

Abstract
Mild exercise activates hippocampal neurons through the glutamatergic pathway and also promotes adult hippocampal neurogenesis (AHN). We hypothesized that such exercise could enhance local androgen synthesis and cause AHN because hippocampal steroid synthesis is facilitated by activated neurons via N-methyl-D-aspartate receptors. Here we addressed this question using a mild-intense treadmill running model that has been shown to be a potent AHN stimulator. A mass-spectrometric analysis demonstrated that hippocampal dihydrotestosterone increased significantly, whereas testosterone levels did not increase significantly after 2 wk of treadmill running in both orchidectomized (ORX) and sham castrated (Sham) male rats. Furthermore, analysis of mRNA expression for the two isoforms of 5α-reductases (srd5a1, srd5a2) and for androgen receptor (AR) revealed that both increased in the hippocampus after exercise, even in ORX rats. All rats were injected twice with 5′-bromo-2′deoxyuridine (50 mg/kg body weight, i.p.) on the day before training. Mild exercise significantly increased AHN in both ORX and Sham rats. Moreover, the increase of doublecortin or 5′-bromo-2′deoxyuridine/NeuN-positive cells in ORX rats was blocked by s.c. flutamide, an AR antagonist. It was also found that application of an estrogen receptor antagonist, tamoxifen, did not suppress exercise-induced AHN. These results support the hypothesis that, in male animals, mild exercise enhances hippocampal synthesis of dihydrotestosterone and increases AHN via androgenenic mediation.
Keywords: neurosteroid, exercise-induced neurogenesis, low intensity exercise, de novo synthesis, paracrine
Gonadal hormones enhance neuronal plasticity, including adult hippocampal neurogenesis (AHN), which occurs in the dentate gyrus throughout life in mammals (1, 2)—as well as having neuroprotective effects (3–6). In particular, the hippocampus is known to be a target of androgen actions (5, 7, 8). Testosterone (T) and dihydrotestosterone (DHT) exert neuroprotective effects through androgen receptors (ARs) in the hippocampus (5, 7, 9), although T can also be converted to estradiol (E2) via aromatase in the brain (10, 11). Intriguingly, current studies revealed that androgens can be synthesized in the hippocampus (10, 12, 13), in contrast to the widespread notion that the testes provide androgens to the brain via the circulatory system. The possibility of local androgen synthesis should be supported by the fact that immunoreactivity for both 5α-reductase 1 and 2 is observed in the dentate granule cell layer of the hippocampus (14). If locally produced by a physiological stimulus such as exercise, then formation of DHT, with its high affinity for ARs, could have beneficial effects on neuronal plasticity, including AHN.
Indeed, we have demonstrated that hippocampal steroid synthesis is facilitated via N-methyl-D-aspartate receptors (15). Furthermore, mild exercise increases regional cerebral blood flow, an index of neuronal activity, through the glutamatergic pathway, and continuous mild exercise enhances AHN (16, 17). These results led us postulate that exercise would enhance androgen synthesis, which may in turn mediate exercise-induced AHN, even though the possible role of androgens in hippocampus plasticity has not been extensively studied, particularly in relation to the effects of exercise.
The mechanism of exercise-induced AHN is still unknown, although some of the beneficial effects of exercise on cognition and mood may be mediated by exercised-induced AHN (18). The participation of several trophic factors such as brain-derived neurotrophic factor (BDNF) and vascular endothelial growth factor (VEGF) has been proposed (18), and their possible androgenic mediation has been reported in adult canaries (19). The dentate gyrus, which expresses ARs (20, 21), is also a target of androgens, and one study has shown that androgens enhance AHN via increased cell survival in an androgen-dependent manner (9). Taking these findings into consideration, androgens may play a significant role in the regulation of exercise-induced AHN.
Furthermore, regarding exercise conditions that activate AHN, much research has demonstrated that both wheel (22) and treadmill (23, 24) running enhance AHN (SI Text, Mild-Intense Treadmill Running Model and Exercise-induced AHN). We used treadmill running, which can be controlled for speed, and developed a mild-intensity treadmill exercise model below the lactate threshold (LT, about 20 m/min) with less blood lactate accumulation and minimal stress response; the LT is a physiologic index of moderate exercise intensity established in rats (25, 26) as in humans (27). We have also shown that hippocampal neuronal activity and neurogenesis is induced by the mild-intensity exercise (16, 17, 28).
Here we address the possibility that hippocampal androgen formation is increased by mild exercise under the LT, which may, in turn, mediate the exercise-induced AHN through ARs. For this purpose, we applied derivatization methods to improve the sensitivity of liquid chromatography with tandem mass spectrometry (LC-MS/MS) (12). We confirmed that mild exercise training for 2 wk below the LT enhances AHN whereas AHN is not enhanced above the LT (SI Text, Mild-Intense Treadmill Running Model and Exercise-induced AHN and Fig. S1]. We thus examined the effects of 2 wk of mild exercise on androgen levels in hippocampus and AHN in male rats.
Results
Experiment I: Levels of DHT, T, and E2 in the Hippocampus and Plasma After Exercise.
