

Abstract
Rats that consume high-energy (HE) diets (i.e., diets high in saturated fats and sugar) show impaired hippocampal-dependent learning and memory (e.g., [1]). To further investigate this effect, we trained rats given restricted access to low-fat lab chow on hippocampal-dependent serial feature-negative (FN) and hippocampal-independent simple discrimination problems. When training was completed, Group Chow received ad libitum lab chow. The remaining rats received ad libitum HE diet. Performance on both discrimination problems was tested following 7, 14, 21 and 28 days of HE diet exposure. FN, but not simple discrimination, was abolished initially for all rats, and then re-emerged for Group Chow. For rats fed HE diet, those that weighed the least and had lowest amount of body fat (HE-diet resistant (HE-DR) rats), performed like Group Chow on both discrimination problems. However, HE diet-induced obese (HE-DIO) rats (i.e., rats that weighed the most weight and had the most body fat) performed like Group Chow on the simple discrimination problem, but were impaired throughout testing on the FN problem. Subsequent assessment of blood-brain barrier (BBB) permeability revealed that concentrations of an exogenously administered dye were elevated in the hippocampus, but not in the striatum or prefrontal cortex for HE-DIO rats relative to the HE-DR and Chow groups. The results indicate that the adverse consequences of HE diet on hippocampal-dependent cognitive functioning are associated with detrimental effects on the BBB and that both of these outcomes vary with sensitivity to HE diet-induced increases in weight and adiposity.
Keywords: hippocampus, high-fat diet, obesity, learning, adiposity, energy balance, cognition
1. Introduction
Intake of high-energy (HE) or “Western” diets (diets high in sugars and saturated fat) is known to promote obesity, Type II diabetes, hyperglycemia, hyperlipidemia, and other metabolic disorders [1]. Furthermore, evidence has emerged which indicates that consuming these diets is also associated with deficits in types of learning and memory that depend on the functional integrity of the hippocampus. For example, Francis and Stevenson [2] reported that performance on neuropsychological tests used to assess hippocampal function was impaired for humans who self-reported consuming high amounts of sugar and saturated fats compared to humans that consumed lower amounts. In contrast, tests of executive function, which is thought be mediated by the prefrontal cortex, did not reveal differences based on self-reported diet composition. However, studies conducted under other conditions (e.g., [3]) have reported a link between intake of high-fat diets and impaired executive function.
The results of Francis and Stephenson are reminiscent of recent findings [4] which showed that learning a hippocampal-dependent Pavlovian serial feature-negative (FN) task was impaired, relative to standard chow-fed controls, for rats that had been maintained on a HE diet for 90 days prior to the beginning of training. This task has the form Tone+, Light→Tone-, where a 5-sec auditory stimulus is followed by the delivery of sucrose pellets on Tone+ trials, but not on Light→Tone- trials where a 5-sec visual stimulus (e.g., a light) precedes the onset of that same auditory cue. Rats show that they have solved the serial FN by exhibiting more conditioned appetitive behavior on Tone+ compared to Light→Tone- trials (e.g.,[5]). Rats with selective lesions to the hippocampus are impaired in solving this problem [6]). In contrast, no differences between the rats fed chow and rats fed HE diet were observed on a concurrently trained, largely hippocampal-independent simple discrimination problem with different 5-sec auditory cues (e.g., clicker, white noise) signaling when sucrose pellets would and would not be presented. This pattern of findings indicated that HE diet had a selective effect on a hippocampal-dependent learning and memory process.
Little is currently known about the mechanisms that might underlie the capacity of HE diets to interfere with hippocampal-dependent memory function. However, Kanoski et al., [4] have observed a relationship between hippocampal dysfunction and the effects of HE diet on blood-brain barrier (BBB) permeability (also see [7]). The BBB helps to maintain brain homeostasis by restricting material flux between the blood and brain interstitial fluids. Complex tight junctions between adjacent cerebral endothelial cells are the structural basis for the BBB. These complex molecular junctions make the brain practically inaccessible for water-soluble or polar molecules, unless they are transported selectively by various transporters on the BBB [8]. In addition to exhibiting performance deficits in a hippocampal-dependent learning and memory task, rats maintained on an HE diet also showed reduced expression of several tight-junction proteins that are known to be essential to the BBB’s integrity. Furthermore, following infusion with sodium fluorescein (NaFl), a small molecule dye that is normally precluded from crossing the BBB, elevated concentrations of NaFl were found in the hippocampi of these same HE diet-fed rats relative to their controls. No differences in the concentration of NaFl were found in striatum or prefrontal cortex, brain regions that have been implicated in cognitive and movement coordination functions that are distinct from those involving the hippocampus [9-11]
The present study extends the work of Kanoski et al., [4] in several ways. First, we trained rats to asymptote on both the serial FN and simple discrimination problems before exposure to the HE diet was initiated. Thus, rather than examining the effects of consuming HE diet on original learning of these hippocampal-dependent and hippocampal-independent discrimination problems, the present study compared the effects of these diets on post-learning retention. This modification allowed us to evaluate the effects HE diet on memory and retrieval of information acquired before, rather than after the onset of diet-induced weight gain. Second, the effects of the HE diets on retention were compared to the effects of regular chow at several time points after both diets were made available ad libitum. This enabled us to study the time course of any HE-diet induced impairment in memory performance. Third, we looked at the effects of HE diet on memory as a function of the susceptibility of the rats to diet-induced increases in body weight, body adiposity, and energy intake. Specifically, consuming a HE diet results in diet-induced obesity for some rats, whereas other rats are resistant to this effect of the HE diet [12]. We compared HE diet-induced obese (DIO) rats with diet resistant (DR) rats and chow controls in terms of their performance on both the serial FN and simple discrimination problems and also with respect to the permeability of the BBB.
Previously, we proposed that HE diets promote hippocampal dysfunction and that this dysfunction in turn promotes increased food intake, body weight and adiposity [1, 12-14]. From the perspective of this “vicious-cycle” model, HE-fed DR rats that eat less and gain less weight and body fat, compared to DIO rats, should also be less impaired on the hippocampal-dependent serial FN problem. Neither DIO nor DR rats should be impaired relative to chow controls on the hippocampal-independent simple discrimination task. Furthermore, if the effects of HE diet on hippocampal functioning also depend on the BBB permeability, a greater permeability should be observed for the DIO compared to the DR phenotype.
2. Methods
2.1. Subjects
The subjects were 24 naïve, male, Sprague-Dawley rats, weighing between 250-275 g upon arrival at the laboratory from Harlan Inc, Indianapolis, IN. The animals were housed individually in a climate-controlled environment under a 12:12 hr light:dark cycle with the light phase beginning at 0700 hrs each day. The rats had free-access water throughout the study, except during experimental sessions as described below. The care and use of all animals in this study was reviewed and approved by the Purdue University Animal Care and Use Committee.
