

Abstract
Pathogens and parasites can induce changes in host or vector behavior that enhance their transmission. In plant systems, such effects are largely restricted to vectors, because they are mobile and may exhibit preferences dependent upon plant host infection status. Here we report the first evidence that acquisition of a plant virus directly alters host selection behavior by its insect vector. We show that the aphid Rhopalosiphum padi, after acquiring Barley yellow dwarf virus (BYDV) during in vitro feeding, prefers noninfected wheat plants, while noninfective aphids also fed in vitro prefer BYDV-infected plants. This behavioral change should promote pathogen spread since noninfective vector preference for infected plants will promote acquisition, while infective vector preference for noninfected hosts will promote transmission. We propose the “Vector Manipulation Hypothesis” to explain the evolution of strategies in plant pathogens to enhance their spread to new hosts. Our findings have implications for disease and vector management.
Pathogenic and parasitic organisms interact with their hosts on a variety of cellular and organismal levels that potentially cause changes in host behavior leading to enhanced transmission1,2,3,4,5. This phenomenon led to the emergence of the “Host Manipulation Hypothesis” (HMH)6. The HMH and its synonyms the adaptive manipulation7 and behavioral manipulation8 hypotheses posit that natural selection on the parasite or pathogen has favored the capacity to elicit host behavior that enhances their transmission. Although examination of the HMH has progressed from descriptive studies to investigations of the mechanisms through which parasites affect host behavior and their consequences for parasite spread9,10, the field remains predominantly focused on animal pathosystems.
Pathogens or parasites can influence the behavior not only of their primary hosts, but also of their vectors. Arthropods are important vectors of both animal and plant pathogens, transmitting thousands of species of pathogens, including viruses, bacteria, phytoplasmas, trypanosomes and Plasmodia2,11. The effects of pathogens on vector biology and behavior have been documented in several pathosystems, including those associated with important human diseases such as malaria, leishmaniasis and sleeping sickness2,5. The observed changes in vector behavior include those related to pathogen transmission. For example, mosquitoes infected with the malaria parasite exhibit increased biting frequency and increased attraction to humans infected with the gametocytes of the parasite compared to noninfected humans1,3.
In contrast to animal pathosystems, plant pathosystems have been less well studied for evidence of host or vector manipulation by pathogens12. While animal pathogens can alter the behavior of both hosts and vectors in ways that increase frequency of host-host or host-vector encounters2,4,5, in plant pathosystems the host is sessile, so the potential for behavioral manipulation is restricted to the vector, the mobile component in these systems. Furthermore, unlike animal pathogens most plant pathogens, including the majority of plant viruses, do not replicate within the vector, so these vectors are not pathogen hosts, sensu stricto.
We previously demonstrated that Barley yellow dwarf virus (BYDV) infecting wheat and Potato leafroll virus (PLRV) infecting potato indirectly induce changes in the host selection behavior of their respective principal aphid vectors, Rhopalosiphum padi and Myzus persicae13,14,15,16. We also have shown that plants infected with these viruses have altered volatile organic compound profiles that elicit greater settling of or arrestment by their noninfective vectors13,14,16,17. Luteoviruses (viruses in the family Luteoviridae), including BYDV and PLRV are persistently transmitted. They are ingested and pass through the midgut or hindgut into the hemocoel, eventually associating with the accessory salivary glands of the vector18. These viruses rely almost exclusively on insect vectors for transmission and require sustained feeding by a vector for their successful acquisition and transmission19. After acquisition, the insect remains a vector for life. Although they do not replicate within the vector, persistently-transmitted viruses interact with the vector at the cellular level during movement among tissues and organs20, with the potential to directly alter vector physiology and behavior.
Preferential settling by vectors onto infected plants, as occurs for BYDV and PLRV, could contribute to enhanced pathogen spread. Models indicate that a preference for infected plants will accelerate pathogen spread, but only when infected plants are rare, not when they are prevalent in a plant population21. Conditional vector preference, however, could enhance pathogen spread regardless of the prevalence of infected plants. Specifically, if noninfective vectors prefer infected plants thereby promoting acquisition, and infective vectors prefer noninfected hosts promoting transmission, overall spread would be accelerated. The possibility of conditional vector preference for pathogen-infected plants has hardly been examined despite its potential importance. Changes in vector behavior that occur after feeding on virus-infected plants could be attributed to direct effects of the acquired virus on the vector, but such direct effects are difficult to distinguish from indirect ones associated with feeding on virus-infected plants. Here we test the hypothesis that a change in host plant selection behavior by an insect vector is the direct result of virus acquisition by the vector. We provide the first experimental evidence that acquisition of a plant virus through in vitro feeding, which eliminates indirect effects of an infected plant host, directly alters subsequent host plant selection behavior of its vector. These findings enhance our understanding of how plant viruses spread to new hosts, with implications for disease and vector management.