On the basis of the LT, the rats were subjected to mild treadmill exercise at a running speed of 13.5 m/min, five times per week for 30 min at a time (Fig. 1A). We measured postexercise changes in the hippocampal gonadal hormones using HPLC and LC-MS/MS (Fig. 1A). Table 1 shows the levels of androgens and estradiol in the hippocampus and plasma after treadmill running. Mass-spectrometric analysis of DHT, T, and E2 levels demonstrated that treadmill running significantly increased hippocampal DHT levels in both the sham castrated (Sham) and orchidectomized (ORX) groups (effect of treadmill exercise: F(1,33) = 5.06, P < 0.05; effect of ORX: F(1,33) = 110.7, P < 0.0001; interaction of treadmill exercise and ORX: F(1,33) = 3.18, P = 0.08) without any changes in T levels. The levels of androgens in hippocampus and plasma after ORX are lower than Sham, with the decrease after ORX being more pronounced in plasma than in the hippocampus, so that the ratios of hippocampus to plasma DHT concentrations are as follows: Sham-sedentary, 23:1; Sham-exercise, 32.3:1; ORX-sedentary, 71.4:1; ORX-exercise, 220:1. We also investigated a possible contribution of estrogens. However, hippocampal E2 levels were unchanged with exercise. Note that plasma levels of DHT and T were also unchanged with exercise in sham rats and were extremely low after ORX.
Fig. 1.

Experimental design. To exclude acute effects of treadmill running, sample collection was performed 2 d after the last training session. (A and C) ORX or Sham was performed 1 wk before exercise training, at which time the rats were 12 wk old. (B and C) On the day before the 2-wk training began, all of the animals were injected twice i.p. (at 8:00 AM and 8:00 PM) with BrdU (50 mg/kg, B.W.) as a short cell-survival marker in this study. Also, rats were administered the flutamide (30 mg/kg, B.W, s.c.) or the estrogen receptor antagonist tamoxifen (1 mg/kg, B.W., s.c.) suspended in sesame oil 2 h before every exercise session. Rats in the control group were injected with sesame oil only. (D) The enzyme 5α-reductases (Srd5a1,2) convert T into DHT. The effects of T and DHT caused by binding to the AR. T is also converted to estradiol (E2) by the enzyme aromatase (P450arom) and the effect of E2 caused by binding to the estrogen receptors (ERα, β). (E) We have divided adult hippocampal neurogenesis into three primary phases: proliferation, differentiation, and survival of cells. Sham: sham castrated; ORX: orchidectomized; flutamide: an androgen receptor antagonist; tamoxifen: an estrogen receptor antagonist.
Table 1.
Effects of treadmill running on hippocampal or plasma gonadal hormone levels
| Sham |
ORX |
|||
| Sedentary | Exercise | Sedentary | Exercise | |
| Hippocampus | ||||
| DHT (nM) | 2.30 ± 0.3 | 3.24 ± 0.2* | 0.10 ± 0.01 | 0.22 ± 0.03* |
| DHT (ng/g wet weight) | 0.67 ± 0.1 | 0.94 ± 0.1* | 0.03 ± 0.002 | 0.06 ± 0.01* |
| Testosterone (nM) | 20.5 ± 2.2 | 19.5 ± 1.6 | 0.77 ± 0.05 | 0.90 ± 0.07 |
| Testosterone (ng/g wet weight) | 5.9 ± 0.7 | 5.6 ± 0.5 | 0.22 ± 0.02 | 0.26 ± 0.02 |
| Estradiol (nM) | 0.68 ± 0.1 | 0.88 ± 0.1 | 0.46 ± 0.05 | 0.33 ± 0.08 |
| Estradiol (ng/g wet weight) | 0.19 ± 0.03 | 0.22 ± 0.04 | 1.29 ± 0.15 | 0.92 ± 0.24 |
| Plasma | ||||
| DHT (nM) | 0.10 ± 0.04 | 0.10 ± 0.03 | 0.0014 ± 0.0005 | 0.0010 ± 0.0002 |
| DHT (pg/mL) | 29.62 ± 11.04 | 30.25 ± 8.32 | 0.40 ± 0.14 | 0.30 ± 0.05 |
| Testosterone (nM) | 5.11 ± 1.8 | 4.6 ± 1.2 | 0.00046 ± 0.0001 | 0.00069 ± 0.0002 |
| Testosterone (pg/mL) | 1472.5 ± 523.7 | 1326.3 ± 353.4 | 0.13 ± 0.03 | 0.20 ± 0.60 |
The steroid levels of Sham rats and ORX rats are shown. The steroids were separated into fractions of DHT, T, and E2 using a normal-phase HPLC system with a silica gel column, and then each steroid was measured using LC-MS/MS. After 2 wk of training, hippocampal DHT levels had increased in both Sham and ORX groups. There were no significant changes in hippocampal T and E2 levels or in plasma T and DHT levels. To compare the hippocampal level of steroids with plasma steroids, we converted to nanomolar concentration via the following estimation. First, 1 mL of plasma (93% is water) is assumed to have 1 g weight, as 1 mL of water is. Second, we assume that the hippocampal tissue having 1 g of wet weight has an approximate volume of 1 mL, as nearly 78% of the brain tissue consists of water (29). Data represent the mean ± SEM (n = 6–14 rats in each group).
*P < 0.05 in comparison with sedentary rats (two-way ANOVA). Sham: sham castrated; ORX: orchidectimized.
Experiment II: Expression of mRNA for Steroidogenic Enzymes and Steroid Receptors in the Hippocampus After Exercise.