2.2. Apparatus
All behavioral training was conducted in 8 conditioning chambers purchased from Med Associates (Georgia VT, USA). The chambers were identical, each with aluminum end walls and clear plexiglass side walls, measuring 21.6 × 21.6 × 27.9 cm and with floors consisting of stainless steel rods (0.48 cm in diameter) spaced 1.9 cm apart. A recessed food-cup was located in the center of one end wall and a 6-W jeweled panel light was located approximately 6 cm above and to the left of the opening for the food cup. Diffuse tone (1500 hz), white noise and clicker (3hz) stimuli (all approximately 78 db) were produced by auditory stimulus generators (Med Associates, ANL-926) located outside the conditioning chambers near the end wall opposite of the food magazine. A motorized pellet dispenser attached to each chamber was used to deliver 45 mg sucrose pellets to the food cup. A computerized infrared monitoring system with a photo transmitter and receptor was positioned in each chamber so that photobeam interruptions would index entries into the recessed area with the food cup. All experimental events (e.g., stimulus presentations, pellet delivery, recording of beam breaks) were controlled using MED PC IV (Med Associates) computer software. The body adiposity of each rat was measured using an EchoMRI-900 magnetic resonance body composition analyzer (Echo Medical Systems, LLC, Houston, TX, USA)
2.3. Procedure
The rats were fed standard rodent laboratory chow (LabDiet, formula 5001) ad libitum for 30 days after their arrival in the lab, after which food rationing was used to gradually reduce the body weight of each rat to 85% of the average weight obtained on the last two days of ad libitum feeding. Behavioral training began when all rats achieved this 85% (+/- 1%) body weight criterion. The rats were weighed immediately prior to each training and probe test session. All rats received their daily food ration immediately after the completion of each training session under food deprivation.
2.3.1. Magazine training
The rats were assigned to three squads of eight rats each with each rat assigned to one of the 8 conditioning chambers for the duration of the study. The rats were then given one 15-min session of magazine training to help habituate them to the apparatus and to learn the location of the food cup. During this session, all rats received 10 presentations of two sucrose pellets, which were delivered according to a variable-time 60-sec schedule (i.e., average of one presentation of two pellets per min).
2.3.2. Discrimination training
All animals received one 90 min session of discrimination training per day. Training sessions were conducted between 0930-1400h. In each session, the rats received two different types of concurrently trained discrimination problems, serial feature negative (FN) and simple (CS+/CS-), each comprising two types of trials, one reinforced and one nonreinforced. For FN discrimination training, rats were given trials in which the presentation of a 5-sec tone terminated with the delivery of two sucrose pellets (on Tone+ trials), whereas no pellets were delivered when a 5-sec illumination of the panel light (L) followed by a 5-sec empty interval (i.e., with no programmed stimulus) preceded presentation of the 5-sec tone (on Light→Tone- trials). For simple CS+/ CS- discrimination training, a 5-sec presentation of the white noise terminated with the delivery of two sucrose pellets (on a CS+ trial) whereas no pellets were delivered after a 5 sec presentation of the clicker (on CS- trials). Each session consisted of one Tone+ trial and three Light→Tone- trials along with one CS+ trial and three CS- trials. Trial orders were randomly selected for each session and the intertrial interval was randomized within the range of 300 to 900 sec (average = 600 sec). Training was completed after 40 sessions.
2.3.3. Diets
Immediately after the end of discrimination training all rats were fed standard lab chow ad libitum for 8 days at which time all rats had regained body weight up 100% of that recorded prior to food rationing. The rats were then assigned to two diet groups that were matched on performance on both FN and simple discrimination performance over the last two sessions of training. One group of 18 rats, received ad libitum powdered high-energy (HE) diet (Harlan Teklad, TD.04489). This nutritionally complete diet has a caloric density of approximately 4.5 kcal/g (≈40% kcal from fat, 38% from carbohydrate and 21% from protein) and contained 270 g/kg casein, 220.5 g/kg glucose, 200 g/kg cornstarch, 50 g/kg cellulose, 170 g/kg lard, 15 g/kg safflower oil. The remaining 6 rats were fed the same type of standard laboratory rodent chow (LabDiet, formula 5001) that was used during training, but now in powdered form. This chow control diet had a caloric density of approximately 3.0 kcal/g (≈13% kcal from fat). Both diets were presented in glass jars fastened inside of the home cage of each rat. All rats were fed their respective diets ad libitum for the remainder the study.
2.3.4. Classification of obesity phenotype
Rats were weighed immediately prior to being given their powdered test diets and were weighed daily thereafter. After 21 days on the test diets, clear differences in body weight and body weight gain emerged between the rats fed the HE diet and the chow-fed controls. The rats fed the HE diet were then further subdivided into three groups of six each based on body weight gain. The six rats that gained the most body weight were classified as diet-induced obese (DIO) whereas the six rats that gained the least amount of weight were classified as DR. The data for middle third of rats with respect to weight gain were discarded.
2.3.5. Discrimination probe testing
All rats were returned to their previously assigned conditioning chambers for probe tests on both the originally-trained FN and simple discrimination problems. The probe tests were given after the rats had been on their diets for 7, 14, 21, and 28 days. Each probe test consisted of two sessions conducted on consecutive days using the same procedures as were used during original training sessions, except that all test sessions were conducted under ad libitum feeding conditions.
2.3.6. Analysis of body adiposity
At the conclusion of each probe test we assessed body fat as a percent of total body mass. Rats were placed in a constraint tube which was then inserted into the EchoMRI-900 for period of approximately 90 sec. During that time, total and % body fat, lean, and fluid was calculated and recorded and the rat was removed from the apparatus.
2.3.7. Measurement of food intake
On the day following each probe test, the food hoppers of each rat were filled with a pre-weighed amount of food. Three days later, the number of grams of food remaining in each hopper was weighed and recorded. The difference in grams (g) between the weight of the food in the hoppers the day after each probe test and the weight recorded three days later was calculated. Using 4.5 kilocalories (kcals)/gas the energy density of the HE diet and 3.0 kcals/g as the energy density of the chow, grams consumed was converted to kcals consumed for each rat.