Results
We first examined host plant selection preferences of infective (reared on virus-infected plants) and noninfective (reared on virus-free plants) R. padi. In dual-choice bioassays using an arena in a platform22 (Fig. 1) infective or noninfective insects were allowed to select BYDV-infected or sham-inoculated wheat plants as their hosts. Sham-inoculated plants are noninfected plants previously fed upon by noninfective aphids and are utilized in our bioassays to account for potential aphid feeding-induced changes in plants23. Infective and noninfective insects were tested simultaneously in separate platforms. Each platform contained a leaf from each plant treatment, BYDV-infected or sham-inoculated, onto which aphids could settle and feed throughout the bioassay. We compared the responses of infective and noninfective aphids by examining the proportion of aphids that settled on BYDV-infected or sham-inoculated plants every 12 h for 72 h. A 72-h time period is sufficiently long for virus acquisition by noninfective aphids to occur when exposed to BYDV-infected plants, while a 12-h time period is unlikely to result in noninfective aphids becoming infective due to the latent period of the virus24,25. We therefore compared aphid responses at the first 12-h observation, and after 72 h when responses were pooled over time. The 12-h observation occurs before additional virus acquisition was expected while the 72-h comparison is more powerful statistically and incorporates a time period more meaningful for transmission dynamics in the field. Noninfective aphids significantly preferred to settle on BYDV-infected wheat compared to infective aphids at the first 12-h observation point (generalized linear model; χ2 = 3.12, p = 0.0774, marginally significant) (Fig. 2a, Supplementary Table S1a) and throughout the duration of the experiment (generalized linear model; χ2 = 19.33, p < 0.0001) (Fig. 2b, Supplementary Table S2a). In contrast, infective aphids significantly preferred to settle on sham-inoculated wheat compared to noninfective aphids at the first observation point (generalized linear model; χ2 = 3.12, p = 0.0774, marginally significant) (Fig. 2a, Supplementary Table S1b) and throughout the duration of the experiment (generalized linear model; χ2 = 20.14, p < 0.0001) (Fig. 2b, Supplementary Table S2b). The time at which the observations were made was not a significant factor when examining the response to BYDV-infected wheat (generalized linear model; χ2 = 4.96, p = 0.4203) (Supplementary Table S2a) or sham-inoculated wheat (generalized linear model; χ2 = 2.15, p = 0.8282) (Supplementary Table S2b). The results suggest that virus acquisition changes vector host plant selection behavior to favor noninfected plants rather than infected plants.
Figure 1. Diagrammatic illustration of the dual-choice bioassay arena used in experiments.
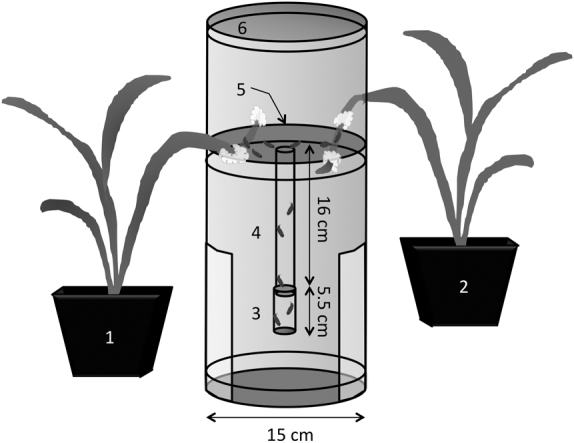
Adapted from Castle et al.22. 1, BYDV-infected wheat; 2, sham-inoculated wheat; 3, vial (5.5 x2.5 cm; Lx D) initially containing 50 aphids; 4, tube (16x2.5 cm; LxD); 5, platform (15 cm; D); 6, lid enclosing the arena.
Figure 2. Mean proportion of infective and noninfective aphids responding in a dual-choice bioassay examining host plant selection preferences to BYDV-infected and sham-inoculated wheat (noninfected plants previously fed upon by noninfective aphids) as influenced by indirect effects of feeding on virus-infected plants.
