

Abstract
Pancreatic cancer is the fourth most common cancer to cause death due to advanced stage at diagnosis and poor response to current treatment. Autophagy is the lysosome-mediated degradation pathway which plays a critical role in cellular defense, quality control, and energy metabolism. Targeting autophagy is now an exciting field for translational cancer research, as autophagy dysfunction is among the hallmarks of cancer. Pancreatic tumors have elevated autophagy under basal conditions when compared with other epithelial cancers. This review describes our current understanding of the interaction between autophagy and pancreatic cancer development, including risk factors (e.g., pancreatitis, smoking, and alcohol use), tumor microenvironment (e.g., hypoxia and stromal cells), and molecular biology (e.g., K-Ras and p53) of pancreatic cancer. The importance of the HMGB1-RAGE pathway in regulation of autophagy and pancreatic cancer is also presented. Finally, we describe current studies involving autophagy inhibition using either pharmacological inhibitors (e.g., chloroquine) or RNA interference of essential autophagy genes that regulate chemotherapy sensitivity in pancreatic cancer. Summarily, autophagy plays multiple roles in the regulation of pancreatic cancer pathogenesis and treatment, although the exact mechanisms remain unknown.
Keywords: Autophagy, pancreatic cancer, oncogene, hypoxia, pancreatitis, HMGB1, RAGE, p53, HIF1α, AMPK
Introduction
Pancreatic cancer is the fourth leading cause of cancer death in the USA for both men and women; 30,500 new cases will occur annually [1]. It is responsible for 5% of all cancer-related deaths; the overall five year survival rate is 3~5% because of advanced stage at diagnosis and poor response to current treatment [2]. Eighty percent of all pancreatic cancers are pancreatic ductal adenocarcinoma (PDAC) [3,4]. Morphologically, PDAC progresses from noninvasive pancreatic lesions termed “Pancreas Intraepithelial Neoplasias” (PanINs) (Figure 1) [5]. PanINs show glandular patterns with ductlike structures and varying degrees of cellular atypia and differentiation. PanINs undergo a neoplastic transformation ranging from early mucinous change (PanIN-1) to frank carcinoma in situ (PanIN-3). Genetically, PDAC progresses as a result of the accumulation of genetic alterations in cellular cancer-causing genes, such as encoding of the small GTPase protein K-Ras and tumor suppressors such as INK4A /p16, p53, BRCA2, and DPC4/SMAD4 (Figure 1) [3]. Of these genetic alterations, mutations of the K-Ras gene occur in over 90% of pancreatic carcinomas and are proposed to be the initiating genetic lesion in PDAC. Epidemiologically, factors such as smoking, alcohol use, diet, obesity, and chronic pancreatitis can increase the risk of acquiring the genetic mutations that may potentially result in pancreatic cancer [6]. Moreover, developmental regulatory pathways, in particular the Wnt, Notch, and Hedgehog pathways, are often activated in PDAC [7]. In addition, deregulation of high mobility group protein box 1 (HMGB1) - the receptor for advanced glycation end products (RAGE) signaling pathway [8,9] and the tumor microenvironment of PDAC (e.g., inflammation and stromal cells) [10,11] are closely associated with pancreatic cancer development. Autophagy, a lysosomal degradation pathway, has been recently demonstrated as active in PDAC and regulates chemotherapy sensitivity during pancreatic treatment [12-19]. Our findings suggest that the HMGB1-RAGE pathway plays critical roles in induction of autophagy, and inhibition of HMGB1-RAGE leads to increased apoptosis and decreased autophagy in pancreatic cancer cells [9,20-23]. Current studies aim to fully understand the emerging role of autophagy in pancreatic cancer formation and progression and determine whether targeting autophagy represents a valid approach to pancreatic cancer treatment in the clinic.
Figure 1.
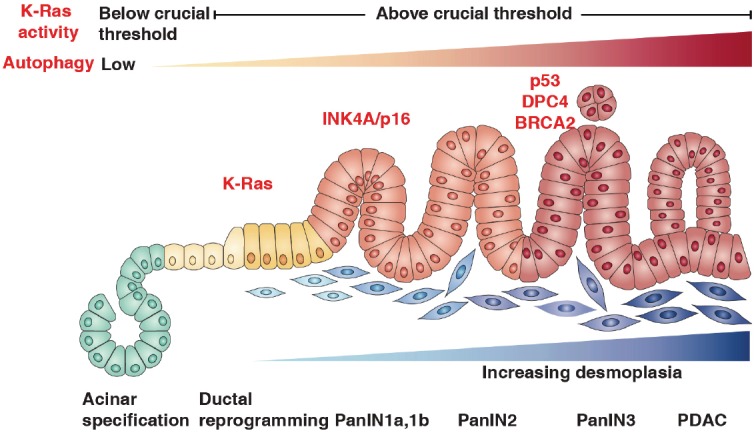
Progression model for pancreatic ductal adenocarcinoma. PDAC progresses from PanINs. The progression from histologically normal ductal epithelium to low-grade PanIN to high-grade PanIN is associated with the accumulation of specific genetic changes. Early changes include K-ras mutations; intermediate changes include INK4A/p16 mutations; and changes associated with either in situ or early invasive cancer include p53, BRCA2, and DPC4/SMAD4. Elevated expression of autophagy in cancer cells has been implicated in the development of PDAC.
Autophagy and cancer
Lysosomes are cellular organelles that contain acid hydrolase enzymes to break down waste materials and cellular debris. Autophagy is a lysosomal degradation pathway by which cytoplasmic components, such as damaged organelles and unused long-lived proteins, are digested within lysosome [24]. There are at least three distinct types of autophagy pathways, including macroautophagy, chaperone-mediated autophagy, and microautophagy [25]. Macroautophagy (referred to hereafter as autophagy) is a dynamic process by which cytoplasmic components are engulfed by an isolation membrane structure termed the phagophore, and then are sequestered in a double or multimembrane structure termed the autophagosome, which is finally delivered to the lysosome to form an autolysosome where their contents are degraded (Figure 2). Potential sources for the phagophore include the Golgi complex, endosomes, ER, mitochondria, and plasma membrane [26]. Although nonselective autophagy is the main autophagic degradation process, evidence of a more selective degradation of particular organelles (e.g. mitochondria) and intracellular bacteria/virus has been found. These processes have been termed mitophagy [27], and xenophagy [28,29], respectively.
Figure 2.

Autophagy and apoptosis: process and function. Autophagy and apoptosis may be triggered by common stimuli. As programmed cell survival, autophagy is an intracellular bulk degradation system, through which cytoplasmic component is delivered to lysosomes to be degraded. The main process of autophagy includes formation and maturation of the phagophore, autophagosome, and autolysosome. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing to LC3-II and can be degraded by autolysosome after binding adaptor protein p62. Autophagy provides amino acids and fatty acids for the synthesis of protein and ATP. In contrast, apoptosis is programmed cell death. There are two main pathways to control apoptosis. The extrinsic pathway is mediated by death receptors such as Fas/CD95. The intrinsic mitochondrial pathway is controlled by Bcl-2 family proteins such as Bax and other regulators, such as p53. Caspase 3 is a caspase protein that is activated by caspase 8 and caspase 9, and in turn, mediates substrate cleavage and cell destruction.
The molecular machinery that controls the autophagy pathway is extremely complex [30,31]. Over 30 different autophagy-related genes (ATGs) have been identified in yeast, and many of these are functionally conserved in higher eukaryotes. Autophagy is induced by the inhibition of mammalian target of rapamycin (mTOR) [32], resulting in the formation of class III phosphoinositide 3-kinase (PI3KC3) -Beclin 1 (also known as Atg6 in yeast) core complex [33], which mediates nucleation of the phagophore in autophagy. After that, two ubiquitin-like conjugation systems (Atg8 and Atg5-Atg12-Atg16) are essential for autophagosome formation. Microtubule-associated protein 1 light chain 3 (LC3), a homologue of Atg8 in yeast, is widely used to monitor autophagy by western blot analysis of turnover of LC3-I to LC3-II, and imaging analysis of LC3 puncta formation under co-treatment with inhibitors of early (e.g., 3-methyladenine) and late (e.g., chloroquine and bafilomycin-A1) autophagy [34]. The lysosomal-associated membrane protein 1 (LAMP1) and LAPM2 and the small GTPase RAB7 are involved in autolyosomal formation.
As programmed cell survival mechanism, autophagy is critical in maintaining a range of normal human physiological processes, such as cellular homeostasis, energetic balance, development, and cellular defense against danger signals [35,36]. Conversely, autophagy is a double-aged sword, implicated in the pathogenesis of diseases including cancer, neurodegenerative diseases, aging, muscle diseases, infection, and immunity diseases [37-41].
