation by long-term prolactin exposure and suckling-induced prolactin surge. Am J Physiol Endocrinol Metab. 2009;297:E609–619. doi: 10.1152/ajpendo.00347.2009. [DOI] [PubMed] [Google Scholar]
- Cui M, F J. Transgenic overexpression of human TRPV6 in intestine increases calcium absorption efficiency and improves bone mass in mice. Journal of Bone and Mineral Research. 2010;25:S59. [Google Scholar]
- Derler I, Hofbauer M, Kahr H, Fritsch R, Muik M, Kepplinger K, Hack ME, Moritz S, Schindl R, Groschner K, Romanin C. Dynamic but not constitutive association of calmodulin with rat TRPV6 channels enables fine tuning of Ca2+-dependent inactivation. J Physiol. 2006;577:31–44. doi: 10.1113/jphysiol.2006.118661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Favus MJ. Factors that influence absorption and secretion of calcium in the small intestine and colon. Am J Physiol. 1985;248:G147–157. doi: 10.1152/ajpgi.1985.248.2.G147. [DOI] [PubMed] [Google Scholar]
- Favus MJ, Angeid-Backman E. Effects of 1,25(OH)2D3 and calcium channel blockers on cecal calcium transport in the rat. Am J Physiol. 1985;248:G676–681. doi: 10.1152/ajpgi.1985.248.6.G676. [DOI] [PubMed] [Google Scholar]
- Favus MJ, Kathpalia SC, Coe FL, Mond AE. Effects of diet calcium and 1,25-dihydroxyvitamin D3 on colon calcium active transport. Am J Physiol. 1980;238:G75–78. doi: 10.1152/ajpgi.1980.238.2.G75. [DOI] [PubMed] [Google Scholar]
- Feher JJ, Wasserman RH. Intestinal calcium-binding protein and calcium absorption in cortisol-treated chicks: effects of vitamin D3 and 1,25-dihydroxyvitamin D3. Endocrinology. 1979;104:547–551. doi: 10.1210/endo-104-2-547. [DOI] [PubMed] [Google Scholar]
- Fujita H, Sugimoto K, Inatomi S, Maeda T, Osanai M, Uchiyama Y, Yamamoto Y, Wada T, Kojima T, Yokozaki H, Yamashita T, Kato S, Sawada N, Chiba H. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol Biol Cell. 2008;19:1912–1921. doi: 10.1091/mbc.E07-09-0973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–1550. doi: 10.1083/jcb.141.7.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halloran BP, Barthell EN, DeLuca HF. Vitamin D metabolism during pregnancy and lactation in the rat. Proc Natl Acad Sci U S A. 1979;76:5549–5553. doi: 10.1073/pnas.76.11.5549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halloran BP, DeLuca HF. Calcium transport in small intestine during pregnancy and lactation. Am J Physiol. 1980;239:E64–68. doi: 10.1152/ajpendo.1980.239.1.E64. [DOI] [PubMed] [Google Scholar]
- Halloran BP, P AA. Vitamin D metabolism in aging. In: P J, G F, Feldman D, editors. Vitamin D. San Diego: 2005. pp. 823–838. [Google Scholar]
- Hirst MA, Feldman D. 1,25-Dihydroxyvitamin D3 receptors in mouse colon. J Steroid Biochem. 1981;14:315–319. doi: 10.1016/0022-4731(81)90147-3. [DOI] [PubMed] [Google Scholar]
- Hoenderop JG, Nilius B, Bindels RJ. Calcium absorption across epithelia. Physiol Rev. 2005;85:373–422. doi: 10.1152/physrev.00003.2004. [DOI] [PubMed] [Google Scholar]
- Huybers S, Naber TH, Bindels RJ, Hoenderop JG. Prednisolone-induced Ca2+ malabsorption is caused by diminished expression of the epithelial Ca2+ channel TRPV6. Am J Physiol Gastrointest Liver Physiol. 2007;292:G92–97. doi: 10.1152/ajpgi.00317.2006. [DOI] [PubMed] [Google Scholar]
- Hylander E, Ladefoged K, Jarnum S. Calcium absorption after intestinal resection. The importance of a preserved colon. Scand J Gastroenterol. 1990;25:705–710. doi: 10.3109/00365529008997596. [DOI] [PubMed] [Google Scholar]
- Kuro-o M, Matsumura Y, Aizawa H, Kawaguchi H, Suga T, Utsugi T, Ohyama Y, Kurabayashi M, Kaname T, Kume E, Iwasaki H, Iida A, Shiraki-Iida T, Nishikawa S, Nagai R, Nabeshima YI. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature. 1997;390:45–51. doi: 10.1038/36285. [DOI] [PubMed] [Google Scholar]