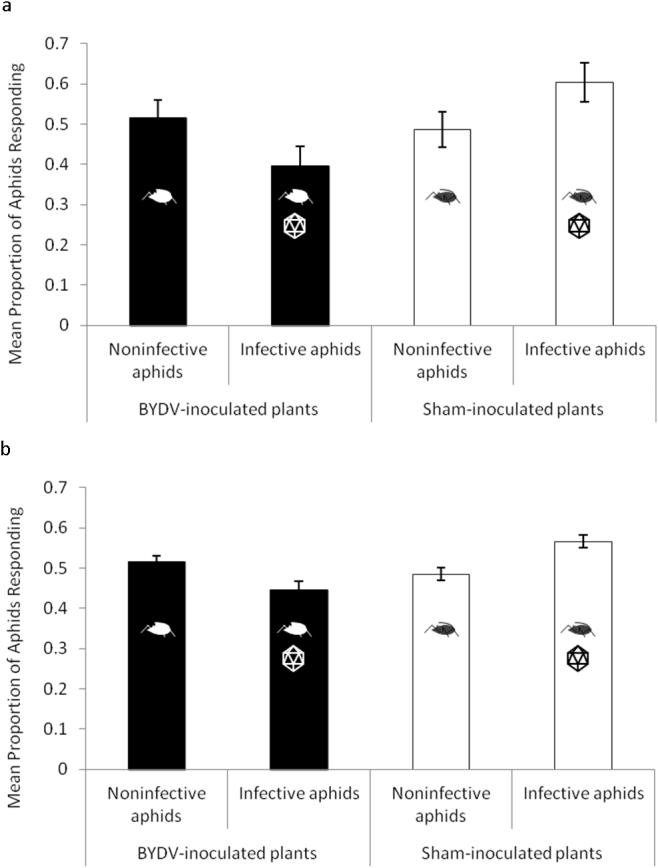
Each replicate (n = 12) consisted of one arena with noninfective aphids paired with one arena of infective aphids, randomized in a complete block design over time. Statistical analyses compared the response of infective and noninfective aphids to the BYDV-infected or sham-inoculated plant treatment. (a) Aphid responses at the first observation point made 12 h after release. Noninfective aphids preferred BYDV-infected wheat compared to infective aphids (generalized linear model; χ2 = 3.12, p = 0.0774, marginally significant). Infective aphids preferred sham-inoculated plants compared to noninfective aphids (generalized linear model; χ2 = 3.12, p = 0.0774, marginally significant). (b) Aphid responses pooled over time (6 observations). Noninfective aphids significantly preferred BYDV-infected wheat compared to infective aphids (generalized linear model; χ2 = 19.33, p < 0.0001). Infective aphids significantly preferred sham-inoculated plants compared to noninfective aphids (generalized linear model; χ2 = 20.14, p < 0.0001). Data are means ± SE following logit transformation. Errors bars are s.e.m.
These behavioral changes could result either from direct effects of acquired virus particles on the aphid, or from insect exposure to cues from virus-infected host plants. To isolate potential direct effects of virus acquisition on the vector we conducted a similar experiment using in vitro feeding to obtain infective and noninfective aphids. Insects were first reared on virus-free plants and subsequently transferred to membrane feeding chambers26 (Fig. 3) that contained artificial phloem with either purified BYDV particles or no virus. Host plant selection preferences of infective and noninfective insects were examined every 12 h for 72 h using an arena as described above. Observation time was not a significant factor when examining the response to BYDV-infected wheat (generalized linear model; χ2 = 2.41, p = 0.7906) (Supplementary Table S2c) or sham-inoculated wheat (generalized linear model; χ2 = 3.66, p = 0.5995) (Supplementary Table S2d). We present the results of the aphid responses at the first 12-h observation point as well as the responses pooled over time. Noninfective aphids significantly preferred BYDV-infected wheat compared to infective aphids at the first observation point (generalized linear model; χ2 = 4.24, p = 0.0394) (Fig. 4a, Supplementary Table S1c), and throughout the duration of the experiment (generalized linear model; χ2 = 16.18, p < 0.0001) (Fig. 4b, Supplementary Table S2c). Similar to the patterns obtained using aphids that acquired virus from plants, infective aphids significantly preferred sham-inoculated wheat compared to noninfective aphids at the first observation point (generalized linear model; χ2 = 5.64, p = 0.0176) (Fig. 4a, Supplementary Table S1d), and throughout the duration of the experiment (generalized linear model; χ2 = 16.32, p < 0.0001) (Fig. 4b, Supplementary Table S2d).
Figure 3. Diagrammatic illustration of a membrane feeding chamber.
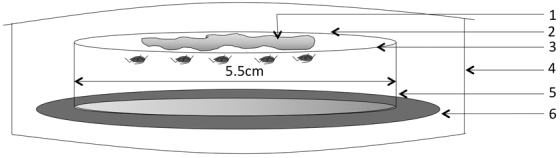
1, artificial diet solution (100 µL); 2, upper layer of Parafilm®; 3, bottom layer of Parafilm®; 4, humid chamber; 5, petri dish (5.5 cm; D); 6, moist filter paper.
Figure 4. Mean proportion of infective and noninfective aphids responding in a dual-choice bioassay examining host plant selection preferences to BYDV-infected and sham-inoculated wheat plants as influenced by direct effects of virus acquisition following membrane feeding.
