

Abstract
Alzheimer's disease (AD) and bipolar disorder (BD) are progressive brain disorders. Upregulated mRNA and protein levels of neuroinflammatory and arachidonic acid (AA) markers with loss of synaptic markers (synaptophysin and drebrin) have been reported in brain tissue from AD and BD patients. We hypothesized that some of these changes are associated with epigenetic modifications of relevant genes. To test this, we measured gene-specific CpG methylation, global DNA methylation and histone modifications in postmortem frontal cortex from BD (n=10) and AD (n=10) patients and respective age-matched controls (10 per group). AD and BD brains showed several epigenetic similarities, including global DNA hypermethylation, and histone H3 phosphorylation. These changes were associated with hypo- and hypermethylation of CpG islands in cyclooxygenase-2 and brain-derived neurotrophic factor promoter regions, respectively. Only the AD brain showed hyper- and hypomethylated CpG islands in promoter regions for cAMP response element-binding protein and nuclear transcription factor kappa B genes, respectively. Only the BD brain demonstrated increased global histone H3 acetylation and hypermethylation of the promotor region for the drebrin-like protein gene. There was no significant epigenetic modification for 12-lipooxygenase or p450 epoxygenase in either illness. Many observed epigenetic changes were inversely related to respective changes in mRNA and protein levels. These epigenetic modifications involving neuroinflammatory, AA cascade and synaptic markers may contribute to progression in AD and BD and identify new targets for drug development.
Keywords: Alzheimer's disease, arachidonic acid cascade, bipolar disorder, DNA methylation, drebrin, synaptophysin
Introduction
Alzheimer's disease (AD) and bipolar disorder (BD) are progressive neuropsychiatric illnesses with overlapping symptoms and neuropathology, including brain atrophy, cognitive impairment, emotional disturbances, neuroinflammation, excitotoxicity and upregulated brain arachidonic acid (AA) metabolism.1, 2, 3 Common behavioral disturbances in AD, aside from memory loss, are apathy, depression, agitation and general withdrawal. Apathy is the most prevalent disturbance, affecting about 70% of AD patients; depression ranks second, occurring in about 54% of patients; and agitation ranks third, appearing in about 50% of patients.4 Progressive neurostructural changes have been reported in adolescent and adult patients with BD,5, 6 associated with cognitive impairment.7 Although genome-wide studies have identified a number of potential risk alleles for BD and late-onset AD, the contribution of each is small and explains only a fraction of the known heritability.8, 9, 10, 11 High throughput genetic analysis confirms that neuropsychiatric disorders are very complex and involve many small interdependent genetic abnormalities that are influenced by polygenic inheritance, epigenetic interactions and pleiotropy.12 Several studies have implicated epigenetic mechanisms in these illnesses.12, 13 In this study, we examined brain epigenetic changes in AD and BD.
Epigenetic modification in the form of DNA methylation involves covalent addition of a methyl group from the methyl donor S-adenosylmethionine to a cytosine base within the DNA. This reaction is catalyzed by a family of DNA methyltransferases (DNMTs), with DNMT1 and DNMT3A being the main enzymes in mammalian brain.14, 15, 16 DNA methylation in the promoter region of a gene has been associated with decreased transcriptional activity.17 CpG islands are extended regions of cytosine and guanine repeats in the promoter region of many mammalian genes. These sites are heavily targeted by DNMTs and are known to modulate gene expression.18
Histones are basic proteins that regulate the compaction of chromatin and can undergo post-translational epigenetic modification by acetylation, methylation, phosphorylation, ubiquitination or sumoylation. Histone acetylation and phosphorylation have been linked to transcriptional activation,19, 20, 21, 22, 23 whereas trimethylation of histone-3K4 is suggested to silence gene expression, although the effects of this modification are still under investigation.24 Epigenetic studies in neuropsychiatric disorders may identify why behavioral phenotypes among patients are highly variable.15, 25
Recently, we reported that AD and BD postmortem frontal cortex (Brodmann area 9) shows upregulation of mRNA and protein levels of neuroinflammatory and arachidonic acid (AA) cascade markers such as AA-selective calcium-independent cytosolic phospholipase A2 (cPLA2)-IVA, secretory PLA2 (sPLA2-IIA and cyclooxygenase-2 (COX-2).1, 26 Loss of the synaptic proteins synaptophysin and drebrin is reported in both diseases,26, 27 and excitotoxicity is linked to increased glutamatergic function with loss of excitatory amino-acid transporters in both disorders.28, 29 Brain-derived neurotrophic factor (BDNF) also is reduced in the AD and BD brain.27, 30 The increased inflammatory and AA markers in BD are associated with increased mRNA and protein levels of transcriptional factor nuclear factor kappa B (NF-κB) subunits.2
It is not known if the altered mRNA and protein levels of AA cascade, neurotrophic, and synaptic protein markers in AD3 and BD1, 27 are associated with epigenetic modifications. To test this possibility, in this study we measured global DNA methylation and promoter region-specific CpG methylation for COX-2, 12-lipoxygenase (12-LOX), p450 expoxygenase, BDNF, CREB (cAMP response element-binding), synaptophysin, drebrin-like protein and NF-κB in the AD and BD frontal cortex tissue in which we had reported changes in mRNA and protein levels of AA cascade, inflammatory and synaptic markers.1, 2, 3, 27 We also measured total tissue histone phosphorylation and acetylation. Additionally, we measured mRNA levels of BDNF and NF-κB subunits in the AD brain samples, as this was not previously determined.26 Revealing epigenetic mechanisms that upregulate neuroinflammatory and AA cascade markers and reduce synaptic markers in these disorders could elucidate etiological mechanisms and identify new targets for pharmacotherapy and platforms for diagnosis. We studied frontal cortex because functional and structural abnormalities have been reported in this region in AD31, 32, 33, 34, 35, 36 and BD patients, and we had studied in previously.37, 38, 39, 40, 41, 42 An abstract of part of this work has been presented.43
Materials and methods
Postmortem brain samples
Frozen postmortem human frontal cortex samples (Brodmann area 9) from 10 AD patients and their 10 age-matched controls, and from 10 BD patients and their 10 age-matched controls, were provided by the Harvard Brain Tissue Resource Center (McLean Hospital, Belmont, MA, USA) under PHS grant number R24MH068855 to JS Rao. The protocol was approved by the Institutional Review Board of McLean Hospital, and by the Office of Human Subjects Research of the NIH (# 4380). Characteristics of the AD, BD and matched control subjects are described in detail elsewhere.2, 26 Briefly, age (years, control: 70.20±2.4 vs AD: 70.60±2.4; control: 43±3.5 vs BD: 49±7.2), postmortem interval (hours, control: 19.16±1.0 vs AD: 19.74±1.0; control: 27±1.5 vs BD: 21±3.0) and brain pH (control: 6.76±0.07 vs AD: 6.84±0.07; control: 6.6±0.16 vs BD: 6.7±0.09) did not differ significantly between the respective groups.
