

Eruption of volcanic rocks at the midocean ridges is the major mechanism by which heat is lost from the interior of the Earth. Approximately one-third of the heat is removed from the spreading centers by convective circulation of seawater (1). The magnitude of this heat loss requires that the entire volume of the oceans circulates through the midocean ridges in approximately 10 million years. Seawater interaction with volcanic rocks at near 400°C results in substantial chemical flux and makes an important contribution to buffering the composition of some elements in seawater. Cations from seawater (Mg+2, Ca+2, and Na+) form hydroxyl-bearing alteration minerals in the volcanic rocks, releasing hydrogen ion to solution. The hot, acidic altered-seawater releases metals (Fe, Mn, Zn, and Cu) and reduced sulfur (H2S) from the volcanic rock; these are transported by hydrothermal solutions to the seafloor and form metallic mineral deposits.
Seafloor hydrothermal vents support ecosystems with enormous biomass and productivity compared with that observed elsewhere in the deep oceans. What is the energy source that fuels these oases of life, and what adaptations allow them to exist in these extreme environments?
Oxidation/reduction (redox) reactions are key to supporting chemosynthesis. The atmosphere and hydrosphere are relatively oxidizing with an abundance of potential electron acceptors (O2, SO4=, and NO3-). In contrast, the basaltic rocks that form the oceanic crust are relatively reduced because of the abundance of ferrous iron. High-temperature fluid/rock interaction forms reduced gases (H2S, H2, and CH4) that dissolve in hydrothermal fluid. Representative redox reactions that produce H2S include:
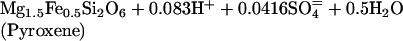 |
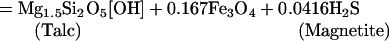 |
1 |
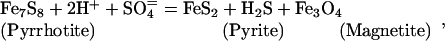 |
2 |
where talc serves as a proxy for a Mg hydroxide component of alteration minerals such as chlorite and amphibole. Sulfur is a particularly important component in the subseafloor redox cycle because there is a transfer of eight electrons (derived from oxidation of Fe+2 to Fe+3) in the reduction of seawater sulfate to sulfide. Microbial reoxidation of H2S near the seafloor releases the stored energy and drives biochemical reactions.
Serpentinization reactions also may be important in supporting chemosynthetic communities (2), especially at off-axis sites. Serpentinization reactions are not limited to the high-temperature portions of seawater circulation systems, and the H2 provided by reactions such as 3 is readily metabolizable by a variety of microbes.
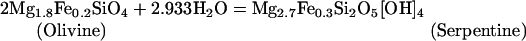 |
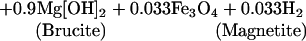 |
3 |
Reduced magmatic gases (H2 and H2S) also are released directly during intrusion and eruption of basaltic magma. Seafloor hydrothermal systems undergo cyclic variations in temperature, vent fluid composition, and biological activity in response to magmatic episodes (3, 4). Volcanic events initially are followed by relative increases in vent fluid temperature and magmatic gases, often accompanied by decreased salinity in hydrothermal fluids as a result of supercritical phase separation of seawater in the subsurface (5) and faster migration of the vapor-like component to the seafloor. Later in the cycle (on the order of a few years) the high-salinity component of the phase-separated fluids reaches the seafloor. The higher chloride content of these fluids enhances metal solubility, but H2S content drops, consistent with solubility control by metal sulfide minerals. Microbial blooms are observed at the initiation of the magmatic cycle, suggesting that the upper oceanic crust is inoculated with microbial communities poised to exploit the chemical energy carried by hydrothermal fluids.
Although there is a potential abundance of chemical energy, deep-sea hydrothermal communities have had to adapt to extreme conditions to exploit this resource. Of particular interest are the hyperthermophiles, which are defined as microorganisms able to grow at 90°C and above. About 20 different types of such organisms are now known (6). They have been found both within the walls of black smoker chimneys and where the hydrothermal vent fluids mix with the surrounding seawater. Classification of the hyperthermophiles has provided new insights into evolution and the origin of life. All but two of the hyperthermophilic genera are classified by ribosomal RNA analyses as Archaea (formerly Archaebacteria), which are the second domain of prokaryotic life, in addition to the bacteria (7). Interestingly, by these phylogenetic analyses, the hyperthermophilic archaea and the two hyperthermophilic bacteria are the most slowly evolving within their domains, suggesting that life may have first evolved when the Earth was much hotter than it is now. Such a thesis is very controversial (8) but indicates that extant life forms are largely the result of temperature adaptations to lower (below hyperthermophilic) temperatures.