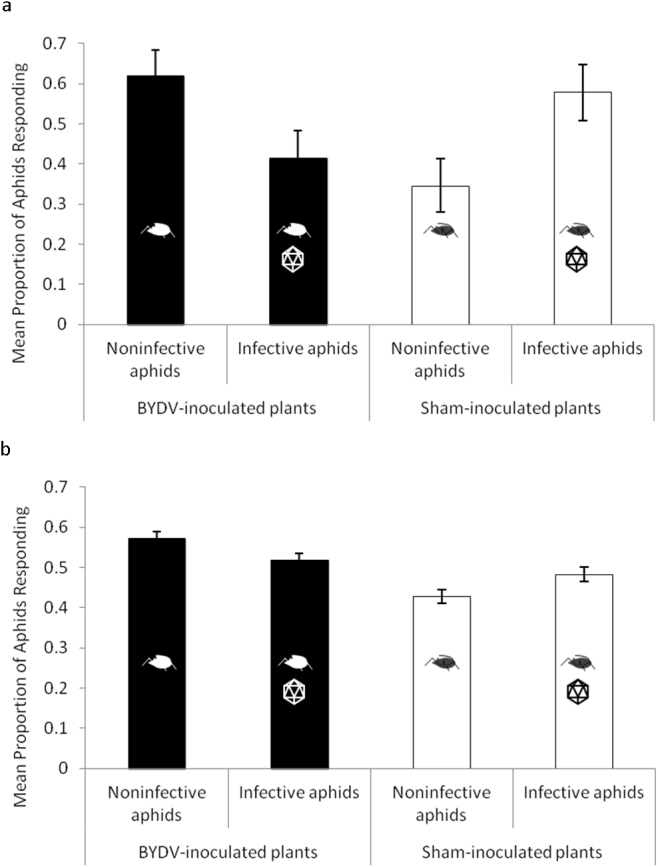
Each replicate (n = 12) consisted of one arena with noninfective aphids paired with one arena of infective aphids, randomized in a complete block design over time. Statistical analyses compared the response of infective and noninfective aphids to the BYDV-infected or sham-inoculated plant treatment. (a) Aphid responses at the first observation point made 12 h after release. Noninfective aphids significantly preferred BYDV-infected wheat compared to infective aphids (generalized linear model; χ2 = 4.24, p = 0.0394). Infective aphids significantly preferred sham-inoculated wheat compared to noninfective aphids (generalized linear model; χ2 = 5.64, p = 0.0176). (b) Aphid responses pooled over time (6 observations). Noninfective aphids significantly preferred BYDV-infected wheat compared to infective aphids (generalized linear model; χ2 = 16.18, p < 0.0001). Infective aphids significantly preferred sham-inoculated wheat compared to noninfective aphids (generalized linear model; χ2 = 16.32, p < 0.0001). Data are means ± SE following logit transformation. Errors bars are s.e.m.
Results from RT-PCR tests verified that our inoculation and acquisition methods were successful (See Supplementary Figure S1). All plants used in the dual choice tests were tested via RT-PCR immediately after the bioassays. Sham-inoculated plants remained virus-free and infected plants remained BYDV-infected, indicating that during the bioassays (72 h) the plant treatments were stable, despite being exposed to potential feeding by infective aphids. Tests of aphids using RT-PCR revealed that infective aphids remained BYDV-infective subsequent to the bioassay, while 25% of noninfective aphids acquired BYDV during the 72-h bioassay when they had access to BYDV-infected plants in the bioassay arena. Although the bioassay design unavoidably results in virus acquisition by some noninfective aphids, the result is a more conservative test of our hypothesis since within-bioassay virus acquisition should tend to diminish detectable differences between the aphid treatments. Furthermore, the aphid responses after 72 h in the bioassay arena are consistent with the preferences observed after 12 h, during which time noninfective aphids almost certainly remained noninfective24,25. The lack of BYDV infection of the sham-inoculated plants after 72 h of exposure to initially noninfective aphids in an arena with BYDV-infected plants also indicates that these aphids did not become infective during the bioassay.
Discussion
Assays utilizing membrane-fed infective aphids yielded results similar to those obtained using aphids that acquired BYDV from infected plants, confirming our hypothesis that changes in host plant selection by the vector are mediated by direct effects of virus acquisition, rather than indirect effects of feeding on infected host plants. Direct effects of virus acquisition on the vector host plant selection behavior in a manner that will promote the spread of the virus is consistent with an evolved strategy in the pathogen of manipulation of its vector. We propose the “Vector Manipulation Hypothesis” (VMH) to explain the evolution of strategies in plant pathogens that enhance their spread to new hosts through their effects on mobile vectors. Selection should favor both direct and indirect mechanisms producing such effects. Vectors that feed on virus-infected host plants exhibit faster growth rates, higher fecundity, greater longevity and/or enhanced production of alate forms of the vector27,28,29,30,31,32,33, which can lead to increased virus spread and are typically attributed to indirect effects of virus infection on host quality. Virus- infection-mediated alterations of the host plant's secondary chemistry can affect vector behavior. Evidence for such indirect effects of pathogens on vector behavior continues to accumulate and is consistent with the VMH13,14,15,16,34,35,36. We provide the first evidence for a direct effect of a plant virus on its vector consistent with the VMH, specifically by influencing the vector's host selection behavior to maximize pathogen spread. In our model pathosystem, noninfective vectors are attracted to virus-infected host plants, which is beneficial as it increases vector fitness23. After virus acquisition virus vector preferences shift to noninfected hosts, maximizing pathogen transmission potential by promoting the movement of infective aphids onto noninfected host plants. Our results offer a specific example of a plant virus directly manipulating its vector in a manner that is likely to maximize pathogen transmission potential between hosts, providing support for the VMH.
