

Abstract
Purpose
Extracellular matrix (ECM) deposits lead to elevated resistance of aqueous humor outflow which play an important role in the development of primary open angle glaucoma (POAG). The TGF-β2 (transforming growth factor β)/Smad (signaling mathers against decapentaplegic) pathway is known to regulate the ECM deposits. In this study, we determined the effect of Smad7 siRNA transfection in inhibiting the expression of ECM components.
Methods
Plasmid containing Smad7 siRNA was used to transfect cultured human trabecular meshwork cells (HTM). Protein expression of Smad7, fibronectin, and laminin was determined using western blot.
Results
Downregulation of Smad7 interrupts the effects of TGF-β2 on the expression of several ECM components. Smad7 siRNA can partially decrease the expression of Smad7, fibronectin, and laminin.
Conclusions
Smad7 plays an important role in regulating the ECM protein in the aqueous outflow pathway.
Introduction
Primary open angle glaucoma (POAG) is a leading cause of blindness in the world [1,2]. Elevated intraocular pressure (IOP), consequence of high resistance to aqueous outflow (AH), is an important risk factor in the development and progression of POAG [2]. Hynes [3] has shown that elevated IOP is associated with increased in outflow resistance in the trabecular meshwork (TM) and is related to elevated deposition of extracellular matrix (ECM) material within the TM. Recent studys have found that transforming growth factor-beta 2 (TGF-β2), known to regulate the ECM metabolism including fibronectin, collagen, and elastin, is elevated in the aqueous humor and TM of the glaucoma patient [3,4]. Since the TGF-β/Smad (signaling mathers against decapentaplegic) pathway is important in regulation of ECM deposition in the TM [5], inhibitory Smad7 could potentially antagonize TGF- β/Smad dependent signaling, which induces degration of TGF-β receptor and prevents phosphorylation of Smad2/3 [6,7].
In t5he present study, we determined effect of Smad7 siRNA in inhibiting the expression of ECM components, including fibronectin and laminin in human trabecular meshwork (HTM) cells.
Methods
Trabecular meshwork cell culture and TGF-β2 treatment
Cultures of HTM cells were established from the eyes of five human donors. The research adhered to the tenets of the Declaration of Helsinki. Written informed consent was obtained from all the patients before tissues were collected. This study and all the procedures were approved by the Ethics Committee of the University of Harbin Medical University. The dissection procedure was performed with sterile instruments under a laminar flow hood. The lens, cornea, retina, iris, and ciliary body were extracted first. Then HTM cells between Descement's membrane and the scleral spur were dissected using fine forceps and placed in a 35 mm2 culture dish where cells were adhered to the plastic. The cell culture medium, Dulbecco’s Modified Eagle’s Medium (DMEM; low glucose) supplemented with 10% fetal bovine serum (FBS), L-glutamine (0.292 mg/ml), penicillin (100 units/ml), streptomycin (0.1 mg/ml), and amphotericin B (4 mg/ml; HyClone Labs, Logan, UT), was changed every 2 days. HTM cells between passages 5 and 8 were [8-10]. For the TGF-β2 (Sigma Aldrich, St. Louis, MO) treatment group, cells were serum starved for 24 h before treatment with 1 ng/ml TGF-β2 for 24 h [11,12].
Construction of plasmid with Smad7 siRNA
Vector pSuppressorNeo (Imgenex, San Diego, CA) is a vector used to generate biologically active siRNAs from the U6promoter. Synthetic oligonucleotide primers (5′- AGG UCA CCA CCA UCC CCA CUU-3′ and 5′-GUG GGG AUG GUG GUG ACC UUU-3′) were annealed and introduced into pSuppressor Neovector [13].