The role of autophagy in cancer is extremely complex [42-46]. Decreased, increased, and mutant ATGs expressions are detected in various tumors. Autophagy was initially as an antioncogenic mechanism in the tumor, because heterozygous Beclin1+/- mice develop spontaneous tumors [47]. Moreover, deletion of other autophagy genes such as UVRAG [48], Atg5 [49] and Bif [50] in mice also increase tumorigenesis. It is believed that autophagy partly pro-motes cancer development through its ability to shape inflammatory reaction [51], metabolic requirements [52], and oxidative stress [53]. In addition, autophagy has a pro-tumor growth effect in some cancers such as pancreatic [12] and BCR-Abl-mediated leukemogenesis [54]. Thus, autophagy may play opposite roles in the early and late cancer development stages. One possible explanation for this difference is that the content of metabolic stress, immune response, and microenvironment is different. Autophagy and apoptosis share common stimuli, signal pathways, and regulators (Figure 2) [55]. Interestingly, apoptosis, as a programmed cell death process, also has dual roles in regulation of tumorigenesis [56]. The inter-relationship between autophagy and apoptosis might decide tumor cell’s fate. In many cases, blockage of autophagy sensitizes tumor cells to anticancer therapy, including chemotherapy, radiation, and immunotherapy [57]. However, under certain conditions such as apoptosis deficiency, autophagy can also be pro-death, termed "autophagic cell death" [58,59].
Autophagy and pancreatic cancer development
Pancreatic tumors have more elevated autophagy under basal conditions than those of other epithelial cancers [60]. A clinicopathological study of 71 archival pancreatic cancer tissues demonstrated that autophagy is activated in pancreatic cancer cells and correlates with poor patient outcome [19]. They found a stronger LC3 intensity expression level in the peripheral area of the pancreatic cancer tissue by immunohistochemical staining with LC3 antibody. Moreover, strong expression of LC3 is associated with enhanced expression of the hypoxia marker carbonic anhydrase IX at the peripheral area, which correlates with poor outcome and a short disease-free period. Another study showed that LC3 is low or absent in normal exocrine pancreas and in low-grade PanIN-1 and PanIN-2 lesions, whereas staining is upregulated and exhibits a vesicular staining pattern in all high-grade PanIN-3 and PDAC (Figure 1). These studies suggested that autophagy may contribute to pancreatic tumor progress. Basal autophagy is significantly elevated compared with non-transformed human pancreatic ductal and other tumor cells, confirmed by monitoring LC3 spot formation in PDAC cell lines and primary tumors [12]. Moreover, knockdown of autophagy genes such as ATG5 and ATG3 by RNA interference significantly inhibits cell growth and colony formation of PDAC cell lines in vitro [12]. Furthermore, chloroquine, an anti-malaria drug with autophagy inhibition activity, suppressed pancreatic tumor growth in xenograft mice [12]. These studies suggest that autophagy can contribute to pancreatic tumor growth and development. In vitro, inhibition of autophagy in pancreatic tumor cells augments production of reactive oxygen species, increases DNA damage, and limits effective metabolism via decreased mitochondrial oxidative phosphorylation [12], suggesting a unique role of autophagy in pancreatic tumorigenesis through regulation of mitochondrial function and energy production. Notably, chloroquine has multiple functions independent from autophagy [61]. Thus, specific autophagic gene deficient mice will confirm whether autophagy promotes pancreatic tumorigenesis. Our recent study indicates that global knockout of RAGE in mice inhibits interleukin-6 (IL-6) / signal transducer and activator of transcription 3 (STAT3) induced autophagy and K-Ras mutant driven PanINs development [62].
Autophagy and pancreatic beta-cell
The pancreas is comprised of separate functional units that regulate two major physiological processes: digestion and glucose metabolism. Blood glucose metabolism is controlled by the pancreatic beta-cells, which are located in the islets of Langerhans. Deficiency in the number of pancreatic beta cells causes diabetes. Beta cell growth is disordered in pancreatic cancer [63]. K-ras in islet cells leads to tumor only with inflammation [64,65]. Recent studies suggest that basal levels of autophagy are also required for maintenance of beta cells homeostasis in the pancreas [66]. Beta cell -specific Atg7 knockout mice had increased apoptosis and reduced proliferation of beta cells, which resulted in reduced beta cell mass and pancreatic insulin content [66]. Moreover, pancreatic islets from mice with Beta cell-specific Atg7 knockout mice had decreased adenosine triphosphate (ATP) production [66]. Islet cell destruction had functional consequences, because the remaining cells were depleted of insulin and the mice developed hyperglycemia that was worsened by a high-fat diet [67]. Autophagy and vacuole membrane protein 1 (VMP1) expression are early cellular events in experimental diabetes [68].
Autophagy and risk factors for pancreatic cancer
Acute pancreatitis precedes chronic pancreatitis, which is a significant risk factor for the development of pancreatic carcinoma [69,70]. Several experimental models of severe acute pancreatitis have been produced in rats or mice by administration of cerulein, bile acid (e.g. taurocholate) or L-arginine, or by feeding mice a choline-deficient, ethionine supplemented diet [71]. Cerulein-induced pancreatitis promotes oncogenic transformation of adult pancreatic acinar cells by oncogenic K-Ras in a mice model [10]. In the early stage of acute pancreatitis, autodigestion is provoked by inappropriate activation of proteolytic enzymes within the acinar cells. These early events are followed by the generation of local inflammatory reaction, as well as systemic inflammatory response syndrome. Recent studies suggest that the impaired autophagic and lysosomal dysfunctions mediate key pathologic responses of pancreatitis, such as acinar cell vacuolation and trypsinogen activation [72-78]. Pancreatitis stimulates autophagy induction and at the same time impairs late stages of autophagy [78]. Inhibition of autophagy by genetic or pharmacologic means markedly improved cerulein-induced pancreatitis. It was indicated that autophagic flux is reduced in pancreatitis due to deficient lysosomal degradation caused by impaired cathepsin processing and activation, and decreased levels of LAMP-1 and -2 [72]. Moreover, there is a decrease in the fully mature (“double-chain”) cathepsin forms and accumulation of their immature forms in pancreatitis [78]. It is unclear how cathepsin processing becomes defective or how this relates to reported decreases in other lysosomal proteins. It is indicated that pancreatitis-like injury spontaneously develops in LAMP-2 knockout mice [79]. Of note, LAMP-2 is up-regulated in many cancer cell lines and human tumors, including pancreatic cancer [80]. Knockdown of LAMP-2 resulted in tumor shrinkage by regulation of p53-mediated glycolytic metabolism [81].
Currently, there are no studies exploring the direct relationship between autophagy and other risk factors factor for pancreatic cancer, only pancreatitis. Alcohol use, smoking, and diet are all implicated as possible pancreatic cancer risk factors. Autophagy is induced by alcohol and prevents liver injury [82]. Cigarette smoke-induced autophagy is involved in early stages of chronic obstructive pulmonary disease progression [83]. A high-fat diet stimulates autophagy in β cell and regulates glucose metabolism [67]. In addition, high-fat diet-induced hepatic steatosis was associated with autophagy [84]. Thus it is interesting to explore whether these factors also induce autophagy in the pancreatic cancer cell.
Autophagy and microenvironment of pancreatic cancer
Accumulating evidence suggests that the tumor microenvironment is in a hypoxic condition [85,86]. The tumor microenvironment includes immune cells (lymphocytes, natural killer cells, and antigen presenting cells), stromal cells (including fibroblasts/stellate cells), and the vasculature (pericytes and the endothelium). The tumor microenvironment directs pancreatic tumor growth [87]. Stromal cells stimulate cancer cell growth and invasion through the chemokine-chemokine receptor axis. The tumor vasculature allows nutrients and oxygen uptake by tumors and tumor-infiltrating immune cells and damage associated molecular pattern molecules (DAMPs) anergize the immune effectors.