Evolution gives no clue, however, as to how life can thrive near and above 100°C. Most microbes, and all eukaryotic cells, cannot survive at temperatures much above 50°C, because of the general instability of biological molecules. The three-dimensional structures of most enzymes and proteins are lost at temperatures much above 70°C, and the double-helical structure of DNA has a comparable lack of stability in in vitro studies. There are also a wide variety of ubiquitous metabolites that are rapidly hydrolyzed at temperatures above 90°C. How do hyperthermophilic cells circumvent these problems?
Although there are some examples of modified pathways and unusual enzymes in hyperthermophiles (9, 10), in general their biochemistry closely resembles that of the mesophilic world. Yet, most enzymes from hyperthermophiles are extremely stable at high temperatures, showing optimal catalytic activity above 100°C with virtually no activity at ambient temperature. They contain exactly the same 20 amino acids as enzymes from conventional organisms, so why are they so stable? Sequence comparisons of analogous proteins from hyperthermophilic and conventional organisms are essentially identical, so the enormous amount of sequence information now becoming available (see www.tigr.org) will be of little use in elucidating stabilizing mechanisms. Comparisons must be made at the level of the three-dimensional structures. Yet, even then, there are no gross structural differences between hyperthermophilic proteins and their mesophilic counterparts, and both forms are stabilized by the same noncovalent interactions (11). The number and extent of such interactions is generally only slightly higher in the hyperthermophilic versions, so extended protein stability at 100°C appears to be the result of very subtle, synergistic and cooperative intramolecular interactions. Moreover, different types of hyperthermophilic protein seem to have unique solutions to the problem. A general mechanism by which any conventional protein could be made stable and functional at temperatures above 100°C may not be forthcoming.
For DNA the stabilizing mechanisms are not quite as mysterious. Denaturation, depurination, and strand cleavage of DNA are greatly minimized by the relatively high salt (Mg, K, and PO4) concentrations typically (although not universally) found in hyperthermophiles (12). Many of these organisms also have histone-like proteins bound to their DNA (13) and some contain a unique enzyme termed DNA reverse gyrase (14), both of which may afford thermal protection. How simple organic metabolites are stabilized, however, remains a mystery. Some may be “channeled” from one enzyme to the next although it is not clear whether this is a widespread mechanism.
Many mesophilic microbes in the hydrothermal vent environment use H2S as an energy source. This molecule is generally highly toxic to aerobic life forms, yet dense populations of organisms flourish in a variety of sulfide-enriched environments. How have they evolved to deal with this extreme environment?
Hydrothermal vent communities can inhabit sulfide-rich habitats because of evolution of detoxification mechanism that often involve microbial symbionts. Detoxification of sulfide through binding to blood-borne components is known in chemosynthetic vestimentiferans and vesicomyid clams and is particularly well characterized for the tube worm Riftia pachyptila (15, 16). The abundant respiratory hemoglobin present in the plume of Riftia is capable of binding oxygen and sulfide simultaneously with very high affinities. The blood transports the respiratory hemoglobin with the tightly bound sulfide to the internal symbiotic bacteria, providing an electron donor for bacterial chemoautotrophy, while also protecting animal tissue by sequestering the toxin as a bound form. Colonies of Riftia are anchored on the rocks where hydrothermal fluid (12–15°C) issues out onto the sea floor. At the base of their tubes, hydrothermal fluid is enriched in H2S and CO2, but is devoid of oxygen. The respiratory plume is extended into the ambient (2°C), oxygen-enriched bottom water. Riftia's unusual microhabitat is the interface between the hydrothermal fluids and the ambient bottom water where essential metabolites can be taken up by the plume and transported to internal bacteria for metabolism. The steep thermal and chemical gradients provide access to the reduced compounds needed to fuel growth and the oxygen needed to burn the fuel.
Tube worms associated with hydrocarbon seeps are long and thin with internal morphology and physiology similar to Riftia. However, they live in an environment where sulfide is present only in the soft sediment, not in the water column. These worms are hypothesized to acquire sulfide across a thin extension of the posterior-most portion of the tube that extends into the sediment (17). Rather than taking up H2S across the plume surface, cold seep tube worms appear to transport sulfide across the body wall.