2.3.8. Analysis of blood-brain barrier (BBB) permeability
Per procedures reported by Kanoski et al., [4], at the conclusion of probe testing, the rats were anesthetized with an intraperitoneal injection of a mixture of ketamine (100mg/kg) and xylazine (10 mg/kg). Under full anesthesia, the right femoral artery was exposed from linear incision and a polyethylene catheter fixed by ligation was introduced into the artery. Sodium fluorescein (10% NaFl, 0.6 ml/kg in saline) was applied from one syringe into the catheter at a rate of approximately 0.2 ml/min. After the NaFl had been applied, the artery was tied with a suture to prevent backflow and the dye was allowed to circulate for 45 min. The rats were then perfused intracardially with 150 ml of sterile saline at a rate of 10 ml/min to wash out the dye remaining in brain capillary. It was determined previously that this infusion rate does not damage the BBB [15]. Following perfusion the rats were decapitated and the brains were quickly removed. After removing the choroid plexus and capillaries by hand, the hippocampus, striatum, and prefrontal cortex were bilaterally extracted over ice and stored at–80°C. For NaFl fluorescence analysis, the tissue samples were weighed, homogenized in 200 μl phosphate buffered saline, and centrifuged for 10 min at 5000 rpm. Supernatants were diluted with 400 μl trichloracetic acid (20%) and then centrifuged for 15 min at 13,600 rpm. Supernatants were then neutralized with sodium hydroxide (NaOH) and fluorescence was measured in duplicates with an ELISA scanner at an excitation wavelength of 480 and emission wavelength of 525. Tissue samples were processed under minimal lighting conditions. Each of the three brain areas were processed and analyzed separately allowing comparison of the two dietary groups within each brain area. A negative control sample lacking brain tissue was used with each brain area scan; the fluorescence value of the negative control was subtracted from the fluorescence intensity value for each sample. Absolute intensity values were divided by the tissue weight for each separate sample.
3. Results
3.1. Discrimination training
Rats showed that they solved the simple (Figure 1a) discrimination problem by exhibiting more conditioned responding (food magazine beam breaks) on reinforced CS+ trials compared nonreinforced CS- trials by the end of training. A within-subjects analysis of variance (ANOVA) with Trial Type (CS+ vs CS) and Session (1-40) as factors obtained significant main effects of Trial Type (F(1, 15) = 46.26, p < 0.01) and Session (F(39, 585) = 1.55, p < 0.05) as well as a significant Trial Type × Session interaction (F(39, 585) = 13.13, p < 0.01). Newman-Keuls test showed that responding on CS+ trials was significantly greater compared to CS- trial beginning on Session 10 through the end of simple discrimination training (ps < 0.05).
Figure 1.

Performance during training of a simple Pavlovian (CS+ vs. CS-) discrimination (Figure 1A) and during training of Pavlovian serial feature negative (L→T- vs. T+) discrimination (Figure 1B) for rats reduced to 85% of their ad libitum body weight by rationing a diet of standard laboratory chow.
Rats also solved the serial FN discrimination problem. Responding on reinforced Tone+ trials (T+ in Figure 1b) exceeded that on Light→Tone- trials (L→T- in Figure 1b) prior to the end of training. This pattern of result yielded significant main effects of Trial Type (F(1, 15) = 25.02, p < 0.01), Session (F(39, 585) = 2.65, p < 0.01), and a significant Trial Type × Session interaction (F(39, 585) = 6.20, p < 0.01). Newman-Keuls tests showed that the difference between Tone+ and Light→Tone- trials was significant on Sessions 29-40 (ps. < 0.05).
For both the simple and the FN discrimination problems, additional ANOVAs with Test Group as a dummy variable were used to determine if discrimination performance for rats that were later designated as HE-DIO, HE-DR and Chow controls during testing (see below) differed at any point during training (data not shown). Both of these ANOVAs obtained significant main effects of Session, Trial Type, and a significant Session × Trial Type interactions (smallest F(39, 585) = 1.58, p. < .05 for the main effect of Sessions on the simple discrimination problem). However, the dummy variable, Group yielded no significant main effect, nor was this variable involved in any significant interactions with Session or Trial Type (largest F(78, 585) = 1.16, p. > .18 for the Session × Trial Type × Group × interaction on the simple discrimination problem). These results indicate that discrimination performance during training was statistically the same for rats which belonged to different diet groups and came to weigh different amounts during subsequent testing.
3.2. Group designation according to diet and body weight
Probe Test Day 21 was when significant differences emerged in mean body weight among the rats fed chow (416.67 g) and rats in the top (449.83 g) and bottom (429.33 g) weight tertiles that were fed the HE diet (F(3, 15) = 3.84, p < .05). Newman-Keuls tests showed that for rats fed the HE diet, rats in the highest weight tertile weighed significantly more than chow fed controls (p < .05) whereas rats in the lowest tertile for body weight did not. Accordingly, rats in the top tertile were designated HE-DIO and rats in the bottom tertile were designated HE-DR based on their weights on Day 21 of probe testing.
3.3 Body weight, body adiposity, and amount consumed
Figure 2 shows mean baseline weights and mean body weights recorded at the time of each probe test throughout the study for the rats in Groups HE-DIO and HE-DR and for the Chow controls. The figure shows, and statistical analysis confirmed, that while there were no significant differences among these groups at baseline, body weight for all groups increased significantly thereafter (Main effect of Test Day, F(4,60) = 528.98, p < .01). Figure 2 also shows that mean body weight for Group HE-DIO was higher compared to groups HE-DR and Chow on each of the subsequent test days (7, 14, 21, and 28) of the study. This pattern of results yielded a significant overall Group × Test Day interaction, F(8, 60) = 16.90, p < .01. Subsequent ANOVAs revealed significant Group × Test Day interactions when Group HE-DIO was compared to Group HE-DR (F4, 40) = 24.25, p < .01) and Chow (F(4, 40) = 32.64, p < .01), respectively. However this interaction was not significant when Group HE-DR and Group Chow were compared (F(4, 40) < 1).
Figure 2.

Mean body weight prior to the beginning of testing (Chow baseline) and on each day of probe testing 7, 14, 21, and 28 after the initiation of ad libitum feeding of HE diet (Groups HE-DIO and HE-DR) and chow (Group Chow).
Mean percent body fat at the time of each probe test also differed among three groups. Figure 3 shows that while body fat for the groups differed little at baseline, mean percent body fat increased for the HE-DIO group and to a lesser extent for the HE-DR group when tested at Day 7, and remained at about the same level for both groups on each subsequent test. In contrast, mean % body fat did not increased relative to baseline on any test day for Group Chow. An ANOVA comparing percent body fat from baseline to probe Test Day 7 obtained significant main effects of Group (F(2, 15) = 18.71, p. < .01) and Test Day (F(4, 60) = 21.96, p. < .01) as well as a significant Group × Day interaction (F(8, 60) = 19.33, p. < .01). Newman-Keuls test confirmed that while mean percent body fat did not differ significantly among the three groups at baseline, Group HE-DIO was higher compared to both Groups HE-DR and Chow and was higher for Group HE-DR compared to Chow on each test after baseline (all ps. < .05).