Transfection HTM with pSup-Smad7 siRNA
HTM transfected with plasmid containing pSup-Smad7 siRNA, empty vector only, or medium were served as experimental, vehicle control, and blank control groups, respectively. Transfection was performed in 60 mm plates using 3 µg (1 µg/µl) vector in 10 µl of Metafectene Pro reagent (Biontex, Martinstried, Germany). After 48 h of transfection, cells were treated with G418 (HyClone Labs) for 2 weeks for positive clone selection. After G418 treatment, several stable transfected cells were cloned. Each clone was screened for expression of HTM by western blot analysis [14].
Western blot analysis
Conditioned medium was collected from HTM cells after treatment with Smad7 siRNA in serum-free medium containing 0.5 mg/ml BSA (HyClone Labs). Protein concentration was measured using absorbance spectroscopy. Protein was separated on a 10% SDS-polyacrylamide gel and transferred to nitrocellulose membranes. After blocking with 5% nonfat milk, membranes were incubated with primary antibodies against Smad7, fibronectin, and laminin (Santa cruz biotechnology Inc., Santa Cruz, CA) overnight at 4 °C, followed by incubation with secondary antibodies. The membrane was then assayed using the enhanced chemiluminescent kit (ECL, Thermo Scientific, Rockford, IL) and scanned with ChemiDoc™Doc XRS+ system (Bio-Rad, Hercules, CA). The density of each band was obtained using Quantity One 4.6.2 basic software (Bio-Rad). Values were expressed as fold change relative to control and normalized to a loading control, glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Santa Cruz biotechnology Inc.) [15].
Statistical analysis
The data were analyzed by the two-tailed Student t-test using SPSS 10.0 (IBM Inc., Beijing, China) and a p<0.05 was considered significant.
Results
Downregulation of Smad7 after transfection with pSup-Smad7 siRNA
Decreased expression of Smad7 was detected in HTM transfected with Smad7 siRNA compared with the TGF-β2 group, control group and TGF-β2 plus vehicle group (p<0.01; Figure 1).
Figure 1.
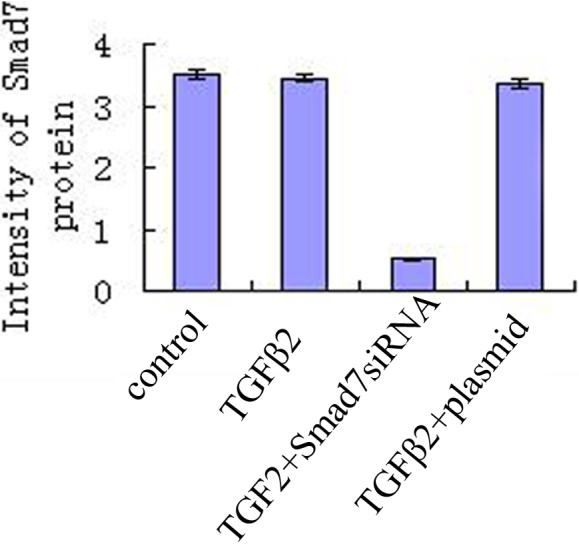
Downregulation of Smad7 after transfection with pSup-Smad7 siRNA. There was significance among the expression of Smad7 in HTM transfected with Smad7 siRNA, TGF-β2 group, control group, and TGF-β2 plus vehicle group (p<0.01).
Downregulation of expression of fibronectin by transfection with pSup-Smad7 siRNA
Fibronectin protein was expressed in the HTM transfected with Smad7 siRNA, TGF-β2 group, control group and TGF-β2 plus vehicle group (p<0.01). Downregulation of fibronectin was detected in HTM transfected with pSup-Smad7 siRNA (Figure 2).
Figure 2.