Genomic DNA isolation
Total genomic DNA was isolated from postmortem frontal cortex of AD and BD patients and controls using a GenElute Mammalian Genomic DNA Miniprep Kit (Sigma Aldrich, St Louis, MO, USA). Briefly, tissue was homogenized in lysis solution T and proteinase K solution, and incubated for 4 h at 55 °C in a shaking water bath. Genomic DNA was isolated according to the manufacturer's instructions.
Gene-specific DNA methylation determination
Gene-specific DNA methylation was determined with a OneStep qMethyl-Lite kit (Zymo Research, Irvine, CA, USA) and methyl primer (SABioscience, Frederick, MD, USA), with minor modifications. Promoter methylation for COX-2, BDNF, NF-κβ, CREB, 12-LOX, p450 epoxygenase, synaptophysin and drebrin-like genes were studied in AD, BD and control brains. Briefly, 20 ng of global DNA was incubated in the presence (test reaction) or absence (reference reaction) of methyl sensitive restriction enzymes (5 U each) (BStUI, HpyCH4IV and HpaII, NEB, Ipswich, MA, USA) at 37 °C for 2 h, followed by real-time reverse transcription PCR (RT-PCR) as described in the manufacturer's instructions. Percentage methylation was calculated using the formula 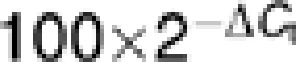 , where ΔCt is the average Ct value from the test reaction minus the average Ct value from the reference reaction. Percentage methylation is relative to each experiment.
, where ΔCt is the average Ct value from the test reaction minus the average Ct value from the reference reaction. Percentage methylation is relative to each experiment.
Total RNA isolation and real-time RT-PCR
Total RNA isolation and RT-PCR was done as described.2 mRNA levels of BDNF, NF-κB p50 and NF-κBp65 were measured by quantitative RT-PCR, using an ABI PRISM 7000 sequence detection system (Applied Biosystems, Carlsbad, CA, USA). Specific primers and probes for BDNF, NF-κB p50 and NF-κB p65 were part of the TaqMan gene expression assays (Applied Biosystems), and consisted of a 20 × mix of unlabeled PCR primers and Taqman minor groove binder probe (FAM dye-labeled). The fold-change in gene expression was determined by the ΔΔCT method.36 Data are expressed as the relative level of the target gene in the AD or BD brain normalized to the endogenous control (β-globulin) and relative to the control (calibrator). Experiments were carried out in duplicate.
Global DNA methylation determination
Global DNA methylation was determined from the genomic DNA of AD, BD and matched controls using an Imprint Methylated DNA Quantification Kit (Sigma Aldrich) following the manufacturer's recommendations. Values are expressed as percent of control.
Global histone, acetylation and phosphorylation determination
Nuclear extracts were prepared from frontal cortices of AD, BD and matched controls as previously described.44 Global acetylation (H3) and phosphorylation (H3) were measured in the nuclear extracts using ELISA kits (Epigentek Group, Farmingdale, NY, USA).
Statistical analysis
Data are expressed as mean±s.e.m. T-tests were used to compare AD and BD with matched control group samples.
Results
Hypomethylated COX-2 promoter region in AD and BD
Both AD and BD compared with respective control frontal cortex (Brodmann area 9) show increased AA cascade markers (protein and mRNA levels of cPLA2-IVA, sPLA2-IIA and COX-2).1, 3 There are no recognized CpG islands reported for PLA2 isoforms. However, the COX-2 promoter CpG region showed decreased methylation in both AD and BD brains (Figures 1a and b). There was no significant change in CpG island methylation for 12-LOX or p450 epoxygenase in either AD or BD (Figures 1c–f).
Figure 1.

Mean levels of CpG methylation at promoter region of cyclooxygenase-2 (COX-2), 12-lipoxygenase (12-LOX) and p450 epoxygenase in frontal cortex of Alzheimer's disease (AD) (a, c, e) and bipolar disorder (BD) patients (b, d, f). Mean levels of brain-derived neurotrophic factor (BDNF) mRNA in AD and control brain samples (g). Mean±s.e.m. (n=10 per group). *P<0.05, **P<0.01, ***P<0.001.