Figure 3.

Mean percent body fat as measured by NMR after initiation ad libitum feeding prior to the beginning of testing (Chow baseline) and on each day of probe testing for rats given ad libitum feeding of HE diet (Groups HE-DIO and HE-DR) and chow (Group Chow).
Mean amount consumed (kcals) and standard error of the mean (SEM) over the 3-day period following each probe test was 252.43 kcals, SEM 14.74 for Group HE-DIO; 201.77 kcals, SEM 15.92 for Group HE-DR; and 187.98 kcals, SEM 17.45 for Group Chow. These data yielded a significant main effect of Diet Group (F(2, 15)= 4.73, p. < .05. Newman-Keuls tests showed that the Group HE-DIO consumed significantly more than Groups HE-DR and Chow (ps. < .05) while intake for these latter two groups did not differ significantly.
3.4. Discrimination probe testing
Figure 4a shows performance for the simple discrimination problem on the last block of acquisition training and on the 7, 14, 21 and 28 day probe tests for rats in HE-DIO, HE-DR, and Chow groups. On the last block of training mean magazine beam interruptions were significantly greater on CS+ trials compared to CS- trials for all groups (F (1, 15) = 89.43, p. < .01, for the main effect of Trial Type). Trial Type did not interact with Group (F (2, 15) < 1) indicating that the magnitude of the difference between CS+ and CS- trials did not differ among the groups prior to when ad lib feeding of their designated diets was initiated.
Figure 4.
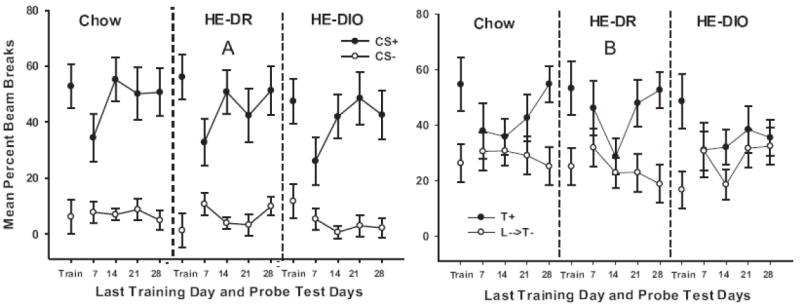
Performance on the simple Pavlovian (CS+ vs. CS-) discrimination (Figure 4A) and the Pavlovian serial feature negative (L→T- vs. T+) discrimination (Figure 4B) during the last day of training under food deprivation and during each probe test after the initiation of ad libitum feeding of HE diet for Groups HE-DIO and HE-DR and of chow for Group Chow.
Figure 4a also shows that simple discrimination performance was similar for all groups on each probe test after the diets were administered. While the magnitude of the difference between CS+ and CS- trials was somewhat smaller on the 7 day probe test compared to that observed at the end of training, the effect was observed for all diet groups. Moreover, simple discrimination performance for all groups returned to the level observed at the end of training on each of the remaining probe test days 14, 21, and 28. This pattern of results yielded a significant main effect of Trial Type (F(1, 15) = 104.78, p < .01) and a significant Trial Type by Test interaction (F(3, 45) = 8.78, p. < .01). However, there was no significant main effect of Group nor did Group interact significantly with Trial Type or Test (largest F(2, 15) < 1 for main effect of Group. Thus, simple discrimination was retained during the probe tests and did not differ as a function of diet.
Figure 4b shows FN discrimination performance on the last block of training and on each probe test for each diet group. The figure shows that the rats in each group responded more on Tone+ trials(T+) than on Light→Tone- (L→T-) trials on the last block of training, before the test diet conditions were introduced. Furthermore, as was the case with the simple discrimination problem, discrimination performance at the end of training differed little between those groups. These conclusions were confirmed by a significant main effect of Trial Type (F(1, 15) = 36.03, p. < .01) and the lack of a significant interaction between Trial Type and Group (F (2, 15) < 1) at the end of training for the FN problem.
Figure 4b also shows that all groups showed substantial decrements in FN discrimination performance on both the 7 and 14 day probe tests. For Groups HE-DR and Chow, discrimination performance began to re-emerge at the 21 and 28 day tests. In contrast, FN discrimination performance remained impaired for Group HE-DIO on all of the probe test days. An ANOVA comparing performance on the last block of training with that on Test Day 7 obtained a significant main effect of Trial Type (F(1, 15) = 26.33, p. < .01) and a significant Trial Type × Phase (last block of training vs. Test Day 7) interaction (F(1, 15) = 13.74, p. < .01). However, a nonsignificant Group × Trial type × Phase interaction (F(2, 15) = 1.04, p. >.37 confirmed that magnitude of the drop in FN discrimination performance from the end of training to Test Day 7 did not differ among the HE-DIO, HE-DR, and Chow groups.
Discrimination performance began to re-emerge over the last 2 probe tests for Groups HE-DR and Chow FN, but not for Group HE-DIO. ANOVA yielded a significant main effect of Trial Type (F(1, 15) = 25.42, p. < .01) and significant Test Day × Trial Type (F(2, 30) = 4.10, p < .05) and Test Day × Trial Type × Group (F(4, 30) = 3.12, p. < .05) interactions. Newman-Keuls tests revealed that for Group Chow responding on Tone+ trials was significantly higher compared to that on Light→Tone- trials on the 28 Day test and was higher on Tone+ trials compared to Light→Tone- on both the 21- and 28-day probe tests for Group HE-DR (ps. < .05). Differences between Tone+ and Light→Tone- trials failed to achieve significance on any probe test day for Group HE-DIO (smallest p-value > .46 for Test Day 21).
3.5. Blood-brain barrier (BBB) permeability
Figure 5 shows absolute fluorescence values for sodium fluorescein (NaFl) from the hippocampus, striatum and prefrontal cortex for rats in the HE-DIO, HE-DR, and Chow control groups. For all groups, intensity values in the hippocampus were higher compared to both of the other brain areas. Furthermore, the results indicate that intensity values in the hippocampus were higher for rats in the HE-DIO group compared to the HE-DR and Chow controls, with these latter groups differing little from each other. Figure 5 also shows that the intensity values in the prefrontal cortex were higher for the HE-DR group compared to both of the other groups. This finding implies that HE-diets may have adverse effects on the brain functions mediated by the prefrontal cortex in rats that are relatively insensitive to the effects of that diet on body weight and adiposity. However, we view this possibility cautiously as more recent unpublished studies in our lab have not replicated this outcome. Confirming these observations, ANOVA obtained significant main effects of Group (F(2, 15) = 4.97, p < 0.05) and Brain Areas (F(2, 30) = 685.35, p < 0.01) as well as a significant Group × Brain Area interaction (F(4, 30) = 12.75, p< 0.01). Across groups, intensity values in the hippocampus were significantly higher compared to both the striatum and the prefrontal cortex (ps. < 0.05). Furthermore, intensity values in the hippocampus were significantly higher for Group HE-DIO compared to groups HE-DR and Chow controls (ps. < 0.01). This latter pattern of differences was not obtained in either the striatum or prefrontal cortex.