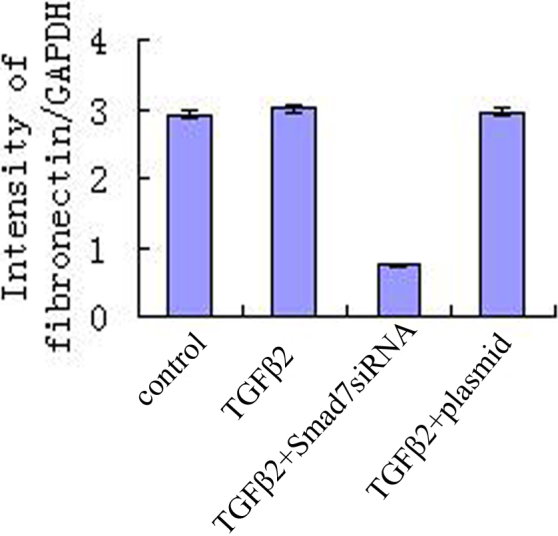
Downregulation expression of fibronectin by transfection with pSup-Smad7 siRNA. Fibronectin is a glycoprotein of the extracellular matrix that binds to membrane-spanning receptor proteins called integrins. There is significance among expression of fibronectin protein in the HTM transfected with Smad7siRNA, TGF-β2 group, control group, and TGF-β2 plus vehicle group (p<0.01). Downregulation of fibronectin was detected in HTM transfected with pSup-Smad7 siRNA.
Inhibition of expression of laminin after transfection with pSup-Smad7 siRNA
Laminin protein was expressed in the HTM transfected with Smad7 siRNA, TGF-β2 group, control group, and TGF-β2 plus vehicle group (p<0.01). Downregulation of laminin was detected in HTM transfected with pSup-Smad7 siRNA (Figure 3).
Figure 3.
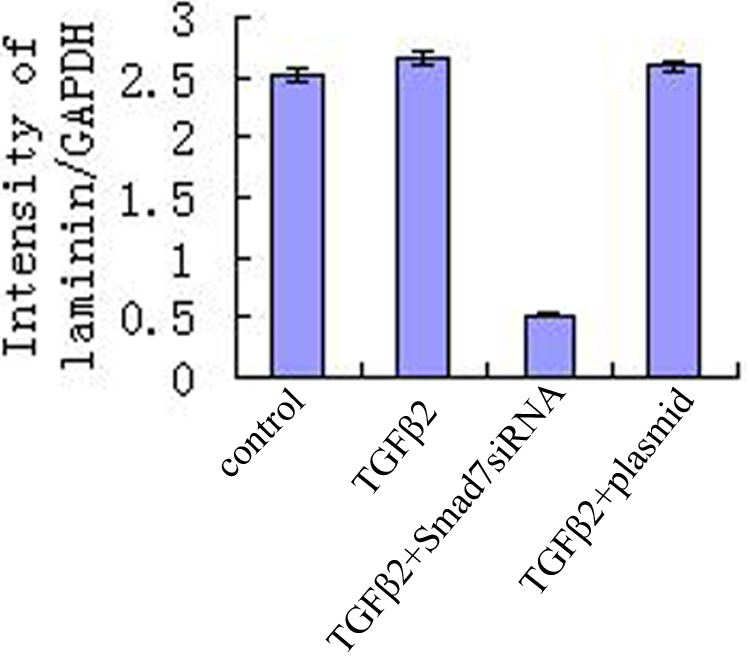
Inhibit expression of laminin after transfection with pSup-Smad7 siRNA. There is significance among expression of laminin protein in the HTM of Smad7siRNA group, TGF-β2 group, control group, and TGF-β2 plus vehicle group (p<0.01). Downregulation of laminin was detected in HTM transfected with pSup-Smad7 siRNA.
Discussion
Abnormal accumulation of ECM components can cause fibrosis [16-18]. Additionally, irregularities in multiple pathways involved in tissue repair and inflammation can lead to the development of heart, kidney, lung fibrosis [19,20]. ECM components such as collagens, fibronectin, laminin, and elastin affect the development of fibrosis [21-23]. Similarly, abnormal accumulation of ECM can lead to elevated resistance in the TM, especially in the juxtacanalicular region, which has been shown to be the major resistance site for the elevated IOP of POAG in the aqueous outflow pathway [24,25].