Autophagy can be induced by hypoxia (Figure 3A). Hypoxia-inducible factor 1 (HIF) is a master regulator of oxygen homeostasis. In many case, hypoxia-induced autophagy is mediated through HIF-1α induction of Bcl-2/adenovirus E1B 19-kDa protein-interacting protein 3 (Bnip3) and BNIP3 like protein (BNIP3L) in several cells, including cancer cells [88-90]. Bnip3 is a member of the Bcl-2 homology domain 3-only subfamily of proapoptotic Bcl-2 proteins and is associated with cell death. However, another study suggests that hypoxia-induced autophagy in tumor cells is AMP-activated protein kinase (AMPK)-mTOR dependent, but not HIF-1α, BNIP3, and BNIP3L dependent [91]. In addition, autophagy may be induced during hypoxia as a result of signals generated by the unfolded protein response in the endoplasmic reticulum. The PKR-like endoplasmic reticulum kinase (PERK)-activating transcription factor 4 (ATF4) pathway is involved in unfolded protein response associated autophagy. Recent studies demonstrate that autophagy mediates survival of pancreatic tumor-initiating cells in a hypoxic microenvironment [92]. They found that physiological enhanced levels of autophagy render pancreatic cancer stem-like cells resistant to induction of apoptosis by hypoxia and starvation [92]. In contrast, inhibition of autophagy induces death of pancreatic cancer stem-like cells and inhibits self-renewal potential [92].
Figure 3.

Pathways of autophagy induction by hypoxia and mutant RAS. (A) During hypoxia, autophagy is activated by sensors that detect low oxygen, unfolded proteins, and energy depletion. Low oxygen induces autophagy involving the HIF1α-mediated up-regulation of BNIP3 and BNIP3L. They compete with Bcl-2 for interaction with Beclin 1, and orient Beclin 1 to autophagosomes. AMPK is a major regulator of energy homeostasis. The kinase is sensitive to the AMP/ATP ratio: binding of AMP activates AMPK and induces phosphorylation by the tumor suppressor LKB1. Activation of AMPK inhibits mTOR-dependent signaling. Inhibition of mTOR induces autophagy. In addition, autophagy may be induced during hypoxia as a result of signals generated by the PERK-ATF4 mediated unfolded protein response in the endoplasmic reticulum. (B) The induction of autophagy by oncogenes such as mutant Ras is importance in tumorigenesis. Ras mutant activates the ROS-dependent JNK signaling pathway and upregulates the Atg5 gene to induce autophagy. Under certain circumstances, Ras mutant activates the MEK/ERK signaling pathway and upregulates the Beclin1 gene to induce autophagic cell death.
Stromal cells have recently gained much attention as a critical determinant of tumor progression and clinical outcome in pancreatic cancer [93]. Several studies have indicated that autophagy in cancer associated fibroblasts promotes tumor cell survival in the tumor stromal microenvironment [94,95]. Caveolae, as a versatile signaling molecule, can bud from the plasma membrane and function in cellular processes such as endocytosis and the transcytosis of molecules. Loss of caveolae-1 expression in cancer-associated fibroblasts results in an activated tumor microenvironment, thereby driving early tumor recurrence, metastasis, and poor clinical outcome in breast and prostate cancers [96,97]. However, overexpression of caveolin-1 correlates with tumor progression markers in PDAC [98]. Hypoxia-induced autophagy and activation of the NF-κB pathway is indicated to drive caveolae-1 degradation in cancer-associated fibroblasts [94]. It is unknown whether stromal cells directly induce cell-mediated autophagy (C-MA) in pancreatic cancer, although lymphocyte-induced C-MA promotes several cancer cell, including pancreatic cancer cell, survival [99].
Autophagy and molecular biology of pancreatic cancer
Since initial recognition of the importance of K-Ras mutations in pancreatic cancer during the late 1980s, understanding of the biologic mechanisms underlying the behavior of this cancer has been growing at an increasingly rapid pace [100]. K-Ras is a member of a highly homologous group of approximately 21 kDa monomeric, membrane-localized GTPases. K-Ras as well as H- and N-Ras is important for cell growth, differentiation, and survival by through a complex interplay of extracellular signals and intracellular signaling cascades. Of them, the Raf/MEK/ERK pathway is the best characterized Ras regulated signaling pathway. Somatic K-Ras mutations, as a transforming protein in human tumors, are found at high rates in pancreatic cancer, lung cancer, and colon cancer. Of note, mutations of the K-Ras gene occur in over 90% of pancreatic carcinomas. Recent studies suggest that autophagy and mitophagy are required for the optimal growth of tumor cells transformed with oncogenic H-RasV12 or K-RasV12 [101-104]. Expression of constitutively active Ras mutants induced autophagy in an ATG5 and Beclin 1 dependent manner [103,105]. Autophagy-defective Ras-transformed cells are not able to proliferate in nutrientdeficient conditions and do not form tumors efficiently in vivo, suggesting a critical role for autophagy in Ras-mediated tumor maintenance [103,104]. However, one study demonstrated that Ras-induced autophagy may cause cell death in breast tumor cells on the verge of oncogenic transformation [105], suggesting a complex relationship between Ras-mediated autophagy and cell death depending on the context [106]. Most solid tumor cells are characterized by energy dependence on aerobic glycolysis accompanied by impairment of the mitochondrial respiratory capacity; these characteristics are consistent with the proposed Warburg effect [107,108]. Ras-transformed cells require autophagy and mitophagy to maintain mitochondrial function [103]. Biochemical analysis demonstrates that autophagy defects result in decreased production of tricarboxylic acid cycle intermediates and impaired oxygen consumption [103] and glycolysis [104].
p53, the most commonly mutated gene in human cancers including pancreatic cancer, plays multiple roles in apoptosis, autophagy, metabolism, cell cycle progression, and many other processes [109]. p53 also regulates autophagy through both transcription-dependent and independent mechanisms. Cytosolic p53 inhibits autophagy through a transcription-independent mechanism [110]. Damage-regulated autophagy modulator (DRAM) and unc-51-like kinase 1 (ULK1) have recently been identified as p53 transcriptionally-regulated genes through which p53 promotes autophagy in contrast to its cytosolic role [111,112]. Interestingly, the endogenous wildtype p53 posttran-scriptionally down-regulates LC3 in colon cancer cells under prolonged starvation [113]. Our findings suggest that HMGB1 and p53 form complexes that regulate the cytoplasmic localization of the reciprocal binding protein and subsequent levels of autophagy and apoptosis [114]. Another important area is delineating whether and how autophagy generally impacts oncogene mutations in pancreatic cancer.
Autophagy and pancreatic cancer treatment
There are three primary treatments for patients with exocrine pancreatic cancer: surgery, radiation therapy, and chemotherapy. Autophagy can be activated by chemotherapy and ionizing radiation in the treatment of pancreatic cancer cells [14-17,115,116]. The cytoprotective role of autophagy following chemotherapy and radiation therapy has been confirmed by many investigators [117]. Gemcitabine, as first-line therapy for patients with advanced pancreas cancer, triggers autophagy in pancreatic cancer cells. Autophagy suppression promotes apoptotic cell death in response to gemcitabine and other stimuli in pancreatic adenocarcinoma [20,117]. Other studies suggested that gemcitabine-induced VMP1-mediated autophagy leads apoptotic cell death [18]. However, zymophagy, a novel selective autophagy pathway which is activated by experimental pancreatitis and mediated by the VMP1-USP9x-p62 pathway, prevents pancreatic cell death [118]. In addition, an enhanced cytotoxic effect of gemcitabine, when combined with cannabinoids, induces reactive oxygen species (ROS)-mediated autophagic cell death in pancreatic tumor cells [119]. Indeed, cannabinoids are also an apoptotic inducer by activation of the endoplasmic reticulum stress pathway in pancreatic cancer cells [120]. ROS act as signaling molecules in various intracellular processes, including apoptosis and autophagy [121]. Mitochondria are a major source of ROS. Radiation therapy and some forms of chemotherapy rely on ROS toxicity to eradicate tumor cells. Inhibition of autophagy or mitophagy increase ROS production, resulting in apoptotic cell death. The concept of autophagic cell death was first established based on observations of increased autophagic markers in dying cells. An increasing number of studies indicate that autophagic cell death is cell death with autophagy rather than cell death by autophagy [59,122].
The efficacy of autophagy inhibitors in preclinical models mostly came from using chloroquine and its derivative hydrochloroquine. Chloroquine is a synthetic 4-aminoquinoline that has been used for 60 years in humans to treat and prevent rheumatoid arthritis, HIV, and malaria in humans. Chloroquine inhibits acidification of the lysosome and therefore prevents autophagy by blocking autophagosome fusion and degradation. In ongoing cancer treatment clinical trials, chloroquine is often used in combination with chemotherapeutic drugs, radiation, and immunotherapy [123]. Of note, a recent study indicated that chloroquine sensitizes breast cancer cells to chemotherapy independent of autophagy [61]. Chloroquine has been demonstrated to inhibit pancreatic tumor growth in a mice model [60,124]. Currently, our ongoing trials examine whether chloroquine or hydro-chloroquine combined with gemcitabine enhances the efficacy of tumor cell killing in pancreatic cancer patients.