Hypermethylated BDNF promoter region in AD and BD
There was a significant decrease in BDNF mRNA in the AD brain (Figure 1g). The BDNF mRNA level in BD brain is published elsewhere and also is decreased significantly.27 Both brains showed a significantly increased methylation state of the promoter region of the BDNF gene (Figures 2a and b). The BDNF transcription factor CREB gene was significantly hypermethylated at its promoter region in the AD but not BD brain (Figures 2c and d).
Figure 2.

Mean CpG methylation at promoter region of brain-derived neurotrophic factor (BDNF), cAMP response element-binding (CREB) factor, synaptophysin and drebrin-like protein in frontal cortex of Alzheimer's disease (AD) (a, c, e, g) and bipolar disorder (BD) (b, d, f, h) patients and respective controls. Mean±s.e.m. (n=10 per group). *P<0.05, **P<0.01, ***P<0.001.
CpG methylation of synaptic markers in AD and BD
The changes observed in the AD and BD brains are associated with changes in the synaptic proteins, synaptophysin and drebrin.3, 15 There is no recognized CpG island region in the drebrin promoter. Instead, we examined the promoter of drebrin-like protein, which is involved in post-synaptic regulation.45 There was a significant increase in DNA methylation at the promoter region of synaptophysin in the AD not BD brain (Figures 2e and f). The promoter region of drebrin-like protein was significantly hypermethylated in BD but not AD brain (Figures 2g and h).
CpG methylation of NF-κB in AD and BD
BD and AD brains exhibit significantly increased mRNA and protein levels of neuroinflammatory markers such as IL-1β and TNF-α, and of markers of astrocytic and microglial activation.2, 3 NF-κB binding sites are present on the promoter region of gene transcripts of AA cascade markers, cPLA2-IVA, sPLA2-IIA and COX-246, 47, 48 and regulate transcription of proinflammatory genes.49, 50 We tested whether altered expression of these markers was associated with altered methylation states in the NF-κB transcription factor promoter region. The AD brain showed significantly decreased methylation of the NF-κB promoter CpG region, but this was not observed in the BD brain (Figures 3a and b). The hypomethylated state of the NF-κB promoter was accompanied by reciprocal increases in NF-κB p50 and p65 subunit mRNA expression (Figure 3c).
Figure 3.

Mean CpG methylation at promoter region of nuclear transcription factor kappa B (NF-κB) in frontal cortex of Alzheimer's disease (AD) (a) and bipolar disorder (BD) (b) patients and respective controls. Relative expression of NF-κB p50 and p65 mRNA levels in AD (c). Mean levels of global DNA methylation, global histone H3 phosphorylation and global histone H3 acetylation in frontal cortex of AD (d, f, h) and BD patients (e, g, i) and controls. Mean±s.e.m. (n=10 per group). *P<0.05, **P<0.01, ***P<0.001.
Increased global DNA methylation and altered global histone modification in AD and BD
The AD and BD brains showed significant increases in global DNA methylation compared with respective control levels (Figures 3d and e). These changes were associated with significant increases in H3 phosphorylation in both cases (Figures 3f and g). Global histone H3 acetylation was increased in the BD but not AD brain (Figures 3h and i).
Correlations with brain variables
Pearson correlations between the gene-specific methylation and histone modification levels in AD and BD brains treated separately on the one hand, and postmortem interval, age and pH on the other, were all statistically insignificant (P>0.05) (Table 2). Mean values of the three parameters did not differ significantly between AD and BD and respective control groups (Table 1).
Table 1. Probabilities and Pearson correlations between epigenetic changes in AD and BD brains and subject age, postmortem interval and pH.
|
Age |
PMI |
pH |
||||
|---|---|---|---|---|---|---|
| P-value | R2 | P-value | R2 | P-value | R2 | |
| Alzheimer's disease | ||||||
| DNA methylation | 0.63 | 0.01 | 0.58 | 0.01 | 0.31 | 0.05 |
| Histone acetylation | 0.32 | 0.05 | 0.510 | 0.02 | 0.08 | 0.16 |
| Histone phosphorylation | 0.25 | 0.07 | 0.61 | 0.01 | 0.37 | 0.04 |
| COX-2 | 0.71 | 0.00 | 0.38 | 0.04 | 0.94 | 0.00 |
| 12-LOX | 0.91 | 0.00 | 0.52 | 0.02 | 0.19 | 0.09 |
| P450 Epox. | 0.37 | 0.04 | 0.13 | 0.11 | 0.88 | 0.00 |
| SYP | 0.75 | 0.00 | 0.54 | 0.02 | 0.63 | 0.01 |
| NF-κB | 0.10 | 0.14 | 0.81 | 0.00 | 0.14 | 0.11 |
| CREB | 0.69 | 0.00 | 0.48 | 0.02 | 0.73 | 0.00 |
| BDNF | 0.25 | 0.07 | 0.21 | 0.08 | 0.52 | 0.02 |
| Drebrin-like protein | 0.37 | 0.04 | 0.08 | 0.15 | 0.88 | 0.00 |
| Bipolar disorder | ||||||
| DNA methylation | 0.83 | 0.002 | 0.83 | 0.00 | 0.10 | 0.13 |
| Histone acetylation | 0.55 | 0.02 | 0.55 | 0.02 | 0.54 | 0.02 |
| Histone phosphorylation | 0.91 | 0.00 | 0.91 | 0.00 | 0.39 | 0.04 |
| COX-2 | 0.79 | 0.00 | 0.05 | 0.18 | 0.44 | 0.03 |
| 12-LOX | 0.73 | 0.00 | 0.73 | 0.00 | 0.21 | 0.08 |
| P450 Epox. | 0.17 | 0.09 | 0.17 | 0.09 | 0.62 | 0.01 |
| SYP | 0.84 | 0.00 | 0.84 | 0.00 | 0.97 | 0.00 |
| NF-κB | 0.85 | 0.00 | 0.85 | 0.00 | 0.66 | 0.01 |
| CREB | 0.14 | 0.11 | 0.14 | 0.11 | 0.99 | 0.00 |
| BDNF | 0.32 | 0.05 | 0.32 | 0.05 | 0.63 | 0.01 |
| Drebrin-like protein | 0.81 | 0.00 | 0.19 | 0.09 | 0.47 | 0.02 |
Abbreviations: AD, Alzheimer's disease; BD, bipolar disorder; BDNF, brain-derived neurotrophic factor; COX-2, cyclooxygenase-2; CREB, cAMP response element-binding; 12-LOX, 12-lipoxygenase; NF-κB, nuclear transcription factor kappa B; SYP, synaptophysin; PMI, post-mortem interval.