TGF-β2 is an important ligand that modulates cell behavior in ocular tissues including enhancing ECM production [26,27], and its gene targets are activated by the translocation of Smad2 and Smad3, and their common mediator Smad4 [28,29]. There are three groups of Smad proteins: receptor-regulated Smads, common-partner Smad, and inhibitory Smads [30-32]. Inhibition of TGF-β family signaling by Smad6 and Smad7 through multiple mechanisms plays a critical role in various physiologic processes [33]. Decreased expression of Smad7 has been reported in patients with scleroderma and inflammatory bowel disease [34].
Our study showed that transfection with pSup-Smad7 siRNA can downregulate Smad7 expression of cultured HTM cells. In summary, this study has delineated the role of Smad7 downregulation in inhibition the expression of ECM protein including fibronectin and laminin of cultured HTM. Downregulation expression of fibronectin and laminin after transfection with pSup-Smad7 siRNA suggest its role in the TGF-β2/Smad pathway in regulating ECM deposits.
In conclusion, the present work demonstrates for the first time that transfection with pSup-Smad7 siRNA can effectively inhibit ECM deposits of HTM treated with TGF-β2 in vitro which indicates it can be novel target for treatment of POAG. Although Smad7 is a powerful way to inhibit the expression of ECM components, further work is needed before this approach can be used for the treatment of human disease.
Acknowledgments
This work was supported by the grant of nature science science foundation of China (81100659), the grant of nature science foundation of Heilongjiang province of China (No. D2007–80), Scientific foundation of education ministry of China (20092307120003), and Postdoctral foundation of China (20080430137, 200902418). Dr. Feng Wang and Dr. Shiguang Zhao contributed equally to the work in this publication and can be considered as co-corresponding authors.
References
- 1.Quigley HA, Broman AT. The number of people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol. 2006;90:262–7. doi: 10.1136/bjo.2005.081224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lütjen-Drecoll E. Morphological changes in glaucomatous eyes and the role of TGFbeta2 for the pathogenesis of the disease. Exp Eye Res. 2005;81:1–4. doi: 10.1016/j.exer.2005.02.008. [DOI] [PubMed] [Google Scholar]
- 3.Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009;326:1216–9. doi: 10.1126/science.1176009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chung EJ, Lee HK, Jung SA, Lee SJ, Chee HY, Sohn YH, Lee JH. Transduction of PTEN proteins using the tat domain modulates TGF-β1-mediated signaling pathways and transdifferentiation in subconjunctival fibroblasts. Invest Ophthalmol Vis Sci. 2012;53:379–86. doi: 10.1167/iovs.11-8491. [DOI] [PubMed] [Google Scholar]
- 5.Takai Y, Tanito M, Ohira A. Multiplex cytokine analysis of aqueous humor in eyes with primary open-angle glaucoma, exfoliation glaucoma, and cataract. Invest Ophthalmol Vis Sci. 2012;53:241–7. doi: 10.1167/iovs.11-8434. [DOI] [PubMed] [Google Scholar]
- 6.Zenkel M, Krysta A, Pasutto F, Juenemann A, Kruse FE, Schlötzer-Schrehardt U. Regulation of lysyl oxidase-like 1 (LOXL1) and elastin-related genes by pathogenic factors associated with pseudoexfoliation syndrome. Invest Ophthalmol Vis Sci. 2011;52:8488–95. doi: 10.1167/iovs.11-8361. [DOI] [PubMed] [Google Scholar]