HMGB1-rage pathway regulates autophagy in pancreatic cancer
HMGB1 belongs to a family of highly conserved proteins that contain HMG box domains. In the nucleus, HMGB1 is important in the regulation of DNA replication, repair, and recombination, as well as gene transcription. During stress, HMGB1 is released to the extracellular environment and functions as a DAMP [125]. The major receptor for HMGB1 is RAGE and TLR. Overexpression of HMGB1 and RAGE have been demonstrated in many tumors, including pancreatic cancer [126,127]. RAGE is a member of the immunoglobulin superfamily and plays important roles in regulation of ROS generation and inflammatory response [128,129]. In pancreatic cancer, RAGE is compatible with PanIn formation [8]. Gemcitabine promotes RAGE expression by ROS-mediated NF-κB activation [20]. This RAGE upregulation protects pancreatic tumor cells against oxidative injury and increases drug resistance by increasing autophagy and decreasing apoptosis (Figure 4) [117]. Knockdown of RAGE by short hairpin RNA inhibits anticancer agents (e.g., gemcitabine, oxaliplatin, and melphalan) induced autophagy by regulates Beclin 1-Vps34 interaction [117]. ROS also trigger the translocation and release of HMGB1 in pancreatic cancer cells [21]. However, reduced HMGB1 promotes autophagy whereas oxidized HMGB1 induces apoptosis [9]. RAGE is required for extracellular HMGB1-mediated autophagy [9]. As a cellular defense mechanism, cytosolic HMGB1 directly interacts with Beclin 1, which is controlled by ULK1 complex (Figure 4) [130,131]. Antioxidant enzymes (e.g., superoxide dismutases) and small-molecule antioxidants (e.g., N-acetyl-L-cysteine) inhibit HMGB1 activation as well as autophagy induction in pancreatic cancer cells [21]. In the nucleus, HMGB1 regulates expression of heat shock protein beta-1 (HSPB1) (Figure 4) [132]. HSPB1 inhibits apoptosis and its phosphorylation promotes autophagy through regulation of the actin cytoskeletal response, which is required to promote the processes of autophagy and mitophagy [132].
Figure 4.

Pathways of autophagy induction by HMGB1 and RAGE. HMGB1 is an autophagy sensor in oxidative stress. HMGB1 plays important intranuclear, cytosolic, and extracellular roles in the regulation of autophagy. Nuclear HMGB1 regulates HSPB1 expression, which is required for the dynamic of mitophagy to control mitochondrial quality. Cytosolic HMGB1 is a Beclin1 binding protein. HMGB1-Beclin1 complex is a downstream signal from ULK1 complex. Interaction between HMGB1 and p53 regulates the level of HMGB1-Beclin1 complex. Loss of p53 promotes HMGB1 cytosolic translocation and increased HMGB1-Beclin1 complex and autophagy. In contrast, loss of HMGB1 promotes p53 cytosolic translocation and inhibits autophagy. Exogenous HMGB1 binds RAGE to active autophagy. ROS such as H2O2 increase activity of NF-κB and subsequently result in RAGE overexpression. This RAGE upregulation protects pancreatic tumor cells against oxidative injury and increases drug resistance by increasing Beclin1 dependent autophagy and decreasing apoptosis. In addition, RAGE is required for IL-6-induced phosporation of STAT3 (pSTAT3) and subsequently, autophagy induction.
Conclusions
Autophagy is a highly conserved mechanism to degrade intracellular components and promote programmed survival of cells in metabolic distress by providing a nutrient source such as ATP and amino acid. Autophagy is associated with pancreatic cancer, a highly fatal cancer, and plays multiple roles in pancreatic cancer pathogenesis and treatment, which may depend on the tumor type, stage, risk factors, and anticancer agents being used in combination with autophagy inhibitor. The K-Ras oncogene and p53 tumor suppressor gene have been demonstrated to regulate autophagy at different levels. In addition, our studies suggest that the HMGB1-RAGE pathway plays a central role in the regulation of autophagy in pancreatic cancer cells. The expression of RAGE is permissive for early pancreatic neoplasia. It is worth highlighting that hydrochloroquine, a potential autophagy inhibitor, is already approved by the U.S. Food and Drug Administration and may prove to be an effective modulator of autophagy in patients with pancreatic cancer.
Acknowledgements
We apologize to the researchers who were not referenced due to space limitations. D.T. is funded by the Department of Surgery, University of Pittsburgh.
References
- 1.Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011: The impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA Cancer J Clin. 2011;61:212–236. doi: 10.3322/caac.20121. [DOI] [PubMed] [Google Scholar]
- 2.Wang Z, Li Y, Ahmad A, Banerjee S, Azmi AS, Kong D, Sarkar FH. Pancreatic cancer: understanding and overcoming chemoresistance. Nat Rev Gastroenterol Hepatol. 2011;8:27–33. doi: 10.1038/nrgastro.2010.188. [DOI] [PubMed] [Google Scholar]
- 3.Hidalgo M. Pancreatic cancer. N Engl J Med. 2010;362:1605–1617. doi: 10.1056/NEJMra0901557. [DOI] [PubMed] [Google Scholar]
- 4.Vincent A, Herman J, Schulick R, Hruban RH, Goggins M. Pancreatic cancer. Lancet. 2011;378:607–620. doi: 10.1016/S0140-6736(10)62307-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hruban RH, Adsay NV. Molecular classification of neoplasms of the pancreas. Hum Pathol. 2009;40:612–623. doi: 10.1016/j.humpath.2009.01.008. [DOI] [PubMed] [Google Scholar]
- 6.Raimondi S, Maisonneuve P, Lowenfels AB. Epidemiology of pancreatic cancer: an overview. Nat Rev Gastroenterol Hepatol. 2009;6:699–708. doi: 10.1038/nrgastro.2009.177. [DOI] [PubMed] [Google Scholar]
- 7.Morris JPt, Wang SC, Hebrok M. KRAS, Hedgehog, Wnt and the twisted developmental biology of pancreatic ductal adenocarcinoma. Nat Rev Cancer. 2010;10:683–695. doi: 10.1038/nrc2899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.DiNorcia J, Lee MK, Moroziewicz DN, Winner M, Suman P, Bao F, Remotti HE, Zou YS, Yan SF, Qiu W, Su GH, Schmidt AM, Allendorf JD. RAGE gene deletion inhibits the development and progression of ductal neoplasia and prolongs survival in a murine model of pancreatic cancer. J Gastrointest Surg. 2012;16:104–112. doi: 10.1007/s11605-011-1754-9. discussion 112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tang D, Kang R, Cheh CW, Livesey KM, Liang X, Schapiro NE, Benschop R, Sparvero LJ, Amoscato AA, Tracey KJ, Zeh HJ, Lotze MT. HMGB1 release and redox regulates autophagy and apoptosis in cancer cells. Oncogene. 2010;29:5299–5310. doi: 10.1038/onc.2010.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Guerra C, Collado M, Navas C, Schuhmacher AJ, Hernandez-Porras I, Canamero M, Rodriguez-Justo M, Serrano M, Barbacid M. Pancreatitis-induced inflammation contributes to pancreatic cancer by inhibiting oncogene-induced senescence. Cancer Cell. 2011;19:728–739. doi: 10.1016/j.ccr.2011.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Neesse A, Michl P, Frese KK, Feig C, Cook N, Jacobetz MA, Lolkema MP, Buchholz M, Olive KP, Gress TM, Tuveson DA. Stromal biology and therapy in pancreatic cancer. Gut. 2011;60:861–868. doi: 10.1136/gut.2010.226092. [DOI] [PubMed] [Google Scholar]