We determined postexercise changes in the mRNA expression of steroid-related enzymes and receptors in the hippocampus using a real-time quantitative PCR (Fig. 1A). For the sequences of the oligonucleotide, see Table S1. Fig. 1D shows steroid metabolism from testosterone. Both of the two isoforms of the enzyme 5α-reductases (srd5a1, srd5a2) convert T into DHT. The enzymatic activity of srd5a2 is higher than that of srd5a1 (30). Both synthase genes increased with mild exercise [srd5a1: effect of treadmill exercise—F(1,17) = 44.93, P < 0.0001 (Fig. 2A); srd5a2: effect of treadmill exercise—F(1,18) = 7.12, P < 0.05 (Fig. 2B)]. Regarding srd5a1, the increase with exercise was lower in the ORX than in the Sham group. There was a significant interaction between treadmill exercise and ORX [interaction: F(1,17) = 7.076, P < 0.05], suggesting that the exercise-induced srd5a1 mRNA expression was attenuated by ORX. The increase of aromatase (P450arom) mRNA expression with exercise was completely blocked by ORX [effect of treadmill exercise; F(1,18) = 1.63, P = 0.22; effect of ORX: F(1,18) = 0.47, P = 0.5; interaction of treadmill exercise and ORX: F(1,18) = 5.88, P < 0.05 (Fig. 2C)]. The Bonferroni post hoc test from two-way ANOVA showed that Sham-exercised rats had significantly higher mRNA expression of P450arom, the enzyme that converts T into E2, than those that were sedentary (P < 0.05). In both Sham and ORX groups, treadmill exercise enhanced mRNA expression of ARs [effect of treadmill exercise: F(1,18) = 9.9, P < 0.01 (Fig. 2D)]. Exercise also significantly up-regulated mRNA expression of ERα [effect of treadmill exercise: F(1,18) = 11.25, P < 0.01 (Fig. 2E), but not of ERβ (Fig. 2F)] in both groups. These data imply that mild exercise promotes hippocampal DHT synthesis independently of circulating androgens. Moreover, although the increase in P450arom thus depends on circulating gonadal hormones, the ERα mRNA increase does not.
Fig. 2.

The effect of treadmill running on hippocampal sex steroidogenesis-related enzymes and steroid receptor mRNA. The mRNA expressions were quantified by real-time PCR. (A, B, and D) Analysis data of mRNA expression for the two isoforms of 5α-reductases (srd5a1: P < 0.0001; srd5a2: P < 0.05), which convert T into DHT, and for ARs (P < 0.01) revealed that both increased in Sham and ORX rats. (C) P450arom, which converts T into E2, mRNA expression was increased in Sham rats only. (E and F) Treadmill exercise increased the mRNA expression of ERα (P < 0.05), but not ERβ, in both Sham and ORX groups. Expressions of mRNA were normalized by GAPDH housekeeping gene as an internal standard. Data represent the mean ± SEM (n = 5–6 rats in each group). *P < 0.05 and ***P < 0.0001 in comparison with sedentary rats (two-way ANOVA). Sham: sham castrated, ORX: orchidectomized.
Experiment III: Suppressive Effects of an AR Antagonist on Exercise-Induced AHN.
To investigate whether androgens have a role in exercise-induced AHN, rats received sesame oil or flutamide [an AR antagonist; 30 mg/kg body weight (BW), s.c.] during 2 wk of mild treadmill exercise (Fig. 1B). We measured the weight of seminal vesicules and prostate to check the effect of the antagonist. The seminal vesicular and prostate weights of flutamide-treated rats were significantly lower compared with those of vehicle rats (Table S2). We have already demonstrated that this exercise protocol is sufficient to increase AHN for mice (16) and rats (Fig. S1). Treadmill running is a forced exercise for rodents, but, when carefully and systematically done after a preconditioning, acclimatization regimen, it causes several beneficial effects in rodents. Indeed, a relatively mild-intensity exercise has already been shown to enhance AHN, with neurogenesis increased by 40–60% compared with control (23, 24), which is similar to our present study (54%).
We assessed the impact of exercise on three primary phases of neurogenesis (proliferation, differentiation, and survival of cells) in the hippocampal dentate gyrus (Fig. 1E). To estimate neuronal survival and maturation, all of the animals were given two (at 8:00 AM and 8:00 PM) i.p. injections of 5-bromo-2-deoxyuridine (BrdU, 50 mg/kg B.W.) on the day before training (Fig. 1B). To monitor cell proliferation and differentiation after 2 wk of training, proliferation was assessed using a specific marker (Ki67, a proliferation marker for neurons and other cell types), and early neuronal differentiation was measured by doublecortin (DCX) immunoreactivity (immature neuron marker) (Fig. 1E). Exercise increased the number of Ki67 immunoreactive cells compared with respective sedentary rats [effect of treadmill exercise: F(1,40) = 39.65, P < 0.0001 (Fig. 3A)], an effect that was unaltered with flutamide treatment. However, the exercise-induced increases of DCX-positive cells (Fig. 3B) and BrdU/NeuN (a neuron survival marker) double-positive cells (Fig. 3C) were blocked by flutamide [DCX: effect of treadmill exercise—F(1,38) = 13.52, P < 0.001; effect of antagonists—F(2,38) = 4.37, P < 0.05; interaction of treadmill exercise and antagonists—F(2,38) = 4.76, P < 0.05; BrdU: effect of treadmill exercise—F(1,40) = 10.95, P < 0.01; effect of antagonists—F(2,40) = 3.7, P < 0.05; interaction of treadmill exercise and antagonists—F(2,40) = 3.7, P < 0.05]. In contrast, the administration of the estrogen-receptor inhibitor tamoxifen (1 mg/kg B.W.) had no effect on the increased number of exercise-induced DCX-positive and BrdU/NeuN double-positive cells (Fig. 3).