Results supportive of the VMH also have been reported from work on nonpersistently-transmitted plant viruses examining effects on noninfective vector behavior. Non-persistently transmitted viruses bind transiently to insect mouthparts20 and interactions in these pathosystems are likely limited to indirect effects on vectors. Recent work with the non-persistently transmitted Cucumber mosaic virus (CMV), which is acquired rapidly during aphid feeding and benefits from rapid vector dispersal, showed that aphids are initially attracted to volatile organic compounds from CMV-infected squash plants, but subsequently prefer to colonize noninfected plants34. Attraction to CMV-infected plants appears to be mediated by their increased emission of volatile organic compounds similar to those from healthy plants. Since CMV can be acquired within a few seconds by an aphid probing on an infected plant, these behaviors can act to enhance virus spread34 and illustrate manipulation of an insect vector by the virus. Interestingly, in addition to manipulating vectors, CMV also may manipulate defensive signaling pathways in plants that could result in enhanced vector survival37.
Our findings highlight the ecological and evolutionary significance of vector manipulation by pathogens and parasites. Effects like those we document for a plant virus, consistent with the VMH, may be widespread since direct and indirect mechanisms that enhance the spread of plant viruses should be favored by natural selection. Furthermore, similar patterns in behavioral changes among vectors of other plant pathogens, such as bacteria and phytoplasmas, which are limited to sessile plant hosts, might also occur. Although our results do not address the specific cellular and molecular mechanisms mediating direct plant virus effects on their vectors, they offer strong quantitative evidence for the VMH, providing a foundation upon which to base further studies of pathogen-mediated manipulation of their vectors and the identification of underlying mechanisms. The evolution of host-vector interactions has recently been suggested to be in part, mediated by virus transmission mechanisms38 underlying the importance of studying such interactions. Greater understanding of host plant-virus-vector interactions has the potential to improve management of vectors and plant diseases in agricultural settings and enhance our understanding of the role plant viruses play in natural settings39, including their effects on ecological processes at the community and ecosystem levels38.
Methods
Virus maintenance and insect rearing
The model system for our study was the wheat-R. padi-BYDV pathosystem. BYDV is exclusively transmitted among Poaceae hosts by aphids, including R. padi, in a persistent circulative manner and the virus does not replicate within the vector40,41. A Washington State isolate of BYDV-PAV maintained by mass transfer of R. padi, the bird-cherry oat aphid, on cv. Sprinter barley plants was used to inoculate wheat plants23. Rophalosiphum padi is the most efficient vector of the BYDV-PAV serotype40. Both the virus and an infective colony of R. padi are maintained at the University of Idaho (UI) Agricultural Biotechnology Laboratory. Aphids were originally obtained from Washington State University and are kept virus-infective through serial transfer23. A noninfective colony of R. padi was derived from the infective colony and is maintained at the UI Manis Entomological Laboratory. Infective and noninfective aphid colonies are reared on Sprinter barley in environmental chambers (20±2°C; 16 h light photoperiod). Aphids from each colony are examined on a regular basis using RT-PCR tests to ensure that the respective colonies remain virus-free or BYDV-PAV infected (see Supplementary Methods).
Plant rearing and inoculation
Winter wheat cv. Lambert was used for all bioassays. Seeds were planted at a density of one per pot in 10.2 cm2 plastic pots. Pots were filled with a mixture of 6:1:0.02 ratio of Sunshine mix #1: sand: Osmocote®, placed on trays in an environmental chamber (20±2°C; 16 h light photoperiod) and bottom watered. After germination, plants were fertilized using a soluble N-P-K fertilizer (15:30:15) biweekly.
Plant inoculations were done at the 2–3 leaf stage (14–16 days after planting). BYDV-infected plants were obtained by caging 10 adult aphids from the infective colony per plant for a 72 h virus inoculation access period23. Cages consisted of a 4-cm long piece of 23 mm dialysis tubing (14.6 mm D, Spectra/Por®) sealed on both ends with a foam stopper. Since BYDV is exclusively insect-transmitted, all BYDV-infected plants are fed-upon by aphids. Insect feeding may induce resistance in plants and potentially affect the response of insects subsequently exposed to such plants23. To account for such potential confounding effects sham-inoculated plants were produced and served as virus-free controls. Sham-inoculation was conducted by caging 10 adult aphids from the noninfective colony per plant for 72 h23.
Infective and noninfective aphid handling
To examine effects of plant virus acquisition from infected plants on host selection behavior, apterous aphids (fourth nymphal stage to early adults) originated from the respective infective and noninfective colonies. Previous research in this pathosystem has focused on aptera14,15,23,25. While alates are important vectors for long-distance dispersal events, apterous aphid behavior can be used to predict severity of epidemics within a field once the virus and vector are established21. Future studies will examine alate behavior in response to BYDV-infection. Aphids for each treatment were individually removed from plants using a number 3 camel's hair brush and placed into vials (2.3x5.5 cm; DxH). Fifty aphids were placed per vial. Vials were capped and aphids starved for one hour prior to the bioassay. A total of 600 aphids of each treatment (infective and noninfective) were tested among 12 replicates of the dual-choice bioassay described below.