- 12.Yang S, Wang X, Contino G, Liesa M, Sahin E, Ying H, Bause A, Li Y, Stommel JM, Dell'antonio G, Mautner J, Tonon G, Haigis M, Shirihai OS, Doglioni C, Bardeesy N, Kimmelman AC. Pancreatic cancers require autophagy for tumor growth. Genes Dev. 2011;25:717–729. doi: 10.1101/gad.2016111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grasso D, Garcia MN, Iovanna JL. Autophagy in pancreatic cancer. Int J Cell Biol. 2012;2012:760498. doi: 10.1155/2012/760498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Naumann P, Fortunato F, Zentgraf H, Buchler MW, Herr I, Werner J. Autophagy and cell death signaling following dietary sulforaphane act independently of each other and require oxidative stress in pancreatic cancer. Int J Oncol. 2011;39:101–109. doi: 10.3892/ijo.2011.1025. [DOI] [PubMed] [Google Scholar]
- 15.Donadelli M, Dando I, Zaniboni T, Costanzo C, Dalla Pozza E, Scupoli MT, Scarpa A, Zappavigna S, Marra M, Abbruzzese A, Bifulco M, Caraglia M, Palmieri M. Gemcitabine/cannabinoid combination triggers autophagy in pancreatic cancer cells through a ROSmediated mechanism. Cell death & disease. 2011;2:e152. doi: 10.1038/cddis.2011.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mukubou H, Tsujimura T, Sasaki R, Ku Y. The role of autophagy in the treatment of pancreatic cancer with gemcitabine and ionizing radiation. Int J Oncol. 2010;37:821–828. doi: 10.3892/ijo_00000732. [DOI] [PubMed] [Google Scholar]
- 17.Cullen JJ. Ascorbate induces autophagy in pancreatic cancer. Autophagy. 2010;6:421–422. doi: 10.4161/auto.6.3.11527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pardo R, Lo Re A, Archange C, Ropolo A, Papademetrio DL, Gonzalez CD, Alvarez EM, Iovanna JL, Vaccaro MI. Gemcitabine induces the VMP1-mediated autophagy pathway to promote apoptotic death in human pancreatic cancer cells. Pancreatology. 2010;10:19–26. doi: 10.1159/000264680. [DOI] [PubMed] [Google Scholar]
- 19.Fujii S, Mitsunaga S, Yamazaki M, Hasebe T, Ishii G, Kojima M, Kinoshita T, Ueno T, Esumi H, Ochiai A. Autophagy is activated in pancreatic cancer cells and correlates with poor patient outcome. Cancer Sci. 2008;99:1813–1819. doi: 10.1111/j.1349-7006.2008.00893.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kang R, Tang D, Livesey KM, Schapiro NE, Lotze MT, Zeh HJ 3rd. The Receptor for Advanced Glycation End-products (RAGE) protects pancreatic tumor cells against oxidative injury. Antioxid Redox Signal. 2011;15:2175–2184. doi: 10.1089/ars.2010.3378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tang D, Kang R, Livesey KM, Zeh HJ 3rd, Lotze MT. High mobility group box 1 (HMGB1) activates an autophagic response to oxidative stress. Antioxid Redox Signal. 2011;15:2185–2195. doi: 10.1089/ars.2010.3666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kang R, Tang D, Lotze MT, Zeh HJ 3rd. RAGE regulates autophagy and apoptosis following oxidative injury. Autophagy. 2011;7:442–444. doi: 10.4161/auto.7.4.14681. [DOI] [PubMed] [Google Scholar]
- 23.Kang R, Tang D, Loze MT, Zeh HJ. Apoptosis to autophagy switch triggered by the MHC class III-encoded receptor for advanced glycation endproducts (RAGE) Autophagy. 2011;7:91–93. doi: 10.1038/cdd.2009.149. [DOI] [PubMed] [Google Scholar]
- 24.Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation. Science. 2000;290:1717–1721. doi: 10.1126/science.290.5497.1717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Klionsky DJ. Autophagy: from phenomenology to molecular understanding in less than a decade. Nat Rev Mol Cell Biol. 2007;8:931–937. doi: 10.1038/nrm2245. [DOI] [PubMed] [Google Scholar]
- 26.Tooze SA, Yoshimori T. The origin of the autophagosomal membrane. Nat Cell Biol. 2010;12:831–835. doi: 10.1038/ncb0910-831. [DOI] [PubMed] [Google Scholar]
- 27.Youle RJ, Narendra DP. Mechanisms of mitophagy. Nat Rev Mol Cell Biol. 2011;12:9–14. doi: 10.1038/nrm3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Knodler LA, Celli J. Eating the strangers within: host control of intracellular bacteria via xenophagy. Cell Microbiol. 2011;13:1319–1327. doi: 10.1111/j.1462-5822.2011.01632.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Levine B. Eating oneself and uninvited guests: autophagy-related pathways in cellular defense. Cell. 2005;120:159–162. doi: 10.1016/j.cell.2005.01.005. [DOI] [PubMed] [Google Scholar]
- 30.He C, Klionsky DJ. Regulation mechanisms and signaling pathways of autophagy. Annu Rev Genet. 2009;43:67–93. doi: 10.1146/annurev-genet-102808-114910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen Y, Klionsky DJ. The regulation of autophagy - unanswered questions. J Cell Sci. 2011;124:161–170. doi: 10.1242/jcs.064576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jung CH, Ro SH, Cao J, Otto NM, Kim DH. mTOR regulation of autophagy. FEBS Lett. 2010;584:1287–1295. doi: 10.1016/j.febslet.2010.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 2011;18:571–580. doi: 10.1038/cdd.2010.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mizushima N, Yoshimori T, Levine B. Methods in mammalian autophagy research. Cell. 2010;140:313–326. doi: 10.1016/j.cell.2010.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mizushima N, Levine B. Autophagy in mammalian development and differentiation. Nat Cell Biol. 2010;12:823–830. doi: 10.1038/ncb0910-823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kroemer G, Marino G, Levine B. Autophagy and the integrated stress response. Mol Cell. 2010;40:280–293. doi: 10.1016/j.molcel.2010.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008;132:27–42. doi: 10.1016/j.cell.2007.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Virgin HW, Levine B. Autophagy genes in immunity. Nat Immunol. 2009;10:461–470. doi: 10.1038/ni.1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Levine B, Mizushima N, Virgin HW. Autophagy in immunity and inflammation. Nature. 2011;469:323–335. doi: 10.1038/nature09782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mizushima N, Levine B, Cuervo AM, Klionsky DJ. Autophagy fights disease through cellular self-digestion. Nature. 2008;451:1069–1075. doi: 10.1038/nature06639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword. Science. 2004;306:990–995. doi: 10.1126/science.1099993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Janku F, McConkey DJ, Hong DS, Kurzrock R. Autophagy as a target for anticancer therapy. Nat Rev Clin Oncol. 2011;8:528–539. doi: 10.1038/nrclinonc.2011.71. [DOI] [PubMed] [Google Scholar]
- 43.Hippert MM, O'Toole PS, Thorburn A. Autophagy in cancer: good, bad, or both? Cancer Res. 2006;66:9349–9351. doi: 10.1158/0008-5472.CAN-06-1597. [DOI] [PubMed] [Google Scholar]
- 44.Kimmelman AC. The dynamic nature of autophagy in cancer. Genes Dev. 2011;25:1999–2010. doi: 10.1101/gad.17558811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kondo Y, Kanzawa T, Sawaya R, Kondo S. The role of autophagy in cancer development and response to therapy. Nat Rev Cancer. 2005;5:726–734. doi: 10.1038/nrc1692. [DOI] [PubMed] [Google Scholar]
- 46.Mathew R, Karantza-Wadsworth V, White E. Role of autophagy in cancer. Nat Rev Cancer. 2007;7:961–967. doi: 10.1038/nrc2254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, Levine B. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature. 1999;402:672–676. doi: 10.1038/45257. [DOI] [PubMed] [Google Scholar]
- 48.Liang C, Feng P, Ku B, Dotan I, Canaani D, Oh BH, Jung JU. Autophagic and tumour suppressor activity of a novel Beclin1-binding protein UVRAG. Nat Cell Biol. 2006;8:688–699. doi: 10.1038/ncb1426. [DOI] [PubMed] [Google Scholar]