Fig. 3.

Effects of sex steroid receptor antagonists on exercise-induced AHN of male rats. (A) Mild exercise significantly increased the number of Ki67+ cells (proliferation marker) in Veh-, Flu-, and Tam-treated rats (P < 0.0001). (B and C) Flu, but not Tam, blocked the increase of the number of DCX+ (immature neuron marker, P < 0.001) and BrdU+/NeuN+ (cell survival and mature neuron marker, P < 0.01) cells with mild exercise. In immunostaining, the Upper and Lower panels show the cells of exercise rats and sedentary rats. (Left) Veh rats. (Center) Tam-treated rats. (Right) Flu-treated rats. Data represent the mean ± SEM (n = 7–8 rats in each group). *P < 0.05, **P < 0.01 ***P < 0.0001 in comparison with sedentary rats (two-way ANOVA and Bonferroni post hoc tests). Veh: vehicle; Tam: tamoxifen; Flu: flutamide.
Experiment IV: Effects of Orchidectomy on Exercise-Induced AHN.
To determine whether testis-derived androgens have a role in exercise-induced AHN, rats were either sham-operated (Sham), orchidectomized (ORX), or ORX+flutamide (ORX+Flu) and divided into sedentary and exercise groups (Fig. 1C). The seminal vesicle and prostate weights of the ORX and ORX+Flu rats were significantly lower compared with those of vehicle rats (Table S2). As in sham rats, exercise increased the number of Ki67 immunoreactive cells in the ORX dentate gyrus compared with respective sedentary rats, but this effect was not blocked by flutamide [effect of treadmill exercise: F(1,39) = 39.83, P < 0.0001 (Fig. 4A)]. However, exercise-induced increases of DCX-positive cells (Fig. 4B) and BrdU/NeuN double-positive cells (Fig. 4C) were prevented by ORX+Flu, but not by ORX (DCX: effect of treadmill exercise—F(1,34) = 44.2, P < 0.0001; effect of treatments—F(2,34) = 13.95, P < 0.0001; interaction of treadmill exercise and treatments—F(2,34) = 13.95, P < 0.0001; BrdU: effect of treadmill exercise—F(1,33) = 17.66, P < 0.01; effect of treatments—F(2,33) = 3.32, P < 0.05; interaction of treadmill exercise and treatments—F(2,33) = 3.32, P < 0.05).
Fig. 4.
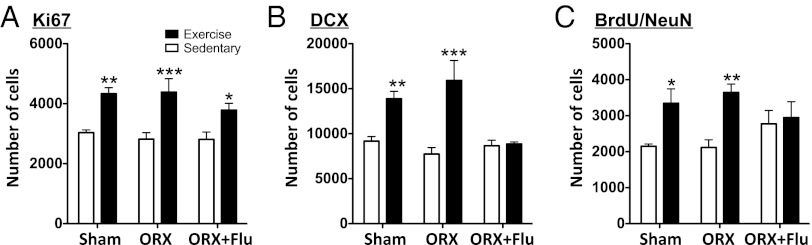
Effects of orchidectomy on exercise-induced AHN of male rats. (A) Mild exercise significantly increased the number of Ki67+ cells in Sham, ORX, and ORX+Flu rats (P < 0.0001). (B and C) Mild exercise increased the number of DCX+ and BrdU+/NeuN+ cells in ORX rats depleted of circulating androgens, and the effects were blocked by Flu. Data represent the mean ± SEM (n = 6–8 rats in each group). *P < 0.05, **P < 0.01, and ***P < 0.0001 in comparison with sedentary rats (two-way ANOVA and Bonferroni post hoc tests). Sham: sham castrated; ORX: orchidectomized; ORX+Flu: orchidectomized+flutamide.
Discussion
This report tests the hypothesis that hippocampal DHT synthesis is stimulated by mild exercise and contributes to exercise-induced AHN through the AR. We found that 2 wk of mild running resulted in increased hippocampal DHT levels, but not in those of T or E2, independently of circulating androgens. Furthermore, we demonstrated, using the AR antagonist, flutamide, that androgens stimulate exercise-induced AHN. In particular, orchidectomized rats with depleted circulating androgens still showed enhanced levels of AHN, which was completely blocked by flutamide, but not by tamoxifen, an estrogen receptor antagonist. These results provide evidence supporting the hypothesis that hippocampal androgen synthesis is stimulated by mild exercise and contributes to stimulation of AHN.
We demonstrated that hippocampal DHT levels and synthases (srd5a1,2) (Fig. 1D) are both increased with mild exercise and found this to be the case after ORX, which depleted circulating androgens. These results support the hypothesis that DHT could be synthesized in the hippocampus. This hypothesis was also supported by the fact that plasma androgens levels were unchanged with exercise with or without ORX. Furthermore, in the hippocampus, DHT is rapidly metabolized to 3α, 5α-androstanediol and inactivated by 3α-HSD (12). Therefore, the hippocampal DHT in the current study is very unlikely to have accumulated from peripheral sources, but was most likely synthesized de novo in the hippocampus.