Discussion
AD and BD are chronic progressive illnesses associated with upregulated mRNA and protein levels of neuroinflammatory markers (GFAP, CD11b, IL-1β) and of brain AA cascade enzymes (cPLA2-IVA, sPLA2-IIA and COX-2), as well as loss of neurotrophic factors (BDNF) and presynaptic and postsynaptic proteins (synaptophysin and drebrin).1, 2, 26, 27 Some of these alterations could be related to epigenetic modifications. The current study revealed that the increased COX-2 expression previously reported in the same AD and BD frontal cortex samples may partly be due to the hypomethylated state of the COX-2 CpG promoter region.26 However, other AA cascade markers, including 12-LOX and p450 epoxygenase, did not have DNA promoter methylation changes. Thus, the changes reported for 12-LOX and p450 epoxygenase mRNA and protein levels26 are unrelated to methylation at their gene promoter region. Protein and mRNA levels of cPLA2-IVA and sPLA2-IIA are upregulated in both disorders.1, 26 Human promoter regions of cPLA2 and sPLA2 isoforms and neuroinflammatory markers are reported to lack CpG islands (http://genome.ucsc.edu/), so we did not test their methylation states. The upregulation of cPLA2-IVA and sPLA2-IIA expression in both illnesses may have other causes, likely related to excitotoxicity and neuroinflammation.2, 3 Consistent with this suggestion, chronic NMDA receptor activation in a rat model for excitotoxicity induced neuroinflammation and increased expression of both enzymes.51, 52, 53
Studies have reported reductions of BDNF and synaptic proteins in both AD and BD brain.3, 27 Both disorders showed reduced mRNA levels of BDNF, which may be related to the observed hypermethylated state of the BDNF promoter region in the same tissues. Although similar promoter methylation patterns were found in COX-2 and BDNF in both AD and BD, other genes exhibited only disease-specific changes.
AD frontal cortex showed disease-specific hypermethylation in the promoter region of CREB, which may exacerbate reduced BDNF. Hypomethylation of NF-κB in the AD cortex may explain reported increased neuroinflammation due to upregulated NF-κB activity associated with its reduced methylation state. Furthermore, altered synaptic plasticity in AD is associated with reduced protein and mRNA levels of synaptophysin, which may be due to the hypermethylated state of its promoter region in AD brain samples. The difference in synaptophysin methylation between AD and BD may reflect a more rapid progression of AD, with clear histological evidence of synaptic loss.54, 55 The BD brain samples showed promoter hypermethylation of drebrin-like protein, which may contribute to the observed reduced drebrin mRNA and protein levels in BD.27 Although loss of drebrin has been reported in AD, the methylation state of drebrin-like protein in the AD brain remained unchanged. Loss of drebrin in BD may be related to epigenetic modifications, whereas in AD it may be related to other factors, including mitogen-activated kinase.56
Statistically significant changes were observed in global DNA methylation in both BD and AD. The significance of this change is not clear. Studies suggest that upregulated global DNA methylation is associated with decreased total gene expression.17 Interestingly, during aging, global chromosomal DNA is progressively hypomethylated, a trend also found in cancer cells.57, 58 Our observation of hypermethylated global DNA in AD and BD brains suggests that these disorders are epigenetically linked to decreased transcriptional activity. Both the AD and BD brain showed increased histone H3 phosphorylation, suggesting an onset of apoptosis and cell death.59 Neuronal damage implied by loss of synaptic proteins27, 28 may be due to the upregulated histone phosphorylation in AD and BD. Global histone H3 acetylation was increased in the BD but not AD brain. Some of the changes may be related to chronic medication. Further studies are required to understand chronic medication effects on epigenetic changes in brain.
Preclinical studies show that chronic administered anti-BD mood stabilizers reduce the activity or mRNA level of COX-2 in rat brain.60, 61, 62 Attenuation of COX-2 expression at the transcriptional or post-transcriptional level by drugs may not be sufficient to override epigenetic mechanisms at the COX-2 promoter region. In AD patients, COX-2 inhibitors failed to improve cognition,63, 64 which may be due to compensatory epigenetic modifications at the COX-2 promoter. Chronic mood-stabilizers and antipsychotic drugs increase neurotrophic factors in rat brain.65, 66, 67 Despite evidence of neuroprotection from these studies, postmortem brains from BD and AD patients showed loss of BDNF.27 Despite the advantages of mood stabilizers and antipsychotic drug treatments for AD and BD patients, chronic treatment with drugs not targeting epigenetic regulation may not provide full recovery. Drugs acting at the cellular level may provide transient protection by correcting neuroinflammatory and synaptic remodeling, but disease progression may reintroduce pathological changes due to epigenetic regulation. Understanding epigenetic mechanisms of genes targeted by current psychiatric drugs may help to establish new or more warranted therapeutic interventions. The basis for the epigenetic changes in these illnesses are not clear, possibly owing to inflammation, excitotoxicity, drug exposure or unknown factors. Revealing the epigenetic modifications may identify underlying mechanisms that influence synaptic loss and disease progression.