Vesicomyid clams living in hydrothermal vents have endosymbiont-containing gills. Vesicomyid blood transports oxygen bound to hemoglobin and contains an extracellular component with a high sulfide binding affinity (18). In this species, sulfide and oxygen acquisition are spatially separated. The foot of the clam is extended down into crevices that vent sulfide-rich water, enabling sulfide uptake and transport through the circulatory system to gills that are bathed in oxygen-rich seawater circulated from above. Although Bathymodiolid mussels living at vents have retained the ability to filter feed they also have abundant microbial symbionts located in their gills. However, these mussels oxidize sulfide to nontoxic thiosulfate, which is used by chemoautotrophic symbionts rather than H2S (19).
Although various sulfide detoxification strategies have evolved, cumulative data suggest that sulfide binding is a particularly effective detoxification mechanism for animals harboring sulfide-oxidizing symbionts. Other marine invertebrates that have no sulfide binding protein appear to depend on sulfide oxidation for detoxification. Both strategies allow large communities of animals to flourish in habitats that typically would be considered inhospitable to life.
Footnotes
This paper is a summary of a session presented at the 11th annual symposium on Frontiers of Science, held November 11–13, 1999, at the Arnold and Mabel Beckman Center of the National Academies of Sciences and Engineering in Irvine, CA.
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.210395997.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.210395997
References
- 1.Humphris S E, Zierenberg R A, Mullineaux L S, Thomson R E, editors. Am Geophys Union Monogr. 1995;91:1–466. [Google Scholar]
- 2.Alt J C, Shanks W C., III J Geophys Res. 1998;103:9917–9929. [Google Scholar]
- 3.Delaney J R, Kelley D S, Lilley M D, Butterfield P A, Baross J A, Wilcock W S D, Embley R W, Summit M. Science. 1998;281:222–230. [PubMed] [Google Scholar]
- 4.Shank T M, Fornari D J, Von Damm K L, Lilley M D, Haymon R M, Lutz R A. Deep-Sea Res. 1998;45:465–515. [Google Scholar]
- 5.Von Damm K L, Oosting S E, Kozlowski R, Buttermore L G, Colodner D C, Edmonds H N, Edmond J M, Grebmeier J M. Nature (London) 1995;375:47–50. [Google Scholar]
- 6.Stetter K O. FEMS Microbiol Rev. 1996;18:149–158. doi: 10.1111/j.1574-6976.2000.tb00562.x. [DOI] [PubMed] [Google Scholar]
- 7.Woese C R, Kandler O, Wheelis M L. Proc Natl Acad Sci USA. 1990;87:4576–4579. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wiegel J, Adams M W W, editors. Thermophiles: The Keys to Molecular Evolution and the Origin of Life? Washington, DC: Taylor and Francis; 1998. [Google Scholar]
- 9.Kengen S W M, Tuininga J E, Debok F A M, Stams A J M, De Vos W M. J Biol Chem. 1995;270:30453–30457. doi: 10.1074/jbc.270.51.30453. [DOI] [PubMed] [Google Scholar]
- 10.Adams M W W, Kletzin A. Adv Protein Chem. 1996;48:101–180. doi: 10.1016/s0065-3233(08)60362-9. [DOI] [PubMed] [Google Scholar]
- 11.Jaenicke R, Bohm G. Curr Opin Struct Biol. 1998;8:738–748. doi: 10.1016/s0959-440x(98)80094-8. [DOI] [PubMed] [Google Scholar]
- 12.Lamosa P, Martins L O, DaCosta M S, Santos H. Appl Environ Microbiol. 1998;64:3591–3598. doi: 10.1128/aem.64.10.3591-3598.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grayling R A, Sandman K, Reeve J N. Adv Protein Chem. 1996;48:437–467. doi: 10.1016/s0065-3233(08)60368-x. [DOI] [PubMed] [Google Scholar]
- 14.Forterre P, Bergerat A, Lopez-Garcia P. FEMS Microbiol Rev. 1996;18:237–248. doi: 10.1111/j.1574-6976.1996.tb00240.x. [DOI] [PubMed] [Google Scholar]
- 15.Arp A J, Childress J J, Fisher C R., Jr Bull Biol Soc Wash. 1985;6:289–300. [Google Scholar]
- 16.Childress J J, Fisher C R. Oceanogr Mar Biol Annu Rev. 1992;30:337–441. [Google Scholar]
- 17.Julian D, Gaill F, Wood E, Arp A J, Fisher C R. J Exp Biol. 1999;202:2245–2257. doi: 10.1242/jeb.202.17.2245. [DOI] [PubMed] [Google Scholar]
- 18.Arp A J, Childress J J, Fisher C R., Jr Physiol Zool. 1984;57:648–662. [Google Scholar]
- 19.Fisher C R, Childress J J, Oremland R S, Bidigare R R. Mar Biol (Berlin) 1987;96:59–71. [Google Scholar]