The source of DHT should not always be restricted to the hippocampus, although dehydroepiandrosterone (DHEA) as adrenal androgen cannot be produced in rats due to their lack of adrenal Cyp17 (31), and both testosterone and DHT levels have been shown to be undetectable in the adrenals. It is possible that pregnenolone or progesterone, for example, coming from the adrenals could serve as substrates for hippocampal DHT. There is the possibility that DHEA produced in the brain mainly during early development but persisting in the adult brain (32) could be a source of androgen. Thus, hippocampal DHT could be synthesized from peripheral androgen precursors and/or synthesized de novo in the hippocampus.
As to the role of androgens in stimulating AHN, there are three principal stages of neurogenesis from progenitor cells in the hippocampus that can be distinguished using markers (33): proliferation, differentiation, and survival (Fig. 1E). Regarding differentiation and survival, we found that exercise stimulated expression of DCX, an early marker of neuronal differentiation, and that flutamide completely blocked exercise stimulation of DCX. We found the same for BrdU/NeuN, which, together, are markers of cell survival and neuronal maturation, indicating androgenic modulation of cell differentiation and survival in exercise-induced AHN. As one of the mechanisms, we considered Wnt/β-catenin signaling. Androgen receptors interact with β-catenin (34), which can enter the cell nucleus and then, via Wnt/β-catenin signaling, trigger the expression of NeuroD1 to promote neuronal differentiation in hippocampal neural progenitors (35). In the hippocampus, ARs thus may act via β-catenin in enhancing AHN. Future studies should address this mechanism.
We found that mild exercise increased Ki67, a proliferation marker, even in flutamide-administrated rats. The results are in agreement with a report by Spritzer and Galea showing that T and DHT, but not E2, enhanced cell survival without affecting cell proliferation in the dentate gyrus of adult male rats (9). The lack of a flutamide effect on Ki67 expression is likely explained by concurrent glial cell or progenitor cell proliferation that is androgen independent and might be stimulated by other mediators, such as IGF-I (36). Because Ki67 labels cells undergoing proliferation within 24 h (31), we gave no exercise for about 2 d (36 h) before euthanasia to exclude the acute effects of the last bout of exercise (37). Thus, the Ki67 results have no bearing on the effects of acute exercise, and we suggest that our results of increased Ki67-positive cells indicate a sustained effect of mild exercise training on enhancement of cell proliferation.
Importantly, we confirmed that the mild exercise significantly increases AHN even in rats with ORX that have depleted circulating androgens. Furthermore, with the administration of flutamide to ORX rats, the effects of exercise on DCX or BrdU/NeuN-positive cells were completely blocked. These results strongly suggest a paracrine/autocrine effect on AHN by hippocampus-synthesized DHT. Androgen might increase BDNF and VEGF (19), resulting in enhancement of neurogenesis. Collectively, these findings imply a stimulatory role for hippocampal DHT in the cascade of molecular mechanisms that underlie exercise-induced AHN.
It is important to consider possible indirect effects of AR on the hippocampus via afferent inputs because circulating testosterone may modulate hippocampal acetylcholine release (38), which is one of the factors that enhance AHN (39). Any contribution of acetylcholine neurons, however, may be excluded, because running-enhanced neurogenesis is unchanged in mice with partial cholinergic denervation (40) and in ORX rats where circulating androgens were eliminated (the current study). There does remain a possibility that hippocampal DHT could stimulate acetylcholine release from terminals in hippocampus via nongenomic AR (21).
Our finding that mild exercise increases hippocampal androgen levels, detected by a sensitive method, may be relevant for the prevention and slowing down of neurodegenerative diseases, such as Alzheimer’s disease, especially in people with a lower level of physical fitness (41). Because the injection of testosterone has been shown to reduce deposits of β-amyloid protein in vitro through the enhanced effects of neprilysin (42), it might be that mild exercise, in mediating paracrine effects through androgens and/or DHT, may, in turn, also reduce the deposits of β-amyloid protein and protect cognitive functions.
In conclusion, androgens, which modulate hippocampal synaptic plasticity (5), including spinogenesis (43) and enhance cognitive function (44), also modulate AHN stimulated by mild exercise below the lactate threshold. The current observations should encourage further research into hippocampal androgens as physiological mediators of neuronal plasticity as well as neuroprotection.
Materials and Methods
For a full description of all materials and methods, see SI Materials and Methods. Each experimental design is shown in Fig. 1.
Animals.
Eleven-week-old adult male Wistar rats (SEASCO Co.) were maintained on a 12-h light/dark schedule (light on at 7:00 AM) and given ad libitum access to food and water. All of the experimental protocols were performed in accordance with the University of Tsukuba Animal Experiment Committee guidelines. Animals were acclimatized to ambient rearing conditions for 7 d (two to three rats per cage) and then randomly assigned to the treadmill running or sedentary control groups.
Exercise Training Protocol.