The Pearson's correlation did not show any significant influence of age, postmortem interval or pH of the samples in either illness. However, the current findings should be interpreted with caution, as only one brain region was studied, and because effects of chronic drug exposure on epigenetic modifications are not clear. Future studies should explore epigenetic modifications by mood stabilizers (lithium, valproate) and atypical antipsychotic drugs (olanzapine) in mouse models to clarify this issue, as genes that do not undergo epigenetic modifications in rats do so in mice (http://genome.ucsc.edu/).
In conclusion, both the BD and AD frontal cortex exhibits altered epigenetic regulation related to neuroinflammation, synaptic integrity, neuroprotection and AA metabolism (Table 2). These changes may modify disease progression and could help in identifying new therapeutic routes for treatment and diagnosis.
Table 2. Summary of epigenetic modifications in AD and BD frontal cortex.
| Epigenetic modification | AD | BD |
|---|---|---|
| Global DNA methylation | ↑ | ↑ |
| Global histone phosphorylation | ↑ | ↑ |
| Global histone H3 acetylation | ↔ | ↑ |
| COX-2 CpG methylation | ↓ | ↓ |
| 12-LOX CpG methylation | ↔ | ↔ |
| p450 Epoxygenase CpG methylation | ↔ | ↔ |
| BDNF CpG methylation | ↑ | ↑ |
| CREB CpG methylation | ↑ | ↔ |
| Synaptophysin CpG methylation | ↑ | ↔ |
| Drebrin-like protein CpG methylation | ↔ | ↑ |
| NF-κB CpG methylation | ↓ | ↔ |
Abbreviations: AD, Alzheimer's disease; BD, bipolar disorder; BDNF, brain-derived neurotrophic factor; COX-2, cyclooxygenase-2; CREB, cAMP response element-binding; 12-LOX, 12-lipoxygenase; NF-κB, nuclear transcription factor kappa B.
Down arrow indicates decrease and up arrow indicates increase, sideways arrow indicates in change.
Acknowledgments
We thank the NIH Fellows Editorial Board for editorial assistance. We thank the Harvard Brain Bank, Boston, MA for providing the postmortem brain samples under PHS grant number R24MH068855. This research was entirely supported by the Intramural Research Program of the National Institute on Aging, National Institutes of Health, Bethesda, MD, USA.
The authors declare no conflict of interest.
Supplementary Material
References
- Kim HW, Rapoport SI, Rao JS. Altered arachidonic acid cascade enzymes in postmortem brain from bipolar disorder patients. Mol Psychiatry. 2011;16:419–428. doi: 10.1038/mp.2009.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao JS, Harry GJ, Rapoport SI, Kim HW. Increased excitotoxicity and neuroinflammatory markers in postmortem frontal cortex from bipolar disorder patients. Mol Psychiatry. 2010;15:384–392. doi: 10.1038/mp.2009.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao JS, Rapoport S, Kim H-W. Altered neuroinflammatory, arachidonic acid cascade and synaptic markers in postmortem Alzheimer's disease brain. Transl Psychiatry. 2011;1:1–9. doi: 10.1038/tp.2011.27. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Frisoni GB, Rozzini L, Gozzetti A, Binetti G, Zanetti O, Bianchetti A, et al. Behavioral syndromes in Alzheimer's disease: description and correlates. Dement Geriatr Cogn Disord. 1999;10:130–138. doi: 10.1159/000017113. [DOI] [PubMed] [Google Scholar]
- Kalmar JH, Wang F, Spencer L, Edmiston E, Lacadie CM, Martin A, et al. Preliminary evidence for progressive prefrontal abnormalities in adolescents and young adults with bipolar disorder. J Int Neuropsychol Soc. 2009;15:476–481. doi: 10.1017/S1355617709090584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lisy ME, Jarvis KB, DelBello MP, Mills NP, Weber WA, Fleck D, et al. Progressive neurostructural changes in adolescent and adult patients with bipolar disorder. Bipolar disorders. 2011;13:396–405. doi: 10.1111/j.1399-5618.2011.00927.x. [DOI] [PubMed] [Google Scholar]
- Hellvin T, Sundet K, Simonsen C, Aminoff SR, Lagerberg TV, Andreassen OA, et al. Neurocognitive functioning in patients recently diagnosed with bipolar disorder. Bipolar disorders. 2012;14:227–238. doi: 10.1111/j.1399-5618.2012.01004.x. [DOI] [PubMed] [Google Scholar]
- Baum AE, Hamshere M, Green E, Cichon S, Rietschel M, Noethen MM, et al. Meta-analysis of two genome-wide association studies of bipolar disorder reveals important points of agreement. Mol Psychiatry. 2008;13:466–467. doi: 10.1038/mp.2008.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, Hunter DJ, et al. Finding the missing heritability of complex diseases. Nature. 2009;461:747–753. doi: 10.1038/nature08494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns LC, Minster RL, Demirci FY, Barmada MM, Ganguli M, Lopez OL, et al. Replication study of genome-wide associated SNPs with late-onset Alzheimer's disease. Am J Med Genet B Neuropsychiatr Genet. 2011;156B:507–512. doi: 10.1002/ajmg.b.31194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craddock N, Owen MJ, O'Donovan MC. The catechol-O-methyl transferase (COMT) gene as a candidate for psychiatric phenotypes: evidence and lessons. Mol Psychiatry. 2006;11:446–458. doi: 10.1038/sj.mp.4001808. [DOI] [PubMed] [Google Scholar]