Aphids for the experiment to assess the direct effects of virus acquisition originated from the noninfective R. padi colony. Tests were conducted using membrane feeding chambers modified after Trębicki et al.26 (Fig. 3), containing artificial diet as described by Ramsey and Jander42. After preparation the diet solution was sieved using a bacteria-proof filter (0.2 µm cellulose acetate). To set up membrane feeding chambers, the bottom halves of glass petri dishes (5.5 cm; D) were first sterilized under UV light for 10 min. Aphids (fourth nymphal stage to early adults) were collected from colony plants using a number 3 camel's hair brush and placed in the petri dishes, 35 aphids per dish. Parafilm®, sterilized with 70% ethanol, was used as membrane material. After placing the aphids in the petri dish, the dish was immediately sealed with a layer of Parafilm® stretched tightly across the dish top. After all dishes were sealed with the first Parafilm® layer 100 µL of artificial diet was pipetted onto the membrane and a second layer of Parafilm® was stretched tightly to sandwich the diet26. The diet was then spread across the surface of the membrane by applying pressure to the top layer with a fingertip. Dishes were placed in a tray with moistened filter paper and the tray covered with cling wrap and placed inside an environmental chamber (20±2°C; 16 h light photoperiod) for 24 h. Non-infective aphids were fed on an amino acid and sucrose diet solution. To obtain infective aphids, insects were fed on the same diet solution that was infused with purified BYDV at a concentration of 100 µg/mL. Virus was purified following a method adapted from Hammond et al.43, and obtained from Dr. Alex Karasev, UI PSES Department. After a 24-h feeding period, aphids were transferred into a vial as described above, starved for one hour and released in a bioassay arena (see below). A total of 840 aphids of each treatment were placed in membrane chambers, 600 were tested among 12 replicates of the dual-choice bioassay described below. The remaining aphids were stored in 70% ethanol at −20°C to verify their status (infective vs. noninfective). Virus-infection status of plants and aphids was determined using RT-PCR (See Supplementary Methods and Supplementary Figures S1–S2).
We recognize that purified virus may contain phloem proteins. Such proteins have been reported to occur in vivo, and were recently reported to play a role in virus transmission44. Additional studies are required to determine if a virus-plant protein complex is present in vitro and if such a complex could contribute to behavioral changes in vectors.
Bioassays to assess aphid preferences
Dual-choice bioassays were performed 40–46 days after plant inoculation, utilizing an arena adapted from Castle et al.22 (Fig. 1). The base of the arena was glued into the lid portion of a 15 cm D petri dish. The platform of the arena consisted of the inverted bottom of the petri dish with a 2.5 cm D hole cut in the center. A clear plastic tube (16x2.5 cm; LxD) was inserted into the bottom of the dish and secured with glue. The arena was wrapped in a heavy weight mylar frame (30.5x46.1 cm; WxL) to add stability to the structure. Holes were cut in the mylar, four (2 cm; D) equally spaced around the top and two (8x8 cm2) in the bottom to access the arena. One leaf still attached to the plant from each treatment (BYDV-infected and sham-inoculated) was inserted through holes on either side of the arena and held in place with a cotton seal. A vial (5.5x2.5 cm; LxD) containing 50 aphids, starved for one hour, was inserted into the bottom of the plastic tube leading to the arena. Apterous infective and noninfective aphids were released simultaneously into separate arenas. Aphids crawled up the tube and emerged onto a platform with one leaf from each treatment on either side (3 cm on either side of where aphids entered the arena). Aphids were able to settle on, feed and move between the two leaves. Aphids were released at the start of a dark period and monitored every 12 h (alternating dark and light times) for a 72-h period. The number of aphids on each leaf was counted at each observation, using a red light when monitoring during the dark cycles14. Assays were conducted in a growth room (14±3°C; 12 h light photoperiod). One replicate consisted of an arena containing infective aphids paired with another arena containing noninfective aphids, constituting a single block. Twelve replicates were performed across time in a randomized complete block repeated measures design.
Data analysis
The proportion of aphids responding to either the BYDV-infected or sham-inoculated plant treatment was compared using a generalized linear model assuming a binomial distribution and logit transformation (SAS, Proc Genmod). Logit transformation was performed to stabilize the variance and meet the assumptions of normality for analysis. Aphids not located on either plant leaf in an arena were considered non responsive and excluded from the analysis. The partial model examined the main effects of replicate (block; n = 12) and aphid treatment (infective or noninfective). The analysis was conducted separately four times, once for each plant treatment (BYDV-infected or sham-inoculated) for the indirect effects experiment (aphids reared on noninfected plants or virus-infected plants) (Supplemental Table S1a-b) and the direct effects experiment (aphids fed on membrane chambers with or without virus) (Supplemental Table S1c-d). The full model examined the main effects of replicate, aphid treatment and time (n = 6) assuming a compound symmetric correlation. The time variable examined observations made at 12, 24, 36, 48, 60 and 72 h after release using a repeated measures design. Observations made at 12, 36, and 60 h were recorded in the dark. Light and dark observations were examined with the model separately and no significant interactions were observed, thus results were pooled in the overall analysis (Supplemental Table S2). All statistical tests (likelihood ratio χ2) were carried out at the alpha = 0.05 level of significance.