Figure 5.
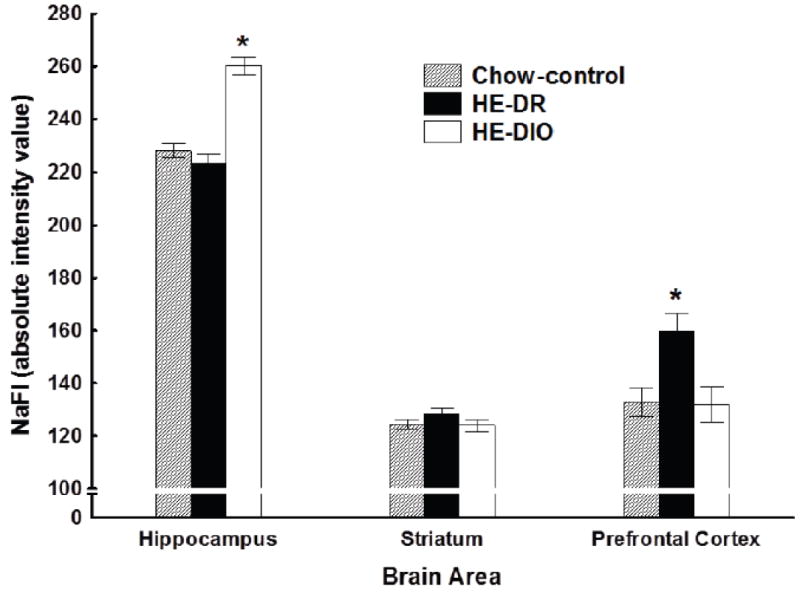
Concentrations of sodium fluorescein (NaFl) in hippocampus, striatum, and prefrontal cortex for rats maintained on ad libitum high-energy (HE) diet (Groups HE-DIO and HE-DR) or on ad libitum chow (Group Chow).
4. Discussion
The present results extend the findings of Kanoski et al., [4] by showing that (a) consuming a HE diet impaired post-learning performance of the serial FN discrimination problem without disrupting performance on a previously learned simple discrimination; (b) HE diet-induced impairment in FN performance emerged within 28 days following the initiation of the diet; (c) the effect of HE diet on FN discrimination performance was associated with level of sensitivity to the effects of HE diet on weight gain; and (d) the HE diet consumption was associated with an increased BBB permeability in select brain regions.
Specifically, rats that gained relatively high amounts of body weight, body fat, and ate more after being placed on a HE diet (Group HE-DIO) exhibited significantly poorer FN discrimination performance when compared to rats that gained significantly less weight and body fat and ate less when maintained on the same diet (Group HE-DR). Furthermore, while FN performance for the HE-DIO rats was also significantly impaired relative to rats maintained ad libitum on standard chow, the performance of HE-DR rats did not differ from these chow controls.
Although the rats in Group HE-DIO were impaired on the serial FN discrimination problem, they were not impaired relative to either Group HE-DR or Chow controls on the simple, CS+ vs. CS- discrimination. This pattern of results makes it difficult to account for Group HE-DIO’s impaired FN performance by appealing to nonspecific effects. For example, the unconditioned stimulus (sucrose pellets), response requirements (magazine photobeam interruption), and apparatus were the same for both the FN and simple discrimination problems. It is also noteworthy that the rats were tested on both types of problems within the same sessions. If the impairment in FN performance was a consequence of deficits in motivation, reward value, behavioral activation, sensory, motor, or other such nonspecific factors, it is difficult to explain why these deficits did not also reduce performance on the simple discrimination problem. Furthermore, when a manipulation (e.g., HE diet consumption) has different effects on performance in two different learning and memory tasks, one can almost always posit, post-hoc, that the performance difference might reflect an interaction between the manipulation and task complexity or cognitive load. However, for the present results this argument is made less persuasive by previous reports that while performance on Pavlovian serial FN tasks are impaired as a consequence of either hippocampal [6] or exposure to an HE-diet [4], performance of the converse serial feature-positive (Light→Tone+, Tone-) problem is not. Even though the two problems differ in their dependence on the hippocampus, there seems to be little a priori basis for claiming that the serial feature-negative (Light→Tone-, Tone+) problem differs appreciably from the serial feature-positive problem (Light→Tone+, Tone-) in terms of effort, complexity, or cognitive load.
Rather than inducing a nonspecific or global deficit in cognitive functioning, the results of this experiment are consistent with an HE diet-induced impairment in hippocampal function for rats in Group HE-DIO. Previous work showed that rats with hippocampal damage are less able to solve a serial FN problem compared to controls for which the hippocampus is intact [6]. In contrast, these lesions appear to have little or no disruptive effect on the performance of simple discrimination problems [6, 16]. The finding of impairment on a hippocampal-dependent discrimination problem but not on a similar concurrently tested hippocampal-independent problem provides evidence that the HE diet produced a specific deficit in hippocampal function for the rats in Group HE-DIO.
This interpretation draws support from our assays of blood-brain barrier (BBB) permeability. Specifically, for the rats in Group HE-DIO, concentration of sodium fluorescein (NaFl), a dye that does not cross the BBB freely, was significantly elevated in the hippocampus of Group HE-DIO compared to both Groups HE-DR and Chow. This pattern was not observed when dye concentration was measured in the striatum and prefrontal cortex of these same groups. It is tempting to speculate that the failure of the rats in Group HE-DIO to recover the FN discrimination during probe testing is likely associated with the effects of increased BBB permeability on hippocampal function.
An interesting feature of the present results is that discriminative responding was reduced on the simple discrimination problem and was abolished on the serial FN task for all groups during the early stages of testing under ad libitum feeding. Deprivation states have long been described as interoceptive contextual stimuli that comprise part of the background for learning about other events [17]. Given that memory retrieval is sensitive to changes in contextual cues [18, 19], including those provided by deprivation states [20], impaired performance by all rats at the outset of testing may have involved a change in contextual cues based on the shift from food deprivation at the end of training to ad libitum feeding during testing. Furthermore, some data suggest that contextual specificity of memory retrieval may depend on the nature or complexity of the discrimination problem [21, 22]. This may be why changing contextual cues related to deprivation state may have had greater effect on FN compared to simple discrimination performance.