- Abdolmaleky HM, Thiagalingam S, Wilcox M. Genetics and epigenetics in major psychiatric disorders: dilemmas, achievements, applications, and future scope. Am J Pharmacogenomics. 2005;5:149–160. doi: 10.2165/00129785-200505030-00002. [DOI] [PubMed] [Google Scholar]
- Cacabelos R. The application of functional genomics to Alzheimer's disease. Pharmacogenomics. 2003;4:597–621. doi: 10.1517/phgs.4.5.597.23795. [DOI] [PubMed] [Google Scholar]
- Laird PW. The power and the promise of DNA methylation markers. Nat Rev Cancer. 2003;3:253–266. doi: 10.1038/nrc1045. [DOI] [PubMed] [Google Scholar]
- Gräff J, Kim D, Dobbin MM, Tsai L-H. Epigenetic regulation of gene expression in physiological and pathological brain processes. Physiol Rev. 2011;91:603–649. doi: 10.1152/physrev.00012.2010. [DOI] [PubMed] [Google Scholar]
- Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Cell. 2007;pp:635–638. doi: 10.1016/j.cell.2007.02.006. [DOI] [PubMed] [Google Scholar]
- Robertson KD, Wolffe AP. DNA methylation in health and disease. Nat Rev Genet. 2000;1:11–19. doi: 10.1038/35049533. [DOI] [PubMed] [Google Scholar]
- Feng J, Zhou Y, Campbell SL, Le T, Li E, Sweatt JD, et al. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat Neurosci. 2010;13:423–430. doi: 10.1038/nn.2514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brownell JE, Zhou J, Ranalli T, Kobayashi R, Edmondson DG, Roth SY, et al. Tetrahymena histone acetyltransferase A: a homolog to yeast Gcn5p linking histone acetylation to gene activation. Cell. 1996;84:843–851. doi: 10.1016/s0092-8674(00)81063-6. [DOI] [PubMed] [Google Scholar]
- Loury R, Sassone-Corsi P. Histone phosphorylation: how to proceed. Methods. 2003;31:40–48. doi: 10.1016/s1046-2023(03)00086-0. [DOI] [PubMed] [Google Scholar]
- Shiio Y, Eisenman RN. Histone sumoylation is associated with transcriptional repression. Proc Natl Acad Sci USA. 2003;100:13225–13230. doi: 10.1073/pnas.1735528100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukuda H, Sano N, Muto S, Horikoshi M. Simple histone acetylation plays a complex role in the regulation of gene expression. Brief Funct Genomic Proteomic. 2006;5:190–208. doi: 10.1093/bfgp/ell032. [DOI] [PubMed] [Google Scholar]
- Shilatifard A. Chromatin modifications by methylation and ubiquitination: implications in the regulation of gene expression. Annu Rev Biochem. 2006;75:243–269. doi: 10.1146/annurev.biochem.75.103004.142422. [DOI] [PubMed] [Google Scholar]
- Li B, Carey M, Workman JL. The role of chromatin during transcription. Cell. 2007;128:707–719. doi: 10.1016/j.cell.2007.01.015. [DOI] [PubMed] [Google Scholar]
- Tsankova N, Renthal W, Kumar A, Nestler EJ. Epigenetic regulation in psychiatric disorders. Nat Rev Neurosci. 2007;8:355–367. doi: 10.1038/nrn2132. [DOI] [PubMed] [Google Scholar]
- Rao JS, Rapoport SI, Kim HW. Altered neuroinflammatory, arachidonic acid cascade and synaptic markers in postmortem Alzheimer's disease brain. Transl Psychiatry. 2011;1:e31. doi: 10.1038/tp.2011.27. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Kim HW, Rapoport SI, Rao JS. Altered expression of apoptotic factors and synaptic markers in postmortem brain from bipolar disorder patients. Neurobiol Dis. 2010;37:596–603. doi: 10.1016/j.nbd.2009.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen KH, Reese EA, Kim HW, Rapoport SI, Rao JS. Disturbed neurotransmitter transporter expression in Alzheimer's disease brain. J Alzheimers Dis. 2011;26:755–766. doi: 10.3233/JAD-2011-110002. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Aoki C, Kojima N, Sabaliauskas N, Shah L, Ahmed TH, Oakford J, et al. Drebrin a knockout eliminates the rapid form of homeostatic synaptic plasticity at excitatory synapses of intact adult cerebral cortex. J Comp Neurol. 2009;517:105–121. doi: 10.1002/cne.22137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aisa B, Gil-Bea FJ, Solas M, Garcia-Alloza M, Chen CP, Lai MK, et al. Altered NCAM expression associated with the cholinergic system in Alzheimer's disease. J Alzheimer's Dis. 2010;20:659–668. doi: 10.3233/JAD-2010-1398. [DOI] [PubMed] [Google Scholar]
- Armstrong RA. Is the clustering of beta-amyloid (A beta) deposits in the frontal cortex of Alzheimer patients determined by blood vessels. Neurosci Lett. 1995;195:121–124. doi: 10.1016/0304-3940(95)11796-y. [DOI] [PubMed] [Google Scholar]