- 49.Takamura A, Komatsu M, Hara T, Sakamoto A, Kishi C, Waguri S, Eishi Y, Hino O, Tanaka K, Mizushima N. Autophagy-deficient mice develop multiple liver tumors. Genes Dev. 2011;25:795–800. doi: 10.1101/gad.2016211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Takahashi Y, Coppola D, Matsushita N, Cualing HD, Sun M, Sato Y, Liang C, Jung JU, Cheng JQ, Mule JJ, Pledger WJ, Wang HG. Bif-1 interacts with Beclin 1 through UVRAG and regulates autophagy and tumorigenesis. Nat Cell Biol. 2007;9:1142–1151. doi: 10.1038/ncb1634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Degenhardt K, Mathew R, Beaudoin B, Bray K, Anderson D, Chen G, Mukherjee C, Shi Y, Gelinas C, Fan Y, Nelson DA, Jin S, White E. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell. 2006;10:51–64. doi: 10.1016/j.ccr.2006.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rabinowitz JD, White E. Autophagy and metabolism. Science. 2010;330:1344–1348. doi: 10.1126/science.1193497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pavlides S, Tsirigos A, Migneco G, Whitaker-Menezes D, Chiavarina B, Flomenberg N, Frank PG, Casimiro MC, Wang C, Pestell RG, Martinez-Outschoorn UE, Howell A, Sotgia F, Lisanti MP. The autophagic tumor stroma model of cancer: Role of oxidative stress and ketone production in fueling tumor cell metabolism. Cell Cycle. 2010;9:3485–3505. doi: 10.4161/cc.9.17.12721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Altman BJ, Jacobs SR, Mason EF, Michalek RD, Macintyre AN, Coloff JL, Ilkayeva O, Jia W, He YW, Rathmell JC. Autophagy is essential to suppress cell stress and to allow BCR-Abl-mediated leukemogenesis. Oncogene. 2011;30:1855–1867. doi: 10.1038/onc.2010.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Maiuri MC, Zalckvar E, Kimchi A, Kroemer G. Self-eating and self-killing: crosstalk between autophagy and apoptosis. Nat Rev Mol Cell Biol. 2007;8:741–752. doi: 10.1038/nrm2239. [DOI] [PubMed] [Google Scholar]
- 56.Tang D, Lotze MT, Kang R, Zeh HJ. Apoptosis promotes early tumorigenesis. Oncogene. 2011;30:1851–1854. doi: 10.1038/onc.2010.573. [DOI] [PubMed] [Google Scholar]
- 57.Amaravadi RK, Lippincott-Schwartz J, Yin XM, Weiss WA, Takebe N, Timmer W, DiPaola RS, Lotze MT, White E. Principles and current strategies for targeting autophagy for cancer treatment. Clin Cancer Res. 2011;17:654–666. doi: 10.1158/1078-0432.CCR-10-2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Shimizu S, Kanaseki T, Mizushima N, Mizuta T, Arakawa-Kobayashi S, Thompson CB, Tsujimoto Y. Role of Bcl-2 family proteins in a non-apoptotic programmed cell death dependent on autophagy genes. Nat Cell Biol. 2004;6:1221–1228. doi: 10.1038/ncb1192. [DOI] [PubMed] [Google Scholar]
- 59.Levine B, Yuan J. Autophagy in cell death: an innocent convict? J Clin Invest. 2005;115:2679–2688. doi: 10.1172/JCI26390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yang S, Kimmelman AC. A critical role for autophagy in pancreatic cancer. Autophagy. 2011;7:912–913. doi: 10.4161/auto.7.8.15762. [DOI] [PubMed] [Google Scholar]
- 61.Maycotte P, Aryal S, Cummings CT, Thorburn J, Morgan MJ, Thorburn A. Chloroquine sensitizes breast cancer cells to chemotherapy independent of autophagy. Autophagy. 2012;8:200–212. doi: 10.4161/auto.8.2.18554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kang R, Loux T, Tang D, Schapiro NE, Vernon P, Livesey KM, Krasinskas A, Lotze MT, Zeh HJ 3rd. The expression of the receptor for advanced glycation endproducts (RAGE) is permissive for early pancreatic neoplasia. Proc Natl Acad Sci USA. 2012;109:7031–7036. doi: 10.1073/pnas.1113865109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang F, Herrington M, Larsson J, Permert J. The relationship between diabetes and pancreatic cancer. Mol Cancer. 2003;2:4. doi: 10.1186/1476-4598-2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Gidekel Friedlander SY, Chu GC, Snyder EL, Girnius N, Dibelius G, Crowley D, Vasile E, De-Pinho RA, Jacks T. Context-dependent transformation of adult pancreatic cells by oncogenic K-Ras. Cancer Cell. 2009;16:379–389. doi: 10.1016/j.ccr.2009.09.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lee KE, Bar-Sagi D. Oncogenic KRas suppresses inflammation-associated senescence of pancreatic ductal cells. Cancer Cell. 2010;18:448–458. doi: 10.1016/j.ccr.2010.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jung HS, Chung KW, Won Kim J, Kim J, Komatsu M, Tanaka K, Nguyen YH, Kang TM, Yoon KH, Kim JW, Jeong YT, Han MS, Lee MK, Kim KW, Shin J, Lee MS. Loss of autophagy diminishes pancreatic beta cell mass and function with resultant hyperglycemia. Cell Metab. 2008;8:318–324. doi: 10.1016/j.cmet.2008.08.013. [DOI] [PubMed] [Google Scholar]
- 67.Ebato C, Uchida T, Arakawa M, Komatsu M, Ueno T, Komiya K, Azuma K, Hirose T, Tanaka K, Kominami E, Kawamori R, Fujitani Y, Watada H. Autophagy is important in islet homeostasis and compensatory increase of beta cell mass in response to high-fat diet. Cell Metab. 2008;8:325–332. doi: 10.1016/j.cmet.2008.08.009. [DOI] [PubMed] [Google Scholar]
- 68.Grasso D, Sacchetti ML, Bruno L, Lo Re A, Iovanna JL, Gonzalez CD, Vaccaro MI. Autophagy and VMP1 expression are early cellular events in experimental diabetes. Pancreatology. 2009;9:81–88. doi: 10.1159/000178878. [DOI] [PubMed] [Google Scholar]
- 69.Ekbom A, McLaughlin JK, Nyren O. Pancreatitis and the risk of pancreatic cancer. N Engl J Med. 1993;329:1502–1503. doi: 10.1056/NEJM199311113292016. [DOI] [PubMed] [Google Scholar]
- 70.Lowenfels AB, Maisonneuve P, Cavallini G, Ammann RW, Lankisch PG, Andersen JR, Dimagno EP, Andren-Sandberg A, Domellof L. Pancreatitis and the risk of pancreatic cancer. International Pancreatitis Study Group. N Engl J Med. 1993;328:1433–1437. doi: 10.1056/NEJM199305203282001. [DOI] [PubMed] [Google Scholar]
- 71.Su KH, Cuthbertson C, Christophi C. Review of experimental animal models of acute pancreatitis. HPB (Oxford) 2006;8:264–286. doi: 10.1080/13651820500467358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Gukovsky I, Pandol SJ, Mareninova OA, Shalbueva N, Jia W, Gukovskaya AS. Impaired autophagy and organellar dysfunction in pancreatitis. J Gastroenterol Hepatol. 2012;27(Suppl 2):27–32. doi: 10.1111/j.1440-1746.2011.07004.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yang S, Bing M, Chen F, Sun Y, Chen H, Chen W. Autophagy Regulation by the Nuclear Factor kappaB Signal Axis in Acute Pancreatitis. Pancreas. 2012;41:367–373. doi: 10.1097/MPA.0b013e31822a9b05. [DOI] [PubMed] [Google Scholar]
- 74.Lugea A, Waldron RT, French SW, Pandol SJ. Drinking and driving pancreatitis: links between endoplasmic reticulum stress and autophagy. Autophagy. 2011;7:783–785. doi: 10.4161/auto.7.7.15594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gukovsky I, Gukovskaya AS. Impaired autophagy underlies key pathological responses of acute pancreatitis. Autophagy. 2010;6:428–429. doi: 10.4161/auto.6.3.11530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fortunato F, Burgers H, Bergmann F, Rieger P, Buchler MW, Kroemer G, Werner J. Impaired autolysosome formation correlates with Lamp-2 depletion: role of apoptosis, autophagy, and necrosis in pancreatitis. Gastroenterology. 2009;137:350–360. 360 e351-355. doi: 10.1053/j.gastro.2009.04.003. [DOI] [PubMed] [Google Scholar]
- 77.Ohmuraya M, Yamamura K. Autophagy and acute pancreatitis: a novel autophagy theory for trypsinogen activation. Autophagy. 2008;4:1060–1062. doi: 10.4161/auto.6825. [DOI] [PubMed] [Google Scholar]