- 7.Tektas OY, Lutjen-Drecoll E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp Eye Res. 2009;88:769–75. doi: 10.1016/j.exer.2008.11.025. [DOI] [PubMed] [Google Scholar]
- 8.Roberts AB, Heine UI, Flanders KC, Sporn MB. Transforming growth factor-beta. Major role in regulation of extracellular matrix. Ann N Y Acad Sci. 1990;580:225–32. doi: 10.1111/j.1749-6632.1990.tb17931.x. [DOI] [PubMed] [Google Scholar]
- 9.Zenkel M, Krysta A, Pasutto F, Juenemann A, Kruse FE, Schlötzer-Schrehardt U. Regulation of lysyl oxidase-like 1 (LOXL1) and elastin-related genes by pathogenic factors associated with pseudoexfoliation syndrome. Invest Ophthalmol Vis Sci. 2011;52:8488–95. doi: 10.1167/iovs.11-8361. [DOI] [PubMed] [Google Scholar]
- 10.Bollinger KE, Crabb JS, Yuan X, Putliwala T, Clark AF, Crabb JW. Quantitative proteomics: TGFβ₂ signaling in trabecular meshwork cells. Invest Ophthalmol Vis Sci. 2011;52:8287–94. doi: 10.1167/iovs.11-8218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Han H, Wecker T, Grehn F, Schlunck G. Elasticity-dependent modulation of TGF-β responses in human trabecular meshwork cells. Invest Ophthalmol Vis Sci. 2011;52:2889–96. doi: 10.1167/iovs.10-6640. [DOI] [PubMed] [Google Scholar]
- 12.Yu AL, Birke K, Moriniere J, Welge-Lüssen U. TGF-{beta}2 induces senescence-associated changes in human trabecular meshwork cells. Invest Ophthalmol Vis Sci. 2010;51:5718–23. doi: 10.1167/iovs.10-5679. [DOI] [PubMed] [Google Scholar]
- 13.Robertson JV, Golesic E, Gauldie J, West-Mays JA. Ocular gene transfer of active TGF-beta induces changes in anterior segment morphology and elevated IOP in rats. Invest Ophthalmol Vis Sci. 2010;51:308–18. doi: 10.1167/iovs.09-3380. [DOI] [PubMed] [Google Scholar]
- 14.Wang F, Qi LX, Su Y, Yan QH, Teng Y. Inhibition of Cell Proliferation of Tenon’s Capsule Fibroblast by S-Phase Kinase-Interacting Protein 2. Targeting SiRNA through Increasing p27 Protein Level. Invest Ophthalmol Vis Sci. 2010;51:1475–82. doi: 10.1167/iovs.09-4363. [DOI] [PubMed] [Google Scholar]
- 15.Su Y, Wang F, Yan Q, Teng Y, Cui H. Inhibition of proliferation of rabbit lens epithelial cells by S-phase kinase-interacting protein 2 targeting small interfering RNA. Mol Vis. 2010;16:907–15. [PMC free article] [PubMed] [Google Scholar]
- 16.Laping NJ, Grygielko E, Mathur A. Inhibition of transforming growth factor (TGF)-beta1-induced extracellular matrix with a novel inhibitor of the TGF-beta type I receptor kinase activity: SB-431542. Mol Pharmacol. 2002;62:58–64. doi: 10.1124/mol.62.1.58. [DOI] [PubMed] [Google Scholar]
- 17.Sawyer JS, Anderson BD, Beight DW. Synthesis and activity of new aryl- and heteroaryl-substituted pyrazole inhibitors of the transforming growth factor-beta type I receptor kinase domain. J Med Chem. 2003;46:3953–6. doi: 10.1021/jm0205705. [DOI] [PubMed] [Google Scholar]
- 18.Tripathi RC, Chan WF, Li J, Tripathi BJ. Trabecular cells express the TGF-beta 2 gene and secrete the cytokine. Exp Eye Res. 1994;58:523–8. doi: 10.1006/exer.1994.1046. [DOI] [PubMed] [Google Scholar]
- 19.Fuchshofer R, Welge-Lussen U, Lutjen-Drecoll E. The effect of TGF-beta2 on human trabecular meshwork extracellular proteolytic system. Exp Eye Res. 2003;77:757–65. doi: 10.1016/s0014-4835(03)00220-3. [DOI] [PubMed] [Google Scholar]