- Armstrong RA, Cairns NJ, Lantos PL. Spatial distribution of diffuse, primitive, and classic amyloid-beta deposits and blood vessels in the upper laminae of the frontal cortex in Alzheimer disease. Alzheimer Dis Assoc Disord. 1998;12:378–383. doi: 10.1097/00002093-199812000-00021. [DOI] [PubMed] [Google Scholar]
- Fischer B, Popa-Wagner A. [Alzheimer disease: involvement of the complement system in cell death. Gene expression of C1q and C3 in the frontal cortex of patients with Alzheimer disease and control probands] Fortschritte der Medizin. 1996;114:161–163. [PubMed] [Google Scholar]
- Nordberg A, Winblad B. Reduced number of [3H]nicotine and [3H]acetylcholine binding sites in the frontal cortex of Alzheimer brains. Neurosci Lett. 1986;72:115–119. doi: 10.1016/0304-3940(86)90629-4. [DOI] [PubMed] [Google Scholar]
- Soininen H, Kosunen O, Helisalmi S, Mannermaa A, Paljarvi L, Talasniemi S, et al. A severe loss of choline acetyltransferase in the frontal cortex of Alzheimer patients carrying apolipoprotein epsilon 4 allele. Neurosci Lett. 1995;187:79–82. doi: 10.1016/0304-3940(95)11343-6. [DOI] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- Lopez-Larson MP, DelBello MP, Zimmerman ME, Schwiers ML, Strakowski SM. Regional prefrontal gray and white matter abnormalities in bipolar disorder. Biol Psychiatry. 2002;52:93–100. doi: 10.1016/s0006-3223(02)01350-1. [DOI] [PubMed] [Google Scholar]
- Lyoo IK, Kim MJ, Stoll AL, Demopulos CM, Parow AM, Dager SR, et al. Frontal lobe gray matter density decreases in bipolar I disorder. Biol Psychiatry. 2004;55:648–651. doi: 10.1016/j.biopsych.2003.10.017. [DOI] [PubMed] [Google Scholar]
- Rajkowska G. Cell pathology in bipolar disorder. Bipolar Disord. 2002;4:105–116. doi: 10.1034/j.1399-5618.2002.01149.x. [DOI] [PubMed] [Google Scholar]
- Buchsbaum MS, Wu J, DeLisi LE, Holcomb H, Kessler R, Johnson J, et al. Frontal cortex and basal ganglia metabolic rates assessed by positron emission tomography with [18F]2-deoxyglucose in affective illness. J Affect Disord. 1986;10:137–152. doi: 10.1016/0165-0327(86)90036-4. [DOI] [PubMed] [Google Scholar]
- Rubinsztein JS, Fletcher PC, Rogers RD, Ho LW, Aigbirhio FI, Paykel ES, et al. Decision-making in mania: a PET study. Brain. 2001;124:2550–2563. doi: 10.1093/brain/124.12.2550. [DOI] [PubMed] [Google Scholar]
- Suhara T, Nakayama K, Inoue O, Fukuda H, Shimizu M, Mori A, et al. D1 dopamine receptor binding in mood disorders measured by positron emission tomography. Psychopharmacology. 1992;106:14–18. doi: 10.1007/BF02253582. [DOI] [PubMed] [Google Scholar]
- Rao JS, Klein S, VL K, Reese EA, Rapoport SI.Epigenetic modifications in postmortem frontal cortex from bipolar disorder and Alzheimer's disease patients50th Annual Meeting of American College of Neuropsychopharmacology; 05 December 2011; Hawaii, USA;2011. p. 226.
- Bazinet RP, Rao JS, Chang L, Rapoport SI, Lee HJ. Chronic carbamazepine decreases the incorporation rate and turnover of arachidonic acid but not docosahexaenoic acid in brain phospholipids of the unanesthetized rat: relevance to bipolar disorder. Biol Psychiatry. 2006;59:401–407. doi: 10.1016/j.biopsych.2005.07.024. [DOI] [PubMed] [Google Scholar]
- Haeckel A, Ahuja R, Gundelfinger ED, Qualmann B, Kessels MM. The actin-binding protein Abp1 controls dendritic spine morphology and is important for spine head and synapse formation. J Neurosci. 2008;28:10031–10044. doi: 10.1523/JNEUROSCI.0336-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morri H, Ozaki M, Watanabe Y. 5′-flanking region surrounding a human cytosolic phospholipase A2 gene. Biochem Biophys Res Commun. 1994;205:6–11. doi: 10.1006/bbrc.1994.2621. [DOI] [PubMed] [Google Scholar]
- Antonio V, Brouillet A, Janvier B, Monne C, Bereziat G, Andreani M, et al. Transcriptional regulation of the rat type IIA phospholipase A2 gene by cAMP and interleukin-1beta in vascular smooth muscle cells: interplay of the CCAAT/enhancer binding protein (C/EBP), nuclear factor-kappaB and Ets transcription factors. Biochem J. 2002;368:415–424. doi: 10.1042/BJ20020658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanabe T, Tohnai N. Cyclooxygenase isozymes and their gene structures and expression. Prostaglandins Other Lipid Mediat. 2002;68–69:95–114. doi: 10.1016/s0090-6980(02)00024-2. [DOI] [PubMed] [Google Scholar]
- Pereira SG, Oakley F. Nuclear factor-kappaB1: regulation and function. Int J Biochem Cell Biol. 2008;40:1425–1430. doi: 10.1016/j.biocel.2007.05.004. [DOI] [PubMed] [Google Scholar]