Author Contributions
N.B.P., L.L.I., and S.D.E. conceived and designed research; L.L.I. performed research and analyzed data; L.L.I., N.B.P., and S.D.E. interpreted results and wrote the paper.
Supplementary Material
Plant Viruses Alter Insect Behavior to Enhance their Spread
Acknowledgments
Our research was supported by USDA-AFRI award 2009-6510405730. We thank A. Busch, S. Eid, E. Kmieciak, J. Knerr, A. Poplawsky, P. Trębicki, and L. Unger for technical assistance, A. Karasev for providing purified BYDV, and B. Price for input on statistical analysis.
References
- Koella J. C., Sorensen F. L. & Anderson R. A. The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 265, 763–768 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurd H. Manipulation of medically important insect vectors by their parasites. Annu. Rev. Entomol. 48, 141–161 (2003). [DOI] [PubMed] [Google Scholar]
- Lacroix R., Mukabana W. R., Gouagna L. C. & Koella J. C. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol 3, e298 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vyas A., Kim S. K., Giacomini N., Boothroyd J. C. & Sapolsky R. M. Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc. Natl. Acad. Sci. U. S. A. 104, 6442–6447 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lefèvre T. & Thomas F. Behind the scene, something else is pulling the strings: Emphasizing parasitic manipulation in vector-borne diseases. Infect., Genet. Evol. 8, 504–519 (2008). [DOI] [PubMed] [Google Scholar]
- Holmes J. C. & Bethel W. M. in Behavioral Aspects of Parasite Transmission (eds Canning E. U., & Wright C. A.) 123–149.– (Academic Press, 1972). [Google Scholar]
- Poulin R. Manipulation of host behaviour by parasites: a weakening paradigm? Proc. R. Soc. Lond. Ser. B. Biol. Sci. 267, 787–792 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas F., Adamo S. & Moore J. Parasitic manipulation: where are we and where should we go? Behav. Processes 68, 185–199 (2005). [DOI] [PubMed] [Google Scholar]
- Poulin R. “Adaptive” changes in the behaviour of parasitized animals: A critical review. Int. J. Parasitol. 25, 1371–1383 (1995). [DOI] [PubMed] [Google Scholar]
- Moore J. & Gotelli N. J. in Parasitism and Host Behaviour (eds Barnard C. J., & Behnke J. M.) 193–233 (Taylor & Francis Ltd., 1990). [Google Scholar]
- Gray S. M. & Banerjee N. Mechanisms of arthropod transmission of plant and animal viruses. Microbiol. Mol. Biol. Rev. 63, 128–148 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosque-Pérez N. A. & Eigenbrode S. The influence of virus-induced changes in plants on aphid vectors: Insights from luteovirus pathosystems. Virus Res 159, 201–205 (2011). [DOI] [PubMed] [Google Scholar]
- Eigenbrode S. D., Ding H., Shiel P. & Berger P. H. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae). Proc. R. Soc. Lond. Ser. B. Biol. Sci. 269, 455–460 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiménez-Martínez E. S. et al. Volatile cues influence the response of Rhopalosiphum padi (Homoptera: Aphididae) to Barley yellow dwarf virus-infected transgenic and untransformed wheat. Environ. Entomol. 33, 1207–1216 (2004). [Google Scholar]
- Medina Ortega K. J., Bosque-Pérez N. A., Ngumbi E., Jiménez-Martínez E. S. & Eigenbrode S. D. Rhopalosiphum padi (Hemiptera: Aphididae) responses to volatile cues from Barley yellow dwarf virus-infected wheat. Environ. Entomol. 38, 836–845 (2009). [DOI] [PubMed] [Google Scholar]
- Werner B. J., Mowry T. M., Bosque-Pérez N. A., Ding H. & Eigenbrode S. Changes in green peach aphid responses to potato leafroll virus-induced volatiles emitted during disease progression. Environ. Entomol. 38, 1429–1438 (2009). [DOI] [PubMed] [Google Scholar]
- Ngumbi E., Eigenbrode S. D., Bosque-Pérez N. A., Ding H. & Rodriguez A. Myzus persicae is arrested more by blends than by individual compounds elevated in headspace of PLRV-infected potato. J. Chem. Ecol. 33, 1733–1747 (2007). [DOI] [PubMed] [Google Scholar]
- Gildow F. Evidence for receptor-mediated endocytosis regulating luteovirus acquisition by aphids. Phytopathology 83, 270–277 (1993). [Google Scholar]
- Sylvester E. S. Circulative and propagative virus transmission by aphids. Annu. Rev. Entomol. 25, 257–286 (1980). [Google Scholar]
- Nault L. R. Arthropod transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 90, 521–541 (1997). [Google Scholar]
- McElhany P., Real L. A. & Power A. G. Vector preference and disease dynamics: A study of Barley yellow dwarf virus. Ecology 76, 444–457 (1995). [Google Scholar]
- Castle S. J., Mowry T. M. & Berger P. H. Differential settling by Myzus persicae (Homoptera: Aphididae) on various virus infected host plants. Ann. Entomol. Soc. Am. 91, 661–667 (1998). [Google Scholar]