Our previous studies revealed that the LT of rats is at a running speed of ∼20 m/min (25, 26). On the basis of the LT, the rats were subjected to mild treadmill exercise at a running speed of 13.5 m/min. We previously showed that hippocampal activation is induced by exercise intensities below the LT (17). The rats were habituated to the treadmill apparatus for 10 min (KN-73; Natsume) before training, and then they were subjected to 2 wk of training, five times per week for 30 min at a time. The training of exercised rats included gradual adaptation to the running. Sedentary rats remained sitting on the treadmill without running for the same amount of time. On the day before training, all of the animals were given two (at 8:00 AM and 8:00 PM) i.p. injections of BrdU (50 mg/kg B.W.) as a short cell-survival marker in this study.
Drug Administration.
Rats were s.c. administered with the androgen receptor antagonist flutamide (30 mg/kg B.W.) or the estrogen receptor antagonist tamoxifen (1 mg/kg B.W.) suspended in sesame oil 2 h before every exercise session. Rats in the control group were injected with sesame oil only. Because rats were subjected to treadmill exercise five times per week for 2 wk, the injections were given 10 times.
Sample Collection.
To exclude the acute effects of treadmill running, sample collection was performed 2 d after the last training session. In the experiment to assess adult hippocampal neurogenesis, the rats were deeply anesthetized with pentobarbital and transcardially perfused with 0.9% saline. Brains were carefully removed and fixed overnight at 4° C with 4% (wt/vol) paraformaldehyde in 0.1 M phosphate buffer and equilibrated in 30% (wt/vol) sucrose.
Mass-Spectrometric Assay of Steroids.
To examine how much androgen and estrogen were synthesized in the hippocampus after exercise, we performed a mass-spectrometric analysis. Detailed procedures are described elsewhere (12) and in the SI Materialsand Methods, Mass-Spectrometric Assay of Steroids. Hippocampi were homogenized for steroid extraction using a hexane:ethylacetate = 2:3 mixture. The steroid extracts were applied to a C18 Amprep solid-phase column (Amersham Biosciences). The androgen or estrogen fraction was purified from the eluted steroids using a normal-phase HPLC system (Jasco) with a silica gel column. For induced ionization, T and DHT were derivatized into T or DHT-picolinoyl (12). To increase the evaporation efficiency, E2 was derivatized into estradiol-3-pentafluorobenzoxy-17-picolinoyl-ester (estradiol-PFBz-picolinoyl). The LC-MS/MS system, which consisted of a reverse-phase LC coupled with an API 5000 triple-stage quadrupole mass spectrometer (Applied Biosystems), was operated with electron spray ionization in the positive-ion mode. The LC chromatographic separation for T, DHT-picolinoyl, or estradiol-PFBz-picolinoyl was performed on a Cadenza CD-C18 column (Imtakt). The MS/MS processes monitored the m/z transition from 396 to 203 for DHT-picolinoyl, from 394 to 253 for T-picolinoyl, and from 558 to 339 for estradiol-PFBz-picolinoyl. DHT-picolinoyl-d3, T-picolinoyl-d3, and 13C4-estradiol-PFBz-picolinoyl were used as internal standards to measure the recovery of steroids and to calibrate the retention time.
Statistical Analysis.
Values are expressed as mean ± SEM and were analyzed using Prism 5 (MDF Co.). Data were analyzed with one-way ANOVAs followed by Dunnet’s post hoc or two-way ANOVAs followed by Bonferroni post hoc tests. Statistical significance was assumed at P values < 0.05.
Supplementary Material
Acknowledgments
We thank M. Iemitsu and K. Aizawa for technical support of real-time quantitative PCR. This work was supported in part by Japan Society for the Promotion of Science (Grants-in-aid for Scientific Research B 20300214 and A 23240091) and for the Body and Mind Integrated Sports Sciences Project (2010–2013) of the Ministry of Education, Culture, Sports, Science and Technology, Japan; and by a grant-in-aid for the Japan Society for the Promotion of Science Fellows (11J01243). Masahiro Okamoto and Takashi Matsui are both Research Fellows of the Japan Society for the Promotion of Science.
Footnotes
The authors declare no conflict of interest.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1210023109/-/DCSupplemental.