- Niederberger E, Schmidtko A, Gao W, Kuhlein H, Ehnert C, Geisslinger G. Impaired acute and inflammatory nociception in mice lacking the p50 subunit of NF-kappaB. Eur J Pharmacol. 2007;559:55–60. doi: 10.1016/j.ejphar.2006.11.074. [DOI] [PubMed] [Google Scholar]
- Rao JS, Ertley RN, Rapoport SI, Bazinet RP, Lee HJ. Chronic NMDA administration to rats up-regulates frontal cortex cytosolic phospholipase A2 and its transcription factor, activator protein-2. J Neurochem. 2007;102:1918–1927. doi: 10.1111/j.1471-4159.2007.04648.x. [DOI] [PubMed] [Google Scholar]
- Li W, Xia J, Sun GY. Cytokine induction of iNOS and sPLA2 in immortalized astrocytes (DITNC): response to genistein and pyrrolidine dithiocarbamate. J Interferon Cytokine Res. 1999;19:121–127. doi: 10.1089/107999099314261. [DOI] [PubMed] [Google Scholar]
- Chang Y, Kim H-W, Rapoport S, Rao J. Chronic NMDA administration increases neuroinflammatory markers in rat frontal cortex: cross-talk between excitotoxicity and neuroinflammation. Neurochem Res. 2008;33:2318–2323. doi: 10.1007/s11064-008-9731-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terry RD. The pathogenesis of Alzheimer disease: an alternative to the amyloid hypothesis. J Neuropathol Exp Neurol. 1996;55:1023–1025. [PubMed] [Google Scholar]
- Scheff SW, Price DA, Schmitt FA, DeKosky ST, Mufson EJ. Synaptic alterations in CA1 in mild Alzheimer disease and mild cognitive impairment. Neurology. 2007;68:1501–1508. doi: 10.1212/01.wnl.0000260698.46517.8f. [DOI] [PubMed] [Google Scholar]
- Maezawa I, Nivison M, Montine KS, Maeda N, Montine TJ. Neurotoxicity from innate immune response is greatest with targeted replacement of E4 allele of apolipoprotein E gene and is mediated by microglial p38MAPK. FASEB J. 2006;20:797–799. doi: 10.1096/fj.05-5423fje. [DOI] [PubMed] [Google Scholar]
- Bellizzi D, D'Aquila P, Montesanto A, Corsonello A, Mari V, Mazzei B, et al. Global DNA methylation in old subjects is correlated with frailty. Age. 2012;34:169–179. doi: 10.1007/s11357-011-9216-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cadieux B, Ching TT, VandenBerg SR, Costello JF. Genome-wide hypomethylation in human glioblastomas associated with specific copy number alteration, methylenetetrahydrofolate reductase allele status, and increased proliferation. Cancer Res. 2006;66:8469–8476. doi: 10.1158/0008-5472.CAN-06-1547. [DOI] [PubMed] [Google Scholar]
- Tikoo K, Lau SS, Monks TJ. Histone H3 phosphorylation is coupled to poly-(ADP-ribosylation) during reactive oxygen species-induced cell death in renal proximal tubular epithelial cells. Mol Pharmacol. 2001;60:394–402. doi: 10.1124/mol.60.2.394. [DOI] [PubMed] [Google Scholar]
- Rapoport SI, Basselin M, Kim HW, Rao JS. Bipolar disorder and mechanisms of action of mood stabilizers. Brain Res Rev. 2009;61:185–209. doi: 10.1016/j.brainresrev.2009.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao JS, Rapoport SI. Mood-stabilizers target the brain arachidonic acid cascade. Curr Mol Pharmacol. 2009;2:207–214. doi: 10.2174/1874467210902020207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao JS, Bazinet RP, Rapoport SI, Lee HJ. Chronic treatment of rats with sodium valproate downregulates frontal cortex NF-kappaB DNA binding activity and COX-2 mRNA. Bipolar Disord. 2007;9:513–520. doi: 10.1111/j.1399-5618.2007.00361.x. [DOI] [PubMed] [Google Scholar]
- Aisen PS, Schafer KA, Grundman M, Pfeiffer E, Sano M, Davis KL, et al. Effects of rofecoxib or naproxen vs placebo on Alzheimer disease progression: a randomized controlled trial. JAMA. 2003;289:2819–2826. doi: 10.1001/jama.289.21.2819. [DOI] [PubMed] [Google Scholar]
- Thal LJ, Ferris SH, Kirby L, Block GA, Lines CR, Yuen E, et al. A randomized, double-blind, study of rofecoxib in patients with mild cognitive impairment. Neuropsychopharmacology. 2005;30:1204–1215. doi: 10.1038/sj.npp.1300690. [DOI] [PubMed] [Google Scholar]
- Chang YC, Rapoport SI, Rao JS. Chronic administration of mood stabilizers upregulates BDNF and bcl-2 expression levels in rat frontal cortex. Neurochem Res. 2009;34:536–541. doi: 10.1007/s11064-008-9817-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang DM. Neuroprotective and neurotrophic actions of the mood stabilizer lithium: can it be used to treat neurodegenerative diseases. Crit Rev Neurobiol. 2004;16:83–90. doi: 10.1615/critrevneurobiol.v16.i12.90. [DOI] [PubMed] [Google Scholar]
- Keilhoff G, Grecksch G, Bernstein HG, Roskoden T, Becker A. Risperidone and haloperidol promote survival of stem cells in the rat hippocampus. Eur Arch Psychiatry Clin Neurosci. 2010;260:151–162. doi: 10.1007/s00406-009-0033-1. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.