Rats with hippocampal damage often show reduced context specificity compared to intact control rats [23]. Thus, the findings that all groups exhibited comparable performance decrements following shift from hunger to satiation at the outset of testing indicates that hippocampal functioning was comparable for all groups at that time. The finding that subsequent recovery of FN performance was impaired for rats in Group HE-DIO, but not for the other groups, suggests that HE diet-induced hippocampal dysfunction began to emerge for this group later in testing and this may have prevented reacquisition of the FN discrimination in the test context. No impairment in recovery from the smaller decrement shown in the simple discrimination task would be expected for Group HE-DIO if performance on that problem does not depend on the hippocampus.
The present results add to a number of epidemiological and experimental studies which suggest that common mechanisms may link obesity and metabolic disease to the emergence of cognitive dysfunction [24-26]. Along with obesity, excessive intake of HE diet is associated with impaired glucoregulation, insulin resistance, reduced neurotrophic factors, and inflammation, among other adverse consequences. Each of these effects could interfere with hippocampal functioning. For example, a variety of data indicate that chronic hyperinsulinemia, concomitant with insulin resistance and poor glycemic control, is associated with decrements in hippocampal-dependent learning and memory and may even set the stage for Alzheimer’s disease or Alzheimer’s-like pathologies [27]. Similarly, consumption of HE diet by rats is accompanied by reduced levels of hippocampal brain-derived neurotrophic factor (BDNF) and reduced performance on hippocampal-dependent spatial [28] and nonspatial [29] learning and memory problems. BDNF has also been postulated to contribute to energy homeostasis by exerting an inhibitory influence on energy intake [30].
HE diet-induced feeding is also associated with low-grade inflammation in peripheral tissues and the hypothalamus [31]. Deficits in memory function and hippocampal inflammation have also been reported in a mouse model of obesity [32]. Furthermore, findings from humans and mouse models [33] suggest that hippocampal inflammation may play a critical role in the etiology of Alzheimer’s Disease. An interesting possibility is that intake of HE-diet contributes to hippocampal inflammation as a consequence of the type of increased BBB permeability that was observed in the present study and by [4].
Understanding the mechanisms that underlie the effects of HE diet-induced obesity on BBB permeability and inflammation is of paramount importance. Recent evidence suggests that elevations in peripheral proinflammatory cytokine compromise BBB function making the brain and hippocampus more accessible to these inflammatory agents as well as to other harmful substances (see [34]). Other findings suggest that elevated levels of proinflammatory cytokines interfere with leptin transport across the BBB (e.g., [35]). Leptin is released into circulation from adipose tissue in direct proportion to the size of the adipose stores. Leptin crosses the BBB via specific transporter mechanisms which enable it enter the brain to participate in the inhibitory control of intake and body adiposity (e.g., [36]). Leptin signaling is impaired in obese humans and rodents, and it appears that at least some of this impairment may be the result of interference with BBB transport ([37, 38]). Also important, impaired leptin transport has been linked to reduced hippocampal-dependent learning and memory function [39]. One recent study found that leptin administered directly into the ventral hippocampus inhibited both food intake and retrieval of food-related memories in rats [40]. Thus, it is conceivable that for the HE-DIO rats in the present study, HE-diet promoted both weight gain and hippocampal-dependent cognitive impairment as a consequence of inflammation-induced BBB dysfunction. Whether these effects are based on increased access to the hippocampus of cytokines or other substances or on interference with leptin transporter and/or other mechanisms that mediate hippocampal function remains to be determined.
It is clear that consuming a HE diet can initiate a chain of events that involves excessive caloric intake, myriad physiological changes and impairment of hippocampal-dependent learning and memory. But it seems that understanding of the causal links in this chain will be incomplete until mechanisms are specified that describe how intake of HE diet initiates such changes in behavior and physiology. Our working model proposes that hippocampal dysfunction may be nearer to the beginning rather than to the end of this chain [1, 13]. Indeed, within a vicious-cycle approach, differences in hippocampal-dependent learning and memory processes may be a cause as well as an effect of increased sensitivity to the adverse effects HE diets on body weight and metabolic activity.
Specifically, this model suggests that intake of HE diet may produce impairments in hippocampal functioning which promotes further increased intake of HE diet. This initiates a vicious cycle of progressively increasing intake, body weight, and body adiposity concomitant with the progressive deterioration of the physiological controls of energy regulation and hippocampal-dependent cognitive processes. This model draws support from findings that (a) selective lesions of the hippocampus can promote food intake and body weight gain [41]; (b) human [42] and nonhuman [43] animals with hippocampal damage are impaired in utilizing cues arising from hunger and satiation to control their intake and appetitive behavior; (c) rats with hippocampal damage exhibit elevated levels of conditioned responding for food [44, 45] and reduced resistance to extinction of previously food-reinforced responses [46-48]; (d) relative to standard chow, consuming HE diets produces impairments in hippocampal-dependent learning and memory which occur prior to the emergence of significant differences in body weight [37, 38].
The results of the present experiment are consistent with this model in that the effects of HE diet on intake, body, weight, and adiposity were attenuated in rats that failed to exhibit hippocampal dependent deficits in discrimination performance, thereby leaving open the possibility that hippocampal dysfunction contributed to energy regulatory disturbances (also see [49]). Determining why these regulatory capacities differ among DIO and DR rats and their human counterparts will be key to improving understanding of both obesity and cognitive dysfunction. Our findings encourage the idea that comparing the hippocampal function of obesity-prone and obesity resistance rats may provide a new avenue on which to approach the links between obesity and cognitive decline.
Highlights.