References
- 1.Cameron HA, Gould E. The control of neuronal birth and survival. In: Shaw C, editor. Receptor Dynamics in Neural Development. Boca Raton, FL: CRC Press; 1996. pp. 141–157. [Google Scholar]
- 2.Gage FH. Mammalian neural stem cells. Science. 2000;287:1433–1438. doi: 10.1126/science.287.5457.1433. [DOI] [PubMed] [Google Scholar]
- 3.Galea LA, Spritzer MD, Barker JM, Pawluski JL. Gonadal hormone modulation of hippocampal neurogenesis in the adult. Hippocampus. 2006;16:225–232. doi: 10.1002/hipo.20154. [DOI] [PubMed] [Google Scholar]
- 4.Gould E, Woolley CS, Frankfurt M, McEwen BS. Gonadal steroids regulate dendritic spine density in hippocampal pyramidal cells in adulthood. J Neurosci. 1990;10:1286–1291. doi: 10.1523/JNEUROSCI.10-04-01286.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.MacLusky NJ, Hajszan T, Prange-Kiel J, Leranth C. Androgen modulation of hippocampal synaptic plasticity. Neuroscience. 2006;138:957–965. doi: 10.1016/j.neuroscience.2005.12.054. [DOI] [PubMed] [Google Scholar]
- 6.Nguyen TV, Jayaraman A, Quaglino A, Pike CJ. Androgens selectively protect against apoptosis in hippocampal neurones. J Neuroendocrinol. 2010;22:1013–1022. doi: 10.1111/j.1365-2826.2010.02044.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hatanaka Y, et al. Androgen rapidly increases dendritic thorns of CA3 neurons in male rat hippocampus. Biochem Biophys Res Commun. 2009;381:728–732. doi: 10.1016/j.bbrc.2009.02.130. [DOI] [PubMed] [Google Scholar]
- 8.Kerr JE, Allore RJ, Beck SG, Handa RJ. Distribution and hormonal regulation of androgen receptor (AR) and AR messenger ribonucleic acid in the rat hippocampus. Endocrinology. 1995;136:3213–3221. doi: 10.1210/endo.136.8.7628354. [DOI] [PubMed] [Google Scholar]
- 9.Spritzer MD, Galea LA. Testosterone and dihydrotestosterone, but not estradiol, enhance survival of new hippocampal neurons in adult male rats. Dev Neurobiol. 2007;67:1321–1333. doi: 10.1002/dneu.20457. [DOI] [PubMed] [Google Scholar]
- 10.Hojo Y, et al. Adult male rat hippocampus synthesizes estradiol from pregnenolone by cytochromes P45017alpha and P450 aromatase localized in neurons. Proc Natl Acad Sci USA. 2004;101:865–870. doi: 10.1073/pnas.2630225100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kretz O, et al. Hippocampal synapses depend on hippocampal estrogen synthesis. J Neurosci. 2004;24:5913–5921. doi: 10.1523/JNEUROSCI.5186-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hojo Y, et al. Comparison between hippocampus-synthesized and circulation-derived sex steroids in the hippocampus. Endocrinology. 2009;150:5106–5112. doi: 10.1210/en.2009-0305. [DOI] [PubMed] [Google Scholar]
- 13.Konkle AT, McCarthy MM. Developmental time course of estradiol, testosterone, and dihydrotestosterone levels in discrete regions of male and female rat brain. Endocrinology. 2011;152:223–235. doi: 10.1210/en.2010-0607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Melcangi RC, Celotti F, Castano P, Martini L. Differential localization of the 5 alpha-reductase and the 3 alpha-hydroxysteroid dehydrogenase in neuronal and glial cultures. Endocrinology. 1993;132:1252–1259. doi: 10.1210/endo.132.3.8440186. [DOI] [PubMed] [Google Scholar]
- 15.Kimoto T, et al. Neurosteroid synthesis by cytochrome p450-containing systems localized in the rat brain hippocampal neurons: N-methyl-D-aspartate and calcium-dependent synthesis. Endocrinology. 2001;142:3578–3589. doi: 10.1210/endo.142.8.8327. [DOI] [PubMed] [Google Scholar]
- 16.Okamoto M, et al. Reduction in paracrine Wnt3 factors during aging causes impaired adult neurogenesis. FASEB J. 2011;25:3570–3582. doi: 10.1096/fj.11-184697. [DOI] [PubMed] [Google Scholar]
- 17.Nishijima T, Okamoto M, Matsui T, Kita I, Soya H. Hippocampal functional hyperemia mediated by NMDA receptor/NO signaling in rats during mild exercise. J Appl Physiol. 2012;112:197–203. doi: 10.1152/japplphysiol.00763.2011. [DOI] [PubMed] [Google Scholar]
- 18.van Praag H. Neurogenesis and exercise: Past and future directions. Neuromolecular Med. 2008;10:128–140. doi: 10.1007/s12017-008-8028-z. [DOI] [PubMed] [Google Scholar]
- 19.Louissaint A, Jr, Rao S, Leventhal C, Goldman SA. Coordinated interaction of neurogenesis and angiogenesis in the adult songbird brain. Neuron. 2002;34:945–960. doi: 10.1016/s0896-6273(02)00722-5. [DOI] [PubMed] [Google Scholar]
- 20.Feng Y, et al. Spatiotemporal expression of androgen receptors in the female rat brain during the oestrous cycle and the impact of exogenous androgen administration: A comparison with gonadally intact males. Mol Cell Endocrinol. 2010;321:161–174. doi: 10.1016/j.mce.2010.02.029. [DOI] [PubMed] [Google Scholar]
- 21.Tabori NE, et al. Ultrastructural evidence that androgen receptors are located at extranuclear sites in the rat hippocampal formation. Neuroscience. 2005;130:151–163. doi: 10.1016/j.neuroscience.2004.08.048. [DOI] [PubMed] [Google Scholar]