- Kurosu H, Ogawa Y, Miyoshi M, Yamamoto M, Nandi A, Rosenblatt KP, Baum MG, Schiavi S, Hu MC, Moe OW, Kuro-o M. Regulation of fibroblast growth factor-23 signaling by klotho. J Biol Chem. 2006;281:6120–6123. doi: 10.1074/jbc.C500457200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutuzova GD, Akhter S, Christakos S, Vanhooke J, Kimmel-Jehan C, Deluca HF. Calbindin D(9k) knockout mice are indistinguishable from wild-type mice in phenotype and serum calcium level. Proc Natl Acad Sci U S A. 2006;103:12377–12381. doi: 10.1073/pnas.0605252103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutuzova GD, Deluca HF. Gene expression profiles in rat intestine identify pathways for 1,25-dihydroxyvitamin D(3) stimulated calcium absorption and clarify its immunomodulatory properties. Arch Biochem Biophys. 2004;432:152–166. doi: 10.1016/j.abb.2004.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutuzova GD, Sundersingh F, Vaughan J, Tadi BP, Ansay SE, Christakos S, Deluca HF. TRPV6 is not required for 1alpha,25-dihydroxyvitamin D3-induced intestinal calcium absorption in vivo. Proc Natl Acad Sci U S A. 2008;105:19655–19659. doi: 10.1073/pnas.0810761105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ledger GA, Burritt MF, Kao PC, O’Fallon WM, Riggs BL, Khosla S. Role of parathyroid hormone in mediating nocturnal and age-related increases in bone resorption. J Clin Endocrinol Metab. 1995;80:3304–3310. doi: 10.1210/jcem.80.11.7593443. [DOI] [PubMed] [Google Scholar]
- Lee DB, Walling MM, Levine BS, Gafter U, Silis V, Hodsman A, Coburn JW. Intestinal and metabolic effect of 1,25-dihydroxyvitamin D3 in normal adult rat. Am J Physiol. 1981;240:G90–96. doi: 10.1152/ajpgi.1981.240.1.G90. [DOI] [PubMed] [Google Scholar]
- Lee GS, Choi KC, Jeung EB. Glucocorticoids differentially regulate expression of duodenal and renal calbindin-D9k through glucocorticoid receptor-mediated pathway in mouse model. Am J Physiol Endocrinol Metab. 2006;290:E299–307. doi: 10.1152/ajpendo.00232.2005. [DOI] [PubMed] [Google Scholar]
- Li YC, Pirro AE, Amling M, Delling G, Baron R, Bronson R, Demay MB. Targeted ablation of the vitamin D receptor: an animal model of vitamin D-dependent rickets type II with alopecia. Proc Natl Acad Sci U S A. 1997;94:9831–9835. doi: 10.1073/pnas.94.18.9831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lieben L, Benn BS, Ajibade D, Stockmans I, Moermans K, Hediger MA, Peng JB, Christakos S, Bouillon R, Carmeliet G. Trpv6 mediates intestinal calcium absorption during calcium restriction and contributes to bone homeostasis. Bone. 2010;47:301–308. doi: 10.1016/j.bone.2010.04.595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meites J, Lu KH, Wuttke W, Welsch CW, Nagasawa H, Quadri SK. Recent studies on functions and control of prolactin secretion in rats. Recent Prog Horm Res. 1972;28:471–526. [PubMed] [Google Scholar]
- O’Loughlin PD, Morris HA. Estrogen deficiency impairs intestinal calcium absorption in the rat. J Physiol. 1998;511:313–322. doi: 10.1111/j.1469-7793.1998.313bi.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pahuja DN, DeLuca HF. Stimulation of intestinal calcium transport and bone calcium mobilization by prolactin in vitamin D-deficient rats. Science. 1981;214:1038–1039. doi: 10.1126/science.7302575. [DOI] [PubMed] [Google Scholar]
- Petith MM, Wilson HD, Schedl HP. Vitamin D dependence of in vivo calcium transport and mucosal calcium binding protein in rat large intestine. Gastroenterology. 1979;76:99–104. [PubMed] [Google Scholar]
- Pike JW, Parker JB, Haussler MR, Boass A, Toverud SV. Dynamic changes in circulating 1,25-dihydroxyvitamin D during reproduction in rats. Science. 1979;204:1427–1429. doi: 10.1126/science.451573. [DOI] [PubMed] [Google Scholar]
- Reid IR. Glucocorticoid osteoporosis--mechanisms and management. Eur J Endocrinol. 1997;137:209–217. doi: 10.1530/eje.0.1370209. [DOI] [PubMed] [Google Scholar]
- Song Y, Peng X, Porta A, Takanaga H, Peng JB, Hediger MA, Fleet JC, Christakos S. Calcium transporter 1 and epithelial calcium channel messenger ribonucleic acid are differentially regulated by 1,25 dihydroxyvitamin D3 in the intestine and kidney of mice. Endocrinology. 2003;144:3885–3894. doi: 10.1210/en.2003-0314. [DOI] [PubMed] [Google Scholar]
- Stumpf WE, Sar M, Reid FA, Tanaka Y, DeLuca HF. Target cells for 1,25-dihydroxyvitamin D3 in intestinal tract, stomach, kidney, skin, pituitary, and parathyroid. Science. 1979;206:1188–1190. doi: 10.1126/science.505004. [DOI] [PubMed] [Google Scholar]