Diet and obesity phenotype interact to alter hippocampal-dependent cognitive function
High-energy diet impaired retention of a hippocampal-dependent discrimination task
This impairment was exhibited by diet-induced obese, but not by diet-resistant rats
Blood-brain barrier (BBB) integrity was compromised for diet-induced obese rats
Obesity and cognitive dysfunction may be linked to diet-induced BBB disruption
Acknowledgments
The authors thank Gerard P. Smith for pointing out the importance of comparing obesity-prone and obesity-resistance rats with respect to their cognitive abilities. The authors also thank Lindsey Schier for her contributions to discussions during the course of this research. This work was supported by grants P01 HD052112 and R01 HD29792 from the NICHD, and grants ES008146 and ES017055 from the NIEHS. The funding sources were not involved with study design, with the collection, analysis and interpretation of data, with the writing of the report, or with the decision to submit the article for publication.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Kanoski SE, Davidson TL. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol Behav. 2011;103:59–68. doi: 10.1016/j.physbeh.2010.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Francis HM, Stevenson RJ. Higher reported saturated fat and refined sugar intake is associated with reduced hippocampal-dependent memory and sensitivity to interoceptive signals. Behav Neurosci. 2011;125:943–55. doi: 10.1037/a0025998. [DOI] [PubMed] [Google Scholar]
- 3.Edwards LM, Murray AJ, Holloway CJ, Carter EE, Kemp GJ, Codreanu I, et al. Short-term consumption of a high-fat diet impairs whole-body efficiency and cognitive function in sedentary men. Faseb J. 2011;25:1088–96. doi: 10.1096/fj.10-171983. [DOI] [PubMed] [Google Scholar]
- 4.Kanoski SE, Zhang YS, Zheng W, Davidson TL. The Effects of a High-Energy Diet on Hippocampal Function and Blood-Brain Barrier Integrity in the Rat. J Alzheimers Dis. 2010;21:207–19. doi: 10.3233/JAD-2010-091414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Morell JR, Davidson TL. Transfer across unconditioned stimuli in serial feature discrimination training. J Exp Psychol -Anim Behav Process. 2002;28:83–96. [PubMed] [Google Scholar]
- 6.Holland PEC, Lamoureux JA, Han JS, Gallagher M. Hippocampal lesions interfere with Pavlovian negative occasion setting. Hippocampus. 1999;9:143–57. doi: 10.1002/(SICI)1098-1063(1999)9:2<143::AID-HIPO6>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 7.Freeman L, Granholm AC. Long-Term Treatment With a High-Fat/High-Cholesterol Diet in Young and Aged Rats: Inflammatory Response, Blood-Brain Barrier Breakdown, and Cognitive Impairment. Cell Transplant. 2011;20:556. [Google Scholar]
- 8.Wolburg H, Lippoldt A. Tight junctions of the blood-brain barrier: development, composition and regulation. Vascul Pharmacol. 2002;38:323–37. doi: 10.1016/s1537-1891(02)00200-8. [DOI] [PubMed] [Google Scholar]
- 9.Devan BD, Hong NS, McDonald RJ. Parallel associative processing in the dorsal striatum: Segregation of stimulus-response and cognitive control subregions. Neurobiol Learn Mem. 2011;96:95–120. doi: 10.1016/j.nlm.2011.06.002. [DOI] [PubMed] [Google Scholar]
- 10.Pennartz CMA, Ito R, Verschure P, Battaglia FP, Robbins TW. The hippocampal-striatal axis in learning, prediction and goal-directed behavior. Trends Neurosci. 2011;34:548–59. doi: 10.1016/j.tins.2011.08.001. [DOI] [PubMed] [Google Scholar]
- 11.Rushworth MFS, Noonan MP, Boorman ED, Walton ME, Behrens TE. Frontal Cortex and Reward-Guided Learning and Decision-Making. Neuron. 2011;70:1054–69. doi: 10.1016/j.neuron.2011.05.014. [DOI] [PubMed] [Google Scholar]
- 12.Madsen AN, Hansen G, Paulsen SJ, Lykkegaard K, Tang-Christensen M, Hansen HS, et al. Long-term characterization of the diet-induced obese and diet-resistant rat model: a polygenetic rat model mimicking the human obesity syndrome. J Endocrinol. 2010;206:287–96. doi: 10.1677/JOE-10-0004. [DOI] [PubMed] [Google Scholar]
- 13.Davidson TL, Kanoski SE, Schier LA, Clegg DJ, Benoit SC, Davidson TL, et al. A potential role for the hippocampus in energy intake and body weight regulation. Current Opinion in Pharmacology. 2007;7:613–6. doi: 10.1016/j.coph.2007.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Davidson TL, Kanoski SE, Walls EK, Jarrard LE, Kanoski SE, Walls EK, et al. Memory inhibition and energy regulation. Physiol Behav. 2005;86:731–46. doi: 10.1016/j.physbeh.2005.09.004. [DOI] [PubMed] [Google Scholar]
- 15.Choi BS, Zheng W. Copper transport to the brain by the blood-brain barrier and blood-CSF barrier. Brain Research. 2009;1248:14–21. doi: 10.1016/j.brainres.2008.10.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Good M, Honey RC. Conditioning And Contextual Retrieval In Hippocampal Rats. Behav Neurosci. 1991;105:499–509. doi: 10.1037//0735-7044.105.4.499. [DOI] [PubMed] [Google Scholar]
- 17.Hirsh R. The hippocampus and contextual retrieval of information from memory: a theory. Behav Biol. 1974;12:421–44. doi: 10.1016/s0091-6773(74)92231-7. [DOI] [PubMed] [Google Scholar]
- 18.Bouton ME, Moody EW. Memory processes in classical conditioning. Neurosci Biobehav Rev. 2004;28:663–74. doi: 10.1016/j.neubiorev.2004.09.001. [DOI] [PubMed] [Google Scholar]
- 19.Urcelay GP, Miller RR. Two Roles of the Context in Pavlovian Fear Conditioning. J Exp Psychol -Anim Behav Process. 2010;36:268–80. doi: 10.1037/a0017298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kennedy PJ, Shapiro ML. Retrieving memories via internal context requires the hippocampus. J Neurosci. 2004;24:6979–85. doi: 10.1523/JNEUROSCI.1388-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nakajima S. Contextual Control Of Pavlovian Feature-Positive And Feature-Negative Discriminations. Anim Learn Behav. 1994;22:34–46. [Google Scholar]
- 22.Nelson JB. Context specificity of excitation and inhibition in ambiguous stimuli. Learning and Motivation. 2002;33:284–310. [Google Scholar]
- 23.Holland PC, Bouton ME. Hippocampus and context in classical conditioning. Current Opinion in Neurobiology. 1999;9:195–202. doi: 10.1016/s0959-4388(99)80027-0. [DOI] [PubMed] [Google Scholar]