- 78.Mareninova OA, Hermann K, French SW, O'Konski MS, Pandol SJ, Webster P, Erickson AH, Katunuma N, Gorelick FS, Gukovsky I, Gukovskaya AS. Impaired autophagic flux mediates acinar cell vacuole formation and trypsinogen activation in rodent models of acute pancreatitis. J Clin Invest. 2009;119:3340–3355. doi: 10.1172/JCI38674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Eskelinen EL, Illert AL, Tanaka Y, Schwarzmann G, Blanz J, Von Figura K, Saftig P. Role of LAMP-2 in lysosome biogenesis and autophagy. Mol Biol Cell. 2002;13:3355–3368. doi: 10.1091/mbc.E02-02-0114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kunzli BM, Berberat PO, Zhu ZW, Martignoni M, Kleeff J, Tempia-Caliera AA, Fukuda M, Zimmermann A, Friess H, Buchler MW. Influences of the lysosomal associated membrane proteins (Lamp-1, Lamp-2) and Mac-2 binding protein (Mac-2-BP) on the prognosis of pancreatic carcinoma. Cancer. 2002;94:228–239. doi: 10.1002/cncr.10162. [DOI] [PubMed] [Google Scholar]
- 81.Kon M, Kiffin R, Koga H, Chapochnick J, Macian F, Varticovski L, Cuervo AM. Chaperonemediated autophagy is required for tumor growth. Sci Transl Med. 2011;3:109ra117. doi: 10.1126/scitranslmed.3003182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ding WX, Li M, Chen X, Ni HM, Lin CW, Gao W, Lu B, Stolz DB, Clemens DL, Yin XM. Autophagy reduces acute ethanol-induced hepatotoxicity and steatosis in mice. Gastroenterology. 2010;139:1740–1752. doi: 10.1053/j.gastro.2010.07.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Chen ZH, Kim HP, Sciurba FC, Lee SJ, Feghali-Bostwick C, Stolz DB, Dhir R, Landreneau RJ, Schuchert MJ, Yousem SA, Nakahira K, Pilewski JM, Lee JS, Zhang Y, Ryter SW, Choi AM. Egr-1 regulates autophagy in cigarette smoke-induced chronic obstructive pulmonary disease. PLoS ONE. 2008;3:e3316. doi: 10.1371/journal.pone.0003316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Charbonneau A, Unson CG, Lavoie JM. High-fat diet-induced hepatic steatosis reduces glucagon receptor content in rat hepatocytes: potential interaction with acute exercise. J Physiol. 2007;579:255–267. doi: 10.1113/jphysiol.2006.121954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Duffy JP, Eibl G, Reber HA, Hines OJ. Influence of hypoxia and neoangiogenesis on the growth of pancreatic cancer. Mol Cancer. 2003;2:12. doi: 10.1186/1476-4598-2-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Niizeki H, Kobayashi M, Horiuchi I, Akakura N, Chen J, Wang J, Hamada JI, Seth P, Katoh H, Watanabe H, Raz A, Hosokawa M. Hypoxia enhances the expression of autocrine motility factor and the motility of human pancreatic cancer cells. Br J Cancer. 2002;86:1914–1919. doi: 10.1038/sj.bjc.6600331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Farrow B, Albo D, Berger DH. The role of the tumor microenvironment in the progression of pancreatic cancer. J Surg Res. 2008;149:319–328. doi: 10.1016/j.jss.2007.12.757. [DOI] [PubMed] [Google Scholar]
- 88.Bellot G, Garcia-Medina R, Gounon P, Chiche J, Roux D, Pouyssegur J, Mazure NM. Hypoxiainduced autophagy is mediated through hypoxia-inducible factor induction of BNIP3 and BNIP3L via their BH3 domains. Mol Cell Biol. 2009;29:2570–2581. doi: 10.1128/MCB.00166-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Zhang H, Bosch-Marce M, Shimoda LA, Tan YS, Baek JH, Wesley JB, Gonzalez FJ, Semenza GL. Mitochondrial autophagy is an HIF-1-dependent adaptive metabolic response to hypoxia. J Biol Chem. 2008;283:10892–10903. doi: 10.1074/jbc.M800102200. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 90.Tracy K, Dibling BC, Spike BT, Knabb JR, Schumacker P, Macleod KF. BNIP3 is an RB/E2F target gene required for hypoxiainduced autophagy. Mol Cell Biol. 2007;27:6229–6242. doi: 10.1128/MCB.02246-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Papandreou I, Lim AL, Laderoute K, Denko NC. Hypoxia signals autophagy in tumor cells via AMPK activity, independent of HIF-1, BNIP3, and BNIP3L. Cell Death Differ. 2008;15:1572–1581. doi: 10.1038/cdd.2008.84. [DOI] [PubMed] [Google Scholar]
- 92.Rausch V, Liu L, Apel A, Rettig T, Gladkich J, Labsch S, Kallifatidis G, Kaczorowski A, Groth A, Gross W, Gebhard MM, Schemmer P, Werner J, Salnikov AV, Zentgraf H, Buchler MW, Herr I. Autophagy mediates survival of pancreatic tumour-initiating cells in a hypoxic microenvironment. J Pathol. 2012 doi: 10.1002/path.3994. [DOI] [PubMed] [Google Scholar]
- 93.Hwang RF, Moore T, Arumugam T, Ramachandran V, Amos KD, Rivera A, Ji B, Evans DB, Logsdon CD. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res. 2008;68:918–926. doi: 10.1158/0008-5472.CAN-07-5714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Martinez-Outschoorn UE, Trimmer C, Lin Z, Whitaker-Menezes D, Chiavarina B, Zhou J, Wang C, Pavlides S, Martinez-Cantarin MP, Capozza F, Witkiewicz AK, Flomenberg N, Howell A, Pestell RG, Caro J, Lisanti MP, Sotgia F. Autophagy in cancer associated fibroblasts promotes tumor cell survival: Role of hypoxia, HIF1 induction and NFkappaB activation in the tumor stromal microenvironment. Cell Cycle. 2010;9:3515–3533. doi: 10.4161/cc.9.17.12928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Pavlides S, Vera I, Gandara R, Sneddon S, Pestell RG, Mercier I, Martinez-Outschoorn UE, Whitaker-Menezes D, Howell A, Sotgia F, Lisanti MP. Warburg Meets Autophagy: Cancer-Associated Fibroblasts Accelerate Tumor Growth and Metastasis via Oxidative Stress, Mitophagy, and Aerobic Glycolysis. Antioxid Redox Signal. 2012;16:1264–1284. doi: 10.1089/ars.2011.4243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Sotgia F, Martinez-Outschoorn UE, Howell A, Pestell RG, Pavlides S, Lisanti MP. Caveolin-1 and cancer metabolism in the tumor microenvironment: markers, models, and mechanisms. Annu Rev Pathol. 2012;7:423–467. doi: 10.1146/annurev-pathol-011811-120856. [DOI] [PubMed] [Google Scholar]
- 97.Martinez-Outschoorn UE, Whitaker-Menezes D, Lin Z, Flomenberg N, Howell A, Pestell RG, Lisanti MP, Sotgia F. Cytokine production and inflammation drive autophagy in the tumor microenvironment: role of stromal caveolin-1 as a key regulator. Cell Cycle. 2011;10:1784–1793. doi: 10.4161/cc.10.11.15674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tanase CP, Dima S, Mihai M, Raducan E, Nicolescu MI, Albulescu L, Voiculescu B, Dumitrascu T, Cruceru LM, Leabu M, Popescu I, Hinescu ME. Caveolin-1 overexpression correlates with tumour progression markers in pancreatic ductal adenocarcinoma. J Mol Histol. 2009;40:23–29. doi: 10.1007/s10735-008-9209-7. [DOI] [PubMed] [Google Scholar]
- 99.Buchser WJ, Laskow TC, Pavlik PJ, Lin HM, Lotze MT. Cell-mediated Autophagy Promotes Cancer Cell Survival. Cancer Res. 2012 doi: 10.1158/0008-5472.CAN-11-3396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Abramson MA, Jazag A, van der Zee JA, Whang EE. The molecular biology of pancreatic cancer. Gastrointest Cancer Res. 2007;1:S7–S12. [PMC free article] [PubMed] [Google Scholar]
- 101.Kim MJ, Woo SJ, Yoon CH, Lee JS, An S, Choi YH, Hwang SG, Yoon G, Lee SJ. Involvement of autophagy in oncogenic K-Ras-induced malignant cell transformation. J Biol Chem. 2011;286:12924–12932. doi: 10.1074/jbc.M110.138958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kim JH, Kim HY, Lee YK, Yoon YS, Xu WG, Yoon JK, Choi SE, Ko YG, Kim MJ, Lee SJ, Wang HJ, Yoon G. Involvement of mitophagy in oncogenic K-Ras-induced transformation: Overcoming a cellular energy deficit from glucose deficiency. Autophagy. 2011;7:1187–1198. doi: 10.4161/auto.7.10.16643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Guo JY, Chen HY, Mathew R, Fan J, Strohecker AM, Karsli-Uzunbas G, Kamphorst JJ, Chen G, Lemons JM, Karantza V, Coller HA, Dipaola RS, Gelinas C, Rabinowitz JD, White E. Activated Ras requires autophagy to maintain oxidative metabolism and tumorigenesis. Genes & development. 2011;25:460–470. doi: 10.1101/gad.2016311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Lock R, Roy S, Kenific CM, Su JS, Salas E, Ronen SM, Debnath J. Autophagy facilitates glycolysis during Ras-mediated oncogenic transformation. Mol Biol Cell. 2011;22:165–178. doi: 10.1091/mbc.E10-06-0500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Elgendy M, Sheridan C, Brumatti G, Martin SJ. Oncogenic Ras-induced expression of Noxa and Beclin-1 promotes autophagic cell death and limits clonogenic survival. Mol Cell. 2011;42:23–35. doi: 10.1016/j.molcel.2011.02.009. [DOI] [PubMed] [Google Scholar]