- Teerapornpuntakit J, Dorkkam N, Wongdee K, Krishnamra N, Charoenphandhu N. Endurance swimming stimulates transepithelial calcium transport and alters the expression of genes related to calcium absorption in the intestine of rats. Am J Physiol Endocrinol Metab. 2009;296:E775–786. doi: 10.1152/ajpendo.90904.2008. [DOI] [PubMed] [Google Scholar]
- Tsujikawa H, Kurotaki Y, Fujimori T, Fukuda K, Nabeshima Y. Klotho, a gene related to a syndrome resembling human premature aging, functions in a negative regulatory circuit of vitamin D endocrine system. Mol Endocrinol. 2003;17:2393–2403. doi: 10.1210/me.2003-0048. [DOI] [PubMed] [Google Scholar]
- Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol. 2001;2:285–293. doi: 10.1038/35067088. [DOI] [PubMed] [Google Scholar]
- Van Cromphaut SJ, Dewerchin M, Hoenderop JG, Stockmans I, Van Herck E, Kato S, Bindels RJ, Collen D, Carmeliet P, Bouillon R, Carmeliet G. Duodenal calcium absorption in vitamin D receptor-knockout mice: functional and molecular aspects. Proc Natl Acad Sci U S A. 2001;98:13324–13329. doi: 10.1073/pnas.231474698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Cromphaut SJ, Rummens K, Stockmans I, Van Herck E, Dijcks FA, Ederveen AG, Carmeliet P, Verhaeghe J, Bouillon R, Carmeliet G. Intestinal calcium transporter genes are upregulated by estrogens and the reproductive cycle through vitamin D receptor-independent mechanisms. J Bone Miner Res. 2003;18:1725–1736. doi: 10.1359/jbmr.2003.18.10.1725. [DOI] [PubMed] [Google Scholar]
- van de Graaf SF, Chang Q, Mensenkamp AR, Hoenderop JG, Bindels RJ. Direct interaction with Rab11a targets the epithelial Ca2+ channels TRPV5 and TRPV6 to the plasma membrane. Mol Cell Biol. 2006;26:303–312. doi: 10.1128/MCB.26.1.303-312.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van de Graaf SF, Hoenderop JG, Gkika D, Lamers D, Prenen J, Rescher U, Gerke V, Staub O, Nilius B, Bindels RJ. Functional expression of the epithelial Ca(2+) channels (TRPV5 and TRPV6) requires association of the S100A10-annexin 2 complex. EMBO J. 2003;22:1478–1487. doi: 10.1093/emboj/cdg162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vergne-Marini P, Parker TF, Pak CY, Hull AR, DeLuca HF, Fordtran JS. Jejunal and ileal absorption in patients with chronic renal disease. Effect of 1alpha-hydroxycholecalciferol. J Clin Invest. 1976;57:861–866. doi: 10.1172/JCI108362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman RH. Vitamin D and intestinal absorption of calcium: a view and overview. In: P JW, Feldman D, Glorieux F, editors. Vitamin D. Acedemin press; San Diego, CA: 2005. pp. 411–428. [Google Scholar]
- Wood RJ, Fleet JC, Cashman K, Bruns ME, Deluca HF. Intestinal calcium absorption in the aged rat: evidence of intestinal resistance to 1,25(OH)2 vitamin D. Endocrinology. 1998;139:3843–3848. doi: 10.1210/endo.139.9.6176. [DOI] [PubMed] [Google Scholar]
- Xue Y, Fleet JC. Intestinal vitamin D receptor is required for normal calcium and bone metabolism in mice. Gastroenterology. 2009;136:1317–1327. e1311–1312. doi: 10.1053/j.gastro.2008.12.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida T, Fujimori T, Nabeshima Y. Mediation of unusually high concentrations of 1,25-dihydroxyvitamin D in homozygous klotho mutant mice by increased expression of renal 1alpha-hydroxylase gene. Endocrinology. 2002;143:683–689. doi: 10.1210/endo.143.2.8657. [DOI] [PubMed] [Google Scholar]
- Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K, Yoshihara Y, Kawakami T, Arioka K, Sato H, Uchiyama Y, Masushige S, Fukamizu A, Matsumoto T, Kato S. Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine hypoplasia and growth retardation after weaning. Nat Genet. 1997;16:391–396. doi: 10.1038/ng0897-391. [DOI] [PubMed] [Google Scholar]
- Zhang W, Na T, Wu G, Jing H, Peng JB. Down-regulation of intestinal apical calcium entry channel TRPV6 by ubiquitin E3 ligase Nedd4-2. J Biol Chem. 2010;285:36586–36596. doi: 10.1074/jbc.M110.175968. [DOI] [PMC free article] [PubMed] [Google Scholar]