- 24.Bruce-Keller AJ, Keller JN, Morrison CD. Obesity and vulnerability of the CNS. Biochim Biophys Acta-Mol Basis Dis. 2009;1792:395–400. doi: 10.1016/j.bbadis.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Craft S. The role of metabolic disorders in Alzheimer disease and vascular dementia: two roads converged. Arch Neurol. 2009;66:300–5. doi: 10.1001/archneurol.2009.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Parrott MD, Greenwood CE. Dietary influences on cognitive function with aging - From high-fat diets to healthful eating. In: Weller NJ, Rattan SIS, editors. Healthy Aging and Longevity. Oxford: Blackwell Publishing; 2007. pp. 389–97. [DOI] [PubMed] [Google Scholar]
- 27.Cholerton B, Baker LD, Craft S. Insulin resistance and pathological brain ageing. Diabet Med. 2011;28:1463–75. doi: 10.1111/j.1464-5491.2011.03464.x. [DOI] [PubMed] [Google Scholar]
- 28.Molteni R, Barnard RJ, Ying Z, Roberts CK, Gomez-Pinilla F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience. 2002;112:803–14. doi: 10.1016/s0306-4522(02)00123-9. [DOI] [PubMed] [Google Scholar]
- 29.Kanoski SE, Meisel RL, Mullins AJ, Davidson TL. The effects of energy-rich diets on discrimination reversal learning and on BDNF in the hippocampus and prefrontal cortex of the rat. Behavioural Brain Research. 2007;182:57–66. doi: 10.1016/j.bbr.2007.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Noble EE, Billington CJ, Kotz CM, Wang C. The lighter side of BDNF. Am J Physiol Regul Integr Comp Physiol. 2011;300:R1053–69. doi: 10.1152/ajpregu.00776.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Thaler JP, Schwartz MW. Minireview: Inflammation and Obesity Pathogenesis: The Hypothalamus Heats Up. Endocrinology. 2010;151:4109–15. doi: 10.1210/en.2010-0336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dinel AL, Andre C, Aubert A, Ferreira G, Laye S, Castanon N. Cognitive and Emotional Alterations Are Related to Hippocampal Inflammation in a Mouse Model of Metabolic Syndrome. Plos One. 2011:6. doi: 10.1371/journal.pone.0024325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cuello AC, Ferretti MT, Leon WC, Iulita MF, Melis T, Ducatenzeiler A, et al. Early-Stage Inflammation and Experimental Therapy in Transgenic Models of the Alzheimer-Like Amyloid Pathology. Neurodegener Dis. 2010;7:96–8. doi: 10.1159/000285514. [DOI] [PubMed] [Google Scholar]
- 34.Fung A, Vizcaychipi M, Lloyd D, Wan Y, Ma D. Central nervous system inflammation in disease related conditions: Mechanistic prospects. Brain Research. 2012 doi: 10.1016/j.brainres.2012.01.061. in press. [DOI] [PubMed] [Google Scholar]
- 35.Hsuchou H, Kastin AJ, Tu H, Abbott NJ, Couraud PO, Pan WH. Role of astrocytic leptin receptor subtypes on leptin permeation across hCMEC/D3 human brain endothelial cells. J Neurochem. 2010;115:1288–98. doi: 10.1111/j.1471-4159.2010.07028.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Woods SC. The Control of Food Intake: Behavioral versus Molecular Perspectives. Cell Metab. 2009;9:489–98. doi: 10.1016/j.cmet.2009.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Banks WA, DiPalma CR, Farrell CL. Impaired transport of leptin across the blood-brain barrier in obesity. Peptides. 1999;20:1341–5. doi: 10.1016/s0196-9781(99)00139-4. [DOI] [PubMed] [Google Scholar]
- 38.Price TO, Farr SA, Yi X, Vinogradov S, Batrakova E, Banks WA, et al. Transport across the Blood-Brain Barrier of Pluronic Leptin. Journal of Pharmacology and Experimental Therapeutics. 2010;333:253–63. doi: 10.1124/jpet.109.158147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Farr SA, Banks WA, Morley JE. Effects of leptin on memory processing. Peptides. 2006;27:1420–5. doi: 10.1016/j.peptides.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 40.Kanoski SE, Hayes MR, Greenwald HS, Fortin SM, Gianessi CA, Gilbert JR, et al. Hippocampal Leptin Signaling Reduces Food Intake and Modulates Food-Related Memory Processing. Neuropsychopharmacology. 2011;36:1859–70. doi: 10.1038/npp.2011.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Davidson TL, Chan K, Jarrard LE, Kanoski SE, Clegg DJ, Benoit SC, et al. Contributions of the hippocampus and medial prefrontal cortex to energy and body weight regulation. Hippocampus. 2009;19:235–52. doi: 10.1002/hipo.20499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hebben N, Corkin S, Eichenbaum H, Shedlack K. Diminished Ability To Interpret And Report Internal States After Bilateral Medial Temporal Resection - Case Hm. Behav Neurosci. 1985;99:1031–9. doi: 10.1037//0735-7044.99.6.1031. [DOI] [PubMed] [Google Scholar]
- 43.Davidson TL, Kanoski SE, Chan K, Clegg DJ, Benoit SC, Jarrard LE. Hippocampal lesions impair retention of discriminative responding based on energy state cues. Behav Neurosci. 2010;124:97–105. doi: 10.1037/a0018402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Clifton PG, Vickers SP, Somerville EM. Little and often: Ingestive behavior patterns following hippocampal lesions in rats. Behav Neurosci. 1998;112:502–11. doi: 10.1037//0735-7044.112.3.502. [DOI] [PubMed] [Google Scholar]
- 45.Murphy RA, McDonald RJ, Baker AG. Lesions of the hippocampus or fornix do not interfere with the relative validity effect on a discrete stimulus or the context. Behavioural Brain Research. 1998;92:55–66. doi: 10.1016/s0166-4328(97)00125-3. [DOI] [PubMed] [Google Scholar]
- 46.Benoit SC, Davidson TL, Chan KH, Trigilio T, Jarrard LE. Pavlovian conditioning and extinction of context cues and punctate CSs in rats with ibotenate lesions of the hippocampus. Psychobiology. 1999;27:26–39. [Google Scholar]
- 47.Kanoski SE, Davidson TL. Different patterns of memory impairments accompany short- and longer-term maintenance on a high-energy diet. Journal of Experimental Psychology: Animal Behavior Processes. 2010;36:313–9. doi: 10.1037/a0017228. [DOI] [PubMed] [Google Scholar]
- 48.Murray AJ, Knight NS, Cochlin LE, McAleese S, Deacon RM, Rawlins JN, et al. Deterioration of physical performance and cognitive function in rats with short-term high-fat feeding. FASEB J. 2009;23:4353–60. doi: 10.1096/fj.09-139691. [DOI] [PubMed] [Google Scholar]
- 49.Smith E, Hay P, Campbell L, Trollor JN. A review of the association between obesity and cognitive function across the lifespan: implications for novel approaches to prevention and treatment. Obes Rev. 2011;12:740–55. doi: 10.1111/j.1467-789X.2011.00920.x. [DOI] [PubMed] [Google Scholar]