- 106.Marino G, Martins I, Kroemer G. Autophagy in Ras-induced malignant transformation: fatal or vital? Mol Cell. 2011;42:1–3. doi: 10.1016/j.molcel.2011.03.003. [DOI] [PubMed] [Google Scholar]
- 107.Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324:1029–1033. doi: 10.1126/science.1160809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Hsu PP, Sabatini DM. Cancer cell metabolism: Warburg and beyond. Cell. 2008;134:703–707. doi: 10.1016/j.cell.2008.08.021. [DOI] [PubMed] [Google Scholar]
- 109.Green DR, Kroemer G. Cytoplasmic functions of the tumour suppressor p53. Nature. 2009;458:1127–1130. doi: 10.1038/nature07986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Tasdemir E, Maiuri MC, Galluzzi L, Vitale I, Djavaheri-Mergny M, D'amelio M, Criollo A, Morselli E, Zhu C, Harper F, Nannmark U, Samara C, Pinton P, Vicencio J, Carnuccio R, Moll U, Madeo F, Paterlini-Brechot P, Rizzuto R, Szabadkai G, Pierron G, Blomgren K, Tavernarakis N, Codogno P, Cecconi F, Kroemer G. Regulation of autophagy by cytoplasmic p53. Nat Cell Biol. 2008;10:676–687. doi: 10.1038/ncb1730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Crighton D, Wilkinson S, O'Prey J, Syed N, Smith P, Harrison PR, Gasco M, Garrone O, Crook T, Ryan KM. DRAM, a p53-induced modulator of autophagy, is critical for apoptosis. Cell. 2006;126:121–134. doi: 10.1016/j.cell.2006.05.034. [DOI] [PubMed] [Google Scholar]
- 112.Gao W, Shen Z, Shang L, Wang X. Upregulation of human autophagy-initiation kinase ULK1 by tumor suppressor p53 contributes to DNAdamage-induced cell death. Cell Death Differ. 2011;18:1598–1607. doi: 10.1038/cdd.2011.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Scherz-Shouval R, Weidberg H, Gonen C, Wilder S, Elazar Z, Oren M. p53-dependent regulation of autophagy protein LC3 supports cancer cell survival under prolonged starvation. Proc Natl Acad Sci USA. 2010;107:18511–18516. doi: 10.1073/pnas.1006124107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Livesey K, Kang R, Vernon P, Buchser W, Loughran P, Watkins SC, Zhang L, Manfredi JJ, Zeh HJ, Li L, Lotze M, Tang D. p53/HMGB1 Complexes Regulate Autophagy and Apoptosis. Cancer Res. 2012 doi: 10.1158/0008-5472.CAN-11-2291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Udelnow A, Kreyes A, Ellinger S, Landfester K, Walther P, Klapperstueck T, Wohlrab J, Henne-Bruns D, Knippschild U, Wurl P. Omeprazole inhibits proliferation and modulates autophagy in pancreatic cancer cells. PLoS ONE. 2011;6:e20143. doi: 10.1371/journal.pone.0020143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Akar U, Ozpolat B, Mehta K, Fok J, Kondo Y, Lopez-Berestein G. Tissue transglutaminase inhibits autophagy in pancreatic cancer cells. Mol Cancer Res. 2007;5:241–249. doi: 10.1158/1541-7786.MCR-06-0229. [DOI] [PubMed] [Google Scholar]
- 117.Kang R, Tang D, Schapiro NE, Livesey KM, Farkas A, Loughran P, Bierhaus A, Lotze MT, Zeh HJ. The receptor for advanced glycation end products (RAGE) sustains autophagy and limits apoptosis, promoting pancreatic tumor cell survival. Cell Death Differ. 2010;17:666–676. doi: 10.1038/cdd.2009.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Grasso D, Ropolo A, Lo Re A, Boggio V, Molejon MI, Iovanna JL, Gonzalez CD, Urrutia R, Vaccaro MI. Zymophagy, a novel selective autophagy pathway mediated by VMP1-USP9x-p62, prevents pancreatic cell death. J Biol Chem. 2011;286:8308–8324. doi: 10.1074/jbc.M110.197301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Vara D, Salazar M, Olea-Herrero N, Guzman M, Velasco G, Diaz-Laviada I. Anti-tumoral action of cannabinoids on hepatocellular carcinoma: role of AMPK-dependent activation of autophagy. Cell Death Differ. 2011;18:1099–1111. doi: 10.1038/cdd.2011.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Carracedo A, Gironella M, Lorente M, Garcia S, Guzman M, Velasco G, Iovanna JL. Cannabinoids induce apoptosis of pancreatic tumor cells via endoplasmic reticulum stress-related genes. Cancer Res. 2006;66:6748–6755. doi: 10.1158/0008-5472.CAN-06-0169. [DOI] [PubMed] [Google Scholar]
- 121.Tang D, Kang R, Zeh HJ 3rd. High-mobility group box 1, oxidative stress, and disease. Antioxid Redox Signal. 2011;14:1315–1335. doi: 10.1089/ars.2010.3356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Kroemer G, Levine B. Autophagic cell death: the story of a misnomer. Nat Rev Mol Cell Biol. 2008;9:1004–1010. doi: 10.1038/nrm2527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Liang X, de Vera ME, Buchser WJ, Romo de Vivar Chavez A, Loughran P, Beer-Stolz D, Basse P, Wang T, van Houten B, Zeh HJ, Lotze M. Inhibiting Autophagy During Interleukin 2 Immunotherapy Promotes Long Term Tumor Regression. Cancer Res. 2012 doi: 10.1158/0008-5472.CAN-12-0320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Yang S, Wang X, Contino G, Liesa M, Sahin E, Ying H, Bause A, Li Y, Stommel JM, Dell'antonio G, Mautner J, Tonon G, Haigis M, Shirihai OS, Doglioni C, Bardeesy N, Kimmelman AC. Pancreatic cancers require autophagy for tumor growth. Genes & development. 2011;25:717–729. doi: 10.1101/gad.2016111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol. 2005;5:331–342. doi: 10.1038/nri1594. [DOI] [PubMed] [Google Scholar]
- 126.Tang D, Kang R, Zeh HJ 3rd. High-mobility group box 1 and cancer. Biochim Biophys Acta. 2010;1799:131–140. doi: 10.1016/j.bbagrm.2009.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ. HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol. 2010;28:367–388. doi: 10.1146/annurev.immunol.021908.132603. [DOI] [PubMed] [Google Scholar]
- 128.Coughlan MT, Thorburn DR, Penfold SA, Laskowski A, Harcourt BE, Sourris KC, Tan AL, Fukami K, Thallas-Bonke V, Nawroth PP, Brownlee M, Bierhaus A, Cooper ME, Forbes JM. RAGE-induced cytosolic ROS promote mitochondrial superoxide generation in diabetes. J Am Soc Nephrol. 2009;20:742–752. doi: 10.1681/ASN.2008050514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Sparvero LJ, Asafu-Adjei D, Kang R, Tang D, Amin N, Im J, Rutledge R, Lin B, Amoscato AA, Zeh HJ, Lotze MT. RAGE (Receptor for Advanced Glycation Endproducts), RAGE ligands, and their role in cancer and inflammation. J Transl Med. 2009;7:17. doi: 10.1186/1479-5876-7-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Tang D, Kang R, Livesey KM, Cheh CW, Farkas A, Loughran P, Hoppe G, Bianchi ME, Tracey KJ, Zeh HJ 3rd. Endogenous HMGB1 regulates autophagy. J Cell Biol. 2010;190:881–892. doi: 10.1083/jcb.200911078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Huang J, Ni J, Liu K, Yu Y, Xie M, Kang R, Vernon P, Cao L, Tang D. HMGB1 promotes drug resistance in osteosarcoma. Cancer Res. 2012;72:230–238. doi: 10.1158/0008-5472.CAN-11-2001. [DOI] [PubMed] [Google Scholar]
- 132.Tang D, Kang R, Livesey KM, Kroemer G, Billiar TR, Van Houten B, Zeh HJ 3rd. High-mobility group box 1 is essential for mitochondrial quality control. Cell Metab. 2011;13:701–711. doi: 10.1016/j.cmet.2011.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]