- Jiménez-Martínez E. S., Bosque-Pérez N. A., Berger P. H. & Zemetra R. S. Life history of the bird cherry-oat aphid, Rhopalosiphum padi (Homoptera: Aphididae) on transgenic and untransformed wheat challenged with Barley yellow dwarf virus. J. Econ. Entomol. 97, 203–212 (2004). [DOI] [PubMed] [Google Scholar]
- Gray S. M., Power A. G., Smith D. M., Seaman A. J. & Altman N. S. Aphid transmission of Barley yellow dwarf virus: acquisition access periods and virus concentration requirements. Phytopathology 81, 539–545 (1991). [Google Scholar]
- Jiménez- Martínez E. S. & Bosque-Pérez N. A. Variation in Barley yellow dwarf virus transmission efficiency by Rhopalosiphum padi (Homoptera: Aphididae) after acquisition from transgenic and non-transformed wheat genotypes. J. Econ. Entomol. 97, 1790–1796 (2004). [PubMed] [Google Scholar]
- Trębicki P., Harding R. M. & Powell K. S. Anti-metabolic effects of Galanthus nivalis agglutinin and wheat germ agglutinin on nymphal stages of the common brown leafhopper using a novel artificial diet system. Entomol. Exp. Appl. 131, 99–105 (2009). [Google Scholar]
- Kennedy J. S. Benefits to aphids from feeding on galled and virus-infected leaves. Nature 168, 825–826 (1951). [DOI] [PubMed] [Google Scholar]
- Baker P. F. Aphid behavior on healthy and on yellow virus infected sugar beet. Ann. Appl. Biol. 48, 384–391 (1960). [Google Scholar]
- Gildow F. E. Increased production of alatae by aphids reared on oats infected with Barley yellow dwarf virus. Ann. Entomol. Soc. Am. 73, 343–347 (1980). [Google Scholar]
- Hodgson C. J. Effects of infection with the cabbage black ringspot strain of turnip mosaic virus on turnip as a host to Myzus persicae and Brevicoryne brassicae. Ann. Appl. Biol. 98, 1–14 (1981). [Google Scholar]
- Ajayi O. & Dewar A. M. The effect of Barley yellow dwarf virus on field populations of the cereal aphids, Sitobion avenae and Metopolophium dirhodum. Ann. Appl. Biol. 103, 1–11 (1983). [Google Scholar]
- Araya J. E. & Foster J. E. Laboratory study on the effects of Barley yellow dwarf virus on the life cycle of Rhopalosiphum padi (L.). J. Plant Dis. Protect. 94, 195–198 (1987). [Google Scholar]
- Fereres A. & Moreno A. Behavioural aspects influencing plant virus transmission by homopteran insects. Virus Res. 141, 158–168 (2009). [DOI] [PubMed] [Google Scholar]
- Mauck K. E., De Moraes C. M. & Mescher M. C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. U. S. A. 107, 3600–3605 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann R. S. et al. Induced release of a plant-defense volatile 'deceptively' attracts insect vectors to plants infected with a bacterial pathogen. PLoS Path. 8, e1002610 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stafford C. A., Walker G. P. & Ullman D. E. Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. U. S. A. 108, 9350–9355 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziebell H. et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant-aphid interactions in tobacco. Sci. Rep. 1, 187; 10.1038/srep00187 (2011) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauck K. E., Bosque-Pérez N. A., Eigenbrode S., De Moraes C. M. & Mescher M. C. Transmission mechanisms shape pathogen effects on host-vector interactions: evidence from plant viruses. Funct. Ecol. 10.1111/j.1365–2435.2012.02026.x. (2012). [Google Scholar]
- Malmstrom C., Melcher U. & Bosque-Pérez N. A. The expanding field of plant virus ecology: Historical foundations, knowledge gaps, and research directions. Virus Res. 159, 84–94 (2011). [DOI] [PubMed] [Google Scholar]
- Gildow F. E., Frank J., Bingham D. & Powell C. Barley yellow dwarf viruses in small grains in Pennsylvania: isolate identification, distribution, and vector efficiency. Plant Dis. 71, 922–926 (1987). [Google Scholar]
- Power A. G. Virus spread and vector dynamics in genetically diverse plant populations. Ecology 72, 232–241 (1991). [Google Scholar]
- Ramsey J. S. & Jander G. Testing nicotine tolerance in aphids using an artificial diet experiment. JoVE, e701 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond J., Lister R. M. & Foster J. E. Purification, identity and some properties of an isolate of Barley yellow dwarf virus from Indiana. J. Gen. Virol. 64, 667–676 (1983). [Google Scholar]
- Bencharki B. et al. Phloem protein partners of Cucurbit aphid borne yellows virus: Possible involvement of phloem proteins in virus transmission by aphids. Mol. Plant. Microbe Interact. 23, 799–810 (2010). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Plant Viruses Alter Insect Behavior to Enhance their Spread