- 22.van Praag H, Christie BR, Sejnowski TJ, Gage FH. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc Natl Acad Sci USA. 1999;96:13427–13431. doi: 10.1073/pnas.96.23.13427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim SH, et al. Treadmill exercise increases cell proliferation without altering of apoptosis in dentate gyrus of Sprague-Dawley rats. Life Sci. 2002;71:1331–1340. doi: 10.1016/s0024-3205(02)01849-0. [DOI] [PubMed] [Google Scholar]
- 24.Trejo JL, Carro E, Torres-Aleman I. Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J Neurosci. 2001;21:1628–1634. doi: 10.1523/JNEUROSCI.21-05-01628.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Soya H, et al. Threshold-like pattern of neuronal activation in the hypothalamus during treadmill running: Establishment of a minimum running stress (MRS) rat model. Neurosci Res. 2007;58:341–348. doi: 10.1016/j.neures.2007.04.004. [DOI] [PubMed] [Google Scholar]
- 26.Saito T, Soya H. Delineation of responsive AVP-containing neurons to running stress in the hypothalamus. Am J Physiol Regul Integr Comp Physiol. 2004;286:R484–R490. doi: 10.1152/ajpregu.00453.2003. [DOI] [PubMed] [Google Scholar]
- 27.Wasserman K, Beaver WL, Whipp BJ. Mechanisms and patterns of blood lactate increase during exercise in man. Med Sci Sports Exerc. 1986;18:344–352. doi: 10.1249/00005768-198606000-00017. [DOI] [PubMed] [Google Scholar]
- 28.Soya H, et al. BDNF induction with mild exercise in the rat hippocampus. Biochem Biophys Res Commun. 2007;358:961–967. doi: 10.1016/j.bbrc.2007.04.173. [DOI] [PubMed] [Google Scholar]
- 29.McIlwain H, Bachelard HS. 1985 Biochemistry and the Central Nervous System (Churchill Livingstone, Edinburgh), 660 pp. [Google Scholar]
- 30.Normington K, Russell DW. Tissue distribution and kinetic characteristics of rat steroid 5 alpha-reductase isozymes. Evidence for distinct physiological functions. J Biol Chem. 1992;267:19548–19554. [PubMed] [Google Scholar]
- 31.Pignatelli D, Xiao F, Gouveia AM, Ferreira JG, Vinson GP. Adrenarche in the rat. J Endocrinol. 2006;191:301–308. doi: 10.1677/joe.1.06972. [DOI] [PubMed] [Google Scholar]
- 32.Compagnone NA, Mellon SH. Neurosteroids: Biosynthesis and function of these novel neuromodulators. Front Neuroendocrinol. 2000;21:1–56. doi: 10.1006/frne.1999.0188. [DOI] [PubMed] [Google Scholar]
- 33.Kempermann G, Jessberger S, Steiner B, Kronenberg G. Milestones of neuronal development in the adult hippocampus. Trends Neurosci. 2004;27:447–452. doi: 10.1016/j.tins.2004.05.013. [DOI] [PubMed] [Google Scholar]
- 34.Yang F, et al. Linking beta-catenin to androgen-signaling pathway. J Biol Chem. 2002;277:11336–11344. doi: 10.1074/jbc.M111962200. [DOI] [PubMed] [Google Scholar]
- 35.Kuwabara T, et al. Wnt-mediated activation of NeuroD1 and retro-elements during adult neurogenesis. Nat Neurosci. 2009;12:1097–1105. doi: 10.1038/nn.2360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Glasper ER, Llorens-Martin MV, Leuner B, Gould E, Trejo JL. Blockade of insulin-like growth factor-I has complex effects on structural plasticity in the hippocampus. Hippocampus. 2010;20:706–712. doi: 10.1002/hipo.20672. [DOI] [PubMed] [Google Scholar]
- 37.Piehl K. Time course for refilling of glycogen stores in human muscle fibres following exercise-induced glycogen depletion. Acta Physiol Scand. 1974;90:297–302. doi: 10.1111/j.1748-1716.1974.tb05592.x. [DOI] [PubMed] [Google Scholar]
- 38.Mitsushima D, Takase K, Funabashi T, Kimura F. Gonadal steroids maintain 24 h acetylcholine release in the hippocampus: Organizational and activational effects in behaving rats. J Neurosci. 2009;29:3808–3815. doi: 10.1523/JNEUROSCI.5301-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mohapel P, Leanza G, Kokaia M, Lindvall O. Forebrain acetylcholine regulates adult hippocampal neurogenesis and learning. Neurobiol Aging. 2005;26:939–946. doi: 10.1016/j.neurobiolaging.2004.07.015. [DOI] [PubMed] [Google Scholar]
- 40.Ho NF, Han SP, Dawe GS. Effect of voluntary running on adult hippocampal neurogenesis in cholinergic lesioned mice. BMC Neurosci. 2009;10:57. doi: 10.1186/1471-2202-10-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Burns JM, Johnson DK, Watts A, Swerdlow RH, Brooks WM. Reduced lean mass in early Alzheimer disease and its association with brain atrophy. Arch Neurol. 2010;67:428–433. doi: 10.1001/archneurol.2010.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yao M, Nguyen TV, Rosario ER, Ramsden M, Pike CJ. Androgens regulate neprilysin expression: Role in reducing beta-amyloid levels. J Neurochem. 2008;105:2477–2488. doi: 10.1111/j.1471-4159.2008.05341.x. [DOI] [PubMed] [Google Scholar]
- 43.Mukai H, et al. Automated analysis of spines from confocal laser microscopy images: Application to the discrimination of androgen and estrogen effects on spinogenesis. Cereb Cortex. 2011;21:2704–2711. doi: 10.1093/cercor/bhr059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Edinger KL, Frye CA. Androgens’ performance-enhancing effects in the inhibitory avoidance and water maze tasks may involve actions at intracellular androgen receptors in the dorsal hippocampus. Neurobiol Learn Mem. 2007;87:201–208. doi: 10.1016/j.nlm.2006.08.008. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.