- 20.Junglas B, Yu AH, Welge-Lussen U, Tamm ER, Fuchshofer R. Connective tissue growth factor induces extracellular matrix deposition in human trabecular meshwork cells. Exp Eye Res. 2009;88:1065–75. doi: 10.1016/j.exer.2009.01.008. [DOI] [PubMed] [Google Scholar]
- 21.Parsons CJ, Takashima M, Rippe RA. Molecular mechanisms of hepatic fibrogenesis. J Gastroenterol Hepatol. 2007;22(suppl 1):S79–84. doi: 10.1111/j.1440-1746.2006.04659.x. [DOI] [PubMed] [Google Scholar]
- 22.Omori S, Kitagawa H, Koike J. Activated extracellular signalregulated kinase correlates with cyst formation and transforming growth factor-beta expression in fetal obstructive uropathy. Kidney Int. 2008;73:1031–7. doi: 10.1038/ki.2008.3. [DOI] [PubMed] [Google Scholar]
- 23.Dobaczewski M, Gonzalez-Quesada C, Frangogiannis NG. The extracellular matrix as a modulator of the inflammatory and reparative response following myocardial infarction. J Mol Cell Cardiol. 2010;48:504–11. doi: 10.1016/j.yjmcc.2009.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gill S, Wight TN, Frevert CW. Proteoglycans: key regulators of pulmonary inflammation and the innate immune response to lung infection. Anat Rec (Hoboken) 2010;293:968–81. doi: 10.1002/ar.21094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Evanko SP, Potter-Perigo S, Johnson PY, Wight TN. Organization of hyaluronan and versican in the extracellular matrix of human fibroblasts treated with the viral mimetic poly I:C. J Histochem Cytochem. 2009;57:1041–60. doi: 10.1369/jhc.2009.953802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ng YY, Hou CC, Wang W, Huang XR, Lan HY. Blockade of NFkappaB activation and renal inflammation by ultrasoundmediated gene transfer of Smad7 in rat remnant kidney. Kidney Int Suppl. 2005;94:S83–91. doi: 10.1111/j.1523-1755.2005.09421.x. [DOI] [PubMed] [Google Scholar]
- 27.Wang W, Huang XR, Li AG. Signaling mechanism of TGF-beta1 in prevention of renal inflammation: role of Smad7. J Am Soc Nephrol. 2005;16:1371–83. doi: 10.1681/ASN.2004121070. [DOI] [PubMed] [Google Scholar]
- 28.Ka SM, Huang XR, Lan HY. Smad7 gene therapy ameliorates an autoimmune crescentic glomerulonephritis in mice. J Am Soc Nephrol. 2007;18:1777–88. doi: 10.1681/ASN.2006080901. [DOI] [PubMed] [Google Scholar]
- 29.Lan HY, Mu W, Tomita N. Inhibition of renal fibrosis by gene transfer of inducible Smad7 using ultrasoundmicrobubble system in rat UUO model. J Am Soc Nephrol. 2003;14:1535–48. doi: 10.1097/01.asn.0000067632.04658.b8. [DOI] [PubMed] [Google Scholar]
- 30.WuDunn D. Mechanobiology of trabecular meshwork cells. Exp Eye Res. 2009;88:718–23. doi: 10.1016/j.exer.2008.11.008. [DOI] [PubMed] [Google Scholar]
- 31.Gasiorowski JZ, Russell P. Biological properties of trabecular meshwork cells. Exp Eye Res. 2009;88:671–5. doi: 10.1016/j.exer.2008.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schwartz MA, DeSimone DW. Cell adhesion receptors in mechanotransduction. Curr Opin Cell Biol. 2008;20:551–6. doi: 10.1016/j.ceb.2008.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Balaban NQ, Schwarz US, Riveline D. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat Cell Biol. 2001;3:466–72. doi: 10.1038/35074532. [DOI] [PubMed] [Google Scholar]
- 34.Discher DE, Janmey P, Wang YL. Tissue cells feel and respond to the stiffness of their substrate. Science. 2005;310:1139–43. doi: 10.1126/science.1116995. [DOI] [PubMed] [